

Abstract
For many genes, proper gene expression requires coordinated and dynamic interactions between multiple regulatory elements, each of which can either promote or silence transcription. In Drosophila, the complexity of the regulatory landscape is further complicated by the tight physical pairing of homologous chromosomes, which can permit regulatory elements to interact in trans, a phenomenon known as transvection. To better understand how gene expression can be programmed through cis- and trans-regulatory interactions, we analyzed transvection effects for a collection of alleles of the eyes absent (eya) gene. We find that trans-activation of a promoter by the eya eye-specific enhancers is broadly supported in many allelic backgrounds, and that the availability of an enhancer to act in trans can be predicted based on the molecular lesion of an eya allele. Furthermore, by manipulating promoter availability in cis and in trans, we demonstrate that the eye-specific enhancers of eya show plasticity in their promoter preference between two different transcriptional start sites, which depends on promoter competition between the two potential targets. Finally, we show that certain alleles of eya demonstrate pairing-sensitive silencing resulting from trans-interactions between Polycomb Response Elements (PREs), and genetic and genomic data support a general role for PcG proteins in mediating transcriptional silencing at eya. Overall, our data highlight how eya gene regulation relies upon a complex but plastic interplay between multiple enhancers, promoters, and PREs.
Author summary
Gene regulation requires interactions between regions of DNA known as regulatory elements, which, in combination, determine where and when a gene will be active or silenced. Some genes use just a few regulatory elements, whereas others rely on highly complex interactions between many different elements that are poorly understood. While we typically imagine regulatory elements interacting with one another along the length of a single chromosome, in a curious phenomenon called transvection, elements can communicate between two different chromosomes that are held in close proximity. Here, we use the study of transvection to better understand how different regulatory elements contribute to the expression of eyes absent (eya), a gene required for proper eye development in Drosophila. Our data show that a class of elements that initiate eya gene expression, called promoters, will compete with one another for activation by eya’s enhancers, a second class of regulatory element, with the promoter that is closest to the enhancers being the favored target for activation. Furthermore, our study of transvection uncovers an important role for a silencing element, called a PRE, in opposing eya gene expression. Overall, our study sheds new light on how different elements combine to produce patterned expression of eya.
Introduction
The eukaryotic genome is rich in regulatory elements whose combined inputs lead to proper execution of programmed patterns of gene expression. Regulatory elements that promote gene expression include promoters, where RNA polymerases begin transcription of genes, and enhancers, which bind to transcriptional activator proteins and are thought to physically interact with promoters via looping, thereby recruiting or activating RNA polymerases [1, 2]. Conversely, other DNA elements play roles in preventing transcription locally, including Polycomb Response Elements (PREs), which bind to complexes of proteins known as the Polycomb Group (PcG) and can ultimately create a silenced chromatin domain via the histone mark H3K27me3 [3, 4]. While our ability to identify these types of regulatory elements has grown with increasing accuracy via the refinement of sophisticated genomic approaches, our understanding of how specific elements interact with one another across diverse tissues remains incomplete.
In Drosophila, specificity of interactions between regulatory sequences is further complicated by the phenomenon of somatic homolog pairing, where homologous chromosomes are held in close proximity in virtually all somatic cells of the organism [5]. A growing body of data supports that somatic homolog pairing permits regulatory elements on one homolog to interact with those on the homologous chromosome, a phenomenon coined transvection by its discoverer, Ed Lewis [6]. The term transvection encompasses several types of pairing-dependent genetic interactions, including those that positively impact gene expression, as is the case when an enhancer on one chromosome acts in trans to activate transcription from a promoter on the homologous chromosome, or that negatively impact gene expression, as observed in some cases when PREs interact in trans, which is thought to increase the efficacy of PcG proteins bound to the PRE in silencing transcription (Fig 1) [5, 7].
Fig 1. Models for transvection via enhancer action in trans and pairing-sensitive silencing.

A, mutant alleles of yellow can be placed into at least three classes based on their molecular lesions: Class A includes deletions of enhancers and insertions of insulator elements between the enhancers and the promoter; Class B includes point mutations and deletions within the core promoter and transposon insertions in either the promoter or the 5’ UTR; Class C includes point mutations and indels in the coding region. Class A and Class B alleles complement when paired, indicating robust enhancer action in trans, whereas complementation between Class A and Class C is either non-existent or too weak to be detected, indicating poor enhancer action in trans. B, a PRE fused to a mini-white transgene typically permits expression in a hemizygous transgenic insertion, but silences expression when insertions are homozygous, reflecting pairing-sensitive silencing.
The study of transvection typically relies on specific mutant backgrounds and/or transgenic organisms with defined constructs placed at equivalent positions on homologous chromosomes. For example, enhancer action in trans has been studied extensively for the yellow gene of Drosophila, which is required for pigmentation of the adult cuticle [8, 9]. The yellow gene has a simple structure, with a single promoter and transcription start site (TSS) and several well-defined tissue-specific enhancers, and is rich in classical alleles that impact gene expression. Notably, enhancer action in trans at yellow appears tightly regulated; intragenic complementation via transvection is observed only between two types of alleles, those in which enhancers are deleted or otherwise prevented from interacting with the yellow promoter in cis (“Class A” alleles), and those in which the promoter region is compromised by deletion, mutation, or nearby transposon insertion (“Class B” alleles) [9] (Fig 1A). In contrast, alleles of yellow that have an intact promoter but carry mutations in the coding region of the gene, known as “Class C” alleles, fail to complement Class A alleles despite carrying functional enhancers that would otherwise be available to act in trans (Fig 1A). The failure of the yellow enhancers of Class C alleles to act in trans has been interpreted to be due to their preference for a promoter in cis; it is only when the cis-promoter is somehow compromised, as in Class B alleles, that the yellow enhancers are released to act in trans, suggesting a hierarchical regulation of potential promoter targets for the yellow enhancers.
Several other examples of enhancer action in trans show evidence that enhancers prefer to act on a promoter in cis relative to a promoter in trans [10–14]. However, in these cases, activation of a trans-promoter is attenuated, but not eliminated, in the presence of a promoter in cis. A simple interpretation is that cis-preference is a global phenomenon that is relevant to many enhancers in Drosophila, and that yellow represents an extreme case where cis-preference is strong enough to reduce trans-activation to undetectable levels. However, aside from analyses at yellow, there has yet to be further characterization of transvection in Drosophila that uses a diverse collection of alleles analogous to the Class A, Class B, and Class C alleles of yellow.
The eyes absent (eya) gene encodes an evolutionarily conserved transcriptional co-activator with protein phosphatase activity that is a component of the Retinal Determination Network (RDN) of transcriptional regulators required for normal eye development in Drosophila [15, 16]. Eye-specific loss of eya function can cause an “eyeless” phenotype in adult flies, whereas ectopic expression of eya can lead to development of eye tissue elsewhere in the body [16, 17]. Eya functions in part via the formation of a complex with the DNA-binding Sine oculis (So) protein, thereby acting as a bipartite transcription factor that regulates RDN gene expression and coordinates other downstream processes of eye differentiation [18].
The eya gene structure contains two major transcriptional start sites, with the first exon of the eya-B (also known as Type I) transcript encoded roughly 10kb upstream of that of eya-A (Type II) (Fig 2A). Each of the alternate first exons splices to common second through fifth exons that together encode the majority of the protein. Presumed null mutations in the coding region result in embryonic lethality due to the requirement of eya activity in diverse tissues at early stages of development, likely via transcription from the eya-A promoter [19–23]. In contrast, retrotransposon insertions into Exon 1B result in homozygous viable flies entirely lacking compound eyes and ocelli, suggesting that the transcript initiated from the eya-B promoter is required for development of eye structures [24]. Upstream of the eya-B promoter is approximately 9 kb of non-coding sequence that carries several enhancers for distinct eye tissues [24–26], and a DNA fragment carrying this 9 kb fragment can fully recapitulate the wild-type eya expression pattern in third instar larval eye discs [25].
Fig 2. Classical alleles of eya.
