

Sleep supports changes in the neuronal representation of emotional experiences to make those experiences less distressing when subsequently recalled. Wassing et al. show that individuals with insomnia disorder activate the anterior cingulate cortex during both new and relived emotional experiences. The findings suggest insufficient adaptation to emotional distress in people who are vulnerable to developing insomnia disorder.
Keywords: self-conscious emotion, insomnia disorder, sleep, ACC, emotional memory
Abstract
Studies suggest that sleep supports persistent changes in the neuronal representation of emotional experiences such that they are remembered better and less distressful when recalled than when they were first experienced. It is conceivable that sleep fragmentation by arousals, a key characteristic of insomnia disorder, could hamper the downregulation of distress. In this study, we sought further support for the idea that insomnia disorder may involve a lasting deficiency to downregulate emotional distress. We used functional MRI in insomnia disorder (n = 27) and normal sleepers (n = 30) to identify how brain activation differs between novel and relived self-conscious emotions. We evaluated whether brain activity elicited by reliving emotional memories from the distant past resembles the activity elicited by novel emotional experiences more in insomnia disorder than in normal sleepers. Limbic areas were activated during novel shameful experiences as compared to neutral experiences in both normal sleepers and insomnia disorder. In normal sleepers, reliving of shameful experiences from the past did not elicit a limbic response. In contrast, participants with insomnia disorder recruited overlapping parts of the limbic circuit, in particular the dorsal anterior cingulate cortex, during both new and relived shameful experiences. The differential activity patterns with new and old emotions in normal sleepers suggest that reactivation of the long-term memory trace does not recruit the limbic circuit. The overlap of activations in insomnia disorder is in line with the hypothesis that the disorder involves a deficiency to dissociate the limbic circuit from the emotional memory trace. Moreover, the findings provide further support for a role of the anterior cingulate cortex in insomnia.
Introduction
With a prevalence of about 7% in the general European population, insomnia disorder is the second most common mental disorder (Wittchen et al., 2011). Genome-wide association studies have commenced to reveal biological pathways involved in insomnia (Hammerschlag et al., 2017; Jansen et al., 2019). Surprisingly, the genetic risk of insomnia disorder is only modestly correlated with the genetic risk of other unfavourable sleep traits. Rather, both genetically and phenotypically, insomnia is most markedly associated with anxiety disorders (Hammerschlag et al., 2017). Moreover, functional annotation analysis found enriched expression of insomnia risk genes in the limbic circuitry of the brain, notably in the anterior cingulate cortex (ACC). The finding that insomnia-risk genes are expressed in limbic circuits again points to a possible key involvement of disturbed emotion processing in insomnia disorder (Jansen et al., 2019). While the name insomnia highlights the sleep issues, the diagnosis requires related daytime complaints as well, and a vast number of objective and subjective measures indicate round-the-clock disturbances (Bonnet and Arand, 2010; Riemann et al., 2010). Hyperarousal resembles the state that healthy volunteers show during short-lived acute anxiety or other emotional distress. The upcoming notion that emotional memory processing deficits are an important aspect of anxiety disorders (Kindt, 2018), may apply equally well to insomnia (Wassing et al., 2016, 2019). The current study sought further support for the idea that insomnia disorder may involve a deficiency to downregulate emotional distress over time.
Several studies indicate that sleep supports the downregulation of emotional distress, although findings are equivocal (for a review see Tempesta et al., 2018). Some studies did not find a sleep-dependent decrease in subjective arousal ratings of the same stimuli presented both before and after periods of sleep or wakefulness (Baran et al., 2012), or both before and after a period of sleep either in early or late in the night (Wagner et al., 2002; Groch et al., 2013). Both enhanced and attenuated subjective changes have been reported, across periods of both rapid eye movement (REM) sleep and REM sleep deprivation (Lara-Carrasco et al., 2009; Gujar et al., 2011; Rosales-Lagarde et al., 2012), and REM sleep may have a specific role in modulating the emotional response (van der Helm et al., 2011; Rosales-Lagarde et al., 2012). Objective measures give somewhat more consistent, but still equivocal results: re-exposure of an emotional stimulus after a period of sleep elicits less autonomic arousal and less activation of the amygdala, than when a stimulus is repeated across a comparable period without sleep (Sterpenich et al., 2007; Pace-Schott et al., 2011; van der Helm et al., 2011; Cunningham et al., 2014). It is important to note that re-exposure can give different results than recall. As compared to unsuccessful recall, successful recall involves the amygdala and its functional connectivity with the hippocampus and ventral medial prefrontal cortex (Sterpenich et al., 2009; Lewis et al., 2011; Payne and Kensinger, 2011). The experimental paradigm thus determines what ‘good’ sleep can do. Sleep may on the one hand favour decreased amygdala activity and subjective emotional intensity at later re-exposure to a stimulus that initially elicited an emotional response (van der Helm et al., 2011). Sleep may on the other hand also favour increased amygdala responses in association with improved recall or discerning emotional from neutral stimuli (Menz et al., 2013). Clearly, one has to carefully interpret the meaning of increases and decreases in activation within the paradigm used.
These findings indicate that recall of an emotional experience activates a memory trace in a network of brain circuits that no longer includes the limbic areas that responded to the initial experience. Indeed, functional MRI studies confirm that the brain network activation in response to a novel experience differs considerably from the activation elicited by later re-exposure to the same experience (Takashima et al., 2006), and also from the activation elicited during internal recall and reliving of the same experience (Dolcos et al., 2005; Daselaar et al., 2008). The encoding and retrieval of emotional memory is initially dependent on the hippocampus and amygdala, but this is followed by consolidation and integration of memory traces into the neocortex and long-term depotentiation of the connections with the limbic circuit (Takashima et al., 2006; Nieuwenhuis and Takashima, 2011; Born and Wilhelm, 2012). The process of long-term memory formation can be facilitated by sleep (Takashima et al., 2006; Wagner et al., 2006; Gais et al., 2007; Yoo et al., 2007; Sterpenich et al., 2009; van der Helm et al., 2011). It is hypothesized that parts of the neuronal activity patterns elicited by the wake experience can be reactivated during sleep, while providing a unique neuromodulatory milieu for synaptic plasticity (Hobson and Pace-Schott, 2002; Poe et al., 2010; van der Helm et al., 2011; Vanderheyden et al., 2014). For example, unperturbed REM sleep is characterized by virtually absent noradrenaline (Vanderheyden et al., 2014). However, if the locus coeruleus fails to completely inactivate during sleep, as indexed by phasic EEG arousals (Foote et al., 1980; Aston-Jones and Bloom, 1981) the resulting presence of noradrenaline perturbs normal synaptic processes and may lead to the formation of long-term emotional memory traces without the dissociation from the limbic system (Vanderheyden et al., 2014; Swift et al., 2018). Corticotrophin-releasing factor (CRF) signalling may be involved in the failure of the locus coeruleus to completely inactivate during sleep. During stress, projections from the amygdala release CRF, which acts on CRF1 receptors of noradrenergic locus coeruleus neurons to stimulate noradrenaline release (Pace-Schott et al., 2015; Bangasser et al., 2016).
Intriguingly, this formation of long-term memory that is facilitated by sleep has very persistent effects. Effects of immediate sleep versus initial wakefulness after learning on subsequent recognition performance have been demonstrated to last for several months in case of neutral visual stimuli (Takashima et al., 2006) or word pairs (Mazza et al., 2016) and even for 4 years in case of emotional text (Wagner et al., 2006). These long-lasting consequences of may be highly relevant for insomnia disorder with its key characteristic of sleep fragmentation by arousals, especially during REM sleep (Merica et al., 1998; Perlis et al., 2001; Feige et al., 2008; Riemann et al., 2012). Indeed, a recent observational study reported that subjects with insomnia have a specific deficit in the overnight resolution of emotional distress, which can consequently linger on for even more than a week (Wassing et al., 2016, 2019). These findings suggest a chronic deficiency to dissociate the emotional response from long-term memory traces in insomnia. The fragmented sleep characteristic of insomnia commences already early in life, long before the onset of a full-blown insomnia disorder. We recently genotyped 200 healthy young volunteers devoid of any sleep disorder and found that a higher polygenic risk score for insomnia (Hammerschlag et al., 2017) predicted less slow wave activity in their sleep EEG (Ghaemmaghami et al., 2018). The hypothesized adverse consequences for the overnight resolution of emotional distress would thus start early in life as well. Accordingly, we hypothesized that the pattern of neuronal activity elicited by reliving emotional memories from the distant past resembles the pattern elicited by novel emotional experiences more in insomnia disorder than in normal sleepers.
