

Summary
Epitope-targeted HIV vaccine design seeks to focus antibody responses to broadly neutralizing antibody (bnAb) sites by sequential immunization. A chimpanzee simian immunodeficiency virus (SIV) envelope (Env) shares a single bnAb site, the variable loop 2 (V2)-apex, with HIV, suggesting its possible utility in an HIV immunization strategy. Here, we generate a chimpanzee SIV Env trimer, MT145K, which displays selective binding to HIV V2-apex bnAbs and precursor versions, but no binding to other HIV specificities. We determine the structure of the MT145K trimer by cryo-EM and show that its architecture is remarkably similar to HIV Env. Immunization of an HIV V2-apex bnAb precursor Ab-expressing knockin mouse with the chimpanzee MT145K trimer induces HIV V2-specific neutralizing responses. Subsequent boosting with an HIV trimer cocktail induces responses that exhibit some virus cross-neutralization. Overall, the chimpanzee MT145K trimer behaves as expected from design both in vitro and in vivo and is an attractive potential component of a sequential immunization regimen to induce V2-apex bnAbs.
Keywords: human immunodeficiency virus, simian immunodeficiency virus, broadly neutralizing antibodies, HIV envelope trimer, chimpanzee SIV envelope trimer, glycan shield, V2-apex bnAb site, immunization strategies, HIV vaccine
Graphical Abstract
Highlights
-
•
A designed chimpanzee SIV Env trimer binds HIV V2-apex bnAbs specifically
-
•
The trimer (MT145K) is engineered to bind inferred unmutated versions of HIV V2-apex bnAbs
-
•
The cryo-EM structure of the SIV MT145K trimer closely resembles that of HIV trimers
-
•
The MT145K SIV trimer induces HIV-specific nAb responses in a favorable animal model
Design of immunogens and strategies that can induce protective broadly neutralizing antibodies (bnAbs) is a priority for HIV vaccine development. Andrabi et al. design a chimpanzee simian immunodeficiency virus (SIV) envelope trimer immunogen that binds specifically to HIV V2-apex bnAbs and their unmutated versions. The SIV trimer immunogen induces HIV-specific neutralizing antibodies (nAbs) in a favorable animal model.
Introduction
The ability to induce HIV envelope (Env) specific broadly neutralizing antibodies (bnAbs) will likely be a key feature of a prophylactic vaccine immunogen. Potent Env-specific bnAbs are produced in a small subset of HIV infected donors, yet attempts to elicit such responses through immunization have failed to date other than in certain animal models (Andrabi et al., 2018, Escolano et al., 2017, Escolano et al., 2016, Haynes and Mascola, 2017, McCoy et al., 2012, Sok et al., 2017, Xu et al., 2018). Previous studies have revealed that HIV bnAb germline-reverted precursors possess unique features that greatly reduce their overall frequencies in the human B cell immune repertoire and, hence, their ability to be targeted by vaccines (Briney et al., 2012, Haynes et al., 2012, Kepler et al., 2014, Klein et al., 2013, Verkoczy et al., 2010, Xiao et al., 2009). Therefore, recent immunogen design approaches that seek to induce bnAb responses by vaccination are taking these rare precursor features into consideration to efficiently activate bnAb precursors and shepherd them along favorable bnAb developmental pathways (Andrabi et al., 2015, Andrabi et al., 2018, Escolano et al., 2016, Gorman et al., 2016, Jardine et al., 2013, McGuire et al., 2013, Saunders et al., 2017, Steichen et al., 2016). These design approaches have shown promise for two of the HIV Env bnAb sites, namely the CD4 binding site (CD4bs) and the V3-N332 glycan site in animal models expressing the appropriate germline precursors (Andrabi et al., 2018, Briney et al., 2016, Dosenovic et al., 2015, Escolano et al., 2016, Jardine et al., 2015, McGuire et al., 2016, Sok et al., 2016a, Steichen et al., 2016, Tian et al., 2016, Williams et al., 2017). Thus, immunogen designs and strategies that can select rare bnAb precursors and reduce off-target B cell responses are valuable for nAb immunofocusing efforts.
One of the Env sites that has shown promise for vaccine targeting is the variable loop 2 (V2)-apex bnAb epitope (Andrabi et al., 2015, Gorman et al., 2016, Moore et al., 2017, Voss et al., 2017). This bnAb epitope sits at the 3-fold axis of the trimer and is primarily formed by a patch rich in positively charged lysine residues and protected by two glycans at HXB2 HIV reference positions N160 and N156/N173 that are part of the Env glycan shield (Andrabi et al., 2017, Bhiman et al., 2015, Bonsignori et al., 2011, Doria-Rose et al., 2014, Gorman et al., 2016, Julien et al., 2013b, Lee et al., 2017, McLellan et al., 2011, Pancera et al., 2013, Walker et al., 2009, Walker et al., 2011). The bnAb precursors targeting this site possess a long anionic heavy-chain complementarity-determining region 3 (CDRH3) that penetrates the glycan shield to reach the protein epitope surface underneath (Bonsignori et al., 2011, Doria-Rose et al., 2014, Landais et al., 2017, Lee et al., 2017, McLellan et al., 2011, Walker et al., 2009, Walker et al., 2011). BnAb prototypes within this class interact with the V2-apex bnAb protein-glycan core epitope through common germline-encoded motifs and are, thus, targetable by a small set of trimers that interact with germline-reverted V2-apex bnAbs, as previously reported by us and others (Andrabi et al., 2015, Gorman et al., 2016). Hence, the germline-priming immunogens to this site could be based directly on native-like trimer configurations (Sanders et al., 2013, Sanders et al., 2015). Other features that favor this site for vaccine targeting include the following: (1) V2-apex bnAbs are elicited frequently in humans that make bnAbs, (2) they emerge early in infection, and (3) they possess relatively low levels of somatic mutation compared to most other HIV Env bnAbs (Bonsignori et al., 2011, Doria-Rose et al., 2014, Georgiev et al., 2013, Kepler et al., 2014, Landais et al., 2016, Landais et al., 2017, Moore et al., 2011, Walker et al., 2009, Wibmer et al., 2013).
Of note, among the major HIV Env bnAb specificities, which include V2-apex, V3-N332, CD4bs, and the gp120-41 interface, the V2-apex is the only bnAb site that consistently exhibits cross-group neutralizing activity with virus Envs derived from HIV groups M, N, O, and P (Braibant et al., 2013, Morgand et al., 2016). Furthermore, V2-apex bnAbs display cross-neutralizing activity with the simian immunodeficiency virus (SIV) isolates that infect chimpanzees (SIVcpzPtt [Pan troglodytes troglodytes], SIVcpzPts [Pan troglodytes schweinfurthii]) and gorillas (SIVgor) (Barbian et al., 2015). The retention of the V2-apex bnAb epitope at the time of species crossover from chimpanzees to humans highlights the biological significance of this region and here we sought to design a trimer based on the SIVcpzPtt Env sequence that could potentially help guide an immunofocused response to the HIV V2-apex bnAb site. We hypothesized that an SIVcpzPtt-based trimer will not only assist in specifically enriching V2-apex-specific B cells but also, owing to V2-apex species cross-conservation, could assist in guiding a V2-focused nAb response when coupled with HIV trimers in a sequential prime-boost immunization strategy. In such an immunization scheme, the overall Env backbone sequence diversity in combination with conservation of the V2-apex bnAb epitope in sequentially administered immunogens is likely to reduce germinal center competition for V2-apex-specific B cells (Havenar-Daughton et al., 2017, Tas et al., 2016, Wang et al., 2015). Such a scheme could not only favor a B cell recall response to the V2-apex bnAb epitope but also reduce off-target Env-specific responses.
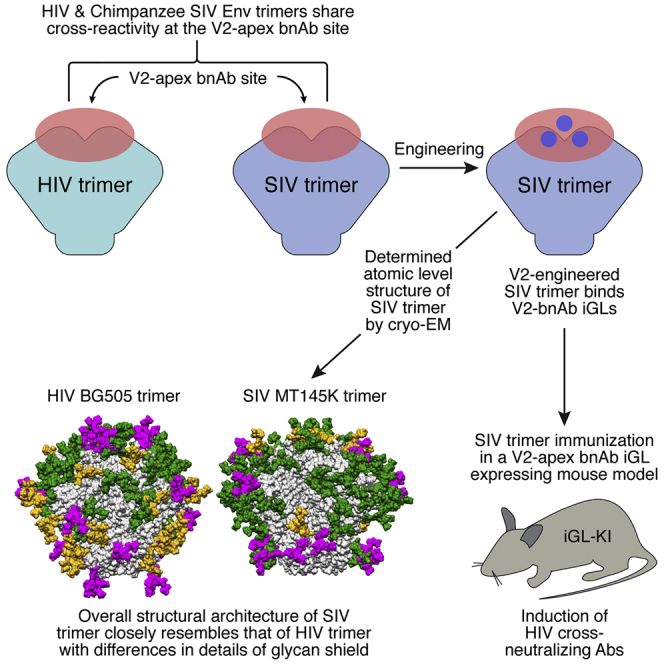
We designed an SIVcpzPtt-based disulfide (SOS), I559P (IP) prefusion stabilized Env ectodomain truncated as residue 664 (SOSIP.664) trimer, MT145K, that displays native trimer-like properties, and selectively binds V2-apex bnAbs as well as their germline reverted precursor versions. We determined a structure of the MT145K trimer by cryo-EM at a global resolution of 4.1 Å and the overall architecture was remarkably similar to HIV Env trimers (Julien et al., 2013a, Lyumkis et al., 2013, Ozorowski et al., 2017, Pancera et al., 2014). In addition, the glycan shield composition of MT145K closely resembled that of HIV Env glycans but was sufficiently different in positioning of the glycans to exclude binding of all HIV bnAbs except for those directed to the V2-apex. MT145K trimer immunization in a V2-apex unmutated common ancestor (UCA)-expressing knockin mouse model revealed induction of a predominantly V2-apex neutralizing Ab response that was reproducible and cross-neutralized a related set of HIV isolates. Overall, the chimpanzee MT145K immunogen shows potential as an immunogen in HIV vaccination strategies.
Results
Selection and Design of a Chimpanzee Env-Derived Trimer
Immunogen templates based on native-like Env trimers offer potential for HIV vaccine development, as they display bnAb epitopes and largely occlude non-native epitopes. However, it remains challenging to induce an epitope-focused bnAb response with Env trimer immunogens, as the bnAb epitopes are relatively immunoquiescent and even very limited exposure of non-desirable epitopes can negatively impact responses to bnAb epitopes (Havenar-Daughton et al., 2017, Wang et al., 2015). Therefore, trimer designs and/or strategies that can mask non-relevant immunodominant epitopes or reduce induction of off-target Ab responses could help guide immunofocused neutralizing responses. In addition, the typical lack of interaction of Env forms with germline-reverted bnAb precursors means difficulties in activating the appropriate B cell lineages. Accordingly, we undertook design of a trimer immunogen that could help guide an epitope-focused Ab response to the V2-apex site of HIV Env. Based on previous studies, we hypothesized that a chimpanzee SIVcpzPtt/Pts or gorilla SIVgor Env sequence-based trimer that shares the V2-apex bnAb epitope with HIV could enrich B cell precursors and boost responses specific to this site (Barbian et al., 2015). Since SIVcpzPtt, among various SIV-species Env sequences, are phylogenetically closest to the HIV Env, we surmised that the SOSIP.664 trimer-stabilizing modifications, which have been used on several HIV Env backgrounds, may function in the stabilization of soluble SIV Env (Gao et al., 1999, Sanders et al., 2013, Sharp and Hahn, 2011).
We incorporated the SOSIP.664 trimer design modifications into four SIVcpzPtt Env sequences: GAB1, MB897, EK505, and MT145 (Figure S1). These isolates have been previously shown to be sensitive to the V2-apex bnAbs, PG9, PG16, and PGT145 (Barbian et al., 2015). Further characterization showed that one of these SIVcpzPtt Env sequences, MT145 SOSIP.664, could be expressed as a soluble Env trimer protein (Figure S1). A PGT145 Ab affinity-purified MT145 trimer was efficiently cleaved into gp120 and gp41 subunits, and revealed well-ordered native-like trimer configurations that were highly thermostable, which are all properties displayed by natively folded HIV soluble trimers (Figure S1) (Pugach et al., 2015, Sanders et al., 2013, Sharma et al., 2015).
Minimally Engineered MT145 (MT145K) Trimer Binds Prototype V2-Apex bnAb Precursors
One property thought to be critical for vaccine immunogens to select rare bnAb precursors is the ability to effectively bind to UCA B cell receptors (Dosenovic et al., 2015, Escolano et al., 2016, Jardine et al., 2015, McGuire et al., 2016, Steichen et al., 2016). Therefore, to gain or improve binding of the V2-apex bnAb inferred precursor Abs to the MT145 Env trimer, we substituted a glutamine (Q) with a lysine (K) residue (HXB2 position 171) in strand C of the V2-apex bnAb core epitope (Figures 1A and 1B). We based this substitution on the presence of a positively charged motif (KKKK) in CRF250 and CP256.SU strand C V2 Env sequences, both of which bind V2-apex bnAb prototype precursors (Andrabi et al., 2015, Bhiman et al., 2015, Doria-Rose et al., 2014, Gorman et al., 2016). ELISA binding revealed strong binding of the mature V2-apex bnAb prototypes with the MT145-WT trimer and weak but detectable binding with one of the UCA Abs, CAP256 UCA (Figure 1C). Strikingly, binding with our V2-engineered MT145 trimer (henceforth referred to as MT145K) not only improved binding to CAP256 UCA Ab but also conferred binding on both PG9 and CH01 inferred germline-reverted (iGL) Abs (Figure 1C). The PG9 and CH01 iGL Abs used here had diversity (D; heavy chain) and joining (J; both heavy and light chains) genes reverted to their corresponding germline gene families in the CDRH3s, in addition to the VH and VL regions reported previously (Andrabi et al., 2015, Gorman et al., 2016).
Figure 1.

Design of a Chimpanzee Env-Stabilized Trimer and Binding to V2-Apex bnAb iGL Abs
(A) Structural arrangement of the V2-apex bnAb core epitope region on the BG505.664 soluble Env trimer (modified from Garces et al., 2015; PDB: 5CEZ). The ribbon representation of V1V2 loop strands that form the trimer apex show a cluster of positively charged lysine-rich peptide regions (HXB2-R166-K171: R or K residues shown as blue spheres) and the two glycans N156 and N160 (depicted in green spheres with lines). The side chains of the positively charged residues intersperse with the side chains of residues from adjacent protomers to form a continuous positively charged surface at the tip of the trimer to provide a minimal V2-apex bnAb epitope.
(B) Amino-acid sequence alignment of strand B and C V2 of HIV CRF250, CAP256.SU, chimpanzee SIV MT145 WT, and its V2-modified variant (Q171K), MT145K. Glutamine (Q) at position 171 (shown in red) was substituted with lysine (K) in MT145 Env to gain binding to V2-apex bnAb inferred germline (iGL) Abs.
(C) ELISA binding of mature V2-apex bnAbs, PG9, CAP256.09, and CH01 and their iGL versions to WT MT145 (red) and MT145K SOSIP trimers.
(D) Octet binding curves (association, 120 s [180–300]; dissociation, 240 s [300–540]) of CAP256 UCA and CH01 iGL Abs and their respective mature Ab versions (CAP256.09 and CH01) to MT145K trimer, its glycan knockout (N160K) variant, K-rich core epitope substituted variants, and the corresponding monomeric gp120. The Abs were immobilized on human IgG Fc capture biosensors and 1uM trimer or gp120 proteins used as analytes. The binding response is shown in nanometers (nm).
Previous mapping studies have defined the HIV core epitope recognized by mature V2-apex bnAbs (Andrabi et al., 2015, Gorman et al., 2016, Landais et al., 2017, Lee et al., 2017, McLellan et al., 2011, Pancera et al., 2013, Walker et al., 2009). To examine the contributions of V2-apex core epitope glycan and protein residues to binding by V2-apex bnAb iGL Ab versions, we generated MT145K strand C peptide and glycan trimer variants that are known to eliminate interactions of V2-apex bnAbs with the Env trimer (Andrabi et al., 2015, McLellan et al., 2011, Pancera et al., 2013). Bio-layer interferometry (BLI or octet) binding analyses of the iGL Abs with these trimer variants showed that glycan and/or peptide epitope requirements of precursor Abs were largely similar to the requirements of corresponding mature Abs (Figure 1D), suggesting that most contacts with the MT145K V2-apex core epitope are already encoded in the germline configuration for this class of bnAbs. Notably, the mature Abs showed slightly more tolerance to changes within the core protein epitope, particularly for the CAP256.09 bnAb, suggesting that part of the affinity maturation in this class of Abs may be to accommodate variation within the bnAb V2-apex core epitope. Overall, the strand C V2-modification in the MT145 SOSIP.664 trimer conferred binding to multiple V2-apex bnAb germline prototypes.
Architecture of the MT145K Trimer
We solved the structure of the MT145K trimer by cryo-EM to a global resolution of ∼4.1 Å (Figure S2; Table S1). To date, our structure represents the only atomic-level structure of an SIV Env trimer. Like other class I fusion proteins, protomers (gp120 and gp41) of MT145K trimerize to form a metastable pre-fusion Env trimer (Figures 2A, 2B, and S3) (Kwon et al., 2015, McLellan et al., 2013, Pallesen et al., 2016, Pallesen et al., 2017, Stevens et al., 2004). The trimer architecture exhibits a mushroom-like shape with subunits gp120 and gp41 constituting the envelope-distal and proximal entities, respectively (Figure 2B). Overall, the MT145K trimer configuration closely resembles that of the trimeric HIV Env spike, with an overall Cα root-mean-square deviation (RMSD) of 1.9 Å (Kwon et al., 2015). The arrangement of the V loops in the MT145K Env trimer is reminiscent of the V-loop arrangement in the HIV Env trimer and is suggestive of a similar role in immune evasion by steric occlusion of underlying conserved epitopes (Julien et al., 2013a, Pancera et al., 2014). Notably, the V1 and V2 loops are largely solvent exposed and occlude access to the underlying V3 loop (Figure 2C). Inaccessibility of the V3 loop is mediated by intra-protomer interactions of V1V2 to V3 and by extensive inter-protomer V1V2 trimer interactions at the apex of the spike. The SIV MT145K Env trimer exhibited well-ordered V2-V5 loops, while V1 is somewhat disordered (Figure 2D).
Figure 2.