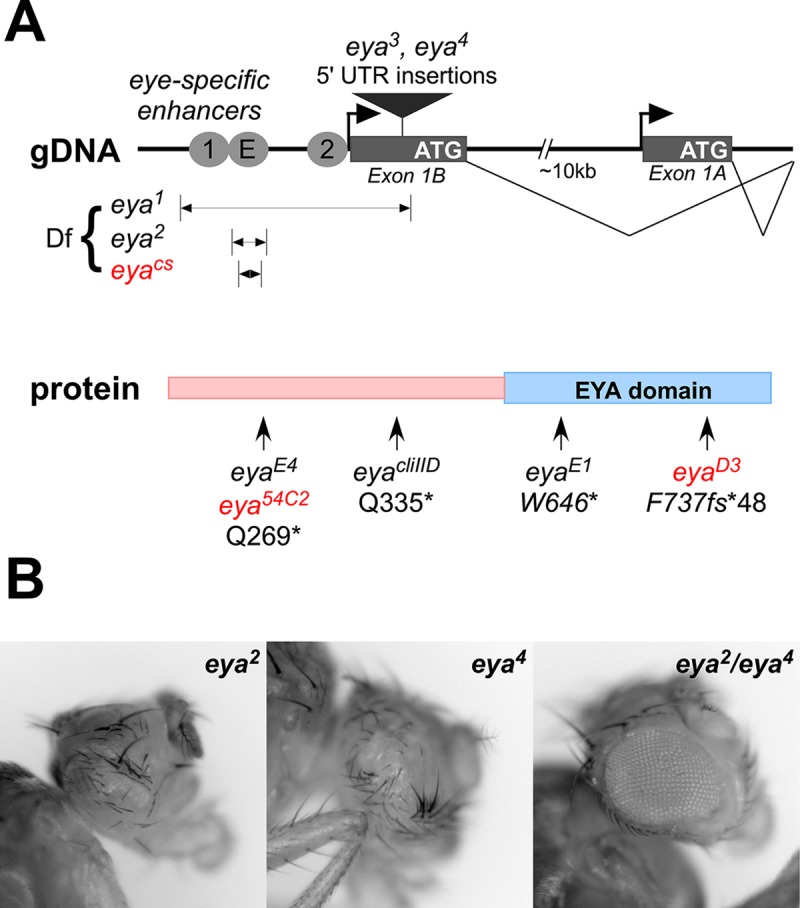
A, top, alleles affecting regulatory sequences mapped to a schematic of the locus. Transcription of eya initiates from two major promoters, eya-B and eya-A, which are separated by approximately 10 kb. Several eye-specific enhancers are located upstream of the eya-B promoter; E is active in early instar discs and is required for eye development, whereas 1 (also known as IAM) and 2 drive expression in later disc tissues [24–26]. Other fragments identified as candidate late enhancers in other regions of the locus are omitted for simplicity. Alleles eya1, eya2, and eyacs delete eye-specific enhancer sequences, and eya3 and eya4 carry retrotransposon insertions into the 5’ UTR of exon 1B. Bottom, alleles with point mutations or indels mapped to the Eya protein structure. Alleles in red had their molecular lesions characterized in this work (see S1 File). The eyaD3 allele carries an indel in exon 5 where a TT doublet is substituted by CCCCCCCCCG, creating a frameshift at F737 and resulting in a C-terminus of 48 random amino acids in place of the usual 23. B, example of transvection between eya2 (Class A) and eya4 (Class B), first demonstrated by Leiserson et al. [27]. Homozygosity of either allele results in eyeless flies, but trans-heterozygotes produce an eye of approximately ¾ size via enhancer action in trans.
Transvection has been previously demonstrated for alleles of eya that are analogous to the Class A and Class B alleles of yellow [27]. Specifically, trans-heterozygous flies carrying eya2, an enhancer deletion, and eya4, an insertion in the 5’ UTR of Exon 1B, result in pairing-dependent rescue of eye development, likely via enhancer action in trans. Furthermore, several other alleles of eya are structurally similar to Class C alleles of yellow, with missense and nonsense mutations in the eya coding region, and several show some degree of complementation with eya Class A alleles that could be consistent with transvection [19, 20].
Here we demonstrate that Class A alleles of eya complement all Class B and Class C alleles tested, demonstrating that enhancer action in trans is less strictly regulated at eya relative to yellow. However, complementation by Class B is consistently stronger than that by Class C across several genetic backgrounds, suggesting that the availability of enhancers to act in trans differs depending on the molecular lesion in eya. Furthermore, by manipulating promoter availability in cis and in trans, we show that the eye-specific enhancers of eya show preference for the eya-B promoter relative to the eya-A promoter, and that the preference depends on competition between the two promoter targets. Finally, genetic interactions between eya Class A alleles demonstrate pairing-sensitive silencing, and genetic and genomic data support a role for PcG proteins in mediating eya transcriptional silencing.
Results
To date, few loci have been studied in detail for transvection effects using a large number of classical mutations. Transvection via enhancer action in trans was previously demonstrated at the eya locus, but only one allelic combination was reported in detail (Fig 2B) [27], whereas many more alleles are described in the literature. Thus, to better understand how regulatory regions can communicate in cis and in trans, we quantitatively characterized transvection effects of a collection of eya alleles.
To begin, we gathered Drosophila stocks carrying known eya alleles and classified them based on their molecular lesions. For those alleles lacking molecular characterization, we identified lesions by sequencing genomic DNA and other methods (Fig 2B; see S1 File). Following the classification scheme of Morris et al. [9], we categorized three alleles, eya1, eya2, and eyacs, as Class A alleles, with each carrying a deletion of enhancer sequences upstream of the eye-specific eya-B promoter [16, 24, 26]. In contrast, the Class B alleles eya3 and eya4 carry transposon insertions in the 5’ UTR near the eya-B promoter [24], and the Class C alleles eyaE1, eyaE4, eyacliIID, eya54C2, and eyaD3 carry point mutations and/or indels in the eya coding region that is common to eya-A and eya-B isoforms [16, 22, 28]. Note that Class C alleles of eya are embryonic lethal as homozygotes or as trans-heterozygotes with other Class C alleles, reflecting the requirement for eya function during embryogenesis [19, 20], whereas Class A and Class B alleles are homozygous viable, suggesting that they primarily impact eye development.
To assess enhancer action in trans, we created flies carrying a Class A allele on one homolog and either a Class B allele or a Class C allele on the other homolog (Fig 3). We initially focused on the strong Class A alleles eya1 and eya2, each of which has a completely eyeless phenotype as a homozygote [16, 27]. To quantify the strength of transvection in trans-heterozygous flies, we scored the number of ommatidia in adult fly eyes from each genotypic combination (see Materials and Methods). All seven Class B and Class C alleles that we tested partially complemented the eyeless phenotype when placed in trans to Class A alleles, consistent with enhancer action in trans being generally permissible for both Class B and Class C alleles of eya (Figs 3 and S2). Notably, Class B alleles consistently showed higher levels of complementation relative to Class C alleles in both eya1 and eya2 backgrounds; specifically, while there was no significant difference in the numbers of ommatidia of flies carrying eya3 and eya4 alleles as trans-heterozygotes with eya1, each had a significantly higher ommatidia count than the Class C alleles in combination with eya1 (adjusted p < 0.05, Kruskal-Wallis test with Dunn’s multiple comparisons test). The same was true of ommatidia counts of Class B and Class C alleles in combination with the Class A allele eya2 with the exception of one comparison (eya3/eya2 vs eyacliIID/eya2, adjusted p = 0.17). Thus, our data support that the strength of complementation between Class A and Class B alleles is consistently greater than that between Class A and Class C alleles.
Fig 3. Transvection via enhancer action in trans is supported by diverse alleles of eya.

A, eyes of representative flies carrying the Class A allele indicated at left and the Class B or Class C allele indicated above. B, quantification of eye development for flies carrying either eya1 (left graph) or eya2 (right graph) and the Class B or Class C alleles indicated below. For each genotype, approximately 20 eyes from 10 flies were scored for the number of ommatidia.
To confirm that the complementation observed between Class A and Class B or C alleles was due to enhancer action in trans, we took advantage of a chromosomal rearrangement of the eya2 allele that was previously shown to disrupt transvection between eya2 and eya4 [27]. All Class B and Class C alleles failed to complement the rearranged eya2 allele ETD2.2 to the extent that they complemented a structurally wild type eya2 chromosome, confirming that intragenic complementation is indeed pairing-dependent (Fig 4). Furthermore, Class B alleles showed partial complementation in combination with ETD2.2, with an average count of 133.3 ± 85.1 ommatidia per eye, whereas Class C alleles showed a near complete failure of complementation with ETD2.2, with the majority of flies being completely eyeless (Fig 4). Importantly, all Class B and Class C alleles completely fail to complement the eyeless phenotype of the Class B allele eya4 (Fig 4A), indicating that differences in transvection observed for Class B vs Class C are unlikely to be due to general differences in the penetrance or expressivity of the eyeless phenotype between the two allele classes [9, 27]. In sum, Class B alleles, which carry insertions near the promoter of the eya-B transcript, support higher levels of transvection with Class A alleles than do Class C alleles, which have no disruptions near the B-transcript promoter.
Fig 4. Transvection at eya via enhancer action in trans requires somatic homolog pairing.

A, eyes of representative flies carrying the allele indicated at left in trans to the Class B or Class C allele indicated above. ETD2.2 is a transvection-disrupting rearrangement of a second chromosome carrying eya2. B, quantification of eye development for flies carrying the indicated alleles. Approximately 20 eyes from 10 flies were scored for each genotype. Note that the data for eya2 crosses are identical to those presented in Fig 3.
