

Abstract
Individual differences in maternal sensitivity to infant distress were examined by testing whether mothers’ physiological responses interacted to predict sensitivity differently in a fear task relative to an anger task, and based on the level of distress displayed by the infant. Mothers’ (N = 208) physiological arousal (SCL arousal) and physiological regulation (RSA withdrawal/augmentation) were measured during a baseline task and while interacting with their 1-year-old infants during tasks that elicited infant fear and infant anger. Infant negative affect and maternal sensitivity were assessed during these interactions, and mothers later reported on their perceptions of infant distress during the tasks. Results indicated that the mother SCL arousal × RSA change interaction predicted sensitivity only with infants who displayed higher distress, and in a different pattern across tasks. In the fear task, for mothers of infants displaying higher distress, SCL arousal related to greater sensitivity only for mothers who displayed RSA withdrawal. In the anger task, for mothers of infants displaying higher distress, SCL arousal related to greater sensitivity only for mothers who displayed RSA augmentation. Further, although infants displayed similar levels of distress in each task, maternal SCL arousal, sensitivity, and perceptions of distress differed across tasks. These results suggest that contextual cues and infant distress cues both contribute to associations between mothers’ physiological responses and maternal sensitivity.
Keywords: parenting, sensitivity, physiology, anger, fear
Maternal sensitivity to infant distress has demonstrated benefits for early socioemotional development (Leerkes, Blankson, & O’Brien, 2012; McElwain & Booth-LaForce, 2006); thus, it is important to understand what qualities or circumstances support sensitive mother-infant interactions. The current study examined several factors that may lead to variation in maternal sensitivity to infant distress, including mothers’ physiological responses to infant distress (SCL arousal and RSA change), the level of distress displayed by the infant, and the type of distress context in which the mother-infant interaction takes place (fear vs. anger). The primary goals of this study were to determine (1) if the interaction of maternal SCL arousal and RSA change is a stronger predictor of maternal sensitivity among mothers whose infants are more highly distressed, and (2) if the nature of the interaction between maternal physiological responses predicting maternal sensitivity varies in a fear context vs. anger context.
Physiological Arousal and Regulation as Predictors of Sensitivity
Mother’s physiological responses to infant distress appear to be meaningful predictors of maternal sensitivity. This study focused on measures of mothers’ autonomic nervous system reactivity. One factor of interest in the current study is an increase in skin conductance level (SCL), which may be considered an indicator of physiological arousal as it is thought to reflect activation of the sympathetic nervous system (SNS) related to emotional arousal and responsivity to aversive stimuli (Gray, 1975). Previous studies have suggested that such sympathetic arousal to infant crying may lead to lower sensitivity, as SCL arousal in response to persistent infant crying and aversive child behavior appears to underlie risk for harsh discipline and child maltreatment (Joosen, Mesman, Bakermans-Kranenburg, & van IJzendoorn, 2013; Wolfe, Fairbank, Kelly, & Bradlyn, 1983). On the other hand, at least one study has indicated that a lack of SCL arousal to infant distress may reflect disengagement from the interaction, and thus also pose risks for maltreatment (Reijman et al., 2014). In contrast, respiratory sinus arrhythmia (RSA) withdrawal, or vagal withdrawal, may be expected to relate to greater sensitivity. Although RSA withdrawal also reflects reactivity to environmental stimuli, it is often labeled as an indicator of regulatory processes at the physiological level, as it is thought to reflect a withdrawal of parasympathetic nervous system (PNS) influence on the heart that allows for coping or regulation in response to environmental challenge (Mills-Koonce et al., 2007; Porges, 2007). Accordingly, previous studies have indicated that RSA withdrawal in response to infant crying and during challenging parent-child interactions may support more positive parenting behavior (Ablow, Marks, Feldman, & Huffman, 2013; Joosen, Mesman, Bakermans-Kranenburg, Pieper, et al., 2013; Mills-Koonce et al., 2007; Moore et al., 2009).
Informed by the perspective that SNS and PNS responses may operate independently (see Bernston, Cacioppo, & Quigley, 1993), more recent work has considered the possibility that measures of physiological arousal and regulation may actually co-act to predict maternal sensitivity. Leerkes and colleagues (Leerkes, Su, Calkins, Supple, & O’Brien, 2016; Leerkes et al., 2015) hypothesized that if a mother is physiologically aroused but well-regulated in the presence of infant distress, this may allow for a motivation to focus less on her own needs in favor of her infants’ needs, which increases the likelihood of sensitive responses in these mothers in contrast to those who are physiologically aroused but poorly regulated. In this framework, the mixed evidence for the role of SCL arousal in (in)sensitive parenting may exist because its influence depends on other physiological mechanisms. In fact, utilizing data from the sample examined in the current study, they found that maternal SCL arousal and RSA withdrawal assessed both prenatally and at 6 months postpartum significantly interacted to predict maternal sensitivity via maternal cognitions about crying. As predicted, arousal related to more adaptive attributions about crying only for mothers who displayed greater RSA withdrawal, and attributions in turn related to sensitivity (Leerkes et al., 2015; 2016).
Other researchers have considered relative influences of SNS and PNS reactivity as predictors of parenting. Sturge-Apple, Skibo, Rogosch, Ignjatovic, and Heinzelman (2011) predicted parenting during a free play task from profiles of mothers’ sympathovagal balance (reflecting relative dominance of SNS to PNS response) through the course of a task intended to be stressful for their toddlers. A moderate reactivity class (relative balance of SNS and PNS activity), displayed the least negative parenting behavior. In contrast, a hyperarousal class (relatively higher SNS activity and lower PNS activity) displayed more harsh, intrusive, and insensitive parenting behavior. A hypoarousal class (relatively higher PNS activity and lower SNS activity), displayed the highest levels of insensitive and disengaged behavior. Similarly, Miller, Kahle, Lopez and Hastings (2015) argued that maternal autonomic coactivation (i.e., high SNS and PNS activation) would lead to supportive social engagement during challenging interactions. They found that greater coactivation related to less harsh or negative parenting during tasks designed to frustrate preschool-aged children.
Collectively, these results suggest that a balance or co-occurrence of sympathetic and parasympathetic responses may predict more adaptive parenting behavior during tasks designed to elicit infant and child distress. Nonetheless, these studies collapsed measures across a range of tasks, or focused on tasks that tended to elicit one form of distress, rather than considering mothers’ responses within different types of distress contexts. It is possible that certain salient characteristics of mother-infant interactions could influence mothers’ perceptions of infant distress, with implications for the association between mothers’ physiological responses and sensitivity. The current study used a framework similar to Leerkes and colleagues, hypothesizing that mothers’ SCL and RSA responses to infant distress interact to predict sensitivity, but examined how two such salient characteristics of mother-infant interaction may alter this association: the level of distress experienced by the infant in the moment, as well as the type of distress context in which the mother-infant interaction takes place.