Cryo-EM Structure of the MT145K Trimer
(A) Schematic showing the MT145K SOSIP soluble trimer design from its full-length gp160 Env sequence. The gp120 constant (C1-C5) and variable (V1-V5) regions and the gp41 regions (fusion peptide, FP; heptad repeat, HR1 and HR2; membrane proximal external region, MPER; transmembrane, TM; and cytoplasmic tail, CT) are indicated. The N-linked glycan positions for each NXT or NXS residue are labeled according to the HIV HXB2 numbering scheme. The SOSIP trimer stabilizing modifications include (i) disulfide bond, A501C-T605C; (ii) R6 cleavage site; (iii) I559P; and (iv) 664-residue truncation in gp41 MPER. The substitution to incorporate a K residue at position 171 (Q171K) to gain binding for V2-apex iGL Abs is indicated in blue.
(B and C) Side (B) and top (C) views of the unliganded MT145K trimer model based on the cryo-EM density map at ∼4.1-Å resolution. Ribbon representations of the MT145K trimer spike, in which the subunits gp120 (cornflower blue) and gp41 (orange) are depicted on one protomer. The gp120 variable loops (V1-V5) positioned to the trimer periphery are depicted in different colors (V1, khaki; V2, red; V3, magenta; V4, yellow; and V5, chartreuse). The fusion peptide region of gp41 is shown in cyan. Glycan sugar residues modeled based on density are represented in forest green stick form.
(D) Superimposition of variable loops (V1-V5) and fusion peptide region for MT145K and unliganded HIV clade A BG505 (PDB: 4ZMJ) SOSIP trimers. The dotted lines indicate regions in the V loops or FP for which the observed electron density was absent or unclear.
(E) Structural comparison of gp41 regions of the MT145K (orange) and BG505 (gray) trimers. The gp41 structural elements overall show a similar arrangement except for the fusion peptide region (colored cyan on MT145K and pink on BG505), which is exposed on the BG505 trimer but remains hidden in a pocket inside the MT145K trimer.
Proximal to the viral membrane is the gp41 subunit that forms the base of the trimer spike and is arranged into heptad repeat-1 (HR1), HR2, and the fusion peptide (FP) (Figure 2B). Similar to the HIV Env trimer, the three C-terminal helices of HR1 are centrally positioned along the trimer axis perpendicular to the viral membrane (Julien et al., 2013a, Lyumkis et al., 2013, Pancera et al., 2014). Intriguingly, the FP region, which has been observed to be solvent exposed on the outside of the HIV Env, is positioned in a pocket inside the MT145K trimer and remains sequestered in all three protomers (Figure 2E).
Conservation of the Glycan Shield on HIV and Chimpanzee SIV Env Trimers
To compare the nature of the glycan shield on SIVcpzPtt Env and HIV Env, we performed site-specific glycan analysis of the MT145K trimer. The overall oligomannose content of the MT145K trimer is similar to HIV Env (Figures 3A and 3B) (Panico et al., 2016, Pritchard et al., 2015). However, although the distributions differ from the HIV clade A strain BG505, which is dominated by Man9GlcNAc2 oligomannose-type glycans, MT145K is predominantly Man8GlcNAc2 (Behrens et al., 2016). In addition, further processing was evident in the MT145K trimer, which showed elevated Man6-7GlcNAc2 structures (Figure 3B). The outer domain of gp120 presents a high density of oligomannose glycans that form the intrinsic mannose patch (Bonomelli et al., 2011), which is a highly conserved feature across the two viral species. The apex of the MT145K trimer possesses oligomannose-type glycans at N160 that correspond to the trimer-associated mannose patch (TAMP) also observed on HIV Env (Behrens et al., 2017). As for HIV, glycans at the base of the trimer at N88 and on gp41 of the MT145K trimer are extensively processed (Figures 3A and 3C).
Figure 3.

Site-Specific Glycoform Composition of MT145K Trimer
(A) Site-specific glycoform quantification of the MT145K SOSIP soluble trimer. MT145K trimers from transiently transfected HEK293F cell expressed supernatants were affinity purified by the quaternary trimer-specific antibody, PGT145. The purified MT145K trimers were treated separately with three proteases—trypsin, chymotrypsin, and elastase—and the digests were enriched for glycopeptides and analyzed by liquid chromatography-electrospray ionization mass spectrometry (LC-ESI MS). The individual glycan compositions of the N-linked glycan sites (n = 26) are represented by bar graphs that indicate the relative abundance of each glycoform species and are derived from the mean of two analytical replicates. The pie charts summarize the proportion of glycoforms for each site and this information is color coded: oligomannose type in green and complex and/or hybrid glycans in pink. The glycoforms at N262 and N268 positions (indicated by “∗”) could not be separately determined by enzymatic digestion and the bars represent the average glycan compositions across both sites.
(B) Hydrophilic interaction ultra-performance liquid chromatography (HILIC-UPLC) profiles of the total N-linked glycans released from the MT145K trimers. The proportions of oligomannose plus hybrid glycan contents and complex-type glycans are represented in green and pink colors, respectively.
(C) Modeled glycan shields for the MT145K and BG505 SOSIP trimers. Man9GlcNAc2 oligomannose-type glycans were docked and rigid-body fitted at each of the corresponding Env glycan positions using the MT145K structure (determined in this study [PDB: 6OHY]) and the unliganded BG505 SOSIP.664 trimer structure (Kwon et al., 2015; PDB: 4ZMJ). Top and side views of the trimers are shown and the individual glycan sites are labeled and color coded based on the content of oligomannose: green, 100%–80%; orange, 79%–20%; and pink, 19%–0%.
The remarkable conservation in the overall architecture of the chimpanzee SIV and HIV Env glycan shield, despite sharing only ∼62% of the amino-acid sequence identity, suggests that the glycan shield has an indispensable role in immune evasion and potentially maintaining the functional integrity of the trimer spike. Indeed, the glycan shield is integral to all lentiviral envelopes and appears to have evolved somewhat specifically to mammalian hosts (Figure S4). Over the course of lentiviral evolution, the Env glycan density shows an overall gradual progression and likely peaked in retroviruses infecting non-human primates and plateaued in HIV Envs (Figure S4) (Zhang et al., 2004). Therefore, the high-density Env glycan shield on HIV must have been established well before chimpanzee SIV crossed into humans. Nevertheless, several glycan positions on HIV Env appear to have subtly shifted after the species crossover: presumably as a result of adaptation to the human immune system (Figure S5).
MT145K Binds V2-Apex bnAbs Almost Exclusively
To define the overall antigenicity of the MT145K trimer, we first assessed the neutralization sensitivity of the MT145K virus (MT145-Q171K) to a broad panel of HIV Env-specific neutralizing and non-neutralizing (nnAbs) mAbs and compared these profiles to those for the clade A BG505 HIV virus (Figures 4A and S6) (Sanders et al., 2013, Voss et al., 2017). Remarkably, the V2-apex bnAbs, but essentially no other bnAbs or nnAbs (except 35O22 gp120-41 interface mAb), exhibited potent neutralizing activities against the MT145K virus (Figures 4A and S6). As previously observed, the BG505 isolate was sensitive to neutralization by all of the bnAbs in the panel, but none of the nnAbs (Figures 4A and S6).
Figure 4.

Antigenic Profile of the MT145K Trimer
(A) HIV Env-specific mAbs were used to characterize the antigenicity of the MT145K Env trimer. MAbs targeting neutralizing and non-neutralizing epitope specificities, including V2-apex, V3-N332, linear V3, CD4bs, CD4i, and the gp120-41 interface were tested with MT145K and BG505 Env-encoding pseudoviruses in a TZM-bl cell-based reporter assay. The reciprocal IC50 neutralization titers for each virus are indicated as dot plots; plots for individual epitope specificities are depicted separately. The neutralization sensitivity comparison of BG505 and MT145K viruses against the mAb panel shows a selectively potent neutralization of MT145K by V2-apex bnAbs but no other bnAbs, except a single gp120-gp41 interface bnAb, 35022. The BG505 virus was neutralized by bnAbs targeting diverse Env sites.
(B) The above mAb panel was further tested with PGT145 Ab-purified MT145K trimer and Galanthus nivalis lectin (GNL)-purified MT145K gp120 monomer by ELISA. The binding, represented as EC50 binding titers, shows selective binding of MT145K by V2-apex bnAbs. Two of the gp120-gp41 interface bnAbs and a CD4i mAb also showed significant binding to the MT145K trimer. Four of the non-neutralizing mAbs specific to a linear V3 epitope exhibited binding to MT145K gp120, but not to the trimer.
Next, we evaluated binding of the MT145K trimer and monomeric gp120 to a panel of mAbs by ELISA and BLI. Consistent with the neutralization results above, bnAbs to the V2-apex site showed robust binding to the MT145K trimer (Figures 4B, S6, and S7), but other bnAbs and nnAbs did not bind, except for a few mAbs that displayed very weak binding (Figures 4B, S6, and S7). PG9, 17b, and some of the linear V3-loop directed mAbs (2557, 3074, 3904, and 14e) (Figures 4B and S6) displayed binding to the MT145K gp120 monomer. The results suggest that the sequence-dependent epitopes for some of the non-neutralizing V3-loop mAbs are present on monomeric MT145K gp120, but are obscured on the MT145K trimer, as indicated by the MT145K structure. Virus neutralization and trimer binding by mAbs is strongly correlated (p = 0.003), consistent with the notion that the MT145K soluble trimer adopts a native-like trimeric Env configuration and displays antigenic properties suitable for a vaccine immunogen.
HIV bnAb Epitopes on Chimpanzee SIV Env
To gain insight into the differences in the HIV Env bnAb epitopes on MT145K SIV Env that may potentially explain the reactivity of V2-apex bnAbs and non-reactivity of HIV bnAbs targeting other Env epitopes, we took advantage of the previously determined structures of human HIV bnAbs in complex with various HIV Env forms and compared the corresponding epitope regions with those on the MT145K Env (Garces et al., 2014, Lee et al., 2016, Lee et al., 2017, Ozorowski et al., 2017, Pejchal et al., 2011, Wu et al., 2010). A lysine-rich patch in strand C of the V2 loop (166RDKKQK171 on BG505 Env) and two nearby glycans N160 and N156 form the core epitope for V2-apex bnAbs on HIV Envs (Figures 5A and S8) (Gorman et al., 2016, Julien et al., 2013b, Lee et al., 2017, McLellan et al., 2011, Pancera et al., 2013). Both of these features are conserved on the MT145K trimer, thus enabling the human V2-apex bnAbs to be effective against the SIV Envs (Figures 5A, S5, and S8) (Barbian et al., 2015).
Figure 5.

A Close-Up View of Regions on the MT145K Trimer That Correspond to Those Recognized by HIV bnAbs on HIV Trimers
(A) V2-apex bnAb binding region: cryo-EM model of PGT145 bnAb (HC, transparent sandy brown; LC, transparent orchid) in complex with the BG505 SOSIP trimer depicting V1V2 loops in ribbon representation (Lee et al., 2017; PDB: 5V8L). The strand C K-rich region (166RDKKQK171; red spheres) and the glycan N160 (forest green sticks) that form the epitope for PGT145 bnAb are indicated. The elements in the core epitope interact with the CDRL1 loop and the long CDRH3 loop that penetrates through glycans to reach the positively charged surface underneath. Both glycan N160 and the positively charged protein residues are conserved between BG505 HIV and MT145K SIV Env trimers.
(B) V3-glycan bnAb binding region: cryo-EM model of PGT128 bnAb (HC, transparent sandy brown; LC, transparent orchid) in complex with the BG505 SOSIP trimer (Lee et al., 2015; PDB: 5ACO). The V3 loop protein backbone residues (324GDIR327; depicted in purple spheres) and the glycans N301 and N332 form the bnAb epitope and are shown to interact with the antibody CDR loops. The MT145K trimer has a glycan at N334 rather than N332 and the glycan points away from the expected location of the PGT128 Ab paratope. In addition, MT145K Env has glycans at two positions, N412 (positioned differently on HIV Env) and N442 (absent on HIV Envs), and particularly the latter will clash with PGT128 CDRH2 and prevent it from interacting with the protein part of the epitope.
(C) The gp120-gp41 interface bnAb binding region: cryo-EM model of PGT151 bnAb bound to a membrane-extracted clade B JRFL Env trimer. The structure depicts PGT151 bnAb CDRs interacting with gp120 and the gp41 interface regions (Lee et al., 2016; PDB: 5FUU). PGT151 CDRH3 interacts with the epitope formed by the protein backbone (in both gp120 and gp41), including the fusion peptide (depicted in pink) and the gp120 (N88, N448) and gp41 (N611 and N637) glycans (not shown). PGT151 Ab CDR loops interact with the FP region on the BG505 trimer. The MT145K trimer FP region (cyan) remains hidden inside the trimer.
(D) Cryo-EM model of two-domain human sCD4 with the B41 SOSIP trimer (Ozorowski et al., 2017; PDB: 5VN3). The structure shows how the Phe43 residue on sCD4 stacks into the Env cavity lining Trp427. This Trp427 cavity is conserved between HIV and MT145K Envs to accommodate CD4 binding.
(E and F) The CD4bs bnAb binding region: crystal structure of VRC01 bnAb in complex with 93TH057 gp120 (Zhou et al., 2010; PDB: 3NGB). The structure depicts VRC01 CDRH3, CDRL3, and CDRL1 loops interacting with the protein residues in loop D (HXB2: 278-282) and the glycan at N276 (E and F, left panels). The MT145K trimer lacks the N276 glycan and bears glycan N236 (unique to SIV Env) in place of N234 that would clash with the VRC01 CDRL1 loop (E and F, right panels). Additionally, the MT145K Env trimer has a longer gp120-V5 loop due to a six-amino-acid insertion at HIV HXB2-456 residue that would shift the loop such that it clashes with the CDRH2 the VRC01 Ab.
Binding of one of the V3-N332 epitope-specific bnAbs, PGT128, predominantly relies on the N332 glycan and a neighboring peptide motif 324GDIR327 at the base of the V3 loop (Figures 5B and S8) (Garces et al., 2014, Gristick et al., 2016, Kong et al., 2013, Pejchal et al., 2011, Sok et al., 2016b). The lack of binding to the MT145K trimer by PGT128 and other bnAbs in this class can be explained by the absence of the N332 glycan on this Env. In contrast, three of the four core protein epitope residues 324G-325D-327R are conserved on the MT145K trimer and, in fact, on other chimpanzee SIV Envs (Figures 5B, S5, and S8). For the PGT128 class bnAbs, the interaction with glycan N332 can be substituted by the N295 glycan observed in some HIV isolates, but not by glycan N334 that is present on the MT145K trimer (Sok et al., 2014a). In fact, the MT145K N334 glycan is positioned in a direction away from the V3-N332 epitope site, making it impossible to facilitate bnAb binding to this epitope. Strikingly, the majority of the known SIVcpz Env sequences possess an N334 glycan in place of the more common N332 glycan on the HIV Env, which appears to be a significant glycan shift upon species crossover as the virus established itself in humans (Figure S5). In addition, the glycans at N442 in the gp120-C4 region and N412 in the gp120-V4 region of MT145K Env may obstruct PGT128 binding. The glycan at N442, unique to the MT145K Env trimer and several other SIVcpz Envs, may clash with CDRH2 of PGT128 and perhaps other bnAbs in this class and may prevent them from accessing the epitope (Figures 5B and S5).
PGT151 represents another glycan-targeting bnAb class (Blattner et al., 2014, Falkowska et al., 2014, Lee et al., 2016) that recognizes several glycans on gp120 (N88 and N448) and gp41 (N611 and N637), as well as the fusion peptide. All glycans and fusion peptide residues that contribute to the PGT151 epitope are conserved between HIV and SIVcpz Envs (Figure S5). Therefore, the lack of PGT151 binding to MT145K is most likely attributable to inaccessibility of the FP on MT145K (Figure 5C).
The CD4bs is conserved between HIV and SIV to the extent that there is cross-species reactivity with sCD4. Human CD4-IgG2 immunoadhesin showed strong binding to the MT145K trimer, indicating a strong cross-species conservation of the Env CD4bs. Phe43 in domain-1 of human sCD4 would fit well inside the Trp427 Env cavity on the MT145K trimer, reminiscent of its interaction with the HIV Env BG505 trimer (Figure 5D) (Ozorowski et al., 2017). However, the MT145K trimer is non-reactive with CD4bs bnAbs. VRC01, one of the bnAbs in this class, binds to the HIV Env CD4bs bnAb epitope formed by discontinuous protein backbone elements including loop D of the gp120-C2 region and bordered by a glycan at N276 (Figures 5E, 5F, S5, and S8) (Wu et al., 2010). MT145K lacks the N276 glycan and the proximal N234 glycan, present in most HIV Envs, but instead has a glycan at position 236. Differences in the loop D sequence (Figure S5) and the glycan at N236, which may clash with VRC01 CDRL1 and CDRL3 loops (Figure 5F), on the MT145K trimer likely impose the biggest impediment to VRC01 binding. Further, the MT145K gp120-V5 loop has a six-amino-acid insertion at HXB2 position 456 compared to HIV Envs that may clash with the VRC01 LC (Figures 5F and S5).
Overall, the non-reactivity of HIV Env bnAbs with the MT145K trimer can be largely ascribed to subtle glycan shifts that have occurred in HIV from chimpanzee SIV Env as the virus established itself in humans.
The Engineered MT145K but Not the MT145-WT Trimer Activates V2-Apex UCA-Expressing B Cell Precursors In Vivo
To determine whether the engineered chimpanzee MT145K trimer could efficiently activate HIV V2-apex Ab germline-encoding precursor B cells in vivo and how it compares with the MT145-WT trimer, we conducted immunization experiments in the CH01 UCA “HC only” knockin (KI) mouse model. This KI-mouse model expresses the pre-rearranged heavy chain (VHDDJH) of the CH01 V2-apex bnAb UCA paired with wild-type (WT) mouse light chains. We immunized two groups of five CH01 UCA heavy chain (HC)-only KI mice, each with two repeated doses (at weeks 0 and 4) of the MT145-WT or MT145K trimer (Figure 6A). To track the development of Ab responses, we performed ELISA assays of the pre-bleed, 2-week (day 14) post-prime (Bleed #1) and 2-week post boost-1 (day 42) (Bleed #2) serum samples with MT145K SOSIP trimer protein and its N160-glycan-eliminated variant (MT145K N160K) (Figure 6B). The N160 glycan is a critical component of the V2-apex bnAb epitope (Lee et al., 2017, McLellan et al., 2011, Pancera et al., 2013, Walker et al., 2009). The pre-bleed serum samples in both immunization groups exhibited weak binding activity with the MT145K trimer that was dependent on the N160 glycan, consistent with the presence of CH01 UCA Abs that do show some binding to the MT145K trimer, as described above (Figure 6B).
Figure 6.

Immunogenicity of MT145-WT Compared to MT145K Trimers in CH01 UCA HC-Only Knockin Mice
(A) Schematic showing immunization schedule of CH01 UCA HC-only KI mice with MT145-WT and engineered MT145K trimers. The CH01 UCA HC-only KI mice were immunized twice with 25 μg of the soluble trimer with glucopyranosyl lipid adjuvant stable emulsion (GLA-SE) as adjuvant. Time points for immunization and bleeds are indicated.