We used functional MRI to evaluate the brain response to novel and past self-conscious emotional experiences in normal sleepers and patients with insomnia disorder. Whereas self-conscious emotions are highly relevant for psychiatry (Feiring and Taska, 2005; Stuewig and McCloskey, 2005; Schalkwijk, 2015), neuroimaging studies are scarce. Therefore, for both normal sleepers and patients with insomnia disorder, we first investigated which brain areas respond to novel self-conscious emotional experiences. Subsequently, we investigated which brain areas respond to reliving self-conscious emotional experiences from the distant past and evaluated how the activation pattern overlaps with activation during novel emotional experiences. The amount of overlap was subsequently compared between normal sleepers and patients with insomnia disorder to address the hypothesis of insufficient dissociation of the limbic circuit from long-term memory traces in insomnia disorder.
Materials and methods
Participants
Between March 2015 and April 2016, 57 volunteers were recruited through the Netherlands Sleep Registry (Benjamins et al., 2017). The inclusion criterion was an age between 18 and 70 years. Exclusion criteria for both groups were any diagnosed current or past neurological or psychiatric disorder, any current sleep disorder (other than insomnia disorder in the patient group), chronic use of medication, the use of sleep mediation during the prior 2 months, and any MRI contraindication. n = 27 [mean ± standard deviation (SD) age = 45.5 ± 13.4 years, 10 males] fulfilled the diagnostic criteria for insomnia disorder, assessed according to the Diagnostic and Statistical Manual of Mental Disorders (DSM-5; American Psychiatric Association, 2013) and International Classification of Sleep Disorders (ICSD3; Diagnostic Classification Steering Committee, 2014), while n = 30 normal sleepers had no complaints (42.4 ± 15.8 years, 17 males). Age was not significantly different between insomnia disorder and normal sleepers [t(55) = 0.80; P = 0.43; Table 1].
Table 1.
Demographic and sleep characteristics of normal sleepers and subjects with insomnia disorder
| NS (n = 30) | ID (n = 27) | t-statistic | P-value | |
|---|---|---|---|---|
| Demographics | ||||
| n, males (%) | 17 (56.7) | 10 (37.0) | χ2 = 2.20 | 0.14 |
| Age, years | 42.4 (15.8) | 45.5 (13.4) | 0.80 | 0.43 |
| ISI (range 0–28) | 5.1 (4.3) | 18.0 (4.8) | 10.65 | 5.6 × 10−15† |
| Polysomnography | ||||
| TIB, min | 477.0 (60.1) | 489.8 (45.7) | 0.89 | 0.38 |
| SOL, min | 13.6 (9.8) | 16.1 (15.1) | 0.74 | 0.46 |
| WASO, min | 35.1 (28.8) | 47.3 (32.8) | 1.49 | 0.14 |
| TST, min | 419.6 (64.4) | 409.5 (63.9) | −0.58 | 0.56 |
| SE (%) | 88.0 (8.9) | 83.6 (10.0) | −1.76 | 0.084 |
| N1, min | 12.9 (8.2) | 14.8 (9.0) | 0.81 | 0.42 |
| N2, min | 193.7 (48.9) | 189.2 (49.0) | −0.34 | 0.73 |
| N3, min | 126.7 (51.0) | 124.6 (41.2) | −0.17 | 0.87 |
| REM, min | 86.3 (37.4) | 81.0 (36.6) | −0.54 | 0.59 |
| Arousal index a | ||||
| Stage N1, n/h | 10.4 (12.8) | 18.8 (21.2) | 1.82 | 0.074 |
| Stage N2, n/h | 5.9 (4.0) | 8.0 (5.0) | 1.78 | 0.082 |
| Stage N3, n/h | 3.0 (1.5) | 3.8 (2.6) | 1.53 | 0.13 |
| Stage REM, n/h | 8.0 (4.8) | 13.0 (7.1) | 3.14 | 0.0027* |
Values are presented as mean (SD) unless otherwise stated.
aThe number of cortical arousals identified as brief wake-like EEG activity per hour spent in each sleep stage.
*Uncorrected significance threshold P < 0.05; †significant after Dunn-Šidák correction for 20 tests, P < 0.0026 (FWE = 0.05). ISI = Insomnia Severity Index; SE = sleep efficiency; SOL = sleep onset latency; TIB = time in bed; WASO = wake after sleep onset.
Subjective complaints assessed with the Insomnia Severity Index were significantly higher in participants diagnosed with insomnia disorder (18.0 ± 4.8) than in normal sleepers [5.1 ± 4.3, t(55) = 10.65, P = 5.6 × 10−15]. Polysomnography recordings were obtained from two consecutive nights in the sleep lab. The first night was regarded as an adaptation night. Standardized sleep scoring (Berry et al., 2015) was performed for the second night. Comparisons of polysomnography parameters between normal sleepers and insomnia disorder revealed no significant differences in the time spent awake or in any sleep stage (0.08 ≤ P ≤ 0.99; Table 1). This finding is in line with the consensus that polysomnography is not required for the routine evaluation of insomnia and that polysomnography abnormalities are not a defining characteristic of insomnia disorder (Standards of Practice Committee of the American Sleep Disorders Association, 1995). Microstructural analysis, however, revealed significantly more REM arousals in insomnia disorder [t(54) = 3.14, P = 0.003; Table 1], supporting the suggestion that this is a robust marker of insomnia (Feige et al., 2008, 2013; Riemann et al., 2012).
The study was performed in accordance with the Declaration of Helsinki and was approved by the ethics review board of the University of Amsterdam, The Netherlands. Written informed consent was obtained prior to study enrolment.
Procedures
Participants were invited to the Netherlands Institute for Neuroscience for an intake interview, structural MRI scans, karaoke-style audio recordings, and retrieval of autobiographical emotional experiences (see below). A week later, they returned for functional MRI assessments between either 07:30 and 10:00 h (n = 27, 14 with insomnia disorder) or 17:00 and 22:00 h (n = 30, 13 with insomnia disorder) counterbalanced across groups [chi-square test; χ2(1) = 0.41, P = 0.52].
Participants were exposed to stimuli aimed at eliciting novel and relived self-conscious emotions, during concurrent functional MRI and galvanic skin response recordings (see Supplementary material for further details). We specifically induced self-conscious emotions, since they are more relevant to psychological functioning than basic emotions (Feiring and Taska, 2005; Stuewig and McCloskey, 2005; Schalkwijk, 2015). We adapted two original paradigms to induce novel emotional experiences (Sturm et al., 2013), and to perform cued recall of autobiographical memories (Wagner et al., 2011). The ‘novel’ emotional stimuli aimed to induce novel shameful experiences by confronting participants with listening to 16-s fragments of their own solo singing, unaccompanied by music. The recording was obtained a week prior to the experiments in a ‘karaoke’ setup, without explicitly stating what the purpose of the recording was. Just prior to the functional MRI experiment, participants were instructed that they would be exposed to recordings of their own and of others, while the two researchers were listening along. The ‘relived’ emotional stimuli aimed to induce reliving of at least five shameful experiences recalled from the distant past. Participants recalled each shameful experience for 16 s aided by the sets of keywords that they had defined themselves a week before (Supplementary material).
To obtain ‘emotion-specific’ brain activation contrasts, trials with emotional stimuli were interleaved with trials with neutral stimuli (Fig. 1). The novel emotional stimuli were interleaved with novel neutral stimuli consisting of fragments of a recording of the same song, sung by a sex-matched semi-professional singer. The relived emotional stimuli were interleaved with relived neutral stimuli: sets of keywords that participants had defined themselves to describe a trivial non-emotional experience from the same period in time as each of the shameful experiences. There was no difference in the average time since occurrence of the autobiographical events between insomnia disorder (16.9 ± 15.8 years) and normal sleepers [17.9 ± 13.19 years, β(standard error, SE) = –0.97 (2.97), t(209) = −0.33, P = 0.74].
Figure 1.

Induction of novel and relived self-conscious emotions. Each trial started with a fixation-cross presented on the screen for 7 to 9 s, after which the emotional or neutral stimulus was presented. (A) The novel stimuli were audio fragments, of either of the participant’s own solo singing (emotional) or of a semi-professional singer (neutral). (B) Reliving of emotional experiences was aided by the presentation of four cue words ‘cue1’ to ‘cue4’ related to either an emotional or a neutral memory. Subjective emotional intensity was assessed with response options ranging from ‘none’ (1) to ‘strong’ (4). To promote dissipation of the induced emotion, each trial was followed by a 1-back task that closed with a rating of the subjective effort required to perform it (same response options).