As an additional test, we assessed the impact of mutations in Cap-H2, a component of the Condensin II complex, on intragenic complementation between alleles of eya. Loss of Cap-H2 has been shown to increase levels of somatic homolog pairing, which can result in elevated levels of transvection relative to a wild type Cap-H2 background [29, 30]. Indeed, flies of genotype eya2/eya4; Cap-H20019/Cap-H25163 have a significantly greater number of ommatidia than eya2/eya4 flies in wild-type Cap-H2 backgrounds (p = 0.02, Mann-Whitney test) (S3 Fig), adding further evidence that intragenic complementation at the eya locus is due to transvection.
Enhancer action in trans reveals plasticity in enhancer-promoter specificity
Prior analyses have demonstrated that, whereas expression during embryogenesis is specific to the eya-A promoter, both the eya-A and eya-B promoters are active in the developing eye disc [20, 26]. Furthermore, qRT-PCR analysis supports that the eye-specific enhancers upstream of the eya-B promoter activate transcription of both transcript types [26]. To better understand transcript-specific expression in the developing eye, we performed in situ hybridization on wild type third instar larval eye discs using probes specific to the first exons of either the eya-A or eya-B transcript. Analysis of the eya-B isoform showed robust expression in progenitor cells anterior to the morphogenetic furrow and in differentiating cells immediately posterior the furrow, with lower levels of expression observed in more mature ommatidial clusters toward the posterior of the disc (Fig 5A). Expression is also seen in the developing ocelli, consistent with a requirement for eya in ocelli development [16]. The pattern of staining for the eya-A transcript appears similar to that of the eya-B transcript, with highest expression seen immediately anterior and posterior to the morphogenetic furrow (Fig 5B). However, the signal for the eya-A transcript is barely detectable above background fluorescence, suggesting that the eye-specific enhancers of eya act predominantly on the eya-B promoter in the developing eye, and only act weakly on the eya-A promoter.
Fig 5. Plasticity in promoter use for enhancer action in trans.

A-F, third instar eye-antennal discs from wild type (A, B), eya4/eya2 (C, D), or eya4/eya1 (E, F) larvae. Discs were subjected to in situ hybridization using probes specific to exon 1B (A, C, E) or exon 1A (B, D, F). Arrowheads indicate the approximate position of the morphogenetic furrow. Robust staining is seen for exon 1B in all cases; Exon 1A shows weak staining in wild type, is generally undetectable above background staining in eya4/eya2, and appears prevalent in eya4/eya1. Schematic diagrams (above) indicate likely promoter usage in each genotype.
The alleles eya3 and eya4 are caused by retrotransposon insertions in the first exon of eya-B (Exon 1B), and do not directly impact the eya-A transcript [24]. Furthermore, in situ hybridization demonstrates that the retroelement in eya4 is transcribed in an eya-like pattern, reflecting that the eye enhancers are functional in this allele [24]. Two plausible models could account for the lack of eye development in eya3 and eya4 flies: first, assuming that the eya-B transcript is rendered non-functional by the retroelement insertions, production of functional mRNA solely from the eya-A promoter could be insufficient to generate eye tissue. Alternatively, it could be that retrotransposon insertion into Exon 1B decreases or prevents communication between eye-specific enhancers and the eya-A promoter, resulting in loss of eya-A transcription and leaving only the non-functional eya-B transcript. To assess these models and thereby gain a better understanding of transcript usage in the developing eye, we employed isoform-specific in situ hybridization in discs that display enhancer action in trans. In eya4/eya2 discs, we observe robust expression of eya-B, likely reflecting strong activation of the retrotransposon-carrying Exon 1B in cis to the functional enhancers of the eya4 chromosome in addition to trans-activation of the functional Exon 1B on the eya2 chromosome (Fig 5C). However, levels of the eya-A transcript appear strongly decreased in eya4/eya2 discs (no detectable signal in 6/7 discs scored) (Fig 5D), suggesting that the retrotransposon insertion into Exon 1B significantly decreases transcription from the eya-A promoter, and that the eya-B promoter is the preferred target in cis and in trans to the eye-specific enhancers.
In contrast to the small enhancer deletion of the eya2 allele, the eya1 deletion removes enhancer sequences and the eya-B promoter [26]; thus, in eya4/eya1 discs, functional eya-B transcript cannot be generated from either chromosome. In contrast to our observations in eya4/eya2 discs, we easily detect RNA signal for the eya-A transcript in eya4/eya1 discs (7/7 discs scored), consistent with a model wherein the eya-A promoter is trans-activated by the functional enhancers of the eya4 chromosome in this background (Fig 5E and 5F). To further support a difference in promoter usage in eya4/eya2 vs. eya4/eya1 discs, we employed isoform-specific quantitative RT-PCR on eye-antennal discs from these genotypes and compared the levels of eya-A transcripts relative to those of eya-B. Notably, eya-A transcript levels in eya4/eya2 discs drop to 56% (95% CI 52.6%-59.9%, n = 3 biological replicates) when compared to transcripts from eya4/eya1 discs, further supporting decreased expression of eya-A relative to eya-B in eya4/eya2 discs. In sum, our data demonstrate that the eye-specific eya enhancers show a preference for the eya-B promoter, and support that the loss of eye development in eya3 and eya4 flies involves a reduction in activation of the eya-A transcript in addition to the insertional disruption of the eya-B transcript. Furthermore, our data suggest that, in the absence of a functional eya-B promoter in trans, the enhancers can switch their specificity to trans-activate the eya-A promoter in order to produce eye tissue.
Plasticity in enhancer-promoter specificity in cis revealed by deletion of a preferred promoter target
To further assess the requirement for the eya-B transcript in eye development, we used CRISPR-Cas9 to completely remove Exon 1B and its associated core promoter from the genome (Fig 6A). Synthetic guide RNAs designed to flank Exon 1B and under the control of the U6 promoter were injected into embryos carrying a source of Cas9, and the progeny of the resulting flies were screened via PCR for the expected deletion, resulting in four independent mutants lacking exon 1B (see Materials and Methods). All four mutant alleles are viable and fertile as homozygotes, consistent with the proposed eye-specific role for Exon 1B. Surprisingly, all mutants lacking Exon 1B develop near-wild type eyes as either homozygotes or in combination with Df(2L)eya (Fig 6B and 6C). To address eye-specific promoter usage in these mutants, we performed isoform-specific in situ hybridization on third instar larval eye-antennal discs that were homozygous for the Exon 1B deletion. We observed no signal above background for the eya-B probe, confirming that the induced deletion prevents transcription of these sequences (Fig 6D and 6E). Remarkably, staining for the eya-A transcript shows robust signal in a pattern similar to that previously observed for eya-B, demonstrating that the loss of the eya-B promoter results in elevated activation of eya-A transcription (Fig 6F and 6G). To further support this observation, we used quantitative RT-PCR to measure levels of eya-A and eya-B expression in wild type and exon 1B-deleted mutant eye-antennal discs (Fig 6H). Consistent with our in situ data, we observed a 3-fold increase in expression of eya-A, and a complete loss of eya-B, in the mutant discs. Thus, our data support a model wherein wild type eye development relies primarily on activation of the eya-B promoter; in the absence of this promoter and its associated first exon, enhancers shift their specificity to the more distal promoter associated with the eya-A transcript, with near complete compensatory expression to support eye development.
Fig 6. Deletion of the eya-B promoter causes a switch in specificity of eye-specific enhancers to the eya-A promoter.

A, strategy for sgRNA design to delete the core promoter and first exon of the eya-B transcript using CRISPR/Cas9. B-C, eyes of representative flies homozygous (B) or hemizygous (C) for the allele eya2m23a, which carries the deletion indicated in part A. Eyes are nearly identical to wild type. D-G, third instar eye-antennal discs from wild type (D, E), or eya2m23a/eya2m23a (F, G) larvae subjected to in situ hybridization using probes specific to exon 1B (D, F) or exon 1A (E, G). Loss of exon 1B in eya2m23a flies leads to robust upregulation of the eya-A transcript. H, quantitative RT-PCR on cDNA isolated from wild type (left) or homozygous eya2m23a (right) third instar eye-antennal discs using primers specific for exon 1B or exon 1A. Deletion of exon 1B results in a roughly 3.5-fold increase in transcripts from the A promoter.
The eyacs allele demonstrates both active and repressive trans-interactions
The Class A allele eyacs is homozygous viable and hypomorphic, with eyacs/eyacs flies showing a reduced adult eye phenotype with variable expressivity (Fig 7A and 7B). Sequence analysis showed that eyacs carries a 115 bp deletion from -806 to -691 relative to the TSS of the B transcript, which is nested within the enhancer deleted by the eya2 allele (-896 to -577) (Fig 2A). To assess whether eyacs can support enhancer action in trans, we created flies with eyacs on one homolog and various Class B or Class C alleles on the other homolog as we had previously done with the Class A alleles eya1 and eya2. Flies carrying trans-heterozygous combinations of eyacs and Class B or Class C alleles show greater numbers of ommatidia than eyacs homozygotes, comfirming increased expression of eya (Fig 7A and 7B). As observed for other Class A alleles, the strength of transvection is higher when eyacs is in trans to Class B alleles relative to Class C alleles, and a transvection-disrupting rearrangement of the chromosome carrying eya4 [27] shows reduced complementation relative to a structurally wild type chromosome carrying eya4, supporting that the observed complementation between eyacs and Class B and Class C alleles are pairing-dependent (Fig 7A and 7B). Thus, the hypomorphic Class A allele eyacs can participate in enhancer action in trans.