(B) ELISA binding of the MT145-WT and MT145K group trimer-immunized CH01 UCA HC-only KI mice serum samples (pre-bleed, Pre; 2 weeks post prime, Bleed #1; and 2 weeks post boost-1, Bleed #2) with soluble MT145K SOSIP and its glycan knockout variant (MT145K N160K) trimers.
(C) Neutralization titrations of the MT145-WT and MT145K group trimer immunized CH01 UCA HC-only KI mice sera (pre-bleed, Pre; post prime, Bleed #1; and post boost-1, Bleed #2) with MT145K virus and a CH01-sensitive virus (Q23_17). The 3-fold diluted sera were tested against the viruses in a TZM-bl reporter cell assay.
(D) The ID50 neutralization titers of the MT145-WT and MT145K group trimer-immunized CH01 UCA HC-only KI mice sera (pre- and post-immunization bleed time points). Neutralization was assessed against the priming immunogen-matched autologous viruses in each group (MT145-WT group, MT145-WT virus; MT145K group, MT145K virus), the N160 glycan knockout variant of the MT145K virus (MT145K N160A), and a highly CH01-sensitive virus, Q23_17. The numerical values shown in the table represent the ID50 neutralization titers of the immune serum samples and were calculated by non-linear regression method from the percent neutralizations of serum titrations with virus.
The immunogen-specific titers of the serum Ab responses post-prime immunizations (Bleed #1 samples) marginally increased in the MT145K group but remained largely unchanged in the MT145-WT trimer immunized group. The serum Ab titers post-boost-1 immunization (Bleed #2) increased in both the groups and were orders of magnitude higher as compared to the pre-bleed or the post-prime Ab binding responses (Figure 6B). At this immunization step, the serum Ab responses in the MT145K trimer immunized group were solely dependent on the N160 glycan while those in the MT145-WT trimer immunization group were mostly independent of the N160 glycan (Figure 6B). Therefore, we conclude that the engineered MT145K trimer, but not the MT145-WT, efficiently triggers the epitope-specific V2-apex bnAb UCA encoding B cell precursors in vivo. Remarkably, immunizations with the Q171K-substituted engineered MT145K trimer also appeared to eliminate the non-V2-apex bnAb site Env-specific off-target B cell responses that were elicited in the MT145-WT trimer immunization group (Figure 6B). The results demonstrate that the activation of the HIV Env bnAb-encoding unmutated B cell precursor by immunogens that display binding to their UCA Ab versions is critical for eliciting epitope-specific Ab responses and the findings are consistent with studies that specifically use germline-targeting immunogen molecules to kick off the bnAb precursor encoding B cell responses in vivo (Dosenovic et al., 2015, Escolano et al., 2016, Jardine et al., 2015, McGuire et al., 2013, Sok et al., 2016a, Steichen et al., 2016, Tian et al., 2016).
Next, we evaluated immune sera for neutralization of autologous and heterologous viruses. Reproducible MT145K autologous virus-specific neutralizing Ab responses were induced in the MT145K immunization group but not in the MT145-WT immunization group (Figures 6C and 6D). As for the ELISA binding responses, the nAb titers in the MT145K trimer immunized group increased at 2 weeks post prime, as indicated by nAb titers against a highly CH01-sensitive HIV Env-encoding virus (Q23_17) and, further, markedly increased after the boost-1 immunization (Figure 6C). At this point, all animals in the MT145K group developed autologous virus-specific nAb responses (Figure 6C). The nAb responses in MT145K trimer-immunized animals mapped to the glycan N160 and strand C K171 residue, both of which form part of the core epitope for V2-apex bnAbs, suggesting that the MT145K trimer successfully primed V2-apex UCA B cells in an epitope-specific manner in vivo.
Overall, we conclude that the engineered chimpanzee MT145K but not the MT145-WT trimer activated the V2-apex-specific bnAb precursor B cells in a UCA-expressing mouse model.
HIV Trimer Cocktail Boosting Recalls Chimpanzee MT145K Trimer-Induced V2-Apex B Cell Responses in the CH01 UCA Model
Evaluation of the utility of the MT145K trimer in a sequential HIV immunization regime will be best carried out in humans. Nevertheless, we were interested to investigate if the chimpanzee SIV MT145K trimer-induced B cell responses could be boosted by HIV trimers that share conservation at the V2-apex bnAb site with the MT145K trimer, in the CH01 UCA KI mice. We further boosted the MT145K trimer immunized CH01 KI mice with a three-trimer cocktail (C108, WITO, and ZM197-ZM233V1V2 SOSIPs) (Figure 7A), derived from HIV subtype AG, B, and C viral isolates previously shown to be sensitive to CH01 bnAb lineage (Andrabi et al., 2015, Bonsignori et al., 2011, Gorman et al., 2016). Following boosting with the HIV trimer cocktail, the week-10 serum Ab responses (Bleed #4: 2-weeks post HIV trimer cocktail boosting) displayed cross-neutralizing activities against a panel of CH01-class bnAb sensitive heterologous HIV isolates (Figure 7B; Table S2). The serum Ab responses mapped entirely to the V2-apex bnAb epitope (Table S2), suggesting a successful nAb recall response to the V2-bnAb site upon HIV trimer boosting. Notably, the MT145K trimer prime boosting itself led to the development of sporadic cross-neutralizing responses against a few CH01-class bnAb sensitive HIV isolates (Bonsignori et al., 2011) (Table S2). However, HIV trimer boosting slightly improved further the neutralization breadth against heterologous viruses (Table S2), suggesting that the boosting immunizations resulted in the development of B cell responses along favorable pathways.
Figure 7.

Boosting Chimpanzee SIV MT145K Trimer-Induced B Cell Responses with an HIV Trimer Cocktail in CH01 UCA HC-Only Knockin Mice
(A) Schematic showing immunization schedule of CH01 UCA HC-only KI mice with a chimpanzee SIV MT145K trimer followed by an HIV Env-derived three-trimer cocktail. A group of five animals was immunized with two doses (prime, week 0; boost-1, week 4) of the MT145K trimer (as also shown in Figure 6). The animals were further boosted (boost-2 at week 8) with an HIV Env-derived three-trimer cocktail (C108, WITO, and ZM197-ZM233V1V2). The V1V2 loops on trimer cartoons are depicted in red to highlight that the region is shared between HIV and SIV Env trimers. The CH01 UCA HC-only KI mice were immunized with 25 μg of the soluble MT145K trimer or HIV three-trimer cocktail (25 μg total) with GLA-SE as adjuvant. Time points for the immunizations and the bleeds are indicated.
(B) The ID50 virus neutralization titers of the Bleed #4 serum samples collected post boost-2 immunization with an HIV three-trimer cocktail. The neutralization of the post-immune sera was assessed against the priming immunogen-matched autologous virus, MT145K, the boosting immunogen-matched, CH01 sensitive viruses, and global panel HIV Env-encoding viruses. Each filled circle in the plot represents virus ID50 neutralization values for the individual animals; an asterisk (∗) indicates that 50% neutralization was not reached at a 1:100 serum dilution.
Interestingly, the trimer-elicited serum Ab responses in the CH01 UCA KI model mapped entirely to the glycan N160, as probed by ELISA binding with MT145K, CRF250 trimers, and their N160-glycan-eliminated trimer variants (Figure S9). The animals, however, did not develop Env backbone-specific off-target B cell responses at any stages of the SIV MT145K or HIV trimer immunizations. This result suggests that the very high frequency of CH01 UCA precursor B cells in the mouse model favors V2-apex responses to the exclusion of off-target responses. Therefore, this model was ultimately found to be unsuitable for evaluation of the advantages of a combined HIV and SIV immunization strategy and an adoptive transfer approach that can generate inter-epitope B cell clonal competition (Abbott et al., 2018, Dosenovic et al., 2018) may be more revealing.
Overall, the analysis of the immune responses revealed that, due to the extraordinary conservation of the V2-apex bnAb epitope region between HIV and chimpanzee SIV, the MT145K trimer successfully primed human V2-apex bnAb UCA-encoding mouse B cells and induced a V2-focused cross-neutralizing HIV Env-specific response that could be further boosted by HIV Env-derived trimers.
Discussion
Vaccination has taken advantage of related viruses from different species, beginning with the use of cowpox as a smallpox vaccine (Riedel, 2005). HIV is too variable and has too many evasion mechanisms for such an approach applied directly to work effectively. Nevertheless, there are HIV-related viruses that have the potential to be exploited in some form in vaccine design. Indeed, the HIV pandemic is believed to have arisen because of a cross-species virus transmission from chimpanzees to humans during the period from 1910 to 1930 (Korber et al., 2000, Sharp and Hahn, 2011, Worobey et al., 2008). The HIV and chimpanzee SIV Envs, the target of potentially protective neutralizing antibodies, display about 60% sequence conservation at the amino-acid level. Importantly, HIV V2-apex bnAbs have been shown to neutralize certain chimpanzee SIV isolates, including the SIVcpzPtt isolate MT145, suggesting cross-species conservation of this epitope (Barbian et al., 2015). Accordingly, we generated a chimpanzee SIV Env trimer (MT145 SOSIP) and showed that it bound HIV V2-apex bnAbs. We then engineered it to bind to germline-reverted V2-apex bnAbs (MT145K SOSIP) so that it might be useful in activating V2-apex precursors.
The cryo-EM structure of MT145K SOSIP trimer revealed that the Env trimers of HIV and chimpanzee SIV have very similar overall architectures. The glycan shield of chimpanzee SIV forms a similarly dense protective layer to antibody recognition of the protein surface as observed in HIV. However, subtle movements in the locations of the glycans appear to contribute to the inability of the great majority of HIV bnAbs to recognize the chimpanzee SIV Env trimer. As noted above, bnAbs to the V2-apex region of the trimer are the exception. We have hypothesized previously (Lee et al., 2017) that the conservation of this region among HIV isolates is to facilitate trimer disassembly during viral entry. It is interesting that the overall V2-apex structure is conserved across the chimpanzee-human species barrier, indicating its critical importance for Env function.
In order to evaluate the MT145K trimer as an immunogen able to activate V2-apex bnAb precursor B cells, we took advantage of the availability of V2-apex bnAb UCA H-chain-only knockin mice. We compared MT145K and MT145 trimers as immunogens. Following two immunizations, MT145K trimers reproducibly elicited Abs able to neutralize the autologous virus and a few V2-apex Ab-sensitive viruses whereas MT145 trimers failed to induce such nAbs. The specificities of the nAbs were dependent on the glycan at N160 and a lysine on strand C of the V2. Boosting with a cocktail of HIV Env trimers successfully recalled the V2-apex-specific nAb responses and generated some enhanced heterologous neutralization. Therefore, from studies in this mouse model, the MT145K trimer appears to be a promising immunogen to induce V2-apex bnAbs. However, further immune evaluation of MT145K vaccine strategies in animal models expressing bnAb-encoding B cell precursors at frequencies closer to those expected in humans will be important (Abbott et al., 2018, Lee et al., 2014, Osborn et al., 2013).
STAR★Methods
Key Resources Table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Monoclonal anti-HIV-1 Env PGT145 | Produced in house; Walker et al., 2011 | NA |
| Monoclonal anti-HIV-1 Env PG9 | Produced in house; Walker et al., 2009 | NA |
| Monoclonal anti-HIV-1 Env PG9 iGL | Produced in house; This paper | N/A |
| Monoclonal anti-HIV-1 Env CAP256.09 | Produced in house; Doria-Rose et al., 2014 | N/A |
| Monoclonal anti-HIV-1 Env CAP256 UCA | produced in house; Doria-Rose et al., 2014 | N/A |
| Monoclonal anti-HIV-1 Env CH01 | Produced in house; Bonsignori et al., 2011 | N/A |
| Monoclonal anti-HIV-1 Env CH01 iGL | Produced in house; This paper | N/A |
| HIV mAbs | NIH AIDS Reagent Program or various research laboratories | N/A |
| Alkaline Phosphatase-AffiniPure Goat Anti-Human IgG, Fc Fragment Specific | Fisher Scientific | Cat# 109-055-098 |
| Bacterial and Virus Strains | ||
| BG505_W6M_C2 | NIH AIDS Reagent Program | N/A |
| MT145K | This paper | N/A |
| Q23_17 | NIH AIDS Reagent Program | N/A |
| WITO4160_33 | NIH AIDS Reagent Program | N/A |
| C1080_C3 | NIH AIDS Reagent Program | N/A |
| ZM233_6 | NIH AIDS Reagent Program | N/A |
| 12 virus panel – global isolates | NIH AIDS Reagent Program | N/A |
| Chemicals, Peptides, and Recombinant Proteins | ||
| X-tremeGENE 9 DNA Transfection Reagent | Sigma-Aldrich | Cat# 6365809001 |
| PEI MAX 40000 | Polysciences, Inc. | Cat# 24765-1 |
| Galanthus nivalis lectin (snow drop),agarose bound | Vector Labs | Cat#AL-1243 |
| DEAE-Dextran | Sigma-Aldrich | Cat# D9885-10G |
| GAB1 SOSIP.664 | This paper | N/A |
| MB897 SOSIP.664 | This paper | N/A |
| EK505 SOSIP.664 | This paper | N/A |
| MT145 SOSIP.664 | This paper | N/A |
| MT145K SOSIP.664 | This paper | N/A |
| Critical Commercial Assays | ||
| QuikChange II XL Site-Directed Mutagenesis Kit | Agilent Technologies | Cat# 200522 |
| Bright-Glo Luciferase Assay System | Promega | Cat# E2610 |
| Deposited Data | ||
| MT145K structure | This study | EMDB # EMD-20074 PDB # 6OHY |
| Experimental Models: Cell Lines | ||
| HEK293T | ATCC | Cat# CRL-3216 |
| FreeStyle 293-F | Thermo Fisher Scientific | Cat# R79007 |
| TZM-bl cells | NIH AIDS Reagent Program | Cat# 8129 |
| Experimental Models: Organisms/Strains | ||
| CH01 UCA HC-only knock-in mouse model | This study | NA |
| Recombinant DNA | ||
| pSG3Δenv plasmid | NIH AIDS Reagent Program | Cat# 11051 |
| Software and Algorithms | ||
| Prism v7.0 | GraphPad | https://www.graphpad.com/scientific-software/prism/ |
| UCSF Chimera | Pettersen et al., 2004 | N/A |
| Other | ||
| Superdex 200 Increase 10/300 GL column | GE Healthcare | Cat# 28990944 |
| Anti-human IgG Fc Capture (AHC) Biosensors | ForteBio | Cat#18-5060 |
| Titan Krios | Thermo Fisher Scientific | N/A |
| K2 Summit | Gatan | N/A |
| Motioncor2 | Zheng et al., 2017 | N/A |
| GCTF | Zhang, 2016 | N/A |
| Relion | Scheres, 2012 | N/A |
| Rosetta | Rosetta Commons | https://www.rosettacommons.org/ |
| Coot | Emsley and Cowtan, 2004 | N/A |
Contact for Reagent and Resource Sharing
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Dennis R. Burton (burton@scripps.edu).
Experimental Model and Subject Details
Knockin Mice
“CH01 UCA homozygous “HC only” (VHDJH+/+) site-directed transgenic, i.e., knock-in (KI) mice, with homozygously knocked-in CH01 UCA VHDJH rearrangements, were generated on the C57BL/6 background based on previously-described IgH locus-directed gene-targeting techniques (Chen et al., 2013, Verkoczy et al., 2011, Verkoczy et al., 2010). All vaccinations were performed using 6-12-week-old mice, and all immunization groups were equally matched for gender. All mice were cared for in a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC), in accordance with NIH guidelines. All animal procedures were approved by the Duke Institutional Animal Care and Use Committee (IACUC), prior to performance.
Cell Lines
Human Embryonic Kidney (HEK293T) cell line is a highly transfectable version of SV40 T-antigen containing 293 human embryonic kidney cells. The FreeStyle 293-F cell line is a version of 293 cells that are adapted to suspension culture in FreeStyle 293 Expression Medium. TZM-bl is a HeLa reporter cell line expresses CD4 receptor and CXCR4 and CCR5 chemokine co-receptors and luciferase and b-galactosidase genes under the control of the HIV-1 promoter, hence is useful to assay in-vitro HIV infection.
Method Details
SIV Envelope Trimer Design, Expression, and Purification
SOSIP.664 HIV Env trimer modification were incorporated into envelope encoding sequences corresponding to four Chimpanzee (SIVcpzPtt) isolates (GAB1 [GenBank: P17281]; MB897 [GenBank: ABU53023]; EK505 [GenBank: ABD19499]; and MT145 [GenBank: ABD19508]) to express as soluble native-like trimers as described previously (Sanders et al., 2013). Briefly, the following modifications were incorporated into these Envs for soluble trimer expression: a) the Env leader sequence was replaced by Tissue Plasminogen Activator (TPA) signal sequence for higher protein expression; b) a disulfide bond was introduced between gp120 and gp41 subunits by substituting residues A501-C and T605-C respectively in gp120 and gp41; c) the gp120 REKR cleavage site was replaced by Furin inducible R6 site (RRRRRR) for enhancing cleavage efficiency between gp120 and gp41; and d) an I559P substitution in gp41 to stabilize the soluble trimer protein. In addition, a GS-linker and a His-tag were added to the gp41ECTO C terminus at HXB2 residue 664 position. The codon-optimized SOSIP.664 gp140 gene constructs were synthesized (Geneart, Life Technologies) and cloned into the phCMV3 vector (Genlantis). Recombinant envelope proteins were expressed in HEK293F cells as described elsewhere (Sanders et al., 2013). Briefly, HIV Env trimers CRF250, WITO, C108, ZM197-ZM233V1V2 and the 4 chimpanzee SIV SOSIP.664 Env-encoding trimer plasmids were cotransfected with a plasmid encoding for Furin (3:1 ratio) into HEK293F cells using PEI-MAX 4000 transfection reagent (Polysciences, Inc.). The secreted soluble trimers proteins were purified from cell supernatants after 5 days using agarose-bound Gallanthus Nivalis Lectin (GNL) (Vector Labs) or CNBr-activated Sepharose 4B bead (GE Healthcare) bound PGT145 bnAb antibody affinity columns as described previously (Pugach et al., 2015). The affinity-purified proteins were size exclusion chromatography (SEC)-purified with a Superdex 200 10/300 GL column (GE Healthcare) in PBS/TBS. The purified trimers for the immunization experiments were quality control tested for antigenicity with a range of HIV Env-specific neutralizing and non-neutralizing mAbs.