To obtain an index of perceived emotional intensity, participants were visually probed with the term ‘intensity’ after each stimulus and allowed 3.5 s to choose one of the response options ‘none’, ‘mild’, ‘quite’, or ‘strong’ using a button-box (Fig. 1). To prevent possible lingering of induced emotions into subsequent trials, participants then had to divert their attention by performing an audio-visual 1-back task for 15 s and rate the amount of effort it took to perform that task. Participants practiced the paradigm prior to the experiment, and were counterbalanced to be first exposed to 20 novel, and subsequently to 20 relived trials, or vice versa. For both the novel and relived stimulus types, the trials were presented in two runs of five emotional and five neutral trials. To prevent stimulus anticipation, half of the emotional trials were followed by neutral trials and half of the neutral trials were followed by emotional trials.
Finally, to confirm that the stimuli induced self-conscious emotions, we asked participants upon completion of each of the runs to rate the intensity of a list of 17 emotions they felt in response to the emotional stimuli (eight negative basic emotions: ‘sadness’, ‘fear’, ‘anger’, ‘upset’, ‘disgust’, ‘rage’, ‘aversion’, and ‘fright’; four positive basic emotions: ‘surprise’, ‘interest’, ‘pleasure’, ‘excitement’; and five self-conscious emotions: ‘shame’, ‘humiliation’, ‘pride’, ‘embarrassment’, and ‘guilt’). Emotion words were presented in random sequence with response options on a Likert-type scale ranging from ‘none’ (0) to ‘strong’ (4).
Brain imaging acquisition and processing
Functional brain data were acquired on a 3-T scanner (Philips Achieva, Philips Healthcare), n = 212 echo-planar images, repetition time: 2500 ms, echo time: 28 ms, voxel-size: 2.5 × 2.5 × 2.5 mm, 43 slices, field-of-view: 240 × 240 mm, flip-angle: 77.2°, parallel imaging SENSE factor 2. A T1-weighted scan was used for anatomical registration and B0 field maps were acquired in between each run to correct the echo planar imaging (EPI) images for magnetic-field distortions (Jenkinson, 2004).
B0 field maps were preprocessed according to previous instructions (Van het Veer et al., 2017). EPI images were motion corrected with MCFLIRT using the middle volume, masked to remove non-brain voxels, spatially smoothed with a Gaussian kernel (5 mm full-width at half-maximum) to reduce noise, and normalized to the grand mean intensity. The ICA-AROMA algorithm was applied to non-aggressively remove motion artefacts (Pruim et al., 2015), and time series extracted from white matter and CSF voxels were used to perform nuisance regression. The EPI time series were high-pass filtered with a cut-off at 1/90 Hz. Transformation matrices were obtained by the combination of a boundary-based registration of the EPI scan to the anatomical scan, and an affine registration of the anatomical scan to the standard-space image (MNI152 T1-weighted anatomical image at 1 mm3, FLIRT). Finally, the EPI images were corrected for subject-specific B0 field distortion maps (Woolrich et al., 2001, 2004; Beckmann et al., 2003).
Statistical analysis
Separately for novel and relived experiences, the subject-level blood oxygen level-dependent (BOLD) responses to emotional stimuli and neutral stimuli were evaluated with general linear models (FSL FEAT v6.00, see Supplementary material; Woolrich et al., 2001). Emotional and neutral stimuli were modelled with two box-car regressors convolved with a double-gamma haemodynamic response function. The emotion-specific BOLD response was obtained for each participant by contrasting the parameter estimates of the two regressors (βemotional − βneutral).
The group-level statistical analyses commenced with a descriptive analysis that evaluated the mean emotion-specific brain responses to novel or to relived experiences within patients with insomnia disorder and within normal sleepers. Then we evaluated our main hypothesis in two steps. First, we tested for differences in the brain response to novel or to relived experiences between normal sleepers and patients with insomnia disorder. Second, we tested for overlap in the brain response to novel or to relived experiences within normal sleepers and within patients with insomnia disorder. Clusters were obtained using the default methods of FSL FEAT, thresholding at |Z| > 3.1 (P < 0.001) and a cluster-based correction for multiple comparisons based on Gaussian Random Field Theory.
The descriptive group-level statistical analyses were performed using a two-way ANOVA (FSL FLAME 1+2; Beckmann et al., 2003; Woolrich et al., 2004) that estimated the mean emotion-specific BOLD response for each combination of stimulus type (‘novel’ or ‘relived’) and group membership (‘insomnia disorder’ or ‘normal sleepers’). Covariates were age and sex, and to account for interindividual differences in perceived emotional intensity (Phan et al., 2003), an additional covariate indicated the participant’s mean intensity difference between the emotional and neutral stimuli.
Subsequently we evaluated our main hypothesis in two steps. First, we defined two masks encompassing the significant clusters in either patients with insomnia disorder or normal sleepers: one mask for novel and the other mask for relived experiences. Separately for novel and relived experiences, we examined the differences in the emotion-specific BOLD response between patients with insomnia disorder and normal sleepers within the corresponding mask using two one-way ANOVAs (FSL FLAME 1+2; Beckmann et al., 2003; Woolrich et al., 2004). We included the covariates age, sex, and perceived emotional intensity, and excluded the covariate if it was not significant.
The second part of our hypothesis stated that the brain response pattern with relived experiences in participants with insomnia disorder would resemble their brain response to novel experiences, because of their deficiency to dissociate the limbic circuit from long-term memory traces. We expected no, or less, overlap in limbic regions in normal sleepers, because their recall of past emotional events would activate long-term memory traces without limbic involvement. This hypothesis was examined separately within patients with insomnia disorder and normal sleepers with conjunction analyses testing for the overlap in emotion-specific BOLD responses to novel and relived experiences (‘easythresh_conj’ algorithm by S. Smith and M. Jenkinson of the FMRIB Image Analysis Group, University of Oxford).
Complementary to the functional MRI analyses, we validated whether the novel and relived emotional stimuli were successful in inducing an emotional response in terms of perceived intensity ratings and autonomic activation (see Supplementary material for skin conductance signal processing). A three-way mixed-effects ANOVA evaluated the fixed-effects of stimulus type (‘novel’ versus ‘relived’), stimulus valence (‘emotional’ versus ‘neutral’), and group-membership (‘insomnia disorder’ versus ‘normal sleepers’) on perceived intensity ratings and event-related galvanic skin responses. Confound regressors were age and sex. We found that the first stimulus elicited greater galvanic skin responses as compared to the subsequent stimuli (P = 0.001), we therefore included a factor indicating each first stimulus in the galvanic skin response models. The random-effects design matrix included a random intercept for each participant, and a random slope for first-stimulus for galvanic skin response models. Standardized β-estimates are reported, and P < 0.05 was considered significant.
Data availability
The data and software/scripts used in this study are available on reasonable request from the corresponding author.
Results
Stimulus validation
In support of the validity of using the tasks for our aims, we confirmed that ‘shame’ was on average the most intensely perceived emotion (Supplementary Table 1). Furthermore, a significant main effect of stimulus valence indicated that emotional stimuli were on average rated more intense than neutral ones [F(1,2223) = 459.57, P = 7.9 × 10−93]. Also galvanic skin response amplitudes were on average greater in response to emotional stimuli than to neutral ones [F(1,502) = 5.02, P = 0.03].
Mixed effect ANOVAs for perceived intensity ratings and galvanic skin responses
A significant three-way interaction [F(1,2223) = 9.41, P = 0.002] indicated that the group difference in perceived intensity ratings between emotional and neutral relived experiences was different for novel experiences. Although the three-way interaction was significant, post hoc t-tests showed that here were no significant group differences in perceived intensity ratings to novel emotional [β (SE) = 0.28 (0.20), t(563) = 1.40, P = 0.16], or novel neutral stimuli [β (SE) = 0.05 (0.12), t(553) = 0.42, P = 0.68]. Also for relived experiences, there were no significant group differences in perceived intensity ratings to emotional [β (SE) = 0.02 (0.15), t(555) = 0.12, P = 0.91], or neutral stimuli [β (SE) = 0.22 (0.16), t(546) = 1.43, P = 0.15].
Furthermore, a significant three-way interaction [F(1,502) = 4.16, P = 0.04] indicated that the group difference in galvanic skin response amplitudes between emotional and neutral relived experiences was different for novel experiences. Post hoc t-tests indicated that while there were no group differences in galvanic skin response amplitudes to novel emotional [β (SE) = 0.17 (0.24), t(127) = 0.70, P = 0.48], or novel neutral stimuli [β (SE) = 0.27 (0.18), t(100) = 1.46, P = 0.15]; relived emotional stimuli elicited greater galvanic skin responses than neutral ones in patients with insomnia disorder [β (SE) = 0.31 (0.11), t(140) = 3.00, P = 0.003], whereas in normal sleepers, reliving emotional or neutral events from the past elicited similar galvanic skin responses [β (SE) = –0.01 (0.11), t(127) = –0.09, P = 0.93].