Infant Distress Level
Infants typically depend on mothers and other caregivers in order to regulate their distress but, consistent with temperament theory, the same stimulus may evoke a different magnitude of emotional reaction in different infants (Calkins, 1994; Rothbart & Bates, 2006). A higher level of distress in the infant may serve as a more impactful stimulus to the parent regarding the infant’s emotional needs, and a stronger basis of emotional cues to which the mother may react at the physiological level and then behavioral level. Thus, the role of mothers’ physiological responses in predicting sensitivity may be moderated by infant distress level, such that their associations with sensitivity are stronger when infants display greater distress. Previous studies have found that greater infant negative affect related to less maternal sensitivity overall (e.g., Leerkes et al., 2015; 2016), potentially more so among mothers with low RSA withdrawal (Mills-Koonce et al., 2007). However, these studies considered physiological responses with younger infants. Given growth and differentiation in infant emotion across the first year of life (Izard, 1978), individual differences in infant distress may be more meaningful to account for when examining relations between mothers’ physiological responses and sensitivity later in infancy than at younger ages. The current study focused on observations when infants were one year of age.
Infant Distress Context
Another important consideration is that mother-infant interactions surrounding infant distress do not all occur under identical circumstances. Different situations are expected to elicit different types of infant emotion (see Goldsmith, Reilly, Lemery, Longley, & Prescott, 1994; Izard, 1978). For example, certain situations are associated with fear responses, such as when infants are exposed to unfamiliar, unpredictable, or threatening stimuli. Others may be associated with anger or frustration, such as when infants are blocked from accessing attractive stimuli. If mothers perceive infant distress differently based on contextual cues, then their physiological responses may relate to sensitivity in a different manner. These perceptual distinctions could also be important from a development standpoint, as anger and fear tend to increase across the first year of life but are not necessarily highly associated (Braungart-Rieker, Hill-Soderlund, & Karrass, 2010; Buss & Goldsmith, 1998; Izard, 1978). Mothers may thus increasingly discern how varying stimuli elicit different forms of distress over the first year of their infants’ lives.
At a conceptual level, researchers acknowledge that emotions should be understood based on their eliciting context, and that parents’ appraisal of situational characteristics and inferences about causes of child behavior both contribute to their momentary parenting goals and to their emotional and behavioral responses to child cues (Cole, Dennis, & Martin, 2004; Dix, 1992). Research suggests that infants’ facial affect does not reliably conform to discrete negative emotion expressions like those in adults; instead, sensitive caregivers are expected to rely on both affective and contextual cues to understand and address infant distress (Oster, Mesman, & Camras, 2012). Additionally, domain-specific approaches to socialization research suggest mechanisms of parenting differ by domains of interaction (Grusec and Davidov, 2010). Applying that perspective, one could argue that infant distress generally motivates the protection domain of parenting, in which parents seek to provide help and comfort and to support appropriate responses to stressors. However, infant fear may more clearly fall into this domain given it is typically elicited with more salient cues of threat. Comparatively, infant anger could activate the guided learning domain, in which parents seek to support the child’s learning of cognitive, physical, social and emotional skills necessary to function well in the social group, for example, by helping the child to manage frustration or to manage a difficult task at hand. Anger may also elicit the control domain in some parents, such that they perceive a conflict between personal and child goals and seek to change child behavior to align with parental goals. This could be the case if mothers perceive infant anger as aversive or antisocial and thus seek to prevent its occurrence.
Porges (2007) also articulated a potential role of situational context in vagal withdrawal and augmentation responses and/or the behaviors these responses motivate. The polyvagal theory proposes the idea of “neuroception” such that the individual’s perception of specific features of the environment may lead to different physiological states that in turn motivate certain types of behavior. Specifically, a physiological state reflecting vagal withdrawal (RSA withdrawal) is thought to support fight-or-flight responses, whereas vagal augmentation is thought to support social engagement responses. These differences could have implications for mothers’ physiological states in response to their infants’ distress. Specifically, in contexts in which mothers perceive cues of infant fear, RSA withdrawal could mobilize maternal flight-or-flight responses relevant to ensuring infant safety and protection. Alternatively, in contexts in which mothers perceive cues of infant anger, maintenance or increase in RSA (i.e., RSA augmentation) could mobilize maternal social engagement responses relevant to assisting infants with problem-solving and goal pursuit. In either case, this specialized vagal response (RSA withdrawal or augmentation) paired with SCL arousal in response to infant distress may predict sensitive responses that are attuned to the needs of the infant within each respective interactive context.
Additionally, mothers’ varying perceptions of infant distress across contexts could have systematic consequences for their reactions at the physiological, cognitive, and behavioral level. First, if mothers perceive fear tasks as eliciting an immediate need for infant protection and survival, they may evidence overall greater physiological arousal than they do in anger tasks, wherein mothers may perceive a need to assist with problem solving and goal pursuit under safe conditions. This could be important to consider if SCL arousal is nonetheless tied to greater sensitivity under certain conditions of RSA change. Secondly, mothers may tend to make less-negative or minimizing attributions about infant distress during fear tasks relative to anger tasks, as mothers may more readily accept infants’ emotional needs related to survival, compared to infant responses that may be viewed as difficult or defiant. Third, given these differing attributions about distress, mothers may display overall greater sensitivity to infant cues during fear tasks relative to anger tasks.
Some researchers have addressed the role of differing situational or distress cues in mothers’ responses to their children’s emotions. In observations of mothers and their toddlers, mothers were found to respond to child fear displays with tenderness above chance level and neutral emotion below chance level, whereas they responded to child anger displays with anger above chance level and happiness below chance level (Denham, 1993). Mothers also endorsed fewer regulatory responses to child anger compared to other emotions, including fear (Casey & Fuller, 1994). It is unclear, however, if these patterns generalize to maternal sensitivity with younger infants. From a physiological standpoint, Mills-Koonce and colleagues (2009) found that RSA change did not moderate the influence of mothers’ stress hormone levels (i.e., resting cortisol) differently across a free play and stressor task in predicting measures of positive and intrusive parenting with their 6-month-old infants. However, Miller et al.’s (2015) findings that SNS and PNS coactivation related to less-negative parenting in situations that may elicit child frustration were based on an argument that PNS activity (i.e., higher RSA) may modulate defensive SNS responses to facilitate social engagement. Nonetheless, to our knowledge, no study has distinguished varying forms of infant distress context (e.g., fear, anger) in the prediction of maternal sensitivity, particularly based on mothers’ physiological responses.
Current Study
The current study examined the interaction of mothers’ physiological arousal and regulation as predictors of maternal sensitivity in a fear context and an anger context, based on infants’ distress level. When infants were one year of age they took part in tasks designed to elicit fear and anger. Mothers were able to interact with their infants after they were exposed to task stimuli, and mothers’ heart period and SCL were recorded while they were observed for maternal sensitivity. Later, mothers watched video recordings of the tasks and rated the emotions they believed their infant felt in the moment and their attributions about their infant’s emotion. These measures were examined concurrently given the expectation that mothers’ responses may be tied to their infants’ in-the-moment distress level, which may not be stable over time.