Antibodies, Expression, and Purification
HIV envelope specific mAbs to a broad range of epitopes were used, including those that target V2-apex, V3-N332, linear V3, CD4bs, CD4i and gp120-41 Env sites. A dengue antibody (DEN3) was used as control Ab for binding experiments. For PG9 and CH01 V2-apex bnAb inferred germline antibody designs, the heavy and the light chain V-gene of the mature Abs were reverted to their corresponding closest inferred germline gene sequence as determined using the ImMunoGeneTics (IMGT) website (http://imgt.cines.fr/) (Brochet et al., 2008). The reverted variable heavy and light chain nucleotide sequences were synthesized by Geneart (Life Technologies) and cloned into corresponding Igγ1, Igκ, and Igλ expression vectors as previously described (Tiller et al., 2008), using the Gibson cloning method (NEB, USA). The antibodies were expressed and purified using methods described previously (Sok et al., 2014b). Briefly, the heavy and light chain encoding plasmids were reconstituted (1:1 ratio) in Opti-MEM (Life Technologies), and cotransfected HEK293F cells (Invitrogen) using 293fectin (Invitrogen). The suspension cells were cultured for 4-5 days in a shaker incubator at 8% CO2, 37.0°C, and 125 rpm. The antibody containing supernatants were harvested, filtered through a 0.22 mm Steriflip units (EMD Millipore) and passed over a protein A or protein G affinity column (GE Healthcare). The bound antibody was eluted from the columns in 0.1 M citric acid, pH 3.0. Column fractions containing IgG were neutralized (2M Tris-base), pooled, and dialyzed against phosphate-buffered saline (PBS), pH 7.4. IgG purity was determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and the concentration was determined by measuring the relative absorbance at 280 nm.
Site-Directed Mutagenesis
The amino-acid point mutations in Env-encoding plasmids were incorporated by using a QuikChange site-directed mutagenesis kit (Agilent Technologies, USA), according to the manufacturer’s instructions. All of the mutations were confirmed by DNA sequence analysis (Eton Bioscience, San Diego, CA).
Differential Scanning Calorimetry
Thermal denaturation was analyzed with a differential scanning calorimetry (DSC) using a MicroCal VP-Capillary DSC instrument (Malvern), at a scanning rate of 1 K/min under 3.0 atmospheres of pressure. Samples were dialyzed in PBS pH 7.4 overnight and protein concentration was adjusted to 0.5 mg/mL prior to measurement. DSC data were analyzed after buffer correction, normalization, and baseline subtraction using MicroCal VP-Capillary DSC analysis software provided by the manufacturer.
Negative Stain Electron Microscopy and Data Treatment
Purified M145K sample was deposited on thin-carbon-coated (Edward Auto 306 carbon evaporator) a C-flat EM grid (Cu400 mesh, 2 μm hole diameter, 2 μm hole spacing) (Protochips, Morrisville, NC, USA) and embedded in 2% (w/V) uranyl formate. The carbon-coated grids were Ar/O2-plasma-cleaned (Gatan Solarus Model 950 Advanced Plasma System; Gatan Inc., Pleasanton, CA, USA) prior to sample deposition. The uranyl-stained EM sample was then inserted into an FEI Tecnai 12 microscope (Thermo Fisher Scientific, Waltham, MA, USA) equipped with a US4000 CMOS detector (Gatan Inc., Pleasanton, CA, USA). The data was collected at 52,000X nominal magnification resulting in a pixel size of 2.05Å at the object level. Data was binned by a factor of 2 prior to data treatment. Projection image identification in the micrographs was performed with a difference-of-Gaussians implementation (Voss et al., 2009). Projection images subsequently underwent 2D alignment and classification by iterative multi-reference alignment/multivariate statistical analysis (Ogura et al., 2003).
CryoEM Sample Preparation, Data Collection, Processing, and Analysis
Purified MT145K sample was deposited on a C-flat EM grid (Cu400 mesh, 2 μm hole diameter, 2 μm hole spacing) (Protochips, Morrisville, NC, USA) that had been Ar/O2-plasma-cleaned (Gatan Solarus Model 950 Advanced Plasma System; Gatan Inc., Pleasanton, CA, USA) prior to sample deposition. Excess buffer was then blotted away from the grid followed by plunging into and vitrification in liquid ethane cooled by liquid nitrogen using a vitrobot (Thermo Fisher Scientific, Waltham, MA, USA). The resulting cryo-EM specimen was transferred into an FEI Titan Krios microscope (Thermo Fisher Scientific, Waltham, MA, USA) equipped with a Gatan K2 Summit direct electron detector (Gatan Inc., Pleasanton, CA, USA). Dose-fractionated data was collected in electron counting mode at a nominal magnification of 29,000X resulting in a pixel size of 1.02 Å at the object level. Micrograph movie frame exposure time was 200ms and each movie micrograph was recorded over 10 s (50 movie frames) corresponding to a total dose of 94e-/Å2. Movie micrograph frames were subsequently aligned (MotionCor2; (Zheng et al., 2017)), dose-weighted and signal-integrated resulting in 1,281 micrographs for further data processing. CTF models were determined using GCTF (Zhang, 2016). Candidate projection images of MT145K were identified using a difference-of-Gaussians implementation (Voss et al., 2009). The resulting set of candidate projection images subsequently underwent 2D alignment and classification by use of Relion 2.1b1 (Scheres, 2012). ∼95,000 projection images corresponding to well-formed class averages of MT145K were selected for further data processing. This data class was iteratively angularly refined and reconstructed using a B41 unliganded Env trimer map rendered at 60 Å resolution as an initial reference (Ozorowski et al., 2017). The data class then underwent 3D classification into six classes with the initial reconstruction rendered at 60 Å resolution as reference. From 3D classification, a subset of 44,301 projection images was selected for final data processing comprising CTF model adjustment at the projection-image level (Zhang, 2016) and angular refinement and reconstruction (Scheres, 2012).
Model Building and Refinement
A homology model (Modeler; (Webb and Sali, 2016)) was generated from sequence alignment of MT145K and BG505 and the structure of the latter (PDB: 4TVP). Significant manual rebuilding followed in Coot (Emsley and Cowtan, 2004). A fragment library was then created from the MT145K sequence containing 200 homologous, non-redundant sequences at each MT145K 7-mer position. Library fragment-based, density-guided, real-space rebuilding was then performed (DiMaio et al., 2015) with 319 decoys. The resulting models were evaluated geometrically (MolProbity; (Chen et al., 2010)) and by fit-to-map (EMRinger; (Barad et al., 2015). The overall best model was selected for further iterations of manual rebuilding and multi-decoy, density-guided, real-space, all-atom Rosetta FastRelax refinement. Finally, glycans were manually built in Coot and restricted, density-guided real-space refinement performed in Phenix 1.12 (Adams et al., 2002) followed by model evaluation by MolProbity, EMRinger and Privateer (Agirre et al., 2015).
Global N-Linked Glycan Analysis
The quantifications and structural characterization of the total glycan pool was achieved by cleaving the N-inked glycans from the surface of the glycoprotein using an in-gel digestion with peptide N-glycosidase F (PNGaseF). The resultant glycans were separated into two aliquots. The first was derivatized with 2-aminobenzoic acid (2-AA) and subjected to HILIC-UPLC analysis using an Acquity UPLC (Waters). To quantify the oligomannose content of the released glycans, the labeled samples were treated with endoglycosidase H (endoH), which selectively cleaves oligomannose glycans. Data analysis and interpretation were performed using Empower software (Waters). The second aliquot of released glycans was subjected to negative ion electrospray ion mobility mass spectrometry using a Synapt G2Si mass spectrometer (Waters). Glycan compositions were determined using collision induced dissociation (CID) fragmentation. Data analysis was performed using Waters Driftscope (version 2.8) software and MassLynxTM (version 4.1). Spectra were interpreted as described previously (Harvey et al., 2009). The glycan compositions were used to generate a sample-specific glycan library that was used to search the glycopeptide data to minimize the number of false-positive assignments in site-specific analysis.
LC-MS Glycopeptide Analysis
Site-specific N-glycosylation analysis was performed using proteolytic digestion followed by tandem LC-MS. Prior to digestion, trimers were denatured, reduced and alkylated by incubation for 1h at room temperature (RT) in a 50 mM Tris/HCl, pH 8.0 buffer containing 6 M urea and 5 mM dithiothreitol (DTT), followed by the addition of 20 mM iodacetamide (IAA) for a further 1h at RT in the dark, and then additional DTT (20 mM) for another 1h, to eliminate any residual IAA. The alkylated trimers were buffer-exchanged into 50 mM Tris/HCl, pH 8.0 using Vivaspin columns (GE healthcare) and digested separately with trypsin, elastase and chymotrypsin (Mass Spectrometry Grade, Promega) at a ratio of 1:30 (w/w). Glycopeptides were selected from the protease-digested samples using the ProteoExtract Glycopeptide Enrichment Kit (Merck Millipore) following the manufacturer’s protocol. Enriched glycopeptides were analyzed by LC-ESI MS on an Orbitrap fusion mass spectrometer (Thermo Fisher Scientific), as previously described (Behrens et al., 2016), using higher energy collisional dissociation (HCD) fragmentation. Data analysis and glycopeptide identification were performed using ByonicTM (Version 2.7) and ByologicTM software (Version 2.3; Protein Metrics Inc.), as previously described (Behrens et al., 2016).
Glycan Modeling
Man9GlcNAc2 oligomannose-type glycans were docked and rigid-body fitted at each of the corresponding Env glycan positions using the MT145K structure presented here or an unliganded BG505 SOSIP.664 structure (PDB: 4ZMJ).
Pseudovirus Production
To produce pseudoviruses, Env-encoding plasmids were cotransfected with an Env-deficient backbone plasmid (pSG3ΔEnv) (1:2 ratio) using X-tremeGENE 9 (Sigma-Aldrich) DNA transfection reagent. Briefly, 1X106 cells in 10ml of Dulbecco’s Modified Eagle Medium (DMEM) were seeded in a 100mm x 20mm cell culture dish (Corning) one day prior to transfection. For transfection, 40μl of X-tremeGENE 9 was added to 700μl of Opti-MEM I reduced serum medium (Thermo Fisher) in tube 1. The Env-encoding plasmid (5 μg) and pSG3ΔEnv (10 μg) were added to tube 2 in 700μl of Opti-MEM. The tube 1 and tube 2 solutions were mixed together and incubated for 25 min at room temperature. Next, the transfection mixture was added to the media with 293T cells seeded previously and then distributed uniformly. All pseudoviruses were harvested 48-72 h posttransfection, filtered through 0.22 mm Steriflip units (EMD Millipore) and aliquoted for use in neutralization assays.
Neutralization Assay
Neutralization was measured by using single-round replication-defective HIV Env-pseudoviruses and TZM-bl target cells (Montefiori, 2005, Seaman et al., 2010). 25ul of 3-fold serially diluted mAbs or serum samples were pre-incubated at 37°C for 1h with 25ul of tissue culture infective dose-50 (TCID50) Env-pseudotyped virus in a half-area 96-well tissue culture plate. TZM-bl target cells (5,000 cells/well) in 50μl of DMEM were added and the plates were allowed to grow in humidified incubator at 37°C and 5% Co2. The luciferase activity of the lysed cells was read on instrument (Biotek) after 2-3 days, by adding lysis buffer followed by Brightglow (Promega). The 50% inhibitory concentration (IC50) or 50% inhibitory doses (ID50) was reported as the antibody concentration or serum dilution required to reduce infection by half.
ELISA Binding Assay
ELISA binding experiments were performed as described previously with minor modification (Sanders et al., 2013). ELISA binding with SOSIP.664 trimer proteins with mAbs was carried out by either capturing the trimer proteins onto the anti-His capture antibodies or on the streptavidin coated plates through biotinylated trimers. For trimer biotinylation, the SOSIP.664 proteins were randomly biotinylated using a 2:1 molar ratio of biotin reagent to trimer using the EZ-link-NHS-PEG4-Biotin kit (Thermo Fisher Scientific, 21324). MaxiSorp plates (Thermo Fisher Scientific) were coated overnight at 4C with 2 ug/mL of anti-His Ab (Thermo Fisher Scientific) or 2 ug/mL streptavidin (Thermo Fisher Scientific). Plates were blocked for 1 hr with 3% BSA and washed three times with 0.05% Tween 20-PBS (PBS-T) (pH 7.4). Anti-His or Streptavidin-coated plates were incubated with biotinylated trimers in 1%BSA plus PBST for 1.5 hr and washed three times with PBST. 3-fold serially diluted mAbs or sera were added starting at a maximum concentration of 10 ug/mL (100ug/ml for iGL Abs) (sera at 1:100 dilution) in 1% BSA plus PBST, and incubated at room temperature (RT) for 1.5 hr. Plates were washed three times with PBST. Alkaline-phosphatase-conjugated goat anti-human IgG Fc secondary antibody (Jackson ImmunoResearch Laboratories) was diluted 1:1000 in 1% BSA PBST and added to plates for 1 hr at RT. Plates were washed three times with PBST and incubated with phosphatase substrate (Sigma) for 15 mins and the absorbance at 405 nm recorded. The 50% binding (EC50) was recorded as the half of the maximum binding activity and was calculated by linear regression method using Prism 6 Software.
Bio-Layer Interferometry (BLI) Binding Assay
The binding experiments of Abs to the affinity purified trimers were performed with an Octet K2 system (ForteBio, Pall Life Sciences). Briefly, the mAbs or IgGs (10 ug/mL in PBST) were immobilizing onto hydrated anti-human IgG-Fc biosensors (AHC: ForteBio) for 60 s to achieve a binding response of at least 1.0. After Ab capture, the sensor was placed in a PBST wash buffer to remove the unbound Ab to establish a baseline signal. Next, the IgG immobilized sensor was dipped into a solution containing SOSIP.664 trimer protein as analyte and incubated for 120 s at 1000 rpm. Following this, the trimer bound to IgG immobilized sensor was removed from the analyte solution and placed into the PBST buffer for 240 s at 1000 rpm. The 2- and 4-minute binding intervals respectively denote the association and dissociation binding curves reported in this study. The sensograms were corrected with the blank reference and fit (1:1 binding kinetics model) with the ForteBio Data Analysis version.9 software using the global fitting function. The data are represented as maximum binding response or the association and dissociation curve fits.
Trimer Protein Immunizations in CH01 UCA HC-Only KI Mice
For the immunization experiments, groups of 5 CH01 UCA HC-only knock-in B cell expressing mice were immunized with 25ug of the individual trimer protein or 25ug total protein of the 3-trimer cocktail (Prime, week-0; Boost-1, week-4 and Boost-2, week-8) along with Glucopyranosyl Lipid Adjuvant in stable emulsion (GLA-SE) as adjuvant. Immunizations were administered intramuscular in the leg of each animal with 25 μg of total trimer immunogens. Blood samples were collected at pre-bleed (Pre), and 2-weeks post-prime (Bleed #1), 2-weeks post boost-1 (Bleed #2), 4-weeks post boost-1 prior to HIV trimer boosting (Bleed #3), and post boost-2 (Bleed #4) immunization time-points for the isolation of sera that were tested for presence of neutralizing antibodies in TZM-bl cell based assay. Serum samples were heat inactivated for potential complement activity at 56°C for 0.5 h. Mice used in this study were approved by Duke University Institutional Animal Care and Use Committee-approved animal protocols.
Quantification and Statistical Analysis
Statistical Analysis
Statistical analysis was performed using Graph Pad Prism 7 for Mac, Graph Pad Software, San Diego, California, USA. The IC50 virus neutralization and the EC50 trimer binding Ab titers with MT145K were compared by two-tailed nonparametric Spearman correlation test with 95% Confidence interval. P value of less than 0.05 were treated as significant. Cryo-EM data were analyzed by a difference-of-Gaussian approach (DoG-Picker) to identify molecular projection images for further processing. Maximum Likelihood optimization (RELION) was utilized to determine a subset of data (44,301 molecular projection images) most suitable for refinement to highest possible resolution and to determine projection image Euler angles and planar shifts.
Data and Software Availability
Cryo-EM reconstructions have been deposited in the Electron Microscopy Data Bank (EMDB) under the accession numbers EMD-20074 and the PDB code for this deposition is 6OHY. The GenBank accession numbers for the heavy and light chain inferred germline versions of PG9 and CH01 broadly neutralizing antibodies reported in this paper are MK825341-MK825344.
Acknowledgments
This work was supported by the International AIDS Vaccine Initiative (IAVI) through the Neutralizing Antibody Consortium SFP1849 (D.R.B., A.B.W., I.A.W., and M.C.), the National Institute of Allergy and Infectious Diseases (Center for HIV/AIDS Vaccine Immunology and Immunogen Discovery grant UM1AI100663 to D.R.B., A.B.W., I.A.W., and M.C.), and the Ragon Institute of MGH, MIT, and Harvard (D.R.B.). This study was made possible by the generous support of the Bill and Melinda Gates Foundation Collaboration for AIDS Vaccine Discovery (CAVD; OPP115782 and OPP1084519) and the American people through USAID. We thank Christina Corbaci and for her help in the preparation of the figures.
Author Contributions
R.A., J.P., J.D.A., J.Z., L.V., A.B.W., and D.R.B. designed the experiments. R.A., J.P., J.D.A., G.S., J.Z., N.d.V., G.G., K.P., C.-Y.S., M.P., A.N., and F.G. performed the experiments. H.B.V., I.A.W., M.C., B.H.H., and B.F.H. contributed critical reagents. R.A., J.P., J.D.A., A.B.W., and D.R.B. analyzed the data and wrote the paper, with inputs from other authors. R.A. and D.R.B. conceived the idea of using SIVcpzPtt Env-derived trimer as an HIV vaccine template.
Declaration of Interests
R.A. and D.R.B. are inventors on a patent application filed by International AIDS Vaccine Initiative (IAVI) and The Scripps Research Institute (TSRI) related to the data presented in this work. All other authors declare no competing interests.
Published: May 21, 2019
Footnotes
Supplemental Information can be found online at https://doi.org/10.1016/j.celrep.2019.04.082.
Contributor Information
Andrew B. Ward, Email: andrew@scripps.edu.
Dennis R. Burton, Email: burton@scripps.edu.