Combined, these findings indicate that the emotional response to novel experiences was similar in patients with insomnia disorder and normal sleepers. For relived experiences on the other hand, we observed a differential response between normal sleepers and patients with insomnia disorder. Normal sleepers experienced emotional and neutral relived stimuli with similar intensity in terms of autonomic responses; patients with insomnia disorder, however, experienced a stronger autonomic response to relived emotional stimuli than to neutral ones.
Emotion-specific BOLD responses to novel and relived experiences
Table 2 provides an overview of the most important functional MRI findings; detailed descriptions of the clusters and local maximally significant voxels are provided in Supplementary Tables 3–8.
Table 2.
Emotion-specific BOLD activations to novel and relived experiences
| Region | Novel | Relived | |||||||
|---|---|---|---|---|---|---|---|---|---|
| H | MNI (mm) | Z | MNI (mm) | Z | |||||
| x | y | z | x | y | z | ||||
| Normal sleepers | |||||||||
| Anterior cingulate gyrus | R | 1 | 17 | 34 | 5.37 | - | - | - | - |
| Insula | R | 39 | 10 | 3 | 4.32 | - | - | - | - |
| L | −40 | 7 | −2 | 4.33 | - | - | - | - | |
| Orbitofrontal cortex | R | 36 | 29 | 0 | 5.13 | - | - | - | - |
| Operculum cortex | R | 47 | 25 | −3 | 5.51 | - | - | - | - |
| L | −47 | 13 | 1 | 4.27 | - | - | - | - | |
| Inferior frontal gyri | R | 49 | 32 | 2 | 5.53 | - | - | - | - |
| L | −54 | 9 | 1 | 4.79 | - | - | - | - | |
| Superior frontal gyrus | R | 4 | 14 | 59 | 7.34 | - | - | - | - |
| Planum temporale | R | 64 | −34 | 18 | 6.29 | - | - | - | - |
| L | −61 | −29 | 9 | 5.53 | - | - | - | - | |
| Anterior supramarginal gyrus | R | 62 | −33 | 40 | 4.41 | - | - | - | - |
| L | −66 | −38 | 23 | 4.35 | - | - | - | - | |
| Posterior supramarginal gyrus | R | 59 | −39 | 18 | 6.55 | - | - | - | - |
| L | −66 | −43 | 12 | 4.96 | −67 | −47 | 26 | 3.99 | |
| Temporal pole | R | 54 | 14 | −6 | 6.19 | - | - | - | - |
| Insomnia disorder | |||||||||
| Amygdala | R | 24 | −4 | −12 | 4.26 | - | - | - | - |
| Anterior cingulate gyrus | R | 3 | 20 | 30 | 5.59 | 1 | 26 | 32 | 4.61 |
| L | −2 | 19 | 36 | 6.40 | −4 | 24 | 31 | 4.70 | |
| Paracingulate gyrus | R | 1 | 22 | 38 | 6.73 | 1 | 21 | 37 | 4.15 |
| L | −4 | 21 | 38 | 6.42 | −4 | 10 | 45 | 4.85 | |
| Supplementary motor area | R | 6 | 2 | 67 | 6.23 | 1 | 2 | 47 | 4.29 |
| L | −1 | −4 | 66 | 5.59 | −4 | 5 | 47 | 4.24 | |
| Insula | R | 45 | 12 | −4 | 6.39 | - | - | - | - |
| L | −37 | −21 | 2 | 4.77 | - | - | - | - | |
| Thalamus | R | 4 | −4 | 8 | 4.60 | - | - | - | - |
| L | −6 | −5 | 8 | 4.99 | - | - | - | - | |
| Orbitofrontal cortex | R | 44 | 23 | −6 | 5.96 | - | - | - | - |
| L | −29 | 11 | −18 | 5.75 | - | - | - | - | |
| Operculum cortex | R | 55 | −28 | 22 | 3.70 | 54 | −26 | 21 | 4.17 |
| L | −53 | −39 | 28 | 5.24 | - | - | - | - | |
| Inferior frontal gyri | R | 51 | 11 | 3 | 4.83 | - | - | - | - |
| L | −48 | 20 | 3 | 4.26 | - | - | - | - | |
| Superior frontal gyrus | R | 8 | 8 | 69 | 5.53 | - | - | - | - |
| L | −2 | 15 | 61 | 5.34 | - | - | - | - | |
| Planum temporale | R | 50 | −31 | 15 | 6.54 | 65 | −25 | 12 | 3.55 |
| L | −44 | −32 | 7 | 5.69 | - | - | - | - | |
| Posterior middle temporal gyrus | R | 52 | −37 | 2 | 4.38 | 53 | −20 | −6 | 4.42 |
| L | −55 | −14 | −9 | 4.12 | - | - | - | - | |
| Anterior supramarginal gyrus | R | - | - | - | - | 66 | −25 | 26 | 4.13 |
| L | −63 | −40 | 29 | 4.66 | - | - | - | - | |
| Posterior supramarginal gyrus | R | 63 | −39 | 18 | 6.41 | 66 | −35 | 21 | 4.32 |
| L | −60 | −43 | 26 | 5.41 | −59 | −51 | 26 | 3.89 | |
| Temporal pole | R | 54 | 8 | −16 | 5.29 | - | - | - | - |
| L | −34 | 23 | −31 | 4.94 | - | - | - | - | |
The table shows the coordinates of within-cluster local maxima of emotion-specific BOLD responses. Values in bold indicate overlap in BOLD responses to novel and relived emotional stimuli confirmed by conjunction analysis. Clusters were obtained by thresholding at |Z| > 3.1 (P < 0.001), and cluster-based correction for multiple comparisons was applied at P < 0.05, using Gaussian Random Field Theory. H = hemipshere; L = left; R = right.
BOLD responses to novel self-conscious emotional experiences
In normal sleepers, eight clusters across known brain networks showed BOLD activations specific to experiencing novel self-conscious emotions. Activations were seen in the sensorimotor network, extended auditory network, and salience network (Fig. 2A). Within the salience network, activation clusters encompassed the bilateral ACC, the bilateral anterior insula, and cortices often associated with the salience network: the right orbitofrontal cortex, and inferior frontal gyri: right triangularis, and bilateral frontal operculum. Deactivations were observed in seven clusters encompassing the posterior default mode network: bilateral superior occipital cortex, and bilateral angular gyrus; as well as in the bilateral middle frontal gyrus and the bilateral middle temporal gyrus.
Figure 2.

Emotion-specific BOLD responses to novel and relived experiences. Group means for the contrast between emotional and neutral stimuli. The mean percent BOLD change is coded by hue and the Z-statistic is coded by opacity. Significant clusters are indicated with black outlines (FLAME 1+2, |Z| > 3.10 and P < 0.05; see Supplementary Tables 5–8 for detailed description of clusters). (A) In normal sleepers, emotion-specific BOLD activations to novel experiences were observed in the salience network, the extended auditory system, and the orbitofrontal cortex. (B) In patients with insomnia, emotion-specific BOLD activations to novel experiences included these areas but covered wider areas and were also seen in the amygdala. (C) In normal sleepers, emotion-specific BOLD activations to relived experiences were observed in the left supramarginal gyrus and just anterior and superior of the caudate nucleus (D) whereas in patients with insomnia, BOLD activations were most pronounced in the dorsal ACC, and pre-SMA, right parietal operculum cortex, right planum temporale, right parietal operculum cortex, and bilateral supramarginal gyrus. Amg = amygdala; CA = caudate nucleus; CB = cerebellum; dACC = dorsal ACC; dlPFC = dorsolateral prefrontal cortex; IFG = inferior frontal gyrus; Ins = insula; LgG = lingual gyrus; mPFC = medial prefrontal cortex; MTG = middle temporal gyrus; OFC = orbital frontal gyrus; Op = operculum cortex; PCC = posterior cingulate cortex; PHG = parahippocampal gyrus; PT = planum temporale; RSC = retrosplenial cortex; SFG = superior frontal gyrus; sLOC = superior lateral occipital cortex; SMG = supramarginal gyrus; Thal = thalamus.
In insomnia disorder, emotion-specific BOLD activations were likewise seen in nine clusters across the same brain networks as in found in normal sleepers, be it more extended and including limbic regions such as the right amygdala, posterior cingulate cortex, bilateral thalamus, and the paralimbic medial PFC (Fig. 1B). No significant deactivations were observed in insomnia disorder.
A one-way ANOVA with age as a confounder revealed no significant differences in the emotion-specific BOLD response to novel experiences between normal sleepers and patients with insomnia disorder. Combined, the findings indicate that novel self-conscious emotions elicit activations of similar magnitude in the salience network and limbic circuit (ACC) in both normal sleepers and patients with insomnia.