To address the primary study goals, we examined infant distress and mothers’ SCL arousal and RSA change as predictors of sensitivity in each task, with two general sets of hypotheses. (1) We hypothesized that in both tasks the interaction between SCL arousal and RSA change would predict sensitivity only with infants who displayed higher distress. (2) Further, we hypothesized that in the fear task, for mothers of infants who displayed high distress, SCL arousal would relate to sensitivity for mothers who displayed more RSA withdrawal. However, we hypothesized that in the anger task, for mothers of infants who displayed high distress, SCL arousal would relate to sensitivity for mothers who displayed RSA augmentation.
The above proposed pattern of effects was predicated on the expectation that mothers perceived the fear task differently than the anger task. Thus, we conducted a set of secondary analyses to confirm the existence of any such context differences in infant distress, mother physiological responses, mother sensitivity, as well as mothers’ identification of infant emotions, and mothers’ attributions about infant crying. We hypothesized that, given the nature of the tasks, infants would display similar mean levels of distress across tasks, but mothers would perceive greater infant anger in the anger task and greater infant fear in the fear task. Due to differences in perceptions of infant fear relative to anger, we hypothesized that mothers would report more negative and minimizing attributions about their infants’ emotion during the anger task and display greater physiological arousal and greater sensitivity in the fear task. Further, we hypothesized that greater infant distress would relate to greater maternal SCL arousal and sensitivity in the fear task, whereas it would relate to less sensitivity and more negative and minimizing attributions in the anger task.
Method
Participants
Participants were 208 primiparous mothers and their children who took part in a longitudinal study in the southeastern United States. Mothers were recruited via flyers and/or presentations in birth education classes, breastfeeding classes, local obstetric practices, clinics, or by referral from other participants. Inclusion criteria for the study included being 18 or older, African-American or European American, fluent in English, and expecting their first child. The original sample (N = 259) consisted of mothers who consented to the study and completed an interview and self-report measures prior to the child’s birth. This study was approved by the institutional review board at (institution and study name masked for blind review).
The current study includes data from laboratory observations of the mother and child when the child was approximately 1 year of age (M = 13.9 months, SD = .98). This study sample included 107 European American mothers, 91 African American mothers, and 10 who self-identified as more than one race. They ranged from 18–44 years of age at the start of the study, M = 25. In terms of education level, 26% of mothers had a high school education or less, 32% had some college up to a 2-year degree, 32% earned a 4-year degree or beyond. Self-reported annual family income ranged from under $2,000 to over $100,000, Median = $35,000. The majority (60%) of mothers were married or living with a partner, 23% were in a relationship but not living with a partner, and 17% were single or divorced. All infants were full term and healthy; 100 (48%) were male and 108 (52%) were female; 102 were European American/White and 106 were non-White (97 African American, 9 multi-racial) based on maternal report.
Participants who did not complete a laboratory visit at the 1-year time point did so primarily because they could not be reached, had moved from the area, or did not have time available to participate. Attrition analyses suggested that those who participated in the 1-year observation did not differ from those who did not on mother race, marital status, or income-to-needs ratio; however, mothers who participated were somewhat older (years at child’s birth M = 25.34, SD = 5.40 compared to M = 23.88, SD =5.403, t(256) = 1.73, p = .09) and reported higher levels of education (M = 3.92, SD = 1.80 compared to M = 3.33, SD = 1.68 on a scale in which 3 = some college and 4 = 2-year college degree, t(255) = 2.12, p = .04).
Procedures
Physiological data collection.
A fetrode was placed on two fingers on mothers’ non-dominant hand to detect electrodermal activity (i.e., skin conductance response) and three disposable electrodes, one on each ribcage and one on the collarbone, were placed on mothers for R-wave detection (i.e., heart period collection). This equipment was connected to a Biolog (UFI, Morro Bay, CA) and worn by mothers throughout the mother-child interaction tasks. An experimenter pressed an event marker on a key fob to signal the start and stop time for each portion of the laboratory tasks with the Biolog.
Mother-infant interaction tasks.
As a baseline task for measuring change in mothers’ physiological responses during the distress tasks, mothers were asked to play with their child as they normally would for 7 minutes using toys set up in the laboratory. Mothers and children then engaged in tasks used to elicit infant fear and anger and assess maternal sensitivity. A toy removal task was utilized to elicit infant anger. The experimenter allowed the child to play with a toy phone for 1 minute, removed the toy and placed it in a clear plastic jar the child was unable to open, then repeatedly prompted the child to get the phone for 4 minutes. A novel person approach was utilized to elicit fear. An experimenter wearing a green ogre costume entered the laboratory and repeatedly approached and withdrew from within 2 feet of the infant while stating a standard script (“Hi, [Child]. What are you doing? I’m an ogre. Do you know what an ogre is?...”) for 4 minutes. In each task, the mother was asked to remain neutral for 1 minute, then could interact with the infant for 3 minutes however she liked except for taking the phone out of the jar or speaking to or touching the ogre. Infants who were distressed by a task were soothed by the mother and/or experimenter before beginning the next task in order to reduce carry-over of infant distress. As such, relatively few infants failed to complete these tasks (2 for the anger task because they were uninterested and 2 for the fear task because they were inconsolable).
Mother emotion interview.
After completing the mother-child interaction tasks, mothers completed an interview about their experiences during each task. Relevant to the current study, mothers were asked to rate how strongly her infant felt irritated/annoyed, frustrated, aggravated, nervous/anxious, angry/mad, worried, disgusted, and afraid/scared on a 4-point scale from 1 = not at all to 4 = very strongly. Mothers were also asked to rate the extent to which she agreed with explanations for why her infant was crying on a 4-point scale from 1 = strongly disagree to 4 = strongly agree. The current study utilized 7 items that reflect negative attributions, including spoiled, difficult temperament, trying to make my life difficult, unreasonable, crying on purpose, selfish, just wanted attention, and 5 items that reflect minimizing attributions, including having a bad day, in a bad mood, tired, hungry, not feeling well (Leerkes & Siepak, 2006).
Measures
Behavioral coding.
The mother-child interaction tasks were continuously coded from digital media files using INTERACT 9 (Mangold, Arnstorf, Germany). Event based coding was used, meaning once a code was activated, it remained active until another code was selected. For all behavioral coding, coders were blind to other data, reliability cases were selected at random, and disagreements were resolved via consensus. Data for the current study were drawn from the portion of the fear and anger tasks in which mothers were allowed to interact with their infants.
Infant distress.
Infant affect in each task was continuously rated on a 7-point scale from 1 = high positive affect to 7 = high negative affect based on vocalizations, facial expressions, and body tension (adapted from Braungart-Rieker & Stifter, 1996). Interrater reliability was assessed on 15% of cases, weighted kappa = .75. Ninety-one percent of infants became distressed (i.e., displayed negative affect, defined as an affect level ≥5) at any point during the distress tasks. Mean scores were calculated across each task, with higher scores reflecting greater distress.
Maternal sensitivity.