Fig 7. The hypomorphic Class A allele eyacs can participate in enhancer action in trans.

A, eyes of representative flies carrying the indicated genotypes. ETD4.3 is a transvection-disrupting rearrangement of a second chromosome carrying eya4. B, quantification of eye development for flies carrying the indicated alleles in trans to eyacs. Consistent with enhancer action in trans, increased eye development is observed when eyacs is placed in trans to Class B or Class C alleles relative to eyacs homozygotes, and the increase is disrupted by the eya4 rearrangement ETD4.3 (compare column 3 to column 6).
In establishing eyacs as a Class A allele, we were surprised to find that the eye phenotypes of flies carrying eyacs trans-heterozygous with the other Class A alleles are more severe than those of eyacs homozygotes, with eyacs/eya2 having a more severe phenotype than eyacs/eya1 (Fig 8A–8E). Furthermore, the eye phenotype of flies carrying eyacs trans-heterozygous with Df(2L)eya, a large deficiency spanning the entire eya locus, does not show an increased severity relative to eyacs homozygotes, but instead shows a more moderate phenotype (Fig 8A and 8E). Thus, eyacs shows repressive trans-interactions with the small deletions carried by other Class A alleles, but not with a large deletion. To determine whether repressive trans-interactions involving eyacs are pairing-dependent, we created trans-heterozygotes between eyacs and the rearranged eya2 allele ETD2.2 (Fig 8D and 8E). Notably, the disruption of pairing between eya alleles caused by the rearrangement carried by ETD2.2 restored partial eye development in these flies, indicating that the repression of eya by other Class A alleles is pairing-sensitive.
Fig 8. Pairing sensitive silencing between Class A alleles reflects a direct role for PcG proteins in regulating eya.

A-D, eyes of representative flies carrying the indicated genotypes. ETD2.2 is a transvection-disrupting rearrangement of a second chromosome carrying eya2. E, quantification of eye development for flies carrying the indicated alleles in trans to eyacs. Consistent with pairing-sensitive silencing, decreased eye development is observed when eyacs is placed in trans to other Class A alleles relative to eyacs homozygotes, and the decrease is disrupted by the eya2 rearrangement ETD2.2 (compare column 4 to column 5). Furthermore, 98.8% of eyacs/eya2 flies are completely eyeless (n = 462; see Panel K), but all (n = 120) eyacs/ETD2.2 flies develop scorable eye tissue. F, genomic features of the eya locus (note that genomic maps are reversed relative to the reference sequence.) Top, Hi-C contact map showing TAD structure, with chromatin color map below. The eya locus occupies a TAD that is primarily blue chromatin, indicative of Polycomb silencing. Below, ChIP-seq peaks for K27-trimethylated histone H3 and individual PcG proteins as assayed from larval disc tissues. Six putative PREs (red boxes) are indicated by the pattern of peaks. G, schematic showing positions of eye-specific enhancers of putative PREs upstream of the eya B promoter. Bars below the schematic indicate the deletions carried by Class A alleles. H-I, eyes of representative flies carrying the indicated genotypes. J, quantification of eye development for flies carrying the indicated genotypes. Completely eyeless flies were not scored in this analysis. Significant increases in the ommatidia count are observed for eyacs/eya1; E(z)S1/+ (p = 0.03, Mann-Whitney test) and eyacs/eya1; Pc4/+ (p = 0.04) relative to eyacs/eya1; +/+. K, scoring of flies that have at least one eye of any size in the indicated genotypes. Asterisks indicates significant difference relative to eyacs/eya2; +/+ (p = 0.008, Mann-Whitney U test) or eyacs/eya1; +/+ (p = 0.03). “N” indicates the number of flies scored, with the number of separate vials in parentheses. L, Models for interactions between eye-specific enhancers and putative PREs. For simplicity, only the E enhancer (required for initiating eye development) and its neighboring PRE are shown; we expect that other local PREs behave similarly. In wild type, we propose that the enhancer has dual roles to block silencing by PREs through an unknown mechanism and to activate transcription primarily from the eya-B promoter. The eyacs allele retains partial enhancer activity, whereas the eya2 allele has no detectable enhancer activity; in eyacs/eya2 flies, pairing between homologous PREs (dual arrows) increases their capacity to silence, resulting in an eyeless phenotype in the paired state, but a partial eye when unpaired by chromosomal rearrangements. Since eya1 deletes some of the putative PREs, pairing-dependent silencing is reduced, and the eyacs/eya1 phenotype is less severe than that of eyacs/eya2.
Previous analyses of pairing-sensitive silencing have revealed a central role for PcG genes [7]. Specifically, known cases of pairing-sensitive silencing are caused by pairing-dependent interactions between PREs on homologous chromosomes, which is thought to augment the recruitment and/or silencing capacity of PcG complexes relative to those at unpaired PREs. To address whether PcG genes may function in pairing-sensitive silencing at the eya locus, we first assessed ChIP-seq binding profiles of key PcG proteins and the histone H3K27me3 mark associated with Polycomb repressive domains (Fig 8F). Indeed, the compartmental domain occupied by eya is rich in H3K27me3 in third instar larval disc tissue and in cultured Drosophila cells [31–33], and carries several putative PREs indicated by peaks of PcG proteins Ph, Psc, Pc, and E(z) [33]. Two putative PREs are located upstream of the eye-specific eya-B promoter and partially overlap the known eye-specific enhancers [26] (Fig 8F and 8G). Notably, the eya2 deletion is predicted to leave all putative PREs intact, whereas the eya1 deletion removes the two putative PREs that are upstream of the eya-B promoter (Fig 8G).
To provide genetic evidence for a role for PcG proteins in pairing-dependent silencing at eya, we used two approaches to assess eye development in backgrounds with reduced expression of two key PcG components, E(z) and Pc. First, we scored the number of ommatidia in developed adult eyes (excluding eyeless flies) for eyacs/eya1 trans-heterozygous flies with and without reduced PcG gene dosage. In this assay, mutations in both E(z) and Pc act as dominant suppressors of pairing-sensitive silencing (Fig 8H–8J), resulting in greater numbers of ommatidia relative to eyacs/eya1 flies with wild type PcG dosage. Secondly, we compared percentages of total adult flies that were completely eyeless; in an eyacs/eya2 background, nearly all (98.8 ± 1.4%) adults completely lack eyes, but loss of a single functional copy of E(z) suppressed the number of eyeless flies to 75.9 ± 2.5% (p = 0.008, Mann-Whitney U test) (Fig 8H). A similar effect is observed in an eyacs/eya1 background (p = 0.02), but not when a functional copy of Pc was removed from either eyacs/eya1 or eyacs/eya2 flies (Fig 8H), perhaps indicating a greater sensitivity for E(z) relative to Pc function at an early stage of eye specification. In sum, our genetic data and genomic analysis support a role for PcG genes in pairing-sensitive silencing at eya.
Based on our data, we favor a model wherein the eya eye-specific enhancers play dual roles, activating transcription of the eya-B and eya-A promoters and opposing the silencing activity of PREs in the domain occupied by eya (Fig 8L). In flies carrying the eyacs deletion, one or both of these activities is partially compromised, resulting in reduced eye growth; when eyacs is placed in trans to the eya2 allele, enhancer activity is further suppressed while pairing between homologous PREs strengthens their silencing capacity, resulting in complete loss of eye tissue. However, when eyacs is placed in trans with eya1 or larger deletions, or with a rearranged eya2 allele, pairing of some homologous PREs is lost, resulting in re-establishment of partial eye development. In sum, our data indicate an important role for PcG genes in regulating a critical eye determining gene.
Discussion
The proper expression of developmental genes represents a complex interplay of interactions between regulatory sequences that include enhancers, promoters, and silencers such as PREs. Growing evidence supports that regulatory interactions depend upon the positions of DNA elements in three-dimensional space and can be influenced by potential competing interactions with neighboring DNA elements. Due to the complexity of interactions in wild type animals, mutations in regulatory sequences are critical in helping us to better understand how the interaction landscape is assembled via the roles played by individual elements. Furthermore, the study of trans-interactions in Drosophila can uncover aspects of gene regulation that are masked or otherwise challenging to understand in other contexts. Here, by examining cis- and trans-interactions between various classes of eya mutants, we demonstrate a hierarchical set of interactions that shed insight on how enhancers may choose between multiple promoter targets, and suggest an important antagonistic relationship between transcriptional activation by eya enhancers and silencing by local PREs.