Supplemental Information
References
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C. Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity. 2018;48:133–146.e6. doi: 10.1016/j.immuni.2017.11.023. [DOI] [PMC free article] [PubMed] [Google Scholar]; Abbott, R.K., Lee, J.H., Menis, S., Skog, P., Rossi, M., Ota, T., Kulp, D.W., Bhullar, D., Kalyuzhniy, O., Havenar-Daughton, C., et al. (2018). Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity 48, 133-146.e6. [DOI] [PMC free article] [PubMed]
- Adams P.D., Grosse-Kunstleve R.W., Hung L.W., Ioerger T.R., McCoy A.J., Moriarty N.W., Read R.J., Sacchettini J.C., Sauter N.K., Terwilliger T.C. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 2002;58:1948–1954. doi: 10.1107/s0907444902016657. [DOI] [PubMed] [Google Scholar]; Adams, P.D., Grosse-Kunstleve, R.W., Hung, L.W., Ioerger, T.R., McCoy, A.J., Moriarty, N.W., Read, R.J., Sacchettini, J.C., Sauter, N.K., and Terwilliger, T.C. (2002). PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948-1954. [DOI] [PubMed]
- Agirre J., Iglesias-Fernández J., Rovira C., Davies G.J., Wilson K.S., Cowtan K.D. Privateer: software for the conformational validation of carbohydrate structures. Nat. Struct. Mol. Biol. 2015;22:833–834. doi: 10.1038/nsmb.3115. [DOI] [PubMed] [Google Scholar]; Agirre, J., Iglesias-Fernandez, J., Rovira, C., Davies, G.J., Wilson, K.S., and Cowtan, K.D. (2015). Privateer: software for the conformational validation of carbohydrate structures. Nat. Struct. Mol. Biol. 22, 833-834. [DOI] [PubMed]
- Andrabi R., Voss J.E., Liang C.H., Briney B., McCoy L.E., Wu C.Y., Wong C.H., Poignard P., Burton D.R. Identification of Common Features in Prototype Broadly Neutralizing Antibodies to HIV Envelope V2 Apex to Facilitate Vaccine Design. Immunity. 2015;43:959–973. doi: 10.1016/j.immuni.2015.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]; Andrabi, R., Voss, J.E., Liang, C.H., Briney, B., McCoy, L.E., Wu, C.Y., Wong, C.H., Poignard, P., and Burton, D.R. (2015). Identification of Common Features in Prototype Broadly Neutralizing Antibodies to HIV Envelope V2 Apex to Facilitate Vaccine Design. Immunity 43, 959-973. [DOI] [PMC free article] [PubMed]
- Andrabi R., Su C.Y., Liang C.H., Shivatare S.S., Briney B., Voss J.E., Nawazi S.K., Wu C.Y., Wong C.H., Burton D.R. Glycans Function as Anchors for Antibodies and Help Drive HIV Broadly Neutralizing Antibody Development. Immunity. 2017;47:524–537.e3. doi: 10.1016/j.immuni.2017.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]; Andrabi, R., Su, C.Y., Liang, C.H., Shivatare, S.S., Briney, B., Voss, J.E., Nawazi, S.K., Wu, C.Y., Wong, C.H., and Burton, D.R. (2017). Glycans Function as Anchors for Antibodies and Help Drive HIV Broadly Neutralizing Antibody Development. Immunity 47, 524-537.e3. [DOI] [PMC free article] [PubMed]
- Andrabi R., Bhiman J.N., Burton D.R. Strategies for a multi-stage neutralizing antibody-based HIV vaccine. Curr. Opin. Immunol. 2018;53:143–151. doi: 10.1016/j.coi.2018.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]; Andrabi, R., Bhiman, J.N., and Burton, D.R. (2018). Strategies for a multi-stage neutralizing antibody-based HIV vaccine. Curr. Opin. Immunol. 53, 143-151. [DOI] [PMC free article] [PubMed]
- Barad B.A., Echols N., Wang R.Y., Cheng Y., DiMaio F., Adams P.D., Fraser J.S. EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods. 2015;12:943–946. doi: 10.1038/nmeth.3541. [DOI] [PMC free article] [PubMed] [Google Scholar]; Barad, B.A., Echols, N., Wang, R.Y., Cheng, Y., DiMaio, F., Adams, P.D., and Fraser, J.S. (2015). EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods 12, 943-946. [DOI] [PMC free article] [PubMed]
- Barbian H.J., Decker J.M., Bibollet-Ruche F., Galimidi R.P., West A.P., Jr., Learn G.H., Parrish N.F., Iyer S.S., Li Y., Pace C.S. Neutralization properties of simian immunodeficiency viruses infecting chimpanzees and gorillas. MBio. 2015;6 doi: 10.1128/mBio.00296-15. e00296-15. [DOI] [PMC free article] [PubMed] [Google Scholar]; Barbian, H.J., Decker, J.M., Bibollet-Ruche, F., Galimidi, R.P., West, A.P., Jr., Learn, G.H., Parrish, N.F., Iyer, S.S., Li, Y., Pace, C.S., et al. (2015). Neutralization properties of simian immunodeficiency viruses infecting chimpanzees and gorillas. MBio 6, e00296-15. [DOI] [PMC free article] [PubMed]
- Behrens A.J., Vasiljevic S., Pritchard L.K., Harvey D.J., Andev R.S., Krumm S.A., Struwe W.B., Cupo A., Kumar A., Zitzmann N. Composition and Antigenic Effects of Individual Glycan Sites of a Trimeric HIV-1 Envelope Glycoprotein. Cell Rep. 2016;14:2695–2706. doi: 10.1016/j.celrep.2016.02.058. [DOI] [PMC free article] [PubMed] [Google Scholar]; Behrens, A.J., Vasiljevic, S., Pritchard, L.K., Harvey, D.J., Andev, R.S., Krumm, S.A., Struwe, W.B., Cupo, A., Kumar, A., Zitzmann, N., et al. (2016). Composition and Antigenic Effects of Individual Glycan Sites of a Trimeric HIV-1 Envelope Glycoprotein. Cell Rep. 14, 2695-2706. [DOI] [PMC free article] [PubMed]
- Behrens A.J., Harvey D.J., Milne E., Cupo A., Kumar A., Zitzmann N., Struwe W.B., Moore J.P., Crispin M. Molecular Architecture of the Cleavage-Dependent Mannose Patch on a Soluble HIV-1 Envelope Glycoprotein Trimer. J. Virol. 2017;91:e01894-16. doi: 10.1128/JVI.01894-16. [DOI] [PMC free article] [PubMed] [Google Scholar]; Behrens, A.J., Harvey, D.J., Milne, E., Cupo, A., Kumar, A., Zitzmann, N., Struwe, W.B., Moore, J.P., and Crispin, M. (2017). Molecular Architecture of the Cleavage-Dependent Mannose Patch on a Soluble HIV-1 Envelope Glycoprotein Trimer. J. Virol. 91, e01894-16. [DOI] [PMC free article] [PubMed]
- Bhiman J.N., Anthony C., Doria-Rose N.A., Karimanzira O., Schramm C.A., Khoza T., Kitchin D., Botha G., Gorman J., Garrett N.J. Viral variants that initiate and drive maturation of V1V2-directed HIV-1 broadly neutralizing antibodies. Nat. Med. 2015;21:1332–1336. doi: 10.1038/nm.3963. [DOI] [PMC free article] [PubMed] [Google Scholar]; Bhiman, J.N., Anthony, C., Doria-Rose, N.A., Karimanzira, O., Schramm, C.A., Khoza, T., Kitchin, D., Botha, G., Gorman, J., Garrett, N.J., et al. (2015). Viral variants that initiate and drive maturation of V1V2-directed HIV-1 broadly neutralizing antibodies. Nat. Med. 21, 1332-1336. [DOI] [PMC free article] [PubMed]
- Blattner C., Lee J.H., Sliepen K., Derking R., Falkowska E., de la Peña A.T., Cupo A., Julien J.P., van Gils M., Lee P.S. Structural delineation of a quaternary, cleavage-dependent epitope at the gp41-gp120 interface on intact HIV-1 Env trimers. Immunity. 2014;40:669–680. doi: 10.1016/j.immuni.2014.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]; Blattner, C., Lee, J.H., Sliepen, K., Derking, R., Falkowska, E., de la Peña, A.T., Cupo, A., Julien, J.P., van Gils, M., Lee, P.S., et al. (2014). Structural delineation of a quaternary, cleavage-dependent epitope at the gp41-gp120 interface on intact HIV-1 Env trimers. Immunity 40, 669-680. [DOI] [PMC free article] [PubMed]
- Bonomelli C., Doores K.J., Dunlop D.C., Thaney V., Dwek R.A., Burton D.R., Crispin M., Scanlan C.N. The glycan shield of HIV is predominantly oligomannose independently of production system or viral clade. PLoS One. 2011;6:e23521. doi: 10.1371/journal.pone.0023521. [DOI] [PMC free article] [PubMed] [Google Scholar]; Bonomelli, C., Doores, K.J., Dunlop, D.C., Thaney, V., Dwek, R.A., Burton, D.R., Crispin, M., and Scanlan, C.N. (2011). The glycan shield of HIV is predominantly oligomannose independently of production system or viral clade. PLoS One 6, e23521. [DOI] [PMC free article] [PubMed]
- Bonsignori M., Hwang K.K., Chen X., Tsao C.Y., Morris L., Gray E., Marshall D.J., Crump J.A., Kapiga S.H., Sam N.E. Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutralizing antibodies and their inferred unmutated common ancestors. J. Virol. 2011;85:9998–10009. doi: 10.1128/JVI.05045-11. [DOI] [PMC free article] [PubMed] [Google Scholar]; Bonsignori, M., Hwang, K.K., Chen, X., Tsao, C.Y., Morris, L., Gray, E., Marshall, D.J., Crump, J.A., Kapiga, S.H., Sam, N.E., et al. (2011). Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutralizing antibodies and their inferred unmutated common ancestors. J. Virol. 85, 9998-10009. [DOI] [PMC free article] [PubMed]
- Braibant M., Gong E.Y., Plantier J.C., Moreau T., Alessandri E., Simon F., Barin F. Cross-group neutralization of HIV-1 and evidence for conservation of the PG9/PG16 epitopes within divergent groups. AIDS. 2013;27:1239–1244. doi: 10.1097/QAD.0b013e32835ecb42. [DOI] [PubMed] [Google Scholar]; Braibant, M., Gong, E.Y., Plantier, J.C., Moreau, T., Alessandri, E., Simon, F., and Barin, F. (2013). Cross-group neutralization of HIV-1 and evidence for conservation of the PG9/PG16 epitopes within divergent groups. AIDS 27, 1239-1244. [DOI] [PubMed]
- Briney B.S., Willis J.R., Crowe J.E., Jr. Human peripheral blood antibodies with long HCDR3s are established primarily at original recombination using a limited subset of germline genes. PLoS One. 2012;7:e36750. doi: 10.1371/journal.pone.0036750. [DOI] [PMC free article] [PubMed] [Google Scholar]; Briney, B.S., Willis, J.R., and Crowe, J.E., Jr. (2012). Human peripheral blood antibodies with long HCDR3s are established primarily at original recombination using a limited subset of germline genes. PLoS One 7, e36750. [DOI] [PMC free article] [PubMed]
- Briney B., Sok D., Jardine J.G., Kulp D.W., Skog P., Menis S., Jacak R., Kalyuzhniy O., de Val N., Sesterhenn F. Tailored Immunogens Direct Affinity Maturation toward HIV Neutralizing Antibodies. Cell. 2016;166:1459–1470.e11. doi: 10.1016/j.cell.2016.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]; Briney, B., Sok, D., Jardine, J.G., Kulp, D.W., Skog, P., Menis, S., Jacak, R., Kalyuzhniy, O., de Val, N., Sesterhenn, F., et al. (2016). Tailored Immunogens Direct Affinity Maturation toward HIV Neutralizing Antibodies. Cell 166, 1459-1470.e11. [DOI] [PMC free article] [PubMed]
- Brochet X., Lefranc M.P., Giudicelli V. IMGT/V-QUEST: the highly customized and integrated system for IG and TR standardized V-J and V-D-J sequence analysis. Nucleic Acids Res. 2008;36:W503–W508. doi: 10.1093/nar/gkn316. [DOI] [PMC free article] [PubMed] [Google Scholar]; Brochet, X., Lefranc, M.P., and Giudicelli, V. (2008). IMGT/V-QUEST: the highly customized and integrated system for IG and TR standardized V-J and V-D-J sequence analysis. Nucleic Acids Res. 36, W503-W508. [DOI] [PMC free article] [PubMed]
- Chen V.B., Arendall W.B., 3rd, Headd J.J., Keedy D.A., Immormino R.M., Kapral G.J., Murray L.W., Richardson J.S., Richardson D.C. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010;66:12–21. doi: 10.1107/S0907444909042073. [DOI] [PMC free article] [PubMed] [Google Scholar]; Chen, V.B., Arendall, W.B., 3rd, Headd, J.J., Keedy, D.A., Immormino, R.M., Kapral, G.J., Murray, L.W., Richardson, J.S., and Richardson, D.C. (2010). MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12-21. [DOI] [PMC free article] [PubMed]
- Chen Y., Zhang J., Hwang K.K., Bouton-Verville H., Xia S.M., Newman A., Ouyang Y.B., Haynes B.F., Verkoczy L. Common tolerance mechanisms, but distinct cross-reactivities associated with gp41 and lipids, limit production of HIV-1 broad neutralizing antibodies 2F5 and 4E10. J. Immunol. 2013;191:1260–1275. doi: 10.4049/jimmunol.1300770. [DOI] [PMC free article] [PubMed] [Google Scholar]; Chen, Y., Zhang, J., Hwang, K.K., Bouton-Verville, H., Xia, S.M., Newman, A., Ouyang, Y.B., Haynes, B.F., and Verkoczy, L. (2013). Common tolerance mechanisms, but distinct cross-reactivities associated with gp41 and lipids, limit production of HIV-1 broad neutralizing antibodies 2F5 and 4E10. J. Immunol. 191, 1260-1275. [DOI] [PMC free article] [PubMed]
- DiMaio F., Song Y., Li X., Brunner M.J., Xu C., Conticello V., Egelman E., Marlovits T., Cheng Y., Baker D. Atomic-accuracy models from 4.5-Å cryo-electron microscopy data with density-guided iterative local refinement. Nat. Methods. 2015;12:361–365. doi: 10.1038/nmeth.3286. [DOI] [PMC free article] [PubMed] [Google Scholar]; DiMaio, F., Song, Y., Li, X., Brunner, M.J., Xu, C., Conticello, V., Egelman, E., Marlovits, T., Cheng, Y., and Baker, D. (2015). Atomic-accuracy models from 4.5-A cryo-electron microscopy data with density-guided iterative local refinement. Nat. Methods 12, 361-365. [DOI] [PMC free article] [PubMed]
- Doria-Rose N.A., Schramm C.A., Gorman J., Moore P.L., Bhiman J.N., DeKosky B.J., Ernandes M.J., Georgiev I.S., Kim H.J., Pancera M., NISC Comparative Sequencing Program Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature. 2014;509:55–62. doi: 10.1038/nature13036. [DOI] [PMC free article] [PubMed] [Google Scholar]; Doria-Rose, N.A., Schramm, C.A., Gorman, J., Moore, P.L., Bhiman, J.N., DeKosky, B.J., Ernandes, M.J., Georgiev, I.S., Kim, H.J., Pancera, M., et al.; NISC Comparative Sequencing Program (2014). Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature 509, 55-62. [DOI] [PMC free article] [PubMed]
- Dosenovic P., von Boehmer L., Escolano A., Jardine J., Freund N.T., Gitlin A.D., McGuire A.T., Kulp D.W., Oliveira T., Scharf L. Immunization for HIV-1 Broadly Neutralizing Antibodies in Human Ig Knockin Mice. Cell. 2015;161:1505–1515. doi: 10.1016/j.cell.2015.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]; Dosenovic, P., von Boehmer, L., Escolano, A., Jardine, J., Freund, N.T., Gitlin, A.D., McGuire, A.T., Kulp, D.W., Oliveira, T., Scharf, L., et al. (2015). Immunization for HIV-1 Broadly Neutralizing Antibodies in Human Ig Knockin Mice. Cell 161, 1505-1515. [DOI] [PMC free article] [PubMed]
- Dosenovic P., Kara E.E., Pettersson A.K., McGuire A.T., Gray M., Hartweger H., Thientosapol E.S., Stamatatos L., Nussenzweig M.C. Anti-HIV-1 B cell responses are dependent on B cell precursor frequency and antigen-binding affinity. Proc. Natl. Acad. Sci. USA. 2018;115:4743–4748. doi: 10.1073/pnas.1803457115. [DOI] [PMC free article] [PubMed] [Google Scholar]; Dosenovic, P., Kara, E.E., Pettersson, A.K., McGuire, A.T., Gray, M., Hartweger, H., Thientosapol, E.S., Stamatatos, L., and Nussenzweig, M.C. (2018). Anti-HIV-1 B cell responses are dependent on B cell precursor frequency and antigen-binding affinity. Proc. Natl. Acad. Sci. USA 115, 4743-4748. [DOI] [PMC free article] [PubMed]
- Emsley P., Cowtan K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004;60:2126–2132. doi: 10.1107/S0907444904019158. [DOI] [PubMed] [Google Scholar]; Emsley, P., and Cowtan, K. (2004). Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126-2132. [DOI] [PubMed]
- Escolano A., Steichen J.M., Dosenovic P., Kulp D.W., Golijanin J., Sok D., Freund N.T., Gitlin A.D., Oliveira T., Araki T. Sequential Immunization Elicits Broadly Neutralizing Anti-HIV-1 Antibodies in Ig Knockin Mice. Cell. 2016;166:1445–1458.e12. doi: 10.1016/j.cell.2016.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]; Escolano, A., Steichen, J.M., Dosenovic, P., Kulp, D.W., Golijanin, J., Sok, D., Freund, N.T., Gitlin, A.D., Oliveira, T., Araki, T., et al. (2016). Sequential Immunization Elicits Broadly Neutralizing Anti-HIV-1 Antibodies in Ig Knockin Mice. Cell 166, 1445-1458.e12. [DOI] [PMC free article] [PubMed]
- Escolano A., Dosenovic P., Nussenzweig M.C. Progress toward active or passive HIV-1 vaccination. J. Exp. Med. 2017;214:3–16. doi: 10.1084/jem.20161765. [DOI] [PMC free article] [PubMed] [Google Scholar]; Escolano, A., Dosenovic, P., and Nussenzweig, M.C. (2017). Progress toward active or passive HIV-1 vaccination. J. Exp. Med. 214, 3-16. [DOI] [PMC free article] [PubMed]
- Falkowska E., Le K.M., Ramos A., Doores K.J., Lee J.H., Blattner C., Ramirez A., Derking R., van Gils M.J., Liang C.H. Broadly neutralizing HIV antibodies define a glycan-dependent epitope on the prefusion conformation of gp41 on cleaved envelope trimers. Immunity. 2014;40:657–668. doi: 10.1016/j.immuni.2014.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]; Falkowska, E., Le, K.M., Ramos, A., Doores, K.J., Lee, J.H., Blattner, C., Ramirez, A., Derking, R., van Gils, M.J., Liang, C.H., et al. (2014). Broadly neutralizing HIV antibodies define a glycan-dependent epitope on the prefusion conformation of gp41 on cleaved envelope trimers. Immunity 40, 657-668. [DOI] [PMC free article] [PubMed]