Maternal sensitivity was rated in the mother-involved portions of the distress-eliciting tasks based on infant affect and mother behavior. The process for determining maternal sensitivity scores was described in detail in (masked for blind review). First, infant affect in each task was continuously rated as described above. Mother behavior was coded in each task using 12 mutually exclusive categories (engagement, routine care, supportive, negative, intrusive, mismatch affect, withdraw, distracted, persistent ineffective, monitor, calming). Interrater reliability was assessed on 13% of cases at 1 year, kappa = .80. Next, the infant affect and maternal behavior code files were merged and the appropriateness and quality of maternal behavior at each moment was assigned a sensitivity rating based on whether concurrent infant was negative, neutral, or positive on an established 3 point scale, 1 = insensitive, 2 = moderately sensitive, and 3 = sensitive. The specific sensitivity rating assigned to each maternal behavior code given the infant’s current affect was consistent with the detailed rating scheme described in (masked for blind review). For example, calming behavior (soothing the child physically or vocally) while the child displays negative affect is assigned a score of 3 (sensitive) because the mother is responding to a clear distress cue from the infant and appropriately attempting to help the infant reduce this distress. Conversely, distracted behavior (uninvolved or minimally involved with child) while the child displays negative affect is assigned a score of 1 (insensitive). The mean level of sensitivity was calculated for each task point based on the sum of time spent at each sensitivity level divided by the total time.
SCL arousal.
Mothers skin conductance level (SCL) was continuously recorded in microsiemens at a sampling rate of 100Hz. SCL data were transferred from the Biolog to a laptop computer. A single imputation of the raw data was then performed to generate complete SCL data for cases missing due to equipment failure (4 cases in the fear task and 6 cases in the anger task among the final analytic sample) using SPSS Version 22 (IBM Corp, Armonk, NY). SCL variables were correlated across all tasks during the visit, so these variables and participant race were entered into the imputation model as predictors. SCL response scores were calculated for each distress task by subtracting average SCL during the baseline free play from average SCL during the task. Higher scores indicated greater SCL arousal from baseline to the task.
RSA change.
Mothers’ electrocardiogram was recorded at a sampling rate of 1 kHz. A data file containing the interbeat intervals (time between R-waves) during the tasks was transferred from the Biolog to a laptop computer for artifact editing (resulting from movement). Standard editing procedures were used by a staff member who was trained by and demonstrated reliability with staff from the Brain-Body Center, University of Illinois at Chicago. Task data requiring artifact editing for >10% of data was removed from the raw data file. Analyses to derive heart period variance were completed using CardioEdit software (Brain-Body Center, University of Illinois at Chicago). Measures of RSA, reflecting vagal tone, were calculated in ms2 using Porges’ (1985) method in 15-s epochs for each task. A band pass filter extracted heart period variance within the frequency band of spontaneous respiration (.12-.40 Hz) in adults. Epoch scores were averaged within task. Vagal withdrawal scores were calculated for each distress task by subtracting average RSA during the task from average RSA during the baseline free play. Higher scores indicated greater RSA withdrawal from baseline to the task, whereas lower scores indicated RSA augmentation. A single imputation of the raw data was then performed to generate complete RSA data for cases with >10% artifact editing (6 cases in the fear task and 4 cases in the anger task) or missing due to equipment failure (7 cases in the fear task and 7 cases in the anger task) using SPSS Version 22 (IBM Corp, Armonk, NY). RSA and heart period measures were correlated across all tasks during the visit, so these variables and participant race were entered into the imputation model as predictors. One mother’s score in the anger task was >3SD below the mean and was replaced with the next lowest score. Four mothers had no usable RSA data due to equipment failure for the entire visit.
Mothers’ ratings of infant emotion and cry attributions.
Mothers’ interview ratings about infant emotions were averaged into subscales reflecting infant fear (nervous/anxious, worried, afraid/scared; fear task α = .77, M = 2.01, SD = .85; anger task α = .66, M = 1.08, SD = .29) and anger (irritated/annoyed, aggravated, frustrated, angry/mad, disgusted; fear task α = .85, M = 1.19, SD = .47; anger task α = .83, M = 1.64, SD = .67). Although alphas above .70 are generally preferred, it is not uncommon to observe lower alphas in scales composed of a small number of items as is the case here (Tavakol & Dennick, 2011). Mothers’ ratings of attributions about her infant’s crying were averaged into subscales for each task reflecting negative (fear task α = .75, M = 1.22, SD = .35; anger task α = .72, M = 1.29, SD = .39) and minimizing (fear task α = .77, M = 1.23, SD = .39; anger task α = .83, M = 1.27, SD = .47) attributions.
Statistical Analyses
Primary study hypotheses regarding the interaction of infant distress and mother physiological responses were tested using two multiple regression models, one predicting maternal sensitivity during the fear task and one predicting sensitivity during the anger task. These models also controlled for mother race and education level. The predictors in each model included the two control variables, infant distress, mother SCL arousal, mother RSA change (higher values reflected RSA withdrawal), and all 2-way and 3-way interactions between infant distress, mother SCL arousal, and mother RSA change. The regression models were run in Mplus 8 (Muthén & Muthén, Los Angeles, CA) using full information maximum likelihood estimation to account for missing data, thus the sample size for these analyses was N = 208. Significant interaction terms were probed in two steps. First, significant 3-way interaction terms were probed by testing the significance of the SCL arousal × RSA change interaction at ±1 SD on infant distress. If the interaction was significant at either value of infant distress, this 2-way interaction was probed by examining the simple slope of SCL arousal at ±1 SD on RSA change at that value (±1 SD) of infant distress.
Secondary study hypotheses regarding context differences in infant and mother responses were examined with a series of multivariate repeated measures ANOVAs. One model tested differences in responses during the tasks by comparing mean infant distress, mother SCL arousal, mother RSA change and maternal sensitivity across contexts. The other model tested the difference in mothers’ perception of infant distress by comparing ratings of infant fear, infant anger, and negative attributions about crying across contexts. Given the nature of the sample, each model controlled for mother race. Additionally, maternal education level was found to relate to both SCL arousal (fear task r = .22, anger task r = .24, ps < .01) and sensitivity (fear task r = .26, anger task r = .29, ps < .01); thus this variable was also included as a covariate. Hypotheses regarding context differences in relations between infant distress and mother responses were tested by comparing partial correlations between infant distress and mothers’ SCL arousal, RSA change, and sensitivity, controlling for mother race and education level. These analyses were conducted in SPSS Statistics 24 (IBM, Armonk, NY). Due to difficulties in task administration and/or equipment failure resulting in missing data, the final sample size for these analyses ranged from N = 193–207. Differences in the magnitude of each correlation in the fear task relative to the anger task were tested as a comparison of non-overlapping correlations in a dependent sample using the corcor package in R (available at comparingcorrelations.org).
Results
Descriptive statistics and correlations among the major variables of interest are presented in Table 1. Infant distress, SCL arousal, RSA change, and sensitivity were each significantly correlated across the anger and fear task. Additionally, mothers’ SCL arousal was significantly correlated with sensitivity, whereas RSA change was not. Infant distress during the anger task was significantly negatively correlated with mothers’ RSA change and sensitivity, but infant distress during the fear task was not significantly related to maternal responses during that task.
Table 1.