Our study was motivated in part by a desire to better understand how gene expression is influenced by trans-interactions, or transvection, in Drosophila. Our data support that the “A, B, C” allele classification put forth by Morris et al. [9] accurately predicts patterns of complementation via enhancer action in trans. Specifically, complementation of Class A alleles by Class B alleles is consistently higher than that by Class C alleles at both yellow and eya. In some Class B alleles of yellow, the core promoter is directly deleted or otherwise mutated, which has led to a model wherein competition by the functional cis-promoters in Class C alleles is a likely explanation for weaker transvection relative to Class B [9, 10]. Other Class B alleles of yellow, and the Class B eya alleles eya3 and eya4, are instead characterized by transposon insertions into the 5’UTR downstream of the promoter, which may cause changes in topology that alter the balance of enhancer-promoter interaction in cis vs trans, although direct evidence for this model is lacking. It should also be noted that variation in strength of enhancer action in trans can be seen within Classes; for example, our data consistently show higher levels of complementation by the Class A allele eya2 relative to that by eya1. This could reflect differences in the specific enhancer elements affected by each deletion, or, alternatively, could result from the availability of the preferred eya-B promoter on the eya2 chromsome vs. its absence on the eya1 chromosome (Fig 5). Similar to these observations, several alleles of Malic Enzyme (Men) would be categorized as Class B since they each carry a promoter deletion, yet they show highly varying levels of trans-activity [34]. Thus, The ABC classes of alleles reflect rough categorizations with respect to an allele’s participation in enhancer action in trans.
To date, transvection effects have been observed for relatively few genes in the Drosophila genome, although transgenic studies using diverse enhancers and genomic locations suggest that enhancer action in trans is widely supported [10, 14, 35–37]. However, we note that genetic interactions consistent with enhancer action in trans have also been reported for two other RDN members, so and eyeless (ey) [38–40]. It may be purely coincidental that a substantial proportion of RDN genes show transvection effects, or, alternatively, somatic homolog pairing and transvection may be of particular importance to gene regulation in the developing eye. Consistent with the latter hypothesis, trans-interactions at the spineless (ss) locus were found to be critical for later fate specifications of photoreceptor types [41]. It will be interesting to explore the potential for trans-interactions at other genes required for eye development to further test this hypothesis.
Plasticity of enhancer-promoter specificity at the eya locus
The presence of multiple eya promoters likely reflects an ancient promoter duplication event, which, like gene duplications, can lead to varying degrees of functional redundancy or sub-functionalization between the alternate TSS [42]. Promoter duplication appears to be widespread in the Drosophila genome; genome-wide mapping of TSS shows that approximately 27% of mapped genes can initiate transcription via two or more promoters, with an average number of 1.4 promoters per gene across all mapped TSS [43]. Comparison to genomes of other Drosophila species suggests that a promoter duplication at eya was a relatively recent event, with evidence of two TSS in the D. melanogaster, D. simulans, and D. ananassae genomes, but not in those of D. pseudoobscura or D. virilis [44, 45].
Prior analyses at eya suggest that the eya-A and eya-B promoters have undergone some degree of sub-functionalization in D. melanogaster, with the eya-B promoter being active primarily within the developing eye disc and the eya-A promoter being more broadly expressed across multiple tissues [20]. Our data support the prior finding that the eya-B promoter is the preferred target of the eye-specific eya enhancers, which are primarily located just upstream of the eya-B promoter, but roughly 10 kb from the eya-A promoter [26]. These observations suggest a simple model wherein specificity can be dictated by relative position; for some enhancers, activity may be highest on nearby promoters, with less activity on more distal promoters. However, our data support that this model is largely driven by promoter competition at the eya locus such that, in the absence of the eya-B promoter, the eya-A promoter becomes a “preferred” target and is highly active. Interestingly, ChIP-seq analysis using antibodies to the insulator protein su(Hw) in embryos suggests the presence of an insulator element in the eya-B first intron that would be predicted to disrupt communication between the eye-specific enhancers and the eya-A promoter [46]. Based on our observations, it is unlikely that this candidate insulator is active in the developing eye disc.
Our data regarding promoter competition and enhancer-promoter proximity is consistent with prior observations where a nearby promoter is preferred to one that is more distal [47–50], and may therefore be generalizable to many enhancers. As a potential caveat in interpreting our data, two other DNA fragments that map close to the eya-A promoter support some degree of transgene expression in the late developing eye disc [26]. These candidate enhancers are not themselves sufficient to rescue eye phenotypes, and are therefore of unknown functional relevance in vivo, but we cannot exclude the possibility that their activity changes in some way upon deletion of the eya-B promoter such that they play a role in the upregulation of eya-A transcription observed in this background. Finally, we note that the current genome annotation supports evidence for a third eya TSS, defining an eya-C transcript that initiates further downstream from the eya-A TSS and is predicted to produce a truncated protein product [45, 51–52]. The biological relevance of this potential promoter and its relationship to the eya-A and eya-B transcripts is as yet unclear.
Pairing-sensitive silencing at eya indicates eya expression is a balance of activating and repressive signals
Our observation of pairing sensitive silencing of eya suggests a direct role for PcG genes in regulating eya expression. In support of this hypothesis, genomic data shows that eya is embedded in a domain of H3K27me3 in cultured cells, embryos, and third instar disc tissues, and distinct peaks of PcG proteins that are characteristic of PREs are found throughout the eya locus [31–33, 53, 54]. Furthermore, reduction in dosage of key PcG proteins, E(z) and Pc, suppresses pairing sensitive silencing of eya, providing genetic evidence for a role for PcG proteins in directly regulating eya expression.
Recently, Erceg et al [55] characterized hundreds of sequences with overlapping PcG binding and enhancer activity, and showed that these fragments can act as enhancers in some cell types and as silencing PREs in others. Notably, one of the candidate PREs upstream of eya overlaps a previously characterized enhancer [26], and the mutations that uncover pairing-sensitive silencing affect sequences in this region. Our data support a model wherein the PRE activity of this region is active in cells outside of the developing eye, silencing eya, whereas the PRE silencing activity is overcome in primordial eye cells in order for eye development to proceed. According to this model, H3K27 methylation would be reduced or suppressed by the activity of the eya enhancers in primordial eye cells (Fig 8L), although we are unable to observe this directly using existing ChIP-seq data derived from mixed larval tissues. Given that several candidate PREs are found across the locus, it is as yet unclear how these different sequences may cooperate and/or interact to determine the transcriptional state of eya in a given tissue.
Interestingly, several other genes of the Retinal Determination Network (RDN) are also characterized by domains of H3K27me3 and localized regions of PcG binding in multiple cell types [31–33, 53, 54]. Furthermore, in addition to eya, RDN genes toy and dac have been identified as having overlapping PRE and enhancer sequences, and a neuronal enhancer from the ey gene was shown to have enhancer activity in some tissues and PRE activity in others [55], suggesting that direct regulation by PcG proteins could be a common feature of RDN genes. According to this model, PcG proteins would maintain RDN genes in an inactive state in non-eye tissues, whereas activation of RDN gene transcription in the developing eye would rely on the coordinated removal of repressive chromatin marks and simultaneous activation of transcription. Consistent with this hypothesis, ChIP-seq analysis shows that binding of the PcG proteins Pho and Ph at the TSS of the RDN genes so and toy is higher in haltere tissues (where the RDN genes are inactive) relative to binding in eye tissue, consistent with reduced binding of PcG proteins at PREs of RDN genes in cells with active expression [56]. However, investigations of roles for PcG proteins in the developing eye are complicated by the widespread pleiotropic effects on gene expression caused by PcG mutations combined with the deeply intertwined regulatory network that determines eye cell fates. For example, clonal loss of E(z) and Pc in cells anterior to the morphogenetic furrow can lead to reduced expression of eya and dachshund (dac), but this is likely due to misexpression of teashirt, which can act as a negative regulator of eya and dac [57, 58]. Similarly, seminal work by Zhu et al. [59] demonstrated a role for PcG proteins in maintaining eye cell fates in the developing eye via the repression of genes that would signal an alternative wing tissue fate. Furthermore, biochemical studies show that Eya protein is a binding partner for Combgap (Cg), a sequence-specific DNA-binding protein that can recruit PcG complexes to PREs, although genetic analyses show that Cg may act in opposition to other PcG complexes in the developing eye [15, 60, 61]. Ultimately, a multifaceted approach involving targeted mutations of individual response elements, combined with transgenic strategies, in backgrounds with altered availability of PcG gene products will likely be required to unravel precise roles for PcG in regulating genes in the developing Drosophila eye.