- Gao F., Bailes E., Robertson D.L., Chen Y., Rodenburg C.M., Michael S.F., Cummins L.B., Arthur L.O., Peeters M., Shaw G.M. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature. 1999;397:436–441. doi: 10.1038/17130. [DOI] [PubMed] [Google Scholar]; Gao, F., Bailes, E., Robertson, D.L., Chen, Y., Rodenburg, C.M., Michael, S.F., Cummins, L.B., Arthur, L.O., Peeters, M., Shaw, G.M., et al. (1999). Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 397, 436-441. [DOI] [PubMed]
- Garces F., Sok D., Kong L., McBride R., Kim H.J., Saye-Francisco K.F., Julien J.P., Hua Y., Cupo A., Moore J.P. Structural evolution of glycan recognition by a family of potent HIV antibodies. Cell. 2014;159:69–79. doi: 10.1016/j.cell.2014.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]; Garces, F., Sok, D., Kong, L., McBride, R., Kim, H.J., Saye-Francisco, K.F., Julien, J.P., Hua, Y., Cupo, A., Moore, J.P., et al. (2014). Structural evolution of glycan recognition by a family of potent HIV antibodies. Cell 159, 69-79. [DOI] [PMC free article] [PubMed]
- Garces F., Lee J.H., de Val N., de la Pena A.T., Kong L., Puchades C., Hua Y., Stanfield R.L., Burton D.R., Moore J.P. Affinity Maturation of a Potent Family of HIV Antibodies Is Primarily Focused on Accommodating or Avoiding Glycans. Immunity. 2015;43:1053–1063. doi: 10.1016/j.immuni.2015.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]; Garces, F., Lee, J.H., de Val, N., de la Pena, A.T., Kong, L., Puchades, C., Hua, Y., Stanfield, R.L., Burton, D.R., Moore, J.P., et al. (2015). Affinity Maturation of a Potent Family of HIV Antibodies Is Primarily Focused on Accommodating or Avoiding Glycans. Immunity 43, 1053-1063. [DOI] [PMC free article] [PubMed]
- Georgiev I.S., Doria-Rose N.A., Zhou T., Kwon Y.D., Staupe R.P., Moquin S., Chuang G.Y., Louder M.K., Schmidt S.D., Altae-Tran H.R. Delineating antibody recognition in polyclonal sera from patterns of HIV-1 isolate neutralization. Science. 2013;340:751–756. doi: 10.1126/science.1233989. [DOI] [PubMed] [Google Scholar]; Georgiev, I.S., Doria-Rose, N.A., Zhou, T., Kwon, Y.D., Staupe, R.P., Moquin, S., Chuang, G.Y., Louder, M.K., Schmidt, S.D., Altae-Tran, H.R., et al. (2013). Delineating antibody recognition in polyclonal sera from patterns of HIV-1 isolate neutralization. Science 340, 751-756. [DOI] [PubMed]
- Gorman J., Soto C., Yang M.M., Davenport T.M., Guttman M., Bailer R.T., Chambers M., Chuang G.Y., DeKosky B.J., Doria-Rose N.A., NISC Comparative Sequencing Program Structures of HIV-1 Env V1V2 with broadly neutralizing antibodies reveal commonalities that enable vaccine design. Nat. Struct. Mol. Biol. 2016;23:81–90. doi: 10.1038/nsmb.3144. [DOI] [PMC free article] [PubMed] [Google Scholar]; Gorman, J., Soto, C., Yang, M.M., Davenport, T.M., Guttman, M., Bailer, R.T., Chambers, M., Chuang, G.Y., DeKosky, B.J., Doria-Rose, N.A., et al.; NISC Comparative Sequencing Program (2016). Structures of HIV-1 Env V1V2 with broadly neutralizing antibodies reveal commonalities that enable vaccine design. Nat. Struct. Mol. Biol. 23, 81-90. [DOI] [PMC free article] [PubMed]
- Gristick H.B., von Boehmer L., West A.P., Jr., Schamber M., Gazumyan A., Golijanin J., Seaman M.S., Fätkenheuer G., Klein F., Nussenzweig M.C., Bjorkman P.J. Natively glycosylated HIV-1 Env structure reveals new mode for antibody recognition of the CD4-binding site. Nat. Struct. Mol. Biol. 2016;23:906–915. doi: 10.1038/nsmb.3291. [DOI] [PMC free article] [PubMed] [Google Scholar]; Gristick, H.B., von Boehmer, L., West, A.P., Jr., Schamber, M., Gazumyan, A., Golijanin, J., Seaman, M.S., Fatkenheuer, G., Klein, F., Nussenzweig, M.C., and Bjorkman, P.J. (2016). Natively glycosylated HIV-1 Env structure reveals new mode for antibody recognition of the CD4-binding site. Nat. Struct. Mol. Biol. 23, 906-915. [DOI] [PMC free article] [PubMed]
- Harvey D.J., Baruah K., Scanlan C.N. Application of negative ion MS/MS to the identification of N-glycans released from carcinoembryonic antigen cell adhesion molecule 1 (CEACAM1) J. Mass Spectrom. 2009;44:50–60. doi: 10.1002/jms.1470. [DOI] [PubMed] [Google Scholar]; Harvey, D.J., Baruah, K., and Scanlan, C.N. (2009). Application of negative ion MS/MS to the identification of N-glycans released from carcinoembryonic antigen cell adhesion molecule 1 (CEACAM1). J. Mass Spectrom. 44, 50-60. [DOI] [PubMed]
- Havenar-Daughton C., Lee J.H., Crotty S. Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol. Rev. 2017;275:49–61. doi: 10.1111/imr.12512. [DOI] [PubMed] [Google Scholar]; Havenar-Daughton, C., Lee, J.H., and Crotty, S. (2017). Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol. Rev. 275, 49-61. [DOI] [PubMed]
- Haynes B.F., Mascola J.R. The quest for an antibody-based HIV vaccine. Immunol. Rev. 2017;275:5–10. doi: 10.1111/imr.12517. [DOI] [PMC free article] [PubMed] [Google Scholar]; Haynes, B.F., and Mascola, J.R. (2017). The quest for an antibody-based HIV vaccine. Immunol. Rev. 275, 5-10. [DOI] [PMC free article] [PubMed]
- Haynes B.F., Kelsoe G., Harrison S.C., Kepler T.B. B-cell-lineage immunogen design in vaccine development with HIV-1 as a case study. Nat. Biotechnol. 2012;30:423–433. doi: 10.1038/nbt.2197. [DOI] [PMC free article] [PubMed] [Google Scholar]; Haynes, B.F., Kelsoe, G., Harrison, S.C., and Kepler, T.B. (2012). B-cell-lineage immunogen design in vaccine development with HIV-1 as a case study. Nat. Biotechnol. 30, 423-433. [DOI] [PMC free article] [PubMed]
- Jardine J., Julien J.P., Menis S., Ota T., Kalyuzhniy O., McGuire A., Sok D., Huang P.S., MacPherson S., Jones M. Rational HIV immunogen design to target specific germline B cell receptors. Science. 2013;340:711–716. doi: 10.1126/science.1234150. [DOI] [PMC free article] [PubMed] [Google Scholar]; Jardine, J., Julien, J.P., Menis, S., Ota, T., Kalyuzhniy, O., McGuire, A., Sok, D., Huang, P.S., MacPherson, S., Jones, M., et al. (2013). Rational HIV immunogen design to target specific germline B cell receptors. Science 340, 711-716. [DOI] [PMC free article] [PubMed]
- Jardine J.G., Ota T., Sok D., Pauthner M., Kulp D.W., Kalyuzhniy O., Skog P.D., Thinnes T.C., Bhullar D., Briney B. HIV-1 VACCINES. Priming a broadly neutralizing antibody response to HIV-1 using a germline-targeting immunogen. Science. 2015;349:156–161. doi: 10.1126/science.aac5894. [DOI] [PMC free article] [PubMed] [Google Scholar]; Jardine, J.G., Ota, T., Sok, D., Pauthner, M., Kulp, D.W., Kalyuzhniy, O., Skog, P.D., Thinnes, T.C., Bhullar, D., Briney, B., et al. (2015). HIV-1 VACCINES. Priming a broadly neutralizing antibody response to HIV-1 using a germline-targeting immunogen. Science 349, 156-161. [DOI] [PMC free article] [PubMed]
- Julien J.P., Cupo A., Sok D., Stanfield R.L., Lyumkis D., Deller M.C., Klasse P.J., Burton D.R., Sanders R.W., Moore J.P. Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science. 2013;342:1477–1483. doi: 10.1126/science.1245625. [DOI] [PMC free article] [PubMed] [Google Scholar]; Julien, J.P., Cupo, A., Sok, D., Stanfield, R.L., Lyumkis, D., Deller, M.C., Klasse, P.J., Burton, D.R., Sanders, R.W., Moore, J.P., et al. (2013a). Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science 342, 1477-1483. [DOI] [PMC free article] [PubMed]
- Julien J.P., Lee J.H., Cupo A., Murin C.D., Derking R., Hoffenberg S., Caulfield M.J., King C.R., Marozsan A.J., Klasse P.J. Asymmetric recognition of the HIV-1 trimer by broadly neutralizing antibody PG9. Proc. Natl. Acad. Sci. USA. 2013;110:4351–4356. doi: 10.1073/pnas.1217537110. [DOI] [PMC free article] [PubMed] [Google Scholar]; Julien, J.P., Lee, J.H., Cupo, A., Murin, C.D., Derking, R., Hoffenberg, S., Caulfield, M.J., King, C.R., Marozsan, A.J., Klasse, P.J., et al. (2013b). Asymmetric recognition of the HIV-1 trimer by broadly neutralizing antibody PG9. Proc. Natl. Acad. Sci. USA 110, 4351-4356. [DOI] [PMC free article] [PubMed]
- Kepler T.B., Liao H.X., Alam S.M., Bhaskarabhatla R., Zhang R., Yandava C., Stewart S., Anasti K., Kelsoe G., Parks R. Immunoglobulin gene insertions and deletions in the affinity maturation of HIV-1 broadly reactive neutralizing antibodies. Cell Host Microbe. 2014;16:304–313. doi: 10.1016/j.chom.2014.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]; Kepler, T.B., Liao, H.X., Alam, S.M., Bhaskarabhatla, R., Zhang, R., Yandava, C., Stewart, S., Anasti, K., Kelsoe, G., Parks, R., et al. (2014). Immunoglobulin gene insertions and deletions in the affinity maturation of HIV-1 broadly reactive neutralizing antibodies. Cell Host Microbe 16, 304-313. [DOI] [PMC free article] [PubMed]
- Klein F., Diskin R., Scheid J.F., Gaebler C., Mouquet H., Georgiev I.S., Pancera M., Zhou T., Incesu R.B., Fu B.Z. Somatic mutations of the immunoglobulin framework are generally required for broad and potent HIV-1 neutralization. Cell. 2013;153:126–138. doi: 10.1016/j.cell.2013.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]; Klein, F., Diskin, R., Scheid, J.F., Gaebler, C., Mouquet, H., Georgiev, I.S., Pancera, M., Zhou, T., Incesu, R.B., Fu, B.Z., et al. (2013). Somatic mutations of the immunoglobulin framework are generally required for broad and potent HIV-1 neutralization. Cell 153, 126-138. [DOI] [PMC free article] [PubMed]
- Kong L., Lee J.H., Doores K.J., Murin C.D., Julien J.P., McBride R., Liu Y., Marozsan A., Cupo A., Klasse P.J. Supersite of immune vulnerability on the glycosylated face of HIV-1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 2013;20:796–803. doi: 10.1038/nsmb.2594. [DOI] [PMC free article] [PubMed] [Google Scholar]; Kong, L., Lee, J.H., Doores, K.J., Murin, C.D., Julien, J.P., McBride, R., Liu, Y., Marozsan, A., Cupo, A., Klasse, P.J., et al. (2013). Supersite of immune vulnerability on the glycosylated face of HIV-1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 20, 796-803. [DOI] [PMC free article] [PubMed]
- Korber B., Muldoon M., Theiler J., Gao F., Gupta R., Lapedes A., Hahn B.H., Wolinsky S., Bhattacharya T. Timing the ancestor of the HIV-1 pandemic strains. Science. 2000;288:1789–1796. doi: 10.1126/science.288.5472.1789. [DOI] [PubMed] [Google Scholar]; Korber, B., Muldoon, M., Theiler, J., Gao, F., Gupta, R., Lapedes, A., Hahn, B.H., Wolinsky, S., and Bhattacharya, T. (2000). Timing the ancestor of the HIV-1 pandemic strains. Science 288, 1789-1796. [DOI] [PubMed]
- Kwon Y.D., Pancera M., Acharya P., Georgiev I.S., Crooks E.T., Gorman J., Joyce M.G., Guttman M., Ma X., Narpala S. Crystal structure, conformational fixation and entry-related interactions of mature ligand-free HIV-1 Env. Nat. Struct. Mol. Biol. 2015;22:522–531. doi: 10.1038/nsmb.3051. [DOI] [PMC free article] [PubMed] [Google Scholar]; Kwon, Y.D., Pancera, M., Acharya, P., Georgiev, I.S., Crooks, E.T., Gorman, J., Joyce, M.G., Guttman, M., Ma, X., Narpala, S., et al. (2015). Crystal structure, conformational fixation and entry-related interactions of mature ligand-free HIV-1 Env. Nat. Struct. Mol. Biol. 22, 522-531. [DOI] [PMC free article] [PubMed]
- Landais E., Huang X., Havenar-Daughton C., Murrell B., Price M.A., Wickramasinghe L., Ramos A., Bian C.B., Simek M., Allen S. Broadly Neutralizing Antibody Responses in a Large Longitudinal Sub-Saharan HIV Primary Infection Cohort. PLoS Pathog. 2016;12:e1005369. doi: 10.1371/journal.ppat.1005369. [DOI] [PMC free article] [PubMed] [Google Scholar]; Landais, E., Huang, X., Havenar-Daughton, C., Murrell, B., Price, M.A., Wickramasinghe, L., Ramos, A., Bian, C.B., Simek, M., Allen, S., et al. (2016). Broadly Neutralizing Antibody Responses in a Large Longitudinal Sub-Saharan HIV Primary Infection Cohort. PLoS Pathog. 12, e1005369. [DOI] [PMC free article] [PubMed]
- Landais E., Murrell B., Briney B., Murrell S., Rantalainen K., Berndsen Z.T., Ramos A., Wickramasinghe L., Smith M.L., Eren K. HIV Envelope Glycoform Heterogeneity and Localized Diversity Govern the Initiation and Maturation of a V2 Apex Broadly Neutralizing Antibody Lineage. Immunity. 2017;47:990–1003.e9. doi: 10.1016/j.immuni.2017.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]; Landais, E., Murrell, B., Briney, B., Murrell, S., Rantalainen, K., Berndsen, Z.T., Ramos, A., Wickramasinghe, L., Smith, M.L., Eren, K., et al. (2017). HIV Envelope Glycoform Heterogeneity and Localized Diversity Govern the Initiation and Maturation of a V2 Apex Broadly Neutralizing Antibody Lineage. Immunity 47, 990-1003.e9. [DOI] [PMC free article] [PubMed]
- Lee E.C., Liang Q., Ali H., Bayliss L., Beasley A., Bloomfield-Gerdes T., Bonoli L., Brown R., Campbell J., Carpenter A. Complete humanization of the mouse immunoglobulin loci enables efficient therapeutic antibody discovery. Nat. Biotechnol. 2014;32:356–363. doi: 10.1038/nbt.2825. [DOI] [PubMed] [Google Scholar]; Lee, E.C., Liang, Q., Ali, H., Bayliss, L., Beasley, A., Bloomfield-Gerdes, T., Bonoli, L., Brown, R., Campbell, J., Carpenter, A., et al. (2014). Complete humanization of the mouse immunoglobulin loci enables efficient therapeutic antibody discovery. Nat. Biotechnol. 32, 356-363. [DOI] [PubMed]
- Lee J.H., de Val N., Lyumkis D., Ward A.B. Model Building and Refinement of a Natively Glycosylated HIV-1 Env Protein by High-Resolution Cryoelectron Microscopy. Structure. 2015;23:1943–1951. doi: 10.1016/j.str.2015.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]; Lee, J.H., de Val, N., Lyumkis, D., and Ward, A.B. (2015). Model Building and Refinement of a Natively Glycosylated HIV-1 Env Protein by High-Resolution Cryoelectron Microscopy. Structure 23, 1943-1951. [DOI] [PMC free article] [PubMed]
- Lee J.H., Ozorowski G., Ward A.B. Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science. 2016;351:1043–1048. doi: 10.1126/science.aad2450. [DOI] [PMC free article] [PubMed] [Google Scholar]; Lee, J.H., Ozorowski, G., and Ward, A.B. (2016). Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science 351, 1043-1048. [DOI] [PMC free article] [PubMed]
- Lee J.H., Andrabi R., Su C.Y., Yasmeen A., Julien J.P., Kong L., Wu N.C., McBride R., Sok D., Pauthner M. A Broadly Neutralizing Antibody Targets the Dynamic HIV Envelope Trimer Apex via a Long, Rigidified, and Anionic β-Hairpin Structure. Immunity. 2017;46:690–702. doi: 10.1016/j.immuni.2017.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]; Lee, J.H., Andrabi, R., Su, C.Y., Yasmeen, A., Julien, J.P., Kong, L., Wu, N.C., McBride, R., Sok, D., Pauthner, M., et al. (2017). A Broadly Neutralizing Antibody Targets the Dynamic HIV Envelope Trimer Apex via a Long, Rigidified, and Anionic β-Hairpin Structure. Immunity 46, 690-702. [DOI] [PMC free article] [PubMed]
- Lyumkis D., Julien J.P., de Val N., Cupo A., Potter C.S., Klasse P.J., Burton D.R., Sanders R.W., Moore J.P., Carragher B. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science. 2013;342:1484–1490. doi: 10.1126/science.1245627. [DOI] [PMC free article] [PubMed] [Google Scholar]; Lyumkis, D., Julien, J.P., de Val, N., Cupo, A., Potter, C.S., Klasse, P.J., Burton, D.R., Sanders, R.W., Moore, J.P., Carragher, B., et al. (2013). Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science 342, 1484-1490. [DOI] [PMC free article] [PubMed]
- McCoy L.E., Quigley A.F., Strokappe N.M., Bulmer-Thomas B., Seaman M.S., Mortier D., Rutten L., Chander N., Edwards C.J., Ketteler R. Potent and broad neutralization of HIV-1 by a llama antibody elicited by immunization. J. Exp. Med. 2012;209:1091–1103. doi: 10.1084/jem.20112655. [DOI] [PMC free article] [PubMed] [Google Scholar]; McCoy, L.E., Quigley, A.F., Strokappe, N.M., Bulmer-Thomas, B., Seaman, M.S., Mortier, D., Rutten, L., Chander, N., Edwards, C.J., Ketteler, R., et al. (2012). Potent and broad neutralization of HIV-1 by a llama antibody elicited by immunization. J. Exp. Med. 209, 1091-1103. [DOI] [PMC free article] [PubMed]