Correlations among Major Study Variables
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
|---|---|---|---|---|---|---|---|---|---|
| 1. Infant distress, Fear task | --- | .10 | −.06 | −.12+ | .04 | −.08 | .04 | −.17* | |
| 2. Infant distress, Anger task | --- | .03 | .07 | −.15* | −.11 | .05 | −.37* | ||
| 3. Maternal SCL arousal, Fear task | --- | .82** | .03 | .06 | .29** | .17* | |||
| 4. Maternal SCL arousal, Anger task | --- | .01 | .10 | .17* | .17* | ||||
| 5. Maternal RSA change1, Fear task | --- | .53** | .11 | .11 | |||||
| 6. Maternal RSA change, Anger task | --- | .07 | .08 | ||||||
| 7. Maternal sensitivity, Fear task | --- | .14* | |||||||
| 8. Maternal sensitivity, Anger task | --- | ||||||||
| M | 4.21 | 4.18 | 2.74 | 1.96 | −.13 | −.10 | 2.50 | 2.24 | |
| SD | .45 | .37 | 2.14 | 1.74 | .83 | .66 | .36 | .30 |
p < .10
p < .05
p < .01
Higher values reflected RSA withdrawal, lower values reflected RSA augmentation.
Primary analyses: Regression models predicting maternal sensitivity.
Fear task.
Results from the regression model predicting maternal sensitivity in the fear task are presented in Table 2. Higher levels of maternal education, greater infant distress, and greater maternal SCL arousal related to greater sensitivity. These effects were qualified by a significant interaction of maternal SCL arousal and RSA change, infant distress and maternal RSA change, as well as a significant three-way interaction of infant distress, maternal SCL arousal and RSA change. The three-way interaction was probed by testing the SCL arousal × RSA change interaction at ±1 SD on infant distress. This test indicated that the interaction was significant at +1 SD (B = .11, SE = .04, p = .005) but not −1 SD (B = −.03, SE = .03, p = .26) on infant distress. Thus, the slope of SCL arousal was tested at ±1 SD on RSA change for mothers of infants who displayed higher distress. Given the distribution of RSA change scores, +1 SD reflected RSA withdrawal and −1 SD reflected RSA augmentation. These tests indicated that SCL arousal related to greater sensitivity in the fear task for mothers who displayed RSA withdrawal (B = .14, SE = .03, p < .001), but not for mothers who displayed RSA augmentation (B = −.04, SE = .04, p = .33), as illustrated in Figure 1.
Table 2.
Multiple Regression Models Predicting Maternal Sensitivity in the Fear Task and Anger Task
| Variables | B | SE(B) | β | R2 |
|---|---|---|---|---|
| Fear Task | .20** | |||
| Mother race | −.05 | .05 | −.07 | |
| Maternal education | .03 | .01 | .15* | |
| Infant distress | .13 | .06 | .16* | |
| Mother SCL arousal | .04 | .01 | .24** | |
| Mother RSA change1 | .02 | .03 | .04 | |
| SCL × RSA | .04 | .02 | .19** | |
| Distress × SCL | .02 | .03 | .05 | |
| Distress × RSA | −.22 | .10 | −.17* | |
| Distress × SCL × RSA | .15 | .06 | .19* | |
| Anger Task | .31** | |||
| Mother race | −.02 | .04 | −.03 | |
| Maternal education | .04 | .01 | .24** | |
| Infant distress | −.29 | .06 | −.35** | |
| Mother SCL arousal | .02 | .01 | .11 | |
| Mother RSA change | .02 | .03 | .05 | |
| SCL × RSA | −.01 | .02 | −.06 | |
| Distress × SCL | .03 | .03 | .07 | |
| Distress × RSA | .09 | .10 | .07 | |
| Distress × SCL × RSA | −.14 | .05 | −.20** |
p < .10
p < .05
p < .01
Higher values reflected RSA withdrawal, lower values reflected RSA augmentation; Mother race was coded 0 = non-White, 1 = White
Figure 1.
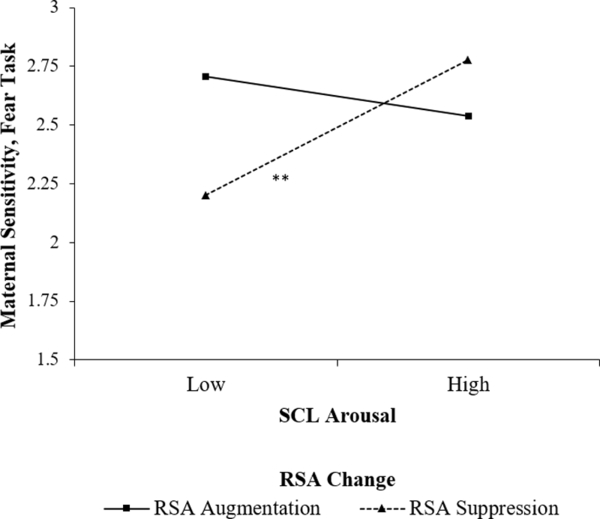
Maternal SCL arousal × RSA change interaction predicting maternal sensitivity during the fear task with infants who displayed higher distress.
Anger task.
Results from the regression model predicting maternal sensitivity in the anger task are presented in Table 2. Higher levels of maternal education related to greater sensitivity. Infant distress related to lower levels of maternal sensitivity, and this effect was qualified by a significant three-way interaction of infant distress, maternal SCL arousal and RSA change. The three-way interaction was probed by testing the SCL arousal × RSA change interaction at ±1 SD on infant distress. This test indicated that the interaction was significant at +1 SD (B = −.07, SE = .02, p = .004) but not −1 SD (B = .04, SE = .02, p = .13) on infant distress. Thus, the slope of SCL arousal was tested at ±1 SD on RSA change for mothers of infants who displayed higher distress. Once again, +1 SD reflected RSA withdrawal and −1 SD reflected RSA augmentation. These tests indicated that SCL arousal related to greater sensitivity in the anger task for mothers who displayed RSA augmentation (B = .07, SE = .02, p < .001), but not for mothers who displayed RSA withdrawal (B = −.01, SE = .02, p = .65), as illustrated in Figure 2.
Figure 2.
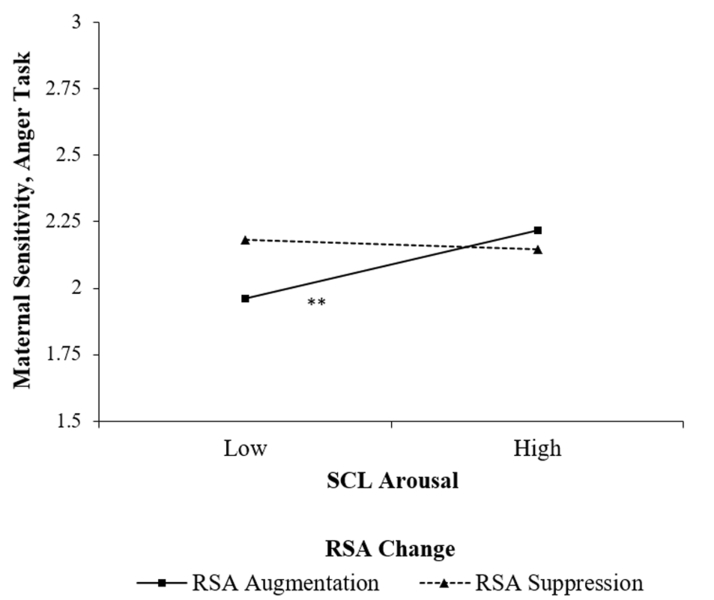
Maternal SCL arousal × RSA change interaction predicting maternal sensitivity during the anger task with infants who displayed higher distress.