BOLD responses to relived self-conscious emotional experiences
In normal sleepers, emotion-specific BOLD responses with reliving memories from the distant past included a cluster in the left supramarginal gyrus, and a cluster just anterior and superior of the caudate nucleus (Fig. 1C). Furthermore, a negative contrast indicated weaker BOLD responses with reliving emotional memories as compared to neutral memories in a cluster encompassing the left posterior parahippocampal and lingual gyrus, in the right ventrolateral prefrontal cortex, in the left inferior temporal-occipital junction, and in two clusters in the posterior default mode network: bilateral retrosplenial cortex and right superior lateral occipital cortex.
Whereas the deactivation pattern was similar in insomnia disorder as in normal sleepers (Supplementary Tables 7 and 8), additional activation clusters were found in insomnia disorder: the ACC, paracingulate gyrus and pre-supplementary motor area (pre-SMA); right parietal operculum cortex, right planum temporale, right parietal operculum cortex, bilateral supramarginal gyrus, and left posterior temporal gyri. Of note, these were all regions that also responded to novel self-conscious emotions (Fig. 2D). A one-way ANOVA with age as a confounder indicated that participants with insomnia disorder showed stronger emotion-specific responses to relived experiences in the dorsal ACC [Pcluster = 0.035, Zmax = 3.72 at MNIxyz = (1, 3, 45) mm].
Similarities in emotion-specific BOLD responses to novel and relived experiences
The findings suggest that normal sleepers activate a markedly different brain circuit while reliving emotional memories from the distant past as compared to when they are exposed to novel emotional experiences. In patients with insomnia, however, the brain circuits recruited with reliving distant emotional memories overlapped with the circuits recruited during a novel emotional experience, notably so for the dorsal ACC. Indeed, a formal conjunction analysis showed no overlap in emotion-specific BOLD responses to novel and relived experiences in normal sleepers, while there was significant overlap in insomnia disorder in two clusters: one cluster encompassing the dorsal ACC, paracingulate gyrus, and part of the pre-SMA [Pcluster = 3.9 × 10−6, Zmax = 4.70 at MNIxyz = (−4, 24, 31) mm; Fig. 3], and a cluster in the right supramarginal gyrus [Pcluster = 0.0005, Zmax = 4.32 at MNIxyz = (66, −35, 21) mm].
Figure 3.
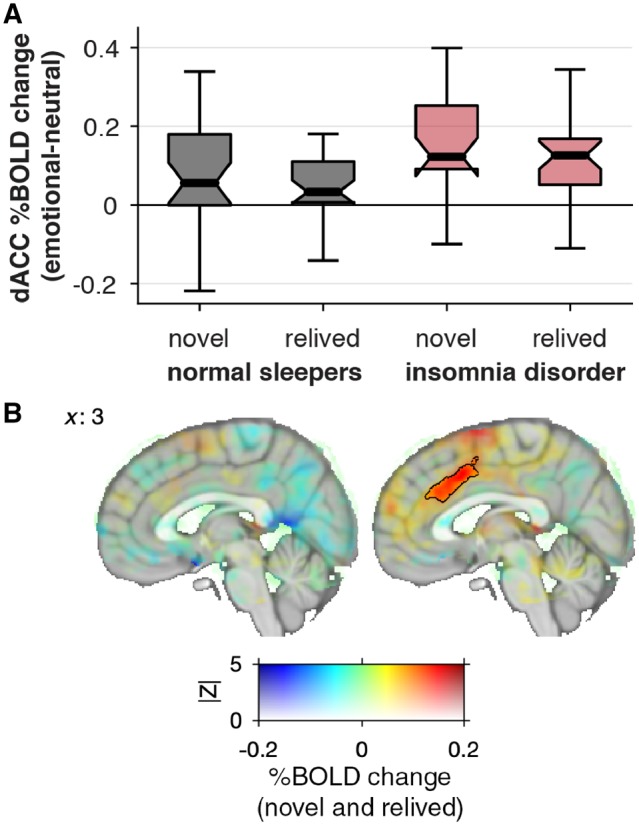
The dorsal ACC responds to both novel and relived self-conscious emotions in patients with insomnia disorder but not in normal sleepers. (A) Boxplots: thick lines indicate the median per cent BOLD change within the dorsal ACC (significant cluster found in insomnia disorder, black outline in B). The box extends from the 25th to the 75th percentile and whiskers extend to the most extreme data point within the range of the box ± 1.5 × IQR. (B) The mean emotion-specific BOLD response to relived and novel experiences is coded by hue (emotional–neutral contrast). The voxel’s Z-statistic for conjunction is coded with opacity (between 0 < |Z| < 5; see ‘Materials and methods’ section). The significantly overlapping cluster (|Z| > 3.10 and P < 0.05) in the dorsal ACC is indicated with black outline; one other cluster was found in the right supramarginal gyrus (not shown; see ‘Results’ section).
Discussion
The current study sought support for the hypothesis that patients suffering from insomnia show insufficient dissociation of the limbic circuit from long-term memory traces. We used functional MRI to evaluate whether participants with insomnia show more overlap in BOLD responses to novel and relived experiences than normal sleepers do.
In normal sleepers, we observed a limbic response in the ACC while experiencing novel emotional distress but not while reliving emotional distress from the distant past. This finding is in line with the hypothesis that the limbic circuit was effectively dissociated from long-term memory traces in normal sleepers. In insomnia disorder, we observed a limbic response in the ACC while experiencing novel emotional distress. In contrast to normal sleepers, however, the BOLD response pattern to relived experiences showed a partial overlap with the response to novel experiences in those with insomnia disorder. Conjunction analysis specifically identified overlap in the bilateral supramarginal gyrus and the dorsal ACC. Group comparison indeed confirmed a significantly stronger dorsal ACC activation in insomnia disorder than in normal sleepers during reliving of self-conscious emotional experiences from the past. These findings are in line with the hypothesis that insomnia disorder involves a deficiency to dissociate the limbic system from emotional long-term memory traces.
The conjunction mask, indicating overlap between brain responses to relived emotional experiences and novel emotional stimuli in insomnia disorder, encompassed the dorsal ACC and part of the pre-SMA. The ACC is a relatively large cortical region that is part of the limbic system, and has functional anatomical connections with cortical regions involved in higher-order cognition and sensorimotor function such as the pre-SMA (Bush et al., 2000; Etkin et al., 2011). Both the pre-SMA and ACC have been implicated in a wide range of functions. Studies for example showed pre-SMA involvement in preparation and (cognitive) modulation of motor responses (Forstmann et al., 2008; Kim et al., 2010), and ACC involvement in pain and basic emotions (Vogt, 2005), prediction error (Botvinick et al., 2004), sensory integration (Etkin et al., 2011), and cognitive control and response modulation (Bush et al., 2000). The ACC has been subdivided into a dorsal region that serves cognitive appraisal and expression of emotions, and a ventral region involved in emotion regulation through its strong connectivity with the limbic regions (Vogt et al., 1992; Devinsky et al., 1995; Bush et al., 2000; Etkin et al., 2011). Interindividual differences in activation of the dorsal ACC while reliving emotional experiences in insomnia disorder was correlated with galvanic skin responses (Supplementary material). This ancillary finding indicates that the dorsal ACC response to relived experiences in insomnia disorder may signify primary somatic reactivity rather than higher-order emotional appraisal. It is tempting to suggest that the dorsal ACC activation while recalling emotional distress from the past in insomnia disorder might represent the neurobiological substrate of the link between long-lasting emotional distress and hyperarousal (Wassing et al., 2016). The bilateral supramarginal gyrus also showed activation during relived self-conscious emotional experiences in insomnia disorder. The wide range of functions attributed to the supramarginal gyrus do not currently allow for a clear interpretation of a possible role in insomnia disorder and emotional distress.
The BOLD response pattern to novel self-conscious emotional experiences in normal sleepers and in insomnia disorder is in line with findings of cognitive appraisal of emotions, which recruits key regions such as the medial prefrontal cortex, dorsal ACC, lateral orbitofrontal cortex, and the inferior frontal gyri (Levesque et al., 2003; Ochsner et al., 2004; Phan et al., 2005; Urry et al., 2006). In insomnia disorder, however, we found additional recruitment of limbic areas with novel emotional experiences, which may signify insufficient top-down cognitive control or the use of maladaptive coping strategies (Etkin and Wager, 2007). Indeed, Morin et al. (2003) found that people with insomnia disorder experience a greater impact of minor daily stressful events and suggested a role of dysfunctional emotional appraisal in the aetiology of insomnia disorder.