Materials and methods
Stocks and fly husbandry
Stocks carrying alleles eya2, eya3, eya4, eyaE1, eyaE4, eyaD1, eyacs, Df(2L)eya, ETD2.2 (a chromosome carrying eya2 and the transvection-disrupting inversion In(2LR)29C;41), and ETD4.3 (an eya4 background carrying a transvection-disrupting cyclical translocation with new order T(2; 3; 4) 30A; 101; 98D) were obtained from Nancy Bonini (Department of Biology, University of Pennsylvania, PA). Stocks carrying eyaD3, eyaD6, and eyaD7 were obtained from Justin Kumar (Department of Biology, Indiana University, IN). Stocks carrying eya137.39, eya117.36, and eya7.42 were provided by Jennifer Jemc Mierisch (Department of Biology, Loyola University, Chicago, IL). A stock carrying eya54C2 was obtained from Denise Montell (Department of Molecular, Cellular, and Developmental Biology, UC Santa Barbara, Santa Barbara, CA). Stocks carrying eya1, eyaEY13242, and eyacliftIID were obtained from the Bloomington Drosophila Stock Center (Indiana University, IN). Stocks carrying Cap-H20019 and Cap-H25163 were provided by Giovanni Bosco (Geisel School of Medicine, Dartmouth College, NH). Stocks carrying Pc1 and E(z)S1 (also known as E(z)60) were obtained from Judy Kassis (NIH). All flies were maintained at 25°C in standard 25 mm-diameter vials containing cornmeal, yeast, sugar, and agar medium with p-hydroxybenzoic acid methyl ester to prevent mold [10].
To assess adult eye development, crosses were established between 1–4 males and 2–5 virgin females of the selected genotypes. Progeny flies were collected 1 to 5 days post-eclosion and frozen for preservation. Fly eyes were imaged using a Canon EOS Rebel Tli digital camera mounted on a Leica MZ7.5 stereomicroscope. For each eye, the number of ommatidia was scored manually from the digital images. Mean count data and standard deviations for crosses examining enhancer action in trans are presented in S3 Table as well as the main text figures. Statistical comparisons were made using Graphpad Prism or R.
Backgrounds carrying PcG mutations occasionally showed suppression of pairing sensitive silencing in late-eclosing flies from vials that were overcrowded, which is similar to observations of sex comb phenotypes induced by other PcG mutations [62]. We did not observe evidence of changes in severity of phenotype for other allelic combinations of eya according to eclosion time or crowding. Nevertheless, we avoided overcrowding and did not score flies beyond day 5 of eclosion.
Identification of molecular lesions in eya alleles
Strategies and analysis for the identification of molecular lesions in alleles of eya are detailed in S1 File.
RNA in situ hybridization
To create transcript-specific RNA probes, exon 1 of the eya-A transcript and exon 1 of the eya-B transcript were each amplified from genomic DNA using primer pairs eyaISA1F/eyaISA1R and eyaISB1F/eyaISB1R, respectively (Primer sequences are provided in S1 Table). For both primer pairs, the reverse primer included a 5’ extension carrying the promoter for T7 RNA polymerase. PCR products were purified using a PCR Purification kit (Qiagen), and 1 μg of each PCR product was used as a template to create digoxygenin-labeled RNA probes using a Dig RNA Labelling Kit (Roche). The products of the reaction were ethanol precipitated and resuspended in 250 μl of 50% formamide/50% TE with 0.1% Tween-20.
For in situ hybridization, eye-antennal discs were dissected in PBS, transferred to a 1.5ml microcentrifuge tube, fixed in 4% formaldehyde in PBS on ice for 20 minutes, then fixed further in 4% formaldehyde/PBS with 0.6% Triton X-100 at room temperature for 20 minutes. After washing in PBS + 0.6% Triton X-100 (3 x 5 min), discs were rinsed with 50% PBS/50% Hybridization Buffer (HB, 50% formamide, 2X SSC, 1X Denhardt’s, 250 μg/ml tRNA, 250 μg/ml salmon sperm DNA, 50 μg/ml heparin sulfate, 5% dextran sulfate, 0.1% Tween-20), then pre-hybridized for 1 hour in 500 μl HB at 52°C. Discs were then incubated overnight in HB with a 1:100 dilution of digoxygenin-labeled probe at 52°C with agitation, followed by four changes of wash solution (50% formamide/2xSSC/0.1% Tween-20) over the next 24 hours at 52°C. Discs were rinsed with PBT (PBS + 0.1% Triton X-100), then incubated in PBT for 30 minutes at room temperature. Next, anti-digoxygenin antibody conjugated to horseradish peroxidase (HRP) (Abcam) was added at a dilution of 1:500, and discs were incubated overnight at 4°C. After four 20-minute washes in PBT at room temperature, discs were developed with a TSA-Plus Cy3 detection kit (Perkin Elmer NEL744E001KT), washed in PBT (3 x 5 minutes) and mounted in Fluoromount G (Electron Microscopy Services). Discs were visualized on either a Zeiss Axio Imager.A2 fluorescence microscope with an AxioCam MRm camera and Zen software, or a Leica SP8 confocal microscope with LASX software.
Quantitative RT-PCR
Assessment of eya mRNA levels via quantitative RT-PCR was carried out as previously described [10]. Briefly, for each sample, 20 imaginal discs were dissected from wandering third instar larvae and frozen at -80°C. Tissue homogenization, genomic DNA elimination, and RNA purification were carried out using an RNeasy plus kit (Qiagen) according to the manufacturer’s protocol. PCR was performed on a StepOne Real-Time PCR system (Applied Biosystems) using cDNA diluted 1:5 into SYBR green PCR Mastermix (Applied Biosystems). Primers were designed to specifically amplify the first exon of either the B transcript (primers eyaRTF1 and eyaRTR1) or the A transcript (primers eyaRT_AF1 and eyaRT_AR1). For discs from CRISPR-edited flies, primers RP49-58F and RP49-175R were used to amplify the housekeeping rp49 cDNA as an internal reference [10]. For discs wherein eye development depended on transvection, RP49 does not present a suitable internal control due to the varying levels of eye tissue relative to the remaining tissues in the disc; in these experiments, the eya-B transcript was used as an internal reference for eya-A to provide a relative measure of eya-A:eya-B transcription. Relative levels of transcript were calculated via the ΔΔCt method using StepOne software.
CRISPR genome editing
To generate a deletion of the eye-specific eya exon 1B, primers were designed to create guide RNAs complementary to sequences 64–84 bp upstream of nucleotide +1 of exon 1B and 537–557 bp downstream of the last nucleotide of exon 1B, spanning roughly 1.1 kb of genomic DNA in total (S1 Table). Guide RNAs were cloned into the plasmid pU6-BbsI-chiRNA as previously described [63], and a mixture of two plasmids carrying upstream- and downstream-targeting guide RNAs (250 ng/μl each) was injected into embryos expressing Cas9 under the control of the Actin5C promoter [64] by BestGene, Inc. From 200 injected embryos, 96 G0 adults eclosed and were crossed to flies carrying the second chromosome balancer CyO. 89 fertile G0 adults were subsequently tested for evidence of an exon 1B deletion via PCR using primers eyaCRISPR34check_F and eyaCRISPR34check_R, which flank the region to be deleted via non-homologous end joining of double strand breaks; 68 (76.4%) of these PCRs produced a single 2.5 kb PCR fragment consistent with unmodified wt DNA, whereas 21 (23.6%) of the PCRs produced additional smaller fragments indicative of putative deletions. Of the 21 G0 flies carrying candidate deletions, four were found to transmit the deletion through the germline, and isogenic stocks were established from three of these. Sequencing of PCR fragments generated from each stock confirmed that each carries a deletion of roughly 1.1 kb spanning the distance between the two guide RNAs and including exon 1B and its promoter.
Supporting information
(DOCX)
Top, eyaD1 fails to complement the eye phenotypes of Class A and Class B alleles. Below, strategy for characterization of eyaD1 using allele-specific PCR. Primer pairs HopFinder_JTR_F2/HopFinder_JTR_R1 (orange) and eya_P_R1_seq/eya_P_F2_seq (blue) show distinct amplification patterns from eya2 and eya4 chromosomes, respectively. PCR from eyaD1/eya2 and eyaD1/eya2 trans-heterozygotes shows no evidence of amplification from the eyaD1 chromosome.
(TIF)
Counts of ommatidia for eya54C2/eya2 and eyaD3/eya2 show complementation consistent with other Class C alleles. Solid line and dark shading represent mean ommatidia counts for eya2 complementation by Class B alleles, dashed line and light shading represent mean ommatidia counts for eya2 complementation by other Class C alleles (see Fig 3). Both alleles completely fail to complement eya4. Data represent n = 20 eyes for each genotype.
(TIF)
Counts of ommatidia are significantly higher in eya2/eya4; Cap-H20019/Cap-H25163, carrying a strong trans-heterozygous loss of function combination of Cap-H2 mutations, than in eya2/eya4; Cap-H20019/+, which carries one wild-type copy of Cap-H2 (p = 0.003, Mann-Whitney test), or eya2/eya4, where both copies of Cap-H2 are wild type (p = 0.02).