Secondary analyses: Context differences.
Results from the multivariate repeated-measures ANOVA testing differences in responses during the tasks indicated that, controlling for maternal race and education, there was no difference across the fear and anger task in infant distress (estimated difference .02, SE = .04, p = .53) and maternal RSA change (estimated difference −.05, SE = .05, p = .38). However, mothers displayed higher levels of SCL arousal (estimated difference .75, SE = .08, p < .001) and sensitivity (estimated difference .25, SE = .03, p < .001) in the fear task relative to the anger task.
Results from the multivariate repeated-measures ANOVA testing differences in mothers’ emotion and attribution ratings indicated that, controlling for maternal race and education, mothers reported higher infant fear in the fear task (estimated difference .93, SE = .06, p < .001) higher infant anger in the anger task (estimated difference .45, SE = .05, p < .001), more negative attributions about infant crying in the anger task (estimated difference .08, SE = .02, p < .001), and more minimizing attributions about infant crying in the anger task (estimated difference .04, SE = .02, p = .05) consistent with hypotheses.
Results from the partial correlations between infant distress and mother responses and attributions during the distress tasks indicated that infant distress was not related to mothers’ SCL arousal (fear r = .03, p = .64; anger r = .07, p = .32; difference z = −.39, p = .69) or RSA change (fear r = .04, p = .59; anger r = −.10, p = .16; difference z = 1.44, p = .15). However, whereas greater infant distress was not significantly related to sensitivity during the fear task (r = .05, p = .50), it was related to lower sensitivity during the anger task (r = −.39, p < .001), and these correlations significantly differed across the tasks (difference z = 4.67, p < .001). Similarly, greater infant distress was not significantly related to mothers’ attributions about the fear task (negative r = .06, p = .42; minimizing r = .03, p = .70), but it was related to more negative and minimizing attributions during the anger task (negative r = .25, p = <.001; minimizing r = .35, p = <.001), and these correlations significantly differed across the tasks (negative difference z = −2.10, p = .04; minimizing difference z = −3.61, p < .001).
Discussion
Previous research suggests that mothers’ autonomic nervous system responses to infant distress may contribute to patterns of maternal sensitivity, and further, that SCL arousal and RSA change may interact to predict sensitivity. However, conceptual arguments about parenting and physiological functioning suggest that different types of infant distress might evoke distinct patterns of maternal responses. The current study utilized observations of mothers and infants in situations designed to elicit infant fear and infant anger, and examined how mothers’ SCL arousal and RSA change predicted sensitivity based on infant distress within each context.
Primary study analyses addressed the possibility that infant distress and mother physiological responses may interact to predict maternal sensitivity in a fear task relative to an anger task. Consistent with hypotheses, mothers’ physiological responses interacted to predict sensitivity primarily when infants displayed relatively greater distress. This suggests that beyond contextual information, infant distress cues serve as a potent stimulus underlying the patterns by which mothers’ physiological responses relate to sensitive behavior. Further, and again as expected, mothers’ RSA change moderated the effects of their SCL arousal in a distinct pattern in each context with infants who displayed greater distress. Importantly, the interaction pattern for both tasks emerged such that the significant simple slope reflected a lower level of sensitivity for mothers who displayed lower SCL arousal. This pattern may reflect the proposition that low levels of sympathetic activity may lead to disengagement from mother-child interactions (Reijman et al., 2014; Sturge-Apple et al., 2016); however, our results suggest this may only be the case for mothers who display specific parasympathetic responses.
In the fear task, when infants displayed greater distress, SCL arousal related to sensitivity for mothers who displayed RSA withdrawal, but not for those who displayed RSA augmentation. Specifically, a lower level of arousal related to lower sensitivity in mothers who displayed RSA withdrawal. This pattern is consistent with proposals made in previous research with this sample that physiological arousal in response to infant distress should relate to greater sensitivity when mothers also display physiological regulation (Leerkes et al., 2015; 2016). However, with older infants, this pattern may be most applicable to situations involving fear or threat stimuli, to the extent that it supports the enactment of maternal flight-or-flight responses that ensure infant protection (Porges, 2007). Mothers whose parasympathetic response reflects preparedness to manage threat (RSA withdrawal) may benefit from sympathetic arousal in order to prioritize infant needs and enact responses that encourage soothing or a sense of security in the infant rather than disengagement (Leerkes et al., 2015; 2016; Sturge-Apple et al., 2011). Conversely, mothers who tend to increase parasympathetic influence (RSA augmentation) may do so because they perceive less anxiety about their ability to appropriately manage infant security. RSA augmentation may thus serve to facilitate sensitive engagement with the infant regardless of SCL response (Miller et al., 2015), perhaps by limiting intrusive protection strategies.
A different descriptive pattern emerged for the anger task. Here, when infants displayed greater distress, SCL arousal related to greater sensitivity for mothers who displayed RSA augmentation, but not for those who displayed RSA withdrawal. Specifically, lower arousal related to lower sensitivity in mothers who displayed RSA augmentation. This task was designed to pose no risk to infant safety and instead to elicit cues of infant goal pursuit and frustration, and mothers displayed lower levels of sensitivity and SCL arousal in this task relative to the fear task. RSA augmentation may reflect a tendency to perceive infant frustration as an experience that may be managed through adaptive social or problem-solving interactions (Porges, 2007), but the experience of SCL arousal in response to infant distress may promote a motivation to adaptively assist the infant with managing the task at hand, whereas low arousal may lead to behaviors reflecting a lack of concern or an aversion to the infants’ frustration (Miller et al., 2015). Mothers’ tendency to display RSA withdrawal suggested a perception that the task involved the presence of an environmental challenge that must be coped with in some manner. However, given the absence of threat cues in this task, this pattern may have motivated less of a fight-or-flight response as much as an adaptive motivation to address the source of frustration with their infant regardless of SCL arousal level. Relative to findings at earlier time points in this sample (e.g., Leerkes et al., 2015; 2016) distinct implications of mothers’ physiological responses to infant anger may emerge later in infants’ emotional development (Izard, 1978).
It is also important to note that mothers’ physiological responses were each strongly correlated across contexts, whereas infant distress and mother sensitivity were not. It is possible that mothers who displayed low SCL arousal to infant distress may have appeared relatively sensitive in one context but not the other depending on their RSA response and their infants’ distress. This suggests that mothers’ relatively stable patterns of physiological responses to infant distress may not confer uniform predispositions toward (mal)adaptive parenting behavior (e.g., Joosen et al., 2013a; 2013b), but instead predict behavior specific to certain contexts. These findings affirm the importance of considering contextual differences in parent-child interactions, both in determining how to support positive parenting behavior, and when understanding parents’ socialization of child outcomes in different domains (see Grusec & Davidov, 2010).