Possible mechanisms underlying disrupted formation of emotional long-term memory in insomnia disorder
Although previous neuroimaging studies are equivocal in the identification of specific brain areas involved in the aetiology of insomnia disorder (Tahmasian et al., 2018), a role of the ACC has frequently been reported. (Plante et al., 2012) applied magnetic resonance spectroscopy and reported that people with insomnia have a low concentration of the inhibitory neurotransmitter GABA in the ACC. (Seo et al., 2018) used a fear conditioning and extinction paradigm and implicated the dorsal ACC in the delayed acquisition of fear extinction observed in insomnia patients as compared to controls. Another notable finding is the activation of the ACC by the claustrum during REM sleep (Renouard et al., 2015; Luppi et al., 2017). This is interesting because both structures show enriched expression of risk genes for insomnia (Jansen et al., 2019), and restless REM sleep is a characteristic sleep EEG signature of insomnia (Feige et al., 2008; Riemann et al., 2012), once more confirmed by the polysomnography findings in the current sample. The arousals that occur during restless REM sleep may indicate a failure to completely inactivate the locus coeruleus (Foote et al., 1980; Aston-Jones and Bloom, 1981). If noradrenaline is present during REM sleep, it could interfere with the synaptic dissociation of the limbic circuit with the formation of long-term memory traces (Vanderheyden et al., 2014; Swift et al., 2018). Although the findings of this cross-sectional study do not provide direct support for this hypothesis, we postulate that this may show markedly in the ACC, which is specifically activated during REM sleep (Renouard et al., 2015; Luppi et al., 2017). But we add that the current findings are also compatible with impaired processing of emotional memories not specifically dependent on sleep.
One alternative interpretation could be that inappropriate ACC activation during recall of past experiences might be explained better by current generalized hyperarousal in insomnia disorder. This alternative hypothesis would predict higher ACC activation in insomnia disorder than in normal sleepers irrespective of whether stimuli concern novel or relived emotional experiences. For novel experiences, however, patients with insomnia disorder did not differ from normal sleepers: neither for emotion-specific galvanic skin response amplitudes or BOLD responses, nor for perceived intensity ratings. Patients with insomnia disorder showed significantly stronger emotion-specific ACC activation and galvanic skin responses exclusively while reliving memories from the distant past. In addition, ancillary analyses using several indices of trait and state-like hyperarousal did not support the alternative hypothesis that a more generalized hyperarousal would better explain our findings (Supplementary material, ‘Sensitivity analysis and specificity of dorsal ACC activation to relived emotional experiences’ section).
Another alternative interpretation could be that inappropriate ACC activation during reliving of past experiences is due to group differences in emotional reactivity during the original experience. We have no access to the emotional impact of the experience when it occurred in the distant past. However, we did assess the emotional impact of a current novel experience. As mentioned above, for novel experiences, insomnia disorder did not differ from normal sleepers: neither for emotion-specific galvanic skin response amplitudes or BOLD responses, nor for perceived intensity ratings. Stronger emotion-specific ACC activation and galvanic skin responses in insomnia disorder were exclusive to reliving memories from the distant past. We moreover evaluated whether insomnia disorder and normal sleepers differed with respect to the currently experienced subjective intensity of relived emotions. A multivariate analysis indicated no group differences in the intensity ratings of 17 emotions obtained directly upon completion of each of the two runs while reliving past experiences [one-way MANOVA; F(1,52) = 1.18, P = 0.72].
Our findings resemble to some extent those of a study on post-traumatic stress disorder (PTSD), another disorder characterized by hyperarousal. During recall of emotional memories, people with PTSD showed a stronger dorsal ACC response than controls that had experienced similarly intense emotional events, but had not developed PTSD (Whalley et al., 2013). The findings suggest that the response of the dorsal ACC in the PTSD group did not seem better explained by a more intense original experience.
Combined, we may tentatively conclude that the dorsal ACC response in insomnia disorder while reliving emotional experiences is not likely to merely result from a more intense emotion during the first experience, nor of generalized hyperarousal. It may signify a deficiency to dissociate the ACC from emotional long-term memory traces.
Several aspects of the present study may limit its interpretation. First, the volunteers were recruited from the Netherlands Sleep Registry (NSR) which is not a random sample from the general population, and may be slightly biased to subjects with a special interest in sleep or participating in scientific research. We purposefully did not sample exclusively from sleep clinics because, unfortunately, awareness for insomnia in general practice is still insufficient (Bjorvatn et al., 2017), and sleep clinic-based studies may over-represent more complex insomnia cases. In fact, as compared to clinics, the sample of the NSR may reach a more diverse population of patients with insomnia disorder, because we stress the need for volunteers covering the whole spectrum of good to bad sleepers. Indeed, the NSR has a uniform insomnia severity distribution: 38% are without insomnia, 29% may have subthreshold insomnia, and 33% have clinical insomnia (Morin et al., 2011; Benjamins et al., 2017).
A further possible limitation is that a small difference in sex distribution occurred because we did not select on sex, and insomnia is more common in females than males. We recruited more female participants with insomnia disorder (63.0%) than female normal sleepers (43.3%) and might alternatively have excluded some of the male controls and/or female insomnia applicants. The difference in sex distribution was non-significant (P = 0.14), and we verified that sex did not affect the emotion-specific BOLD response, amplitude of galvanic skin responses or perceived intensity ratings (Supplementary material, ‘Effects of age and sex on perceived intensity ratings and galvanic skin response amplitudes’ and ‘Effects of age and sex on the emotion-specific BOLD response’ sections). Another possible limitation is that we elicited novel and relived emotional distress through two different stimulus modalities; auditory to elicit novel distress and visual to cue the requested start of reliving past distress. Therefore, novel and relived experiences activated different sensory areas (Supplementary Table 4). Moreover, a sensory stimulus is required to elicit novel distress, but not for reliving past distress. Relived emotions may also have acquired secondary emotional loading, like regret, in contrast to the more primary novel emotions. Our design aimed to distill the emotional component of the induced experiences, by contrasting brain activity during novel and relived experienced with brain activity of corresponding neutral experiences. Some within-subject variability in the specificity of emotional and neutral distant memories and between-subject variability in other processes is likely to have remained and may have lowered the signal to noise ratio in our assessments. This did not preclude our finding of systematic differences in subjective ratings and in brain responses in areas involved in salience and emotion. A final possible limitation is that we do not have access to the detailed sleep history since the original occurrence of the relived stimuli. We found no evidence that features of sleep the night before our experimental probing of emotional circuits were associated with either the dorsal ACC BOLD response, the galvanic skin response amplitudes, or perceived intensity ratings (Supplementary Table 2). Given the long time scale of past emotional experiences in this study (on average about 15 years ago), and the large night-to-night variability of sleep in insomnia (Buysse et al., 2010), we considered it unlikely that polysomnography variables derived from a single night would reflect trait-like sleep characteristics across years of insomnia. Sleep features obtained during a single night do not adequately represent the proposed more chronic process we hypothesized to underlie the failure to dissociate the dorsal ACC from the emotional memory trace. Our hypothesis is that people who are diagnosed with insomnia later in life, may have shown signs of restless sleep and insufficient dissociation of ACC involvement and dissipation of emotional distress already early in life. The hypothesis is supported by two observations. First, retrospective sleep history assessment since one’s earliest memories, indicates that people with a current diagnosis are more likely to have experienced sleep complaints already early in life (unpublished data; Benjamins et al., 2017). Second, we recently genotyped 200 healthy young volunteers devoid of any sleep disorder diagnosis and found that a higher polygenic risk score for insomnia (Hammerschlag et al., 2017) predicted less slow wave activity in their current polysomnography (Ghaemmaghami et al., 2018). Notwithstanding the study’s sampling and design limitations, the major strengths were the considerable sample size (n = 57) in comparison to most neuroimaging studies, careful design of the stimulus presentation paradigm and its validation through autonomic-activity assessments and self-rated intensity of emotions. Furthermore, we controlled for known confounders including time of day and differences in perceived emotional intensity between stimulus modalities. Finally, we report a chronic insufficiency to dissociate the dorsal ACC from long-term memory traces in insomnia, a finding that was further corroborated by showing that this dorsal ACC response was associated with autonomic reactivity, which provides a possible neurobiological link between long-lasting emotional distress and hyperarousal.
In conclusion, the present study adds to the converging body of evidence that implicate the ACC as an important cortical area in the mechanisms involved in insomnia. Our findings suggest involvement of the dorsal ACC in insufficient long-term adaptation to emotional memory in people suffering from insomnia. Future studies into fundamental mechanisms underlying insomnia could evaluate whether REM-specific activation of the ACC (Renouard et al., 2015) is involved in the overnight dissociation of its functional connectivity, and whether such processes are perturbed in case of a failing shutdown of noradrenergic activity during REM sleep (Vanderheyden et al., 2014; Swift et al., 2018).