(TIF)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
We thank Nancy Bonini, Judy Kassis, Gio Bosco, Justin Kumar, Denise Montell, Jennifer Jemc Mierisch, and the Bloomington Drosophila Stock Center for fly stocks, advice, and stimulating conversations.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by funding to JRB: Faculty Early Development (CAREER) award 1349779, National Science Foundation, www.nsf.gov; National Institute of General Medical Sciences of the National Institutes of Health P20 GM0103423, www.nih.gov The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Long HK, Prescott SL, Wysocka J. Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution. Cell. 2016;167(5):1170–87. Epub 2016/11/20. 10.1016/j.cell.2016.09.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Shlyueva D, Stampfel G, Stark A. Transcriptional enhancers: from properties to genome-wide predictions. Nat Rev Genet. 2014;15(4):272–86. Epub 2014/03/13. 10.1038/nrg3682 . [DOI] [PubMed] [Google Scholar]
- 3.Schuettengruber B, Bourbon HM, Di Croce L, Cavalli G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell. 2017;171(1):34–57. Epub 2017/09/25. 10.1016/j.cell.2017.08.002 . [DOI] [PubMed] [Google Scholar]
- 4.Kassis JA, Kennison JA, Tamkun JW. Polycomb and Trithorax Group Genes in Drosophila. Genetics. 2017;206(4):1699–725. Epub 2017/08/06. 10.1534/genetics.115.185116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Joyce EF, Erceg J, Wu CT. Pairing and anti-pairing: a balancing act in the diploid genome. Curr Opin Genet Dev. 2016;37:119–28. Epub 2016/04/12. 10.1016/j.gde.2016.03.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lewis EB. The Theory and Application of a New Method of Detecting Chromosomal Rearrangements in Drosophila melanogaster. The American Naturalist. 1954;88(841):225. [Google Scholar]
- 7.Kassis J. Pairing-sensitive silencing, polycomb group response elements, and transposon homing in Drosophila. Homology Effects. Advances in Genetics 2002. p. 421–38. [DOI] [PubMed] [Google Scholar]
- 8.Geyer PK, Green MM, Corces VG. Tissue-specific transcriptional enhancers may act in trans on the gene located in the homologous chromosome: the molecular basis of transvection in Drosophila. EMBO J. 1990;9(7):2247–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Morris JR, Chen J, Filandrinos ST, Dunn RC, Fisk R, Geyer PK, et al. An analysis of transvection at the yellow locus of Drosophila melanogaster. Genetics. 1999;151(2):633–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bateman JR, Johnson JE, Locke MN. Comparing enhancer action in cis and in trans. Genetics. 2012;191(4):1143–55. Epub 2012/06/01. 10.1534/genetics.112.140954 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Casares F, Bender W, Merriam J, Sanchez-Herrero E. Interactions of Drosophila Ultrabithorax regulatory regions with native and foreign promoters. Genetics. 1997;145(1):123–37. Epub 1997/01/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gohl D, Muller M, Pirrotta V, Affolter M, Schedl P. Enhancer blocking and transvection at the Drosophila apterous locus. Genetics. 2008;178(1):127–43. Epub 2008/01/19. 10.1534/genetics.107.077768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Martinez-Laborda A, Gonzalez-Reyes A, Morata G. Trans regulation in the Ultrabithorax gene of Drosophila: alterations in the promoter enhance transvection. The EMBO journal. 1992;11(10):3645–52. papers2://publication/uuid/880FF385-876B-4452-8071-FC0A476C67CA. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mellert DJ, Truman JW. Transvection is common throughout the Drosophila genome. Genetics. 2012;191(4):1129–41. Epub 2012/06/01. 10.1534/genetics.112.140475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Davis TL, Rebay I. Master regulators in development: Views from the Drosophila retinal determination and mammalian pluripotency gene networks. Developmental biology. 2017;421(2):93–107. Epub 2016/12/17. 10.1016/j.ydbio.2016.12.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bonini NM, Leiserson WM, Benzer S. The eyes absent gene: genetic control of cell survival and differentiation in the developing Drosophila eye. Cell. 1993;72(3):379–95. . [DOI] [PubMed] [Google Scholar]
- 17.Bonini NM, Bui QT, Gray-Board GL, Warrick JM. The Drosophila eyes absent gene directs ectopic eye formation in a pathway conserved between flies and vertebrates. Development. 1997;124(23):4819–26. Epub 1998/01/15. . [DOI] [PubMed] [Google Scholar]
- 18.Kumar JP. The molecular circuitry governing retinal determination. Biochim Biophys Acta. 2009;1789(4):306–14. Epub 2008/11/18. 10.1016/j.bbagrm.2008.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bonini NM, Leiserson WM, Benzer S. Multiple roles of the eyes absent gene in Drosophila. Developmental biology. 1998;196(1):42–57. 10.1006/dbio.1997.8845 . [DOI] [PubMed] [Google Scholar]
- 20.Leiserson WM, Benzer S, Bonini NM. Dual functions of the Drosophila eyes absent gene in the eye and embryo. Mech Dev. 1998;73(2):193–202. . [DOI] [PubMed] [Google Scholar]
- 21.Weyers JJ, Milutinovich AB, Takeda Y, Jemc JC, Van Doren M. A genetic screen for mutations affecting gonad formation in Drosophila reveals a role for the slit/robo pathway. Developmental biology. 2011;353(2):217–28. 10.1016/j.ydbio.2011.02.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bai J. Eyes Absent, a key repressor of polar cell fate during Drosophila oogenesis. Development. 2002;129(23):5377–88. 10.1242/dev.00115 [DOI] [PubMed] [Google Scholar]
- 23.Xiong W, Dabbouseh NM, Rebay I. Interactions with the Abelson tyrosine kinase reveal compartmentalization of eyes absent function between nucleus and cytoplasm. Dev Cell. 2009;16(2):271–9. 10.1016/j.devcel.2008.12.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zimmerman JE, Bui QT, Liu H, Bonini NM. Molecular genetic analysis of Drosophila eyes absent mutants reveals an eye enhancer element. Genetics. 2000;154(1):237–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Karandikar UC, Jin M, Jusiak B, Kwak S, Chen R, Mardon G. Drosophila eyes absent is required for normal cone and pigment cell development. PLoS One. 2014;9(7):e102143 10.1371/journal.pone.0102143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Weasner BM, Weasner BP, Neuman SD, Bashirullah A, Kumar JP. Retinal Expression of the Drosophila eyes absent Gene Is Controlled by Several Cooperatively Acting Cis-regulatory Elements. PLoS Genet. 2016;12(12):e1006462 10.1371/journal.pgen.1006462 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Leiserson WM, Bonini NM, Benzer S. Transvection at the eyes absent gene of Drosophila. Genetics. 1994;138(4):1171–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bui QT, Zimmerman JE, Liu H, Bonini NM. Molecular analysis of Drosophila eyes absent mutants reveals features of the conserved Eya domain. Genetics. 2000;155(2):709–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hartl TA, Smith HF, Bosco G. Chromosome alignment and transvection are antagonized by condensin II. Science. 2008;322(5906):1384–7. 10.1126/science.1164216 . [DOI] [PubMed] [Google Scholar]
- 30.Joyce EF, Williams BR, Xie T, Wu CT. Identification of genes that promote or antagonize somatic homolog pairing using a high-throughput FISH-based screen. PLoS Genet. 2012;8(5):e1002667 10.1371/journal.pgen.1002667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Celniker SE, Dillon LA, Gerstein MB, Gunsalus KC, Henikoff S, Karpen GH, et al. Unlocking the secrets of the genome. Nature. 2009;459(7249):927–30. 10.1038/459927a [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Filion GJ, van Bemmel JG, Braunschweig U, Talhout W, Kind J, Ward LD, et al. Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell. 2010;143(2):212–24. Epub 2010/10/05. 10.1016/j.cell.2010.09.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brown JL, Sun MA, Kassis JA. Global changes of H3K27me3 domains and Polycomb group protein distribution in the absence of recruiters Spps or Pho. Proc Natl Acad Sci U S A. 2018;115(8):E1839–E48. Epub 2018/02/13. 10.1073/pnas.1716299115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lum TE, Merritt TJ. Nonclassical regulation of transcription: interchromosomal interactions at the malic enzyme locus of Drosophila melanogaster. Genetics. 2011;189(3):837–49. Epub 2011/09/09. 10.1534/genetics.111.133231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Blick AJ, Mayer-Hirshfeld I, Malibiran BR, Cooper MA, Martino PA, Johnson JE, et al. The Capacity to Act in Trans Varies Among Drosophila Enhancers. Genetics. 2016;203(1):203–18. Epub 2016/03/18. 10.1534/genetics.115.185645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen JL, Huisinga KL, Viering MM, Ou SA, Wu CT, Geyer PK. Enhancer action in trans is permitted throughout the Drosophila genome. Proc Natl Acad Sci USA. 2002;99(6):3723–8. 10.1073/pnas.062447999 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kravchenko E, Savitskaya E, Kravchuk O, Parshikov A, Georgiev P, Savitsky M. Pairing between gypsy insulators facilitates the enhancer action in trans throughout the Drosophila genome. Mol Cell Biol. 2005;25(21):9283–91. Epub 2005/10/18. 10.1128/MCB.25.21.9283-9291.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Callaerts P, Leng S, Clements J, Benassayag C, Cribbs D, Kang YY, et al. Drosophila Pax-6/eyeless is essential for normal adult brain structure and function. J Neurobiol. 2001;46(2):73–88. Epub 2001/01/12. . [DOI] [PubMed] [Google Scholar]