Secondary study analyses examined context differences in infant and maternal responses in order to confirm our expectations surrounding mothers’ differing perceptions of each context. Consistent with hypotheses, mothers did in fact report greater infant fear in the fear task and anger in the anger task, suggesting that mothers utilized contextual information to make inferences about the nature of their infants’ distress (Dix, 1992; Oster et al., 2012). Mothers also reported more negative and minimizing attributions about the anger task relative to the fear task, and displayed higher levels of both SCL arousal and sensitivity in the fear task relative to the anger task. Further, infant distress was not associated with sensitivity in the fear task; however, mothers tended to display lower levels of sensitivity and report more negative and minimizing attributions during the anger task with infants who displayed higher distress.
Taken together, these secondary findings may indicate that situations involving infant fear elicit greater SCL arousal, less-negative attributions, and a greater readiness to respond sensitively in mothers than situations involving infant anger. Importantly, these differences emerged despite the fact that infants displayed similar mean levels of distress. Thus, the current results provide some evidence for our proposition that mothers may be more reactive to or more accepting of infant distress in situations designed to elicit fear compared to anger. Children’s anger and fear reactivity both play important but potentially differing roles in children’s development of emotion regulation skills and their risk for later behavior problems (Calkins, 1994). Systematic differences in maternal perceptions, physiological functioning, and sensitivity across tasks that elicit these different emotions may be important to consider in the study of these developmental pathways. Nonetheless, these results also indicated that infant distress did not relate to mothers’ SCL arousal in either task. Although mothers’ arousal levels differed across context, it is possible that distress related to patterns of SCL arousal × RSA change rather than each measure alone. This cannot be determined from the present analyses, as the regression of a statistical interaction on another variable cannot be interpreted in a meaningful way. This possibility could, however, be addressed through person-oriented approaches.
The results of this study may also point to the increasing importance of situational cues for maternal physiological responses and sensitivity through infancy. Context-specific patterns were not addressed in earlier studies using this dataset, but additional analyses (available upon request) suggest the SCL arousal × RSA change interaction did not predict sensitivity differently by context and/or infant distress level when infants were younger. Thus, it seems plausible that, due to greater experience with parenting and/or greater differentiation of infant emotion (Izard, 1978), mothers’ perceptions of infant distress could become more distinct over time and more reliably tied to the context in which distress is displayed. Mothers may also begin to display distinctions in their tolerance for different forms of distress, as evidenced by different perceptions of anger compared to fear. These developmental distinctions may help to explain the different patterns of association between physiological responses and sensitivity at this age.
The strengths of this study include the use of observational and mother-report measures at both the physiological and behavioral level. These data were collected in a racially and socioeconomically diverse sample, allowing for greater generalizability of the results. Hypothesized context differences in mothers’ perceptions of infant emotions were addressed in secondary analyses, supporting our interpretation that the different pattern of relations between mothers’ physiological responses and sensitivity across contexts is in part due to differences in mothers’ perceptions and responses within each context. Additionally, this study addressed individual differences in both mothers and infants as predictors of maternal behavior, providing relatively more precise information about patterns of functioning in mother-child dyads than would measures examined only in mothers or only through main effects.
We also acknowledge several study limitations. This study examined concurrent measures of infant and mother responses in order to understand relations between in-the-moment responses; however, it is unclear from these cross-sectional findings whether context-based patterns of maternal physiology and infant distress have longitudinal implications for mothers’ later sensitivity, or if these patterns emerge only within the same point in development. We also examined infant and mother responses as average levels across task time rather than utilizing contingency or time-series methods. Both of these characteristics limit the ability to infer causal relations between these variables. Additionally, the infant distress tasks were conducted in the same order for all infants; thus, fear task measures may have been affected by carryover of infant distress and/or mother fatigue from the anger task. However, all distressed infants were soothed between tasks as necessary and infant distress level was not significantly correlated across tasks, providing some evidence against the presence of systematic carryover effects. Lastly, our primary analyses concerned patterns of effects within each distress context, but did not address a formal test of differences in effects across contexts (i.e., a 4-way interaction of context, infant affect, SCL arousal, and RSA change). We acknowledge our speculation about such context differences remains descriptive. Future studies with larger samples may be better-powered to address these patterns in a repeated-measures framework.
In terms of future research, further exploration of differences in mothers’ responses to infant fear and anger are warranted. One major priority is to address this complex pattern of findings in replication studies before any robust inferences may be made. It is possible that lower-order interaction effects will receive more consistent support. Moving forward, it is also possible these distinct patterns have implications for child outcomes, if mothers are less eager or effective in managing developmentally-appropriate infant anger. Similarly, although not a focus of this study, mothers’ perceptions of infant distress may relate to their patterns of physiological responses in different situations, which in turn relate to sensitivity. This underscores the idea that mothers’ cognitive and physiological reactions to infant distress could be significant targets for parenting interventions (Leerkes et al., 2016), particularly for mothers of infants and children who display higher emotional reactivity. Lastly, broader methodological approaches could be addressed, such as examining a larger number or range of distress tasks, and examining if the current pattern of results emerges with older children and/or in longitudinal study designs.
In conclusion, these results provide meaningful insight into the role of situational context in mothers’ physiological and behavioral responses to infant distress. Mothers appeared to readily utilize contextual information in their perceptions of infant emotional responses, and these perceptions may underlie systematic variation in their responses to infant distress in different situations. Further, mothers’ patterns of physiological responses related to sensitivity differently based on the infants’ level of distress, with apparent differences by infant distress type. Collectively, mothers’ physiological responses to infant distress may not relate to sensitive responses uniformly across contexts, and researchers should acknowledge this variation as a potential contributor to larger patterns of infant emotional development.
Acknowledgments
This work was supported by the Eunice Kennedy Shriver National Institute for Child Health and Human Development (NICHD) through R01HD058578 awarded to the second author, and a postdoctoral fellowship (T32-HD07376) through the Center for Developmental Science, University of North Carolina at Chapel Hill, to the first author. Results from this study were previously presented at the 2018 Biennial Meeting of the International Congress on Infant Studies. The contents of this article are the sole responsibility of the authors and do not necessarily reflect the views of NICHD. We are grateful to the participants for their time and Dr. Regan Burney and project staff for their dedication.