Supplementary Material
Acknowledgements
The authors give special thanks to the volunteers for their dedicated participation in this study, and to the students involved in recruitment and assessments: C. Dekker, V. Hohn, M. Korpelshoek, K. Müller, L. Nijboer, C. Oosterwijk, S. Riemsma, M. Spaander, G. Thijsma, and N. Veerman.
Glossary
Abbreviations
- ACC
anterior cingulate cortex
- BOLD
blood oxygen level-dependent
- REM
rapid eye movement
- SMA
supplementary motor area
Funding
This work was supported by ZONMW Neuropsychoanalysis Fund Grant 16.561.0001 (to F.S. and E.J.W.V.S.), VICI Grant 453.07.001 (to E.J.W.V.S.), all part of the Netherlands Organization of Scientific Research (NWO); by the Grant 253/2012 of the Bial Foundation (to L.M.T.); and by the European Research Council ERC-ADG-2014–671084 INSOMNIA and ERC-2016-PoC-737634 INSOMNIA BEAT IT Grants (to E.J.W.V.S.).
Competing interests
All authors declare no conflict of interest.
References
- American Psychiatric Association. DSM-5: diagnostic and statistical manual of mental disorders. 5th edn. Washington, DC: American Psychiatric Press; 2013. [Google Scholar]
- Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci 1981; 1: 876–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bangasser DA, Wiersielis KR, Khantsis S. Sex differences in the locus coeruleus-norepinephrine system and its regulation by stress. Brain Res 2016; 1641: 177–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baran B, Pace-Schott EF, Ericson C, Spencer RM. Processing of emotional reactivity and emotional memory over sleep. J Neurosci 2012; 32: 1035–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beckmann CF, Jenkinson M, Smith SM. General multilevel linear modeling for group analysis in FMRI. Neuroimage 2003; 20: 1052–63. [DOI] [PubMed] [Google Scholar]
- Benjamins JS, Migliorati F, Dekker K, Wassing R, Moens S, Blanken TF et al. . Insomnia heterogeneity: characteristics to consider for data-driven multivariate subtyping. Sleep Med Rev 2017; 36: 71–81. [DOI] [PubMed] [Google Scholar]
- Berry RB, Brooks R, Gamaldo CE, Harding SM, Lloyd RM, Marcus CL. et al. The AASM manual for the scoring of sleep and associated events: rules, terminology and technical specifications, version 2.2. Darien, Illinois: American Academy of Sleep Medicine, 2015. [Google Scholar]
- Bjorvatn B, Meland E, Flo E, Mildestvedt T. High prevalence of insomnia and hypnotic use in patients visiting their general practitioner. Fam Pract 2017; 34: 20–4. [DOI] [PubMed] [Google Scholar]
- Bonnet MH, Arand DL. Hyperarousal and insomnia: state of the science. Sleep Med Rev 2010; 14: 9–15. [DOI] [PubMed] [Google Scholar]
- Born J, Wilhelm I. System consolidation of memory during sleep. Psychol Res 2012; 76: 192–203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick MM, Cohen JD, Carter CS. Conflict monitoring and anterior cingulate cortex: an update. Trends Cogn Sci 2004; 8: 539–46. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci 2000; 4: 215–22. [DOI] [PubMed] [Google Scholar]
- Buysse DJ, Cheng Y, Germain A, Moul DE, Franzen PL, Fletcher M et al. . Night-to-night sleep variability in older adults with and without chronic insomnia. Sleep Med 2010; 11: 56–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham TJ, Crowell CR, Alger SE, Kensinger EA, Villano MA, Mattingly SM et al. . Psychophysiological arousal at encoding leads to reduced reactivity but enhanced emotional memory following sleep. Neurobiol Learn Mem 2014; 114: 155–64. [DOI] [PubMed] [Google Scholar]
- Daselaar SM, Rice HJ, Greenberg DL, Cabeza R, LaBar KS, Rubin DC. The spatiotemporal dynamics of autobiographical memory: neural correlates of recall, emotional intensity, and reliving. Cereb Cortex 2008; 18: 217–29. [DOI] [PubMed] [Google Scholar]
- Devinsky O, Morrell MJ, Vogt BA. Contributions of anterior cingulate cortex to behaviour. Brain 1995; 118 (Pt 1): 279–306. [DOI] [PubMed] [Google Scholar]
- Diagnostic Classification Steering Committee. ICSD3—international classification of sleep disorders: diagnostic and coding manual. 3rd edn Rochester, MN: American Sleep Disorders Association; 2014. [Google Scholar]
- Dolcos F, LaBar KS, Cabeza R. Remembering one year later: role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proc Natl Acad Sci USA 2005; 102: 2626–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A, Egner T, Kalisch R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn Sci 2011; 15: 85–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A, Wager TD. Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am J Psychiatry 2007; 164: 1476–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feige B, Al-Shajlawi A, Nissen C, Voderholzer U, Hornyak M, Spiegelhalder K et al. . Does REM sleep contribute to subjective wake time in primary insomnia? A comparison of polysomnographic and subjective sleep in 100 patients. J Sleep Res 2008; 17: 180–90. [DOI] [PubMed] [Google Scholar]
- Feige B, Baglioni C, Spiegelhalder K, Hirscher V, Nissen C, Riemann D. The microstructure of sleep in primary insomnia: an overview and extension. Int J Psychophysiol 2013; 89: 171–80. [DOI] [PubMed] [Google Scholar]
- Feiring C, Taska LS. The persistence of shame following sexual abuse: a longitudinal look at risk and recovery. Child Maltreat 2005; 10: 337–49. [DOI] [PubMed] [Google Scholar]
- Foote SL, Aston-Jones G, Bloom FE. Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proc Natl Acad Sci USA 1980; 77: 3033–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forstmann BU, Dutilh G, Brown S, Neumann J, von Cramon DY, Ridderinkhof KR et al. . Striatum and pre-SMA facilitate decision-making under time pressure. Proc Natl Acad Sci USA 2008; 105: 17538–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gais S, Albouy G, Boly M, Dang-Vu TT, Darsaud A, Desseilles M et al. . Sleep transforms the cerebral trace of declarative memories. Proc Natl Acad Sci USA 2007; 104: 18778–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghaemmaghami P, Muto V, Jaspar M, Meyer C, Elansary M, VanEgroo M et al. . The genetic liability for insomnia is associated with lower amount of slow wave sleep in young and healthy individuals. In: Frontiers in Neuroscience Conference Abstract: Belgian Brain Congress 2018—Belgian Brain Council 2018. [Google Scholar]
- Groch S, Wilhelm I, Diekelmann S, Born J. The role of REM sleep in the processing of emotional memories: evidence from behavior and event-related potentials. Neurobiol Learn Mem 2013; 99: 1–9. [DOI] [PubMed] [Google Scholar]
- Gujar N, McDonald SA, Nishida M, Walker MP. A role for REM sleep in recalibrating the sensitivity of the human brain to specific emotions. Cereb Cortex 2011; 21: 115–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammerschlag AR, Stringer S, de Leeuw CA, Sniekers S, Taskesen E, Watanabe K et al. . Genome-wide association analysis of insomnia complaints identifies risk genes and genetic overlap with psychiatric and metabolic traits. Nat Genet 2017; 49: 1584–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobson JA, Pace-Schott EF. The cognitive neuroscience of sleep: neuronal systems, consciousness and learning. Nat Rev Neurosci 2002; 3: 679–93. [DOI] [PubMed] [Google Scholar]
- Jansen PR, Watanabe K, Stringer S, Skene N, Bryois J, Hammerschlag AR et al. . Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nat Genet 2019; 51: 394–403. [DOI] [PubMed] [Google Scholar]
- Jenkinson M. Improving the registration of B0-disorted EPI images using calculated cost function weights. Neuroimage 2004; 22: e1544–5. [Google Scholar]
- Kim JH, Lee JM, Jo HJ, Kim SH, Lee JH, Kim ST et al. . Defining functional SMA and pre-SMA subregions in human MFC using resting state fMRI: functional connectivity-based parcellation method. Neuroimage 2010; 49: 2375–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kindt M. The surprising subtleties of changing fear memory: a challenge for translational science. Philos Trans R Soc Lond B Biol Sci 2018; 373: 20170033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lara-Carrasco J, Nielsen TA, Solomonova E, Levrier K, Popova A. Overnight emotional adaptation to negative stimuli is altered by REM sleep deprivation and is correlated with intervening dream emotions. J Sleep Res 2009; 18: 178–87. [DOI] [PubMed] [Google Scholar]