- McGuire A.T., Hoot S., Dreyer A.M., Lippy A., Stuart A., Cohen K.W., Jardine J., Menis S., Scheid J.F., West A.P. Engineering HIV envelope protein to activate germline B cell receptors of broadly neutralizing anti-CD4 binding site antibodies. J. Exp. Med. 2013;210:655–663. doi: 10.1084/jem.20122824. [DOI] [PMC free article] [PubMed] [Google Scholar]; McGuire, A.T., Hoot, S., Dreyer, A.M., Lippy, A., Stuart, A., Cohen, K.W., Jardine, J., Menis, S., Scheid, J.F., West, A.P., et al. (2013). Engineering HIV envelope protein to activate germline B cell receptors of broadly neutralizing anti-CD4 binding site antibodies. J. Exp. Med. 210, 655-663. [DOI] [PMC free article] [PubMed]
- McGuire A.T., Gray M.D., Dosenovic P., Gitlin A.D., Freund N.T., Petersen J., Correnti C., Johnsen W., Kegel R., Stuart A.B. Specifically modified Env immunogens activate B-cell precursors of broadly neutralizing HIV-1 antibodies in transgenic mice. Nat. Commun. 2016;7:10618. doi: 10.1038/ncomms10618. [DOI] [PMC free article] [PubMed] [Google Scholar]; McGuire, A.T., Gray, M.D., Dosenovic, P., Gitlin, A.D., Freund, N.T., Petersen, J., Correnti, C., Johnsen, W., Kegel, R., Stuart, A.B., et al. (2016). Specifically modified Env immunogens activate B-cell precursors of broadly neutralizing HIV-1 antibodies in transgenic mice. Nat. Commun. 7, 10618. [DOI] [PMC free article] [PubMed]
- McLellan J.S., Pancera M., Carrico C., Gorman J., Julien J.P., Khayat R., Louder R., Pejchal R., Sastry M., Dai K. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature. 2011;480:336–343. doi: 10.1038/nature10696. [DOI] [PMC free article] [PubMed] [Google Scholar]; McLellan, J.S., Pancera, M., Carrico, C., Gorman, J., Julien, J.P., Khayat, R., Louder, R., Pejchal, R., Sastry, M., Dai, K., et al. (2011). Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 480, 336-343. [DOI] [PMC free article] [PubMed]
- McLellan J.S., Chen M., Leung S., Graepel K.W., Du X., Yang Y., Zhou T., Baxa U., Yasuda E., Beaumont T. Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science. 2013;340:1113–1117. doi: 10.1126/science.1234914. [DOI] [PMC free article] [PubMed] [Google Scholar]; McLellan, J.S., Chen, M., Leung, S., Graepel, K.W., Du, X., Yang, Y., Zhou, T., Baxa, U., Yasuda, E., Beaumont, T., et al. (2013). Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science 340, 1113-1117. [DOI] [PMC free article] [PubMed]
- Montefiori D.C. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays. Curr. Protoc. Immunol. 2005;Chapter 12 doi: 10.1002/0471142735.im1211s64. Unit 12.11. [DOI] [PubMed] [Google Scholar]; Montefiori, D.C. (2005). Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays. Curr. Protoc. Immunol. Chapter 12, Unit 12.11. [DOI] [PubMed]
- Moore P.L., Gray E.S., Sheward D., Madiga M., Ranchobe N., Lai Z., Honnen W.J., Nonyane M., Tumba N., Hermanus T., CAPRISA 002 Study Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol. 2011;85:3128–3141. doi: 10.1128/JVI.02658-10. [DOI] [PMC free article] [PubMed] [Google Scholar]; Moore, P.L., Gray, E.S., Sheward, D., Madiga, M., Ranchobe, N., Lai, Z., Honnen, W.J., Nonyane, M., Tumba, N., Hermanus, T., et al.; CAPRISA 002 Study (2011). Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol. 85, 3128-3141. [DOI] [PMC free article] [PubMed]
- Moore P.L., Gorman J., Doria-Rose N.A., Morris L. Ontogeny-based immunogens for the induction of V2-directed HIV broadly neutralizing antibodies. Immunol. Rev. 2017;275:217–229. doi: 10.1111/imr.12501. [DOI] [PMC free article] [PubMed] [Google Scholar]; Moore, P.L., Gorman, J., Doria-Rose, N.A., and Morris, L. (2017). Ontogeny-based immunogens for the induction of V2-directed HIV broadly neutralizing antibodies. Immunol. Rev. 275, 217-229. [DOI] [PMC free article] [PubMed]
- Morgand M., Bouvin-Pley M., Plantier J.C., Moreau A., Alessandri E., Simon F., Pace C.S., Pancera M., Ho D.D., Poignard P. V1/V2 Neutralizing Epitope is Conserved in Divergent Non-M Groups of HIV-1. J. Acquir. Immune Defic. Syndr. 2016;71:237–245. doi: 10.1097/QAI.0000000000000854. [DOI] [PMC free article] [PubMed] [Google Scholar]; Morgand, M., Bouvin-Pley, M., Plantier, J.C., Moreau, A., Alessandri, E., Simon, F., Pace, C.S., Pancera, M., Ho, D.D., Poignard, P., et al. (2016). V1/V2 Neutralizing Epitope is Conserved in Divergent Non-M Groups of HIV-1. J. Acquir. Immune Defic. Syndr. 71, 237-245. [DOI] [PMC free article] [PubMed]
- Ogura T., Iwasaki K., Sato C. Topology representing network enables highly accurate classification of protein images taken by cryo electron-microscope without masking. J. Struct. Biol. 2003;143:185–200. doi: 10.1016/j.jsb.2003.08.005. [DOI] [PubMed] [Google Scholar]; Ogura, T., Iwasaki, K., and Sato, C. (2003). Topology representing network enables highly accurate classification of protein images taken by cryo electron-microscope without masking. J. Struct. Biol. 143, 185-200. [DOI] [PubMed]
- Osborn M.J., Ma B., Avis S., Binnie A., Dilley J., Yang X., Lindquist K., Ménoret S., Iscache A.L., Ouisse L.H. High-affinity IgG antibodies develop naturally in Ig-knockout rats carrying germline human IgH/Igκ/Igλ loci bearing the rat CH region. J. Immunol. 2013;190:1481–1490. doi: 10.4049/jimmunol.1203041. [DOI] [PMC free article] [PubMed] [Google Scholar]; Osborn, M.J., Ma, B., Avis, S., Binnie, A., Dilley, J., Yang, X., Lindquist, K., Menoret, S., Iscache, A.L., Ouisse, L.H., et al. (2013). High-affinity IgG antibodies develop naturally in Ig-knockout rats carrying germline human IgH/Igκ/Igλ loci bearing the rat CH region. J. Immunol. 190, 1481-1490. [DOI] [PMC free article] [PubMed]
- Ozorowski G., Pallesen J., de Val N., Lyumkis D., Cottrell C.A., Torres J.L., Copps J., Stanfield R.L., Cupo A., Pugach P. Open and closed structures reveal allostery and pliability in the HIV-1 envelope spike. Nature. 2017;547:360–363. doi: 10.1038/nature23010. [DOI] [PMC free article] [PubMed] [Google Scholar]; Ozorowski, G., Pallesen, J., de Val, N., Lyumkis, D., Cottrell, C.A., Torres, J.L., Copps, J., Stanfield, R.L., Cupo, A., Pugach, P., et al. (2017). Open and closed structures reveal allostery and pliability in the HIV-1 envelope spike. Nature 547, 360-363. [DOI] [PMC free article] [PubMed]
- Pallesen J., Murin C.D., de Val N., Cottrell C.A., Hastie K.M., Turner H.L., Fusco M.L., Flyak A.I., Zeitlin L., Crowe J.E., Jr. Structures of Ebola virus GP and sGP in complex with therapeutic antibodies. Nat. Microbiol. 2016;1:16128. doi: 10.1038/nmicrobiol.2016.128. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pallesen, J., Murin, C.D., de Val, N., Cottrell, C.A., Hastie, K.M., Turner, H.L., Fusco, M.L., Flyak, A.I., Zeitlin, L., Crowe, J.E., Jr., et al. (2016). Structures of Ebola virus GP and sGP in complex with therapeutic antibodies. Nat. Microbiol. 1, 16128. [DOI] [PMC free article] [PubMed]
- Pallesen J., Wang N., Corbett K.S., Wrapp D., Kirchdoerfer R.N., Turner H.L., Cottrell C.A., Becker M.M., Wang L., Shi W. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA. 2017;114:E7348–E7357. doi: 10.1073/pnas.1707304114. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pallesen, J., Wang, N., Corbett, K.S., Wrapp, D., Kirchdoerfer, R.N., Turner, H.L., Cottrell, C.A., Becker, M.M., Wang, L., Shi, W., et al. (2017). Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA 114, E7348-E7357. [DOI] [PMC free article] [PubMed]
- Pancera M., Shahzad-Ul-Hussan S., Doria-Rose N.A., McLellan J.S., Bailer R.T., Dai K., Loesgen S., Louder M.K., Staupe R.P., Yang Y. Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat. Struct. Mol. Biol. 2013;20:804–813. doi: 10.1038/nsmb.2600. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pancera, M., Shahzad-Ul-Hussan, S., Doria-Rose, N.A., McLellan, J.S., Bailer, R.T., Dai, K., Loesgen, S., Louder, M.K., Staupe, R.P., Yang, Y., et al. (2013). Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat. Struct. Mol. Biol. 20, 804-813. [DOI] [PMC free article] [PubMed]
- Pancera M., Zhou T., Druz A., Georgiev I.S., Soto C., Gorman J., Huang J., Acharya P., Chuang G.Y., Ofek G. Structure and immune recognition of trimeric pre-fusion HIV-1 Env. Nature. 2014;514:455–461. doi: 10.1038/nature13808. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pancera, M., Zhou, T., Druz, A., Georgiev, I.S., Soto, C., Gorman, J., Huang, J., Acharya, P., Chuang, G.Y., Ofek, G., et al. (2014). Structure and immune recognition of trimeric pre-fusion HIV-1 Env. Nature 514, 455-461. [DOI] [PMC free article] [PubMed]
- Panico M., Bouché L., Binet D., O’Connor M.J., Rahman D., Pang P.C., Canis K., North S.J., Desrosiers R.C., Chertova E. Mapping the complete glycoproteome of virion-derived HIV-1 gp120 provides insights into broadly neutralizing antibody binding. Sci. Rep. 2016;6:32956. doi: 10.1038/srep32956. [DOI] [PMC free article] [PubMed] [Google Scholar]; Panico, M., Bouche, L., Binet, D., O’Connor, M.J., Rahman, D., Pang, P.C., Canis, K., North, S.J., Desrosiers, R.C., Chertova, E., et al. (2016). Mapping the complete glycoproteome of virion-derived HIV-1 gp120 provides insights into broadly neutralizing antibody binding. Sci. Rep. 6, 32956. [DOI] [PMC free article] [PubMed]
- Pejchal R., Doores K.J., Walker L.M., Khayat R., Huang P.S., Wang S.K., Stanfield R.L., Julien J.P., Ramos A., Crispin M. A potent and broad neutralizing antibody recognizes and penetrates the HIV glycan shield. Science. 2011;334:1097–1103. doi: 10.1126/science.1213256. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pejchal, R., Doores, K.J., Walker, L.M., Khayat, R., Huang, P.S., Wang, S.K., Stanfield, R.L., Julien, J.P., Ramos, A., Crispin, M., et al. (2011). A potent and broad neutralizing antibody recognizes and penetrates the HIV glycan shield. Science 334, 1097-1103. [DOI] [PMC free article] [PubMed]
- Pettersen E.F., Goddard T.D., Huang C.C., Couch G.S., Greenblatt D.M., Meng E.C., Ferrin T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004;25:1605–1612. doi: 10.1002/jcc.20084. [DOI] [PubMed] [Google Scholar]; Pettersen, E.F., Goddard, T.D., Huang, C.C., Couch, G.S., Greenblatt, D.M., Meng, E.C., and Ferrin, T.E. (2004). UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605-1612. [DOI] [PubMed]
- Pritchard L.K., Harvey D.J., Bonomelli C., Crispin M., Doores K.J. Cell- and Protein-Directed Glycosylation of Native Cleaved HIV-1 Envelope. J. Virol. 2015;89:8932–8944. doi: 10.1128/JVI.01190-15. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pritchard, L.K., Harvey, D.J., Bonomelli, C., Crispin, M., and Doores, K.J. (2015). Cell- and Protein-Directed Glycosylation of Native Cleaved HIV-1 Envelope. J. Virol. 89, 8932-8944. [DOI] [PMC free article] [PubMed]
- Pugach P., Ozorowski G., Cupo A., Ringe R., Yasmeen A., de Val N., Derking R., Kim H.J., Korzun J., Golabek M. A native-like SOSIP.664 trimer based on an HIV-1 subtype B env gene. J. Virol. 2015;89:3380–3395. doi: 10.1128/JVI.03473-14. [DOI] [PMC free article] [PubMed] [Google Scholar]; Pugach, P., Ozorowski, G., Cupo, A., Ringe, R., Yasmeen, A., de Val, N., Derking, R., Kim, H.J., Korzun, J., Golabek, M., et al. (2015). A native-like SOSIP.664 trimer based on an HIV-1 subtype B env gene. J. Virol. 89, 3380-3395. [DOI] [PMC free article] [PubMed]
- Riedel S. Edward Jenner and the history of smallpox and vaccination. Proc. Bayl. Univ. Med. Cent. 2005;18:21–25. doi: 10.1080/08998280.2005.11928028. [DOI] [PMC free article] [PubMed] [Google Scholar]; Riedel, S. (2005). Edward Jenner and the history of smallpox and vaccination. Proc. Bayl. Univ. Med. Cent. 18, 21-25. [DOI] [PMC free article] [PubMed]
- Sanders R.W., Derking R., Cupo A., Julien J.P., Yasmeen A., de Val N., Kim H.J., Blattner C., de la Peña A.T., Korzun J. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 2013;9:e1003618. doi: 10.1371/journal.ppat.1003618. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sanders, R.W., Derking, R., Cupo, A., Julien, J.P., Yasmeen, A., de Val, N., Kim, H.J., Blattner, C., de la Peña, A.T., Korzun, J., et al. (2013). A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 9, e1003618. [DOI] [PMC free article] [PubMed]
- Sanders R.W., van Gils M.J., Derking R., Sok D., Ketas T.J., Burger J.A., Ozorowski G., Cupo A., Simonich C., Goo L. HIV-1 VACCINES. HIV-1 neutralizing antibodies induced by native-like envelope trimers. Science. 2015;349:aac4223. doi: 10.1126/science.aac4223. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sanders, R.W., van Gils, M.J., Derking, R., Sok, D., Ketas, T.J., Burger, J.A., Ozorowski, G., Cupo, A., Simonich, C., Goo, L., et al. (2015). HIV-1 VACCINES. HIV-1 neutralizing antibodies induced by native-like envelope trimers. Science 349, aac4223. [DOI] [PMC free article] [PubMed]
- Saunders K.O., Verkoczy L.K., Jiang C., Zhang J., Parks R., Chen H., Housman M., Bouton-Verville H., Shen X., Trama A.M. Vaccine Induction of Heterologous Tier 2 HIV-1 Neutralizing Antibodies in Animal Models. Cell Rep. 2017;21:3681–3690. doi: 10.1016/j.celrep.2017.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]; Saunders, K.O., Verkoczy, L.K., Jiang, C., Zhang, J., Parks, R., Chen, H., Housman, M., Bouton-Verville, H., Shen, X., Trama, A.M., et al. (2017). Vaccine Induction of Heterologous Tier 2 HIV-1 Neutralizing Antibodies in Animal Models. Cell Rep. 21, 3681-3690. [DOI] [PMC free article] [PubMed]
- Scheres S.H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 2012;180:519–530. doi: 10.1016/j.jsb.2012.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]; Scheres, S.H. (2012). RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 180, 519-530. [DOI] [PMC free article] [PubMed]
- Seaman M.S., Janes H., Hawkins N., Grandpre L.E., Devoy C., Giri A., Coffey R.T., Harris L., Wood B., Daniels M.G. Tiered categorization of a diverse panel of HIV-1 Env pseudoviruses for assessment of neutralizing antibodies. J. Virol. 2010;84:1439–1452. doi: 10.1128/JVI.02108-09. [DOI] [PMC free article] [PubMed] [Google Scholar]; Seaman, M.S., Janes, H., Hawkins, N., Grandpre, L.E., Devoy, C., Giri, A., Coffey, R.T., Harris, L., Wood, B., Daniels, M.G., et al. (2010). Tiered categorization of a diverse panel of HIV-1 Env pseudoviruses for assessment of neutralizing antibodies. J. Virol. 84, 1439-1452. [DOI] [PMC free article] [PubMed]
- Sharma S.K., de Val N., Bale S., Guenaga J., Tran K., Feng Y., Dubrovskaya V., Ward A.B., Wyatt R.T. Cleavage-independent HIV-1 Env trimers engineered as soluble native spike mimetics for vaccine design. Cell Rep. 2015;11:539–550. doi: 10.1016/j.celrep.2015.03.047. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sharma, S.K., de Val, N., Bale, S., Guenaga, J., Tran, K., Feng, Y., Dubrovskaya, V., Ward, A.B., and Wyatt, R.T. (2015). Cleavage-independent HIV-1 Env trimers engineered as soluble native spike mimetics for vaccine design. Cell Rep. 11, 539-550. [DOI] [PMC free article] [PubMed]
- Sharp P.M., Hahn B.H. Origins of HIV and the AIDS pandemic. Cold Spring Harb. Perspect. Med. 2011;1:a006841. doi: 10.1101/cshperspect.a006841. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sharp, P.M., and Hahn, B.H. (2011). Origins of HIV and the AIDS pandemic. Cold Spring Harb. Perspect. Med. 1, a006841. [DOI] [PMC free article] [PubMed]
- Sok D., Doores K.J., Briney B., Le K.M., Saye-Francisco K.L., Ramos A., Kulp D.W., Julien J.P., Menis S., Wickramasinghe L. Promiscuous glycan site recognition by antibodies to the high-mannose patch of gp120 broadens neutralization of HIV. Sci. Transl. Med. 2014;6:236ra63. doi: 10.1126/scitranslmed.3008104. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sok, D., Doores, K.J., Briney, B., Le, K.M., Saye-Francisco, K.L., Ramos, A., Kulp, D.W., Julien, J.P., Menis, S., Wickramasinghe, L., et al. (2014a). Promiscuous glycan site recognition by antibodies to the high-mannose patch of gp120 broadens neutralization of HIV. Sci. Transl. Med. 6, 236ra63. [DOI] [PMC free article] [PubMed]