References
- Ablow JC, Marks AK, Feldman SS, & Huffman LC (2013). Associations between first-time expectant women’s representations of attachment and their physiological reactivity to infant cry. Child Development, 84, 1373–1391. doi:0.1111/cdev.12135 [DOI] [PubMed] [Google Scholar]
- Berntson GG, Cacioppo JT, & Quigley KS (1993). Cardiac psychophysiology and autonomic space in humans: Empirical perspectives and conceptual implications. Psychological Bulletin, 114, 296–322. doi: 10.1037/0033-2909.114.2.296 [DOI] [PubMed] [Google Scholar]
- Braungart-Rieker JM, Hill-Soderlund AL, & Karrass J (2010). Fear and anger reactivity trajectories from 4 to 16 months: The roles of temperament, regulation, and maternal sensitivity. Developmental Psychology, 46, 791–804. doi: 10.1037/a0019673 [DOI] [PubMed] [Google Scholar]
- Buss KA, & Goldsmith HH (1998). Fear and anger regulation in infancy: Effects on the temporal dynamics of affective expression. Child Development, 69, 359–374. doi: 10.2307/1132171 [DOI] [PubMed] [Google Scholar]
- Calkins SD (1994). Origins and outcomes of individual differences in emotion regulation. In Fox NA (Ed.), The development of emotion regulation: Biological and behavioral considerations. Monographs of the Society for Research in Child Development, 59(2–3), 53–72. [PubMed] [Google Scholar]
- Casey RJ, & Fuller LL (1994). Maternal regulation of children’s emotions. Journal of Nonverbal Behavior, 18, 57–89. doi: 10.1007/BF02169079 [DOI] [Google Scholar]
- Cole P, Martin S, & Dennis T (2004). Emotion regulation as a scientific construct: Methodological challenges and directions for child development research. Child Development, 75, 317–333. doi: 10.1111/j.1467-8624.2004.00673.x [DOI] [PubMed] [Google Scholar]
- Denham SA (1993). Maternal emotional responsiveness and toddlers’ social-emotional competence. Journal of Child Psychology and Psychiatry, 34, 715–728. doi: 10.1111/j.1469-7610.1993.tb01066.x [DOI] [PubMed] [Google Scholar]
- Dix T (1992). Parenting on behalf of the child: Empathic goals in the regulation of responsive parenting. In Sigel IE, McGillicuddy-DeLisi AV, & Goodnow JJ (Eds.), Parental belief systems: The psychological consequences for children (pp. 319–346). Mahwah, NJ: Erlbaum. [Google Scholar]
- Goldsmith HH, Reilly HH, Lemery KS, Longley S, & Prescott A (1994). Manual for the Preschool Laboratory Temperament Assessment Battery (Lab-TAB). Unpublished manuscript Madison: University of Wisconsin. [Google Scholar]
- Gray JA (1975). Elements of a two-process theory of learning Oxford, UK: Academic Press. [Google Scholar]
- Izard CE (1978). On the ontogenesis of emotions and emotion-cognition relationships in infancy. In Lewis M & Rosenblum LA (Eds.) The development of affect (pp. 389–413). New York, NY: Plenum Press. [Google Scholar]
- Joosen KJ, Mesman J, Bakermans-Kranenburg MJ, Pieper S, Zeskind PS, & van IJzendoorn MH (2013). Physiological reactivity to infant crying and observed maternal sensitivity. Infancy, 18, 414–431. doi: 10.1111/j.1532-7078.2012.00122.x [DOI] [Google Scholar]
- Joosen KJ, Mesman J, Bakermans-Kranenburg MJ, & van IJzendoorn MH (2013). Maternal overreactive sympathetic nervous system responses to repeated infant crying predicts risk for impulsive harsh discipline of infants. Child Maltreatment, 18, 252–263. doi: 10.1177/1077559513494762 [DOI] [PubMed] [Google Scholar]
- Leerkes EM, Blankson AN, & O’Brien M (2009). Differential effects of maternal sensitivity to infant distress and nondistress on social-emotional functioning. Child Development, 80, 762–775. doi: 10.1111/j.1467-8624.2009.01296.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leerkes EM, & Siepak KJ (2006). Attachment linked predictors of women’s emotional and cognitive responses to infant distress. Attachment & Human Development, 8, 11–32. doi: 10.1080/14616730600594450 [DOI] [PubMed] [Google Scholar]
- Leerkes EM, Su J, Calkins SD, Supple AJ, and O’Brien M (2016). Pathways by which mothers’ physiological arousal and regulation while caregiving predict sensitivity to infant distress. Journal of Family Psychology, 30, 769–779. doi: 10.1037/fam0000185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leerkes EM, Supple AJ, O’Brien M, Calkins SD, Haltigan JD, Wong MS, and Fortuna K (2015). Antecedents of maternal sensitivity during distressing tasks: Integrating attachment, social information processing, and psychobiological perspectives. Child Development, 86, 94–111. doi: 10.1111/cdev.12288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McElwain NL, & Booth-LaForce C (2006). Maternal sensitivity to infant distress and nondistress as predictors of infant-mother attachment security. Journal of Family Psychology, 20, 247–255. doi: 10.1037/0893-3200.20.2.247 [DOI] [PubMed] [Google Scholar]
- Mesman J, Oster H, & Camras L (2012). Parental sensitivity to infant distress: what do discrete negative emotions have to do with it? Attachment & Human Development, 14, 337–348. doi: 10.1080/14616734.2012.691649 [DOI] [PubMed] [Google Scholar]
- Mills-Koonce WR, Gariépy J, Propper C, Sutton K, Calkins S, Moore G, & Cox M (2007). Infant and parent factors associated with early maternal sensitivity: A caregiver-attachment systems approach. Infant Behavior & Development, 30, 114–126. doi: 10.1016/j.infbeh.2006.11.010 [DOI] [PubMed] [Google Scholar]
- Mills-Koonce WR, Propper C, Gariepy J, Barnett M, Moore GA, Calkins S, & Cox MJ (2009). Psychophysiological correlates of parenting behavior in mothers of young children. Developmental Psychobiology, 51, 650–661. doi: 10.1002/dev.20400 [DOI] [PubMed] [Google Scholar]
- Moore GA, Hill-Soderlund AL, Propper CB, Calkins SD, Mills-Koonce WR, & Cox MJ (2009). Mother-infant vagal regulation in the face-to-face still-face paradigm is moderated by maternal sensitivity. Child Development, 80, 209–223. doi: 10.1111/j.1467-8624.2008.01255.x [DOI] [PubMed] [Google Scholar]
- Porges SW (2007). The polyvagal perspective. Biological Psychology, 74, 116–143. doi: 10.1016/j.biopsycho.2006.06.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reijman S, Alink LRA, Compier-de Block LHCG, Werner CD, Maras A, Rijnberk C,…Bakermans-Kranenburg MJ. (2014). Autonomic reactivity to infant crying in maltreating mothers. Child Maltreatment, 19, 101–112. doi: 10.1177/1077559514538115 [DOI] [PubMed] [Google Scholar]
- Rothbart MK, & Bates JE (2006). Temperament. In Damon W & Lerner R (Eds.), Handbook of child psychology: Vol. 3, social, emotional and personality development (6th ed., pp. 99–166). New York, NY: Wiley. [Google Scholar]
- Tavakol M & Dennick R (2011). Making sense of Cronbach’s alpha. International Journal of Medical Education, 2, 53–55. doi: 10.5116/ijme.4dfb.8dfd [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturge-Apple ML, Skibo MA, Rogosch FA, Ignjatovic Z, & Heinzelman W (2011). The impact of allostatic load on maternal sympathovagal functioning in stressful child contexts: Implications for problematic parenting. Development and Psychopathology, 23, 831–844. doi: 10.1017/S0954579411000332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfe DA, Fairbank JA, Kelly JA, & Bradlyn AS (1983). Child abusive parents’ physiological responses to stressful and non-stressful behavior in children. Behavioral Assessment, 5, 363–371. [Google Scholar]