- 39.Cheyette BN, Green PJ, Martin K, Garren H, Hartenstein V, Zipursky SL. The Drosophila sine oculis locus encodes a homeodomain-containing protein required for the development of the entire visual system. Neuron. 1994;12(5):977–96. Epub 1994/05/01. . [DOI] [PubMed] [Google Scholar]
- 40.Serikaku MA, O'Tousa JE. sine oculis is a homeobox gene required for Drosophila visual system development. Genetics. 1994;138(4):1137–50. Epub 1994/12/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Johnston RJ Jr., Desplan C. Interchromosomal communication coordinates intrinsically stochastic expression between alleles. Science. 2014;343(6171):661–5. Epub 2014/02/08. 10.1126/science.1243039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Main BJ, Smith AD, Jang H, Nuzhdin SV. Transcription start site evolution in Drosophila. Mol Biol Evol. 2013;30(8):1966–74. Epub 2013/05/08. 10.1093/molbev/mst085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hoskins RA, Landolin JM, Brown JB, Sandler JE, Takahashi H, Lassmann T, et al. Genome-wide analysis of promoter architecture in Drosophila melanogaster. Genome Res. 2011;21(2):182–92. Epub 2010/12/24. 10.1101/gr.112466.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Drosophila 12 Genomes C, Clark AG, Eisen MB, Smith DR, Bergman CM, Oliver B, et al. Evolution of genes and genomes on the Drosophila phylogeny. Nature. 2007;450(7167):203–18. Epub 2007/11/13. 10.1038/nature06341 . [DOI] [PubMed] [Google Scholar]
- 45.Thurmond J, Goodman JL, Strelets VB, Attrill H, Gramates LS, Marygold SJ, et al. FlyBase 2.0: the next generation. Nucleic Acids Res. 2019;47(D1):D759–D65. Epub 2018/10/27. 10.1093/nar/gky1003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Negre N, Brown CD, Shah PK, Kheradpour P, Morrison CA, Henikoff JG, et al. A comprehensive map of insulator elements for the Drosophila genome. PLoS Genet. 2010;6(1):e1000814 Epub 2010/01/20. 10.1371/journal.pgen.1000814 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kwon D, Mucci D, Langlais KK, Americo JL, DeVido SK, Cheng Y, et al. Enhancer-promoter communication at the Drosophila engrailed locus. Development. 2009;136(18):3067–75. Epub 2009/08/14. 10.1242/dev.036426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dillon N, Trimborn T, Strouboulis J, Fraser P, Grosveld F. The effect of distance on long-range chromatin interactions. Mol Cell. 1997;1(1):131–9. Epub 1998/07/11. . [DOI] [PubMed] [Google Scholar]
- 49.Kmita M, Fraudeau N, Herault Y, Duboule D. Serial deletions and duplications suggest a mechanism for the collinearity of Hoxd genes in limbs. Nature. 2002;420(6912):145–50. Epub 2002/11/15. 10.1038/nature01189 . [DOI] [PubMed] [Google Scholar]
- 50.Fukaya T, Lim B, Levine M. Enhancer Control of Transcriptional Bursting. Cell. 2016;166(2):358–68. Epub 2016/06/14. 10.1016/j.cell.2016.05.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Leader DP, Krause SA, Pandit A, Davies SA, Dow JAT. FlyAtlas 2: a new version of the Drosophila melanogaster expression atlas with RNA-Seq, miRNA-Seq and sex-specific data. Nucleic Acids Res. 2018;46(D1):D809–D15. Epub 2017/10/27. 10.1093/nar/gkx976 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hoskins RA, Carlson JW, Wan KH, Park S, Mendez I, Galle SE, et al. The Release 6 reference sequence of the Drosophila melanogaster genome. Genome Res. 2015;25(3):445–58. Epub 2015/01/16. 10.1101/gr.185579.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kharchenko PV, Alekseyenko AA, Schwartz YB, Minoda A, Riddle NC, Ernst J, et al. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. Nature. 2011;471(7339):480–5. Epub 2010/12/24. 10.1038/nature09725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Schwartz YB, Kahn TG, Nix DA, Li XY, Bourgon R, Biggin M, et al. Genome-wide analysis of Polycomb targets in Drosophila melanogaster. Nat Genet. 2006;38(6):700–5. Epub 2006/05/30. 10.1038/ng1817 . [DOI] [PubMed] [Google Scholar]
- 55.Erceg J, Pakozdi T, Marco-Ferreres R, Ghavi-Helm Y, Girardot C, Bracken AP, et al. Dual functionality of cis-regulatory elements as developmental enhancers and Polycomb response elements. Genes Dev. 2017;31(6):590–602. Epub 2017/04/07. 10.1101/gad.292870.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Schuettengruber B, Ganapathi M, Leblanc B, Portoso M, Jaschek R, Tolhuis B, et al. Functional anatomy of polycomb and trithorax chromatin landscapes in Drosophila embryos. PLoS Biol. 2009;7(1):e13 Epub 2009/01/16. 10.1371/journal.pbio.1000013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Janody F, Lee JD, Jahren N, Hazelett DJ, Benlali A, Miura GI, et al. A mosaic genetic screen reveals distinct roles for trithorax and polycomb group genes in Drosophila eye development. Genetics. 2004;166(1):187–200. Epub 2004/03/17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bessa J, Gebelein B, Pichaud F, Casares F, Mann RS. Combinatorial control of Drosophila eye development by eyeless, homothorax, and teashirt. Genes Dev. 2002;16(18):2415–27. Epub 2002/09/17. 10.1101/gad.1009002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhu J, Ordway AJ, Weber L, Buddika K, Kumar JP. Polycomb group (PcG) proteins and Pax6 cooperate to inhibit in vivo reprogramming of the developing Drosophila eye. Development. 2018;145(7). Epub 2018/03/14. 10.1242/dev.160754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ray P, De S, Mitra A, Bezstarosti K, Demmers JA, Pfeifer K, et al. Combgap contributes to recruitment of Polycomb group proteins in Drosophila. Proc Natl Acad Sci U S A. 2016;113(14):3826–31. Epub 2016/03/24. 10.1073/pnas.1520926113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Davis TL, Rebay I. Pleiotropy in Drosophila organogenesis: Mechanistic insights from Combgap and the retinal determination gene network. Fly (Austin). 2018;12(1):62–70. Epub 2017/11/11. 10.1080/19336934.2017.1402994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kennison JA, Russell MA. Dosage-Dependent Modifiers of Homoeotic Mutations in Drosophila melanogaster. Genetics. 1987;116(1):75–86. Epub 1987/05/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gratz SJ, Cummings AM, Nguyen JN, Hamm DC, Donohue LK, Harrison MM, et al. Genome engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease. Genetics. 2013;194(4):1029–35. Epub 2013/05/28. 10.1534/genetics.113.152710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Port F, Chen HM, Lee T, Bullock SL. Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc Natl Acad Sci U S A. 2014;111(29):E2967–76. Epub 2014/07/09. 10.1073/pnas.1405500111 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
Top, eyaD1 fails to complement the eye phenotypes of Class A and Class B alleles. Below, strategy for characterization of eyaD1 using allele-specific PCR. Primer pairs HopFinder_JTR_F2/HopFinder_JTR_R1 (orange) and eya_P_R1_seq/eya_P_F2_seq (blue) show distinct amplification patterns from eya2 and eya4 chromosomes, respectively. PCR from eyaD1/eya2 and eyaD1/eya2 trans-heterozygotes shows no evidence of amplification from the eyaD1 chromosome.
(TIF)
Counts of ommatidia for eya54C2/eya2 and eyaD3/eya2 show complementation consistent with other Class C alleles. Solid line and dark shading represent mean ommatidia counts for eya2 complementation by Class B alleles, dashed line and light shading represent mean ommatidia counts for eya2 complementation by other Class C alleles (see Fig 3). Both alleles completely fail to complement eya4. Data represent n = 20 eyes for each genotype.
(TIF)
Counts of ommatidia are significantly higher in eya2/eya4; Cap-H20019/Cap-H25163, carrying a strong trans-heterozygous loss of function combination of Cap-H2 mutations, than in eya2/eya4; Cap-H20019/+, which carries one wild-type copy of Cap-H2 (p = 0.003, Mann-Whitney test), or eya2/eya4, where both copies of Cap-H2 are wild type (p = 0.02).
(TIF)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.