- Sok D., van Gils M.J., Pauthner M., Julien J.P., Saye-Francisco K.L., Hsueh J., Briney B., Lee J.H., Le K.M., Lee P.S. Recombinant HIV envelope trimer selects for quaternary-dependent antibodies targeting the trimer apex. Proc. Natl. Acad. Sci. USA. 2014;111:17624–17629. doi: 10.1073/pnas.1415789111. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sok, D., van Gils, M.J., Pauthner, M., Julien, J.P., Saye-Francisco, K.L., Hsueh, J., Briney, B., Lee, J.H., Le, K.M., Lee, P.S., et al. (2014b). Recombinant HIV envelope trimer selects for quaternary-dependent antibodies targeting the trimer apex. Proc. Natl. Acad. Sci. USA 111, 17624-17629. [DOI] [PMC free article] [PubMed]
- Sok D., Briney B., Jardine J.G., Kulp D.W., Menis S., Pauthner M., Wood A., Lee E.C., Le K.M., Jones M. Priming HIV-1 broadly neutralizing antibody precursors in human Ig loci transgenic mice. Science. 2016;353:1557–1560. doi: 10.1126/science.aah3945. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sok, D., Briney, B., Jardine, J.G., Kulp, D.W., Menis, S., Pauthner, M., Wood, A., Lee, E.C., Le, K.M., Jones, M., et al. (2016a). Priming HIV-1 broadly neutralizing antibody precursors in human Ig loci transgenic mice. Science 353, 1557-1560. [DOI] [PMC free article] [PubMed]
- Sok D., Pauthner M., Briney B., Lee J.H., Saye-Francisco K.L., Hsueh J., Ramos A., Le K.M., Jones M., Jardine J.G. A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity. 2016;45:31–45. doi: 10.1016/j.immuni.2016.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sok, D., Pauthner, M., Briney, B., Lee, J.H., Saye-Francisco, K.L., Hsueh, J., Ramos, A., Le, K.M., Jones, M., Jardine, J.G., et al. (2016b). A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity 45, 31-45. [DOI] [PMC free article] [PubMed]
- Sok D., Le K.M., Vadnais M., Saye-Francisco K.L., Jardine J.G., Torres J.L., Berndsen Z.T., Kong L., Stanfield R., Ruiz J. Rapid elicitation of broadly neutralizing antibodies to HIV by immunization in cows. Nature. 2017;548:108–111. doi: 10.1038/nature23301. [DOI] [PMC free article] [PubMed] [Google Scholar]; Sok, D., Le, K.M., Vadnais, M., Saye-Francisco, K.L., Jardine, J.G., Torres, J.L., Berndsen, Z.T., Kong, L., Stanfield, R., Ruiz, J., et al. (2017). Rapid elicitation of broadly neutralizing antibodies to HIV by immunization in cows. Nature 548, 108-111. [DOI] [PMC free article] [PubMed]
- Steichen J.M., Kulp D.W., Tokatlian T., Escolano A., Dosenovic P., Stanfield R.L., McCoy L.E., Ozorowski G., Hu X., Kalyuzhniy O. HIV Vaccine Design to Target Germline Precursors of Glycan-Dependent Broadly Neutralizing Antibodies. Immunity. 2016;45:483–496. doi: 10.1016/j.immuni.2016.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]; Steichen, J.M., Kulp, D.W., Tokatlian, T., Escolano, A., Dosenovic, P., Stanfield, R.L., McCoy, L.E., Ozorowski, G., Hu, X., Kalyuzhniy, O., et al. (2016). HIV Vaccine Design to Target Germline Precursors of Glycan-Dependent Broadly Neutralizing Antibodies. Immunity 45, 483-496. [DOI] [PMC free article] [PubMed]
- Stevens J., Corper A.L., Basler C.F., Taubenberger J.K., Palese P., Wilson I.A. Structure of the uncleaved human H1 hemagglutinin from the extinct 1918 influenza virus. Science. 2004;303:1866–1870. doi: 10.1126/science.1093373. [DOI] [PubMed] [Google Scholar]; Stevens, J., Corper, A.L., Basler, C.F., Taubenberger, J.K., Palese, P., and Wilson, I.A. (2004). Structure of the uncleaved human H1 hemagglutinin from the extinct 1918 influenza virus. Science 303, 1866-1870. [DOI] [PubMed]
- Tas J.M., Mesin L., Pasqual G., Targ S., Jacobsen J.T., Mano Y.M., Chen C.S., Weill J.C., Reynaud C.A., Browne E.P. Visualizing antibody affinity maturation in germinal centers. Science. 2016;351:1048–1054. doi: 10.1126/science.aad3439. [DOI] [PMC free article] [PubMed] [Google Scholar]; Tas, J.M., Mesin, L., Pasqual, G., Targ, S., Jacobsen, J.T., Mano, Y.M., Chen, C.S., Weill, J.C., Reynaud, C.A., Browne, E.P., et al. (2016). Visualizing antibody affinity maturation in germinal centers. Science 351, 1048-1054. [DOI] [PMC free article] [PubMed]
- Tian M., Cheng C., Chen X., Duan H., Cheng H.L., Dao M., Sheng Z., Kimble M., Wang L., Lin S. Induction of HIV Neutralizing Antibody Lineages in Mice with Diverse Precursor Repertoires. Cell. 2016;166:1471–1484.e18. doi: 10.1016/j.cell.2016.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]; Tian, M., Cheng, C., Chen, X., Duan, H., Cheng, H.L., Dao, M., Sheng, Z., Kimble, M., Wang, L., Lin, S., et al. (2016). Induction of HIV Neutralizing Antibody Lineages in Mice with Diverse Precursor Repertoires. Cell 166, 1471-1484.e18. [DOI] [PMC free article] [PubMed]
- Tiller T., Meffre E., Yurasov S., Tsuiji M., Nussenzweig M.C., Wardemann H. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. J. Immunol. Methods. 2008;329:112–124. doi: 10.1016/j.jim.2007.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]; Tiller, T., Meffre, E., Yurasov, S., Tsuiji, M., Nussenzweig, M.C., and Wardemann, H. (2008). Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. J. Immunol. Methods 329, 112-124. [DOI] [PMC free article] [PubMed]
- Verkoczy L., Diaz M., Holl T.M., Ouyang Y.B., Bouton-Verville H., Alam S.M., Liao H.X., Kelsoe G., Haynes B.F. Autoreactivity in an HIV-1 broadly reactive neutralizing antibody variable region heavy chain induces immunologic tolerance. Proc. Natl. Acad. Sci. USA. 2010;107:181–186. doi: 10.1073/pnas.0912914107. [DOI] [PMC free article] [PubMed] [Google Scholar]; Verkoczy, L., Diaz, M., Holl, T.M., Ouyang, Y.B., Bouton-Verville, H., Alam, S.M., Liao, H.X., Kelsoe, G., and Haynes, B.F. (2010). Autoreactivity in an HIV-1 broadly reactive neutralizing antibody variable region heavy chain induces immunologic tolerance. Proc. Natl. Acad. Sci. USA 107, 181-186. [DOI] [PMC free article] [PubMed]
- Verkoczy L., Chen Y., Bouton-Verville H., Zhang J., Diaz M., Hutchinson J., Ouyang Y.B., Alam S.M., Holl T.M., Hwang K.K. Rescue of HIV-1 broad neutralizing antibody-expressing B cells in 2F5 VH x VL knockin mice reveals multiple tolerance controls. J. Immunol. 2011;187:3785–3797. doi: 10.4049/jimmunol.1101633. [DOI] [PMC free article] [PubMed] [Google Scholar]; Verkoczy, L., Chen, Y., Bouton-Verville, H., Zhang, J., Diaz, M., Hutchinson, J., Ouyang, Y.B., Alam, S.M., Holl, T.M., Hwang, K.K., et al. (2011). Rescue of HIV-1 broad neutralizing antibody-expressing B cells in 2F5 VH x VL knockin mice reveals multiple tolerance controls. J. Immunol. 187, 3785-3797. [DOI] [PMC free article] [PubMed]
- Voss J.E., Andrabi R., McCoy L.E., de Val N., Fuller R.P., Messmer T., Su C.Y., Sok D., Khan S.N., Garces F. Elicitation of Neutralizing Antibodies Targeting the V2 Apex of the HIV Envelope Trimer in a Wild-Type Animal Model. Cell Rep. 2017;21:222–235. doi: 10.1016/j.celrep.2017.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]; Voss, J.E., Andrabi, R., McCoy, L.E., de Val, N., Fuller, R.P., Messmer, T., Su, C.Y., Sok, D., Khan, S.N., Garces, F., et al. (2017). Elicitation of Neutralizing Antibodies Targeting the V2 Apex of the HIV Envelope Trimer in a Wild-Type Animal Model. Cell Rep. 21, 222-235. [DOI] [PMC free article] [PubMed]
- Voss N.R., Yoshioka C.K., Radermacher M., Potter C.S., Carragher B. DoG Picker and TiltPicker: software tools to facilitate particle selection in single particle electron microscopy. J. Struct. Biol. 2009;166:205–213. doi: 10.1016/j.jsb.2009.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]; Voss, N.R., Yoshioka, C.K., Radermacher, M., Potter, C.S., and Carragher, B. (2009). DoG Picker and TiltPicker: software tools to facilitate particle selection in single particle electron microscopy. J. Struct. Biol. 166, 205-213. [DOI] [PMC free article] [PubMed]
- Walker L.M., Phogat S.K., Chan-Hui P.Y., Wagner D., Phung P., Goss J.L., Wrin T., Simek M.D., Fling S., Mitcham J.L., Protocol G Principal Investigators Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science. 2009;326:285–289. doi: 10.1126/science.1178746. [DOI] [PMC free article] [PubMed] [Google Scholar]; Walker, L.M., Phogat, S.K., Chan-Hui, P.Y., Wagner, D., Phung, P., Goss, J.L., Wrin, T., Simek, M.D., Fling, S., Mitcham, J.L., et al.; Protocol G Principal Investigators (2009). Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 326, 285-289. [DOI] [PMC free article] [PubMed]
- Walker L.M., Huber M., Doores K.J., Falkowska E., Pejchal R., Julien J.P., Wang S.K., Ramos A., Chan-Hui P.Y., Moyle M., Protocol G Principal Investigators Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature. 2011;477:466–470. doi: 10.1038/nature10373. [DOI] [PMC free article] [PubMed] [Google Scholar]; Walker, L.M., Huber, M., Doores, K.J., Falkowska, E., Pejchal, R., Julien, J.P., Wang, S.K., Ramos, A., Chan-Hui, P.Y., Moyle, M., et al.; Protocol G Principal Investigators (2011). Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 477, 466-470. [DOI] [PMC free article] [PubMed]
- Wang S., Mata-Fink J., Kriegsman B., Hanson M., Irvine D.J., Eisen H.N., Burton D.R., Wittrup K.D., Kardar M., Chakraborty A.K. Manipulating the selection forces during affinity maturation to generate cross-reactive HIV antibodies. Cell. 2015;160:785–797. doi: 10.1016/j.cell.2015.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]; Wang, S., Mata-Fink, J., Kriegsman, B., Hanson, M., Irvine, D.J., Eisen, H.N., Burton, D.R., Wittrup, K.D., Kardar, M., and Chakraborty, A.K. (2015). Manipulating the selection forces during affinity maturation to generate cross-reactive HIV antibodies. Cell 160, 785-797. [DOI] [PMC free article] [PubMed]
- Webb B., Sali A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinformatics. 2016;Chapter 5 doi: 10.1002/0471250953.bi0506s15. Unit-5.6. [DOI] [PMC free article] [PubMed] [Google Scholar]; Webb, B., and Sali, A. (2016). Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinformatics Chapter 5, Unit-5.6. [DOI] [PMC free article] [PubMed]
- Wibmer C.K., Bhiman J.N., Gray E.S., Tumba N., Abdool Karim S.S., Williamson C., Morris L., Moore P.L. Viral escape from HIV-1 neutralizing antibodies drives increased plasma neutralization breadth through sequential recognition of multiple epitopes and immunotypes. PLoS Pathog. 2013;9:e1003738. doi: 10.1371/journal.ppat.1003738. [DOI] [PMC free article] [PubMed] [Google Scholar]; Wibmer, C.K., Bhiman, J.N., Gray, E.S., Tumba, N., Abdool Karim, S.S., Williamson, C., Morris, L., and Moore, P.L. (2013). Viral escape from HIV-1 neutralizing antibodies drives increased plasma neutralization breadth through sequential recognition of multiple epitopes and immunotypes. PLoS Pathog. 9, e1003738. [DOI] [PMC free article] [PubMed]
- Williams W.B., Zhang J., Jiang C., Nicely N.I., Fera D., Luo K., Moody M.A., Liao H.X., Alam S.M., Kepler T.B. Initiation of HIV neutralizing B cell lineages with sequential envelope immunizations. Nat. Commun. 2017;8:1732. doi: 10.1038/s41467-017-01336-3. [DOI] [PMC free article] [PubMed] [Google Scholar]; Williams, W.B., Zhang, J., Jiang, C., Nicely, N.I., Fera, D., Luo, K., Moody, M.A., Liao, H.X., Alam, S.M., Kepler, T.B., et al. (2017). Initiation of HIV neutralizing B cell lineages with sequential envelope immunizations. Nat. Commun. 8, 1732. [DOI] [PMC free article] [PubMed]
- Worobey M., Gemmel M., Teuwen D.E., Haselkorn T., Kunstman K., Bunce M., Muyembe J.J., Kabongo J.M., Kalengayi R.M., Van Marck E. Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature. 2008;455:661–664. doi: 10.1038/nature07390. [DOI] [PMC free article] [PubMed] [Google Scholar]; Worobey, M., Gemmel, M., Teuwen, D.E., Haselkorn, T., Kunstman, K., Bunce, M., Muyembe, J.J., Kabongo, J.M., Kalengayi, R.M., Van Marck, E., et al. (2008). Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature 455, 661-664. [DOI] [PMC free article] [PubMed]
- Wu X., Yang Z.Y., Li Y., Hogerkorp C.M., Schief W.R., Seaman M.S., Zhou T., Schmidt S.D., Wu L., Xu L. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science. 2010;329:856–861. doi: 10.1126/science.1187659. [DOI] [PMC free article] [PubMed] [Google Scholar]; Wu, X., Yang, Z.Y., Li, Y., Hogerkorp, C.M., Schief, W.R., Seaman, M.S., Zhou, T., Schmidt, S.D., Wu, L., Xu, L., et al. (2010). Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 329, 856-861. [DOI] [PMC free article] [PubMed]
- Xiao X., Chen W., Feng Y., Zhu Z., Prabakaran P., Wang Y., Zhang M.Y., Longo N.S., Dimitrov D.S. Germline-like predecessors of broadly neutralizing antibodies lack measurable binding to HIV-1 envelope glycoproteins: implications for evasion of immune responses and design of vaccine immunogens. Biochem. Biophys. Res. Commun. 2009;390:404–409. doi: 10.1016/j.bbrc.2009.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]; Xiao, X., Chen, W., Feng, Y., Zhu, Z., Prabakaran, P., Wang, Y., Zhang, M.Y., Longo, N.S., and Dimitrov, D.S. (2009). Germline-like predecessors of broadly neutralizing antibodies lack measurable binding to HIV-1 envelope glycoproteins: implications for evasion of immune responses and design of vaccine immunogens. Biochem. Biophys. Res. Commun. 390, 404-409. [DOI] [PMC free article] [PubMed]
- Xu K., Acharya P., Kong R., Cheng C., Chuang G.Y., Liu K., Louder M.K., O’Dell S., Rawi R., Sastry M. Epitope-based vaccine design yields fusion peptide-directed antibodies that neutralize diverse strains of HIV-1. Nat. Med. 2018;24:857–867. doi: 10.1038/s41591-018-0042-6. [DOI] [PMC free article] [PubMed] [Google Scholar]; Xu, K., Acharya, P., Kong, R., Cheng, C., Chuang, G.Y., Liu, K., Louder, M.K., O’Dell, S., Rawi, R., Sastry, M., et al. (2018). Epitope-based vaccine design yields fusion peptide-directed antibodies that neutralize diverse strains of HIV-1. Nat. Med. 24, 857-867. [DOI] [PMC free article] [PubMed]
- Zhang K. Gctf: Real-time CTF determination and correction. J. Struct. Biol. 2016;193:1–12. doi: 10.1016/j.jsb.2015.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]; Zhang, K. (2016). Gctf: Real-time CTF determination and correction. J. Struct. Biol. 193, 1-12. [DOI] [PMC free article] [PubMed]
- Zhang M., Gaschen B., Blay W., Foley B., Haigwood N., Kuiken C., Korber B. Tracking global patterns of N-linked glycosylation site variation in highly variable viral glycoproteins: HIV, SIV, and HCV envelopes and influenza hemagglutinin. Glycobiology. 2004;14:1229–1246. doi: 10.1093/glycob/cwh106. [DOI] [PubMed] [Google Scholar]; Zhang, M., Gaschen, B., Blay, W., Foley, B., Haigwood, N., Kuiken, C., and Korber, B. (2004). Tracking global patterns of N-linked glycosylation site variation in highly variable viral glycoproteins: HIV, SIV, and HCV envelopes and influenza hemagglutinin. Glycobiology 14, 1229-1246. [DOI] [PubMed]
- Zheng S.Q., Palovcak E., Armache J.P., Verba K.A., Cheng Y., Agard D.A. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods. 2017;14:331–332. doi: 10.1038/nmeth.4193. [DOI] [PMC free article] [PubMed] [Google Scholar]; Zheng, S.Q., Palovcak, E., Armache, J.P., Verba, K.A., Cheng, Y., and Agard, D.A. (2017). MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331-332. [DOI] [PMC free article] [PubMed]
- Zhou T., Georgiev I., Wu X., Yang Z.Y., Dai K., Finzi A., Kwon Y.D., Scheid J.F., Shi W., Xu L. Structural basis for broad and potent neutralization of HIV-1 by antibody VRC01. Science. 2010;329:811–817. doi: 10.1126/science.1192819. [DOI] [PMC free article] [PubMed] [Google Scholar]; Zhou, T., Georgiev, I., Wu, X., Yang, Z.Y., Dai, K., Finzi, A., Kwon, Y.D., Scheid, J.F., Shi, W., Xu, L., et al. (2010). Structural basis for broad and potent neutralization of HIV-1 by antibody VRC01. Science 329, 811-817. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Cryo-EM reconstructions have been deposited in the Electron Microscopy Data Bank (EMDB) under the accession numbers EMD-20074 and the PDB code for this deposition is 6OHY. The GenBank accession numbers for the heavy and light chain inferred germline versions of PG9 and CH01 broadly neutralizing antibodies reported in this paper are MK825341-MK825344.