- Levesque J, Eugene F, Joanette Y, Paquette V, Mensour B, Beaudoin G et al. . Neural circuitry underlying voluntary suppression of sadness. Biol Psychiatry 2003; 53: 502–10. [DOI] [PubMed] [Google Scholar]
- Lewis PA, Cairney S, Manning L, Critchley HD. The impact of overnight consolidation upon memory for emotional and neutral encoding contexts. Neuropsychologia 2011; 49: 2619–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luppi PH, Billwiller F, Fort P. Selective activation of a few limbic structures during paradoxical (REM) sleep by the claustrum and the supramammillary nucleus: evidence and function. Curr Opin Neurobiol 2017; 44: 59–64. [DOI] [PubMed] [Google Scholar]
- Mazza S, Gerbier E, Gustin MP, Kasikci Z, Koenig O, Toppino TC et al. . Relearn faster and retain longer. Psychol Sci 2016; 27: 1321–30. [DOI] [PubMed] [Google Scholar]
- Menz MM, Rihm JS, Salari N, Born J, Kalisch R, Pape HC et al. . The role of sleep and sleep deprivation in consolidating fear memories. Neuroimage 2013; 75: 87–96. [DOI] [PubMed] [Google Scholar]
- Merica H, Blois R, Gaillard JM. Spectral characteristics of sleep EEG in chronic insomnia. Eur J Neurosci 1998; 10: 1826–34. [DOI] [PubMed] [Google Scholar]
- Morin CM, Belleville G, Belanger L, Ivers H. The Insomnia Severity Index: psychometric indicators to detect insomnia cases and evaluate treatment response. Sleep 2011; 34: 601–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin CM, Rodrigue S, Ivers H. Role of stress, arousal, and coping skills in primary insomnia. Psychosom Med 2003; 65: 259–67. [DOI] [PubMed] [Google Scholar]
- Nieuwenhuis IL, Takashima A. The role of the ventromedial prefrontal cortex in memory consolidation. Behav Brain Res 2011; 218: 325–34. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Ray RD, Cooper JC, Robertson ER, Chopra S, Gabrieli JD et al. . For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion. Neuroimage 2004; 23: 483–99. [DOI] [PubMed] [Google Scholar]
- Pace-Schott EF, Germain A, Milad MR. Sleep and REM sleep disturbance in the pathophysiology of PTSD: the role of extinction memory. Biol Mood Anxiety Disord 2015; 5: 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pace-Schott EF, Shepherd E, Spencer RM, Marcello M, Tucker M, Propper RE et al. . Napping promotes inter-session habituation to emotional stimuli. Neurobiol Learn Mem 2011; 95: 24–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Payne JD, Kensinger EA. Sleep leads to changes in the emotional memory trace: evidence from FMRI. J Cogn Neurosci 2011; 23: 1285–97. [DOI] [PubMed] [Google Scholar]
- Perlis ML, Smith MT, Andrews PJ, Orff H, Giles DE. Beta/gamma EEG activity in patients with primary and secondary insomnia and good sleeper controls. Sleep 2001; 24: 110–7. [DOI] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Moore GJ, Uhde TW, Tancer ME. Neural substrates for voluntary suppression of negative affect: a functional magnetic resonance imaging study. Biol Psychiatry 2005; 57: 210–9. [DOI] [PubMed] [Google Scholar]
- Phan KL, Taylor SF, Welsh RC, Decker LR, Noll DC, Nichols TE et al. . Activation of the medial prefrontal cortex and extended amygdala by individual ratings of emotional arousal: a fMRI study. Biol Psychiatry 2003; 53: 211–5. [DOI] [PubMed] [Google Scholar]
- Plante DT, Jensen JE, Schoerning L, Winkelman JW. Reduced gamma-aminobutyric acid in occipital and anterior cingulate cortices in primary insomnia: a link to major depressive disorder? Neuropsychopharmacology 2012; 37: 1548–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poe GR, Walsh CM, Bjorness TE. Cognitive neuroscience of sleep. Prog Brain Res 2010; 185: 1–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pruim RHR, Mennes M, van Rooij D, Llera A, Buitelaar JK, Beckmann CF. ICA-AROMA: a robust ICA-based strategy for removing motion artifacts from fMRI data. Neuroimage 2015; 112: 267–77. [DOI] [PubMed] [Google Scholar]
- Renouard L, Billwiller F, Ogawa K, Clement O, Camargo N, Abdelkarim M et al. . The supramammillary nucleus and the claustrum activate the cortex during REM sleep. Sci Adv 2015; 1: e1400177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riemann D, Spiegelhalder K, Feige B, Voderholzer U, Berger M, Perlis M et al. . The hyperarousal model of insomnia: a review of the concept and its evidence. Sleep Med Rev 2010; 14: 19–31. [DOI] [PubMed] [Google Scholar]
- Riemann D, Spiegelhalder K, Nissen C, Hirscher V, Baglioni C, Feige B. REM sleep instability—a new pathway for insomnia? Pharmacopsychiatry 2012; 45: 167–76. [DOI] [PubMed] [Google Scholar]
- Rosales-Lagarde A, Armony JL, Del Rio-Portilla Y, Trejo-Martinez D, Conde R, Corsi-Cabrera M. Enhanced emotional reactivity after selective REM sleep deprivation in humans: an fMRI study. Front Behav Neurosci 2012; 6: 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schalkwijk F. The conscience and self-conscious emotions in adolescence: an integrative approach. East Sussex, UK: Routledge; 2015. [Google Scholar]
- Seo J, Moore KN, Gazecki S, Bottary RM, Milad MR, Song H et al. . Delayed fear extinction in individuals with insomnia disorder. Sleep 2018; 41: zsy095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Standards of Practice Committee of the American Sleep Disorders Association. Practice parameters for the use of polysomnography in the evaluation of insomnia. Sleep 1995; 18: 55–7. [PubMed] [Google Scholar]
- Sterpenich V, Albouy G, Boly M, Vandewalle G, Darsaud A, Balteau E et al. . Sleep-related hippocampo-cortical interplay during emotional memory recollection. PLoS Biol 2007; 5: e282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sterpenich V, Albouy G, Darsaud A, Schmidt C, Vandewalle G, Dang Vu TT et al. . Sleep promotes the neural reorganization of remote emotional memory. J Neurosci 2009; 29: 5143–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuewig J, McCloskey LA. The relation of child maltreatment to shame and guilt among adolescents: psychological routes to depression and delinquency. Child Maltreat 2005; 10: 324–36. [DOI] [PubMed] [Google Scholar]
- Sturm VE, Sollberger M, Seeley WW, Rankin KP, Ascher EA, Rosen HJ et al. . Role of right pregenual anterior cingulate cortex in self-conscious emotional reactivity. Soc Cogn Affect Neurosci 2013; 8: 468–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swift KM, Gross BA, Frazer MA, Bauer DS, Clark KJD, Vazey EM et al. . Abnormal locus coeruleus sleep activity alters sleep signatures of memory consolidation and impairs place cell stability and spatial memory. Curr Biol 2018; 28: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tahmasian M, Noori K, Samea F, Zarei M, Spiegelhalder K, Eickhoff SB et al. . A lack of consistent brain alterations in insomnia disorder: An activation likelihood estimation meta-analysis. Sleep Med Rev 2018; 42: 111–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takashima A, Petersson KM, Rutters F, Tendolkar I, Jensen O, Zwarts MJ et al. . Declarative memory consolidation in humans: a prospective functional magnetic resonance imaging study. Proc Natl Acad Sci USA 2006; 103: 756–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tempesta D, Socci V, De Gennaro L, Ferrara M. Sleep and emotional processing. Sleep Med Rev 2018; 40: 183–95. [DOI] [PubMed] [Google Scholar]
- Urry HL, van Reekum CM, Johnstone T, Kalin NH, Thurow ME, Schaefer HS et al. . Amygdala and ventromedial prefrontal cortex are inversely coupled during regulation of negative affect and predict the diurnal pattern of cortisol secretion among older adults. J Neurosci 2006; 26: 4415–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Helm E, Yao J, Dutt S, Rao V, Saletin JM, Walker MP. REM sleep depotentiates amygdala activity to previous emotional experiences. Curr Biol 2011; 21: 2029–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van het Veer A, Hafkemeijer A, Van Steenbergen H, Bas-Hoogendam JM. Philips B0 Unwarping in FSL FEAT on Shark (or locally). 2017. www.osf.io/hks7x (3 December 2018, date last accessed).
- Vanderheyden WM, Poe GR, Liberzon I. Trauma exposure and sleep: using a rodent model to understand sleep function in PTSD. Exp Brain Res 2014; 232: 1575–84. [DOI] [PubMed] [Google Scholar]
- Vogt BA. Pain and emotion interactions in subregions of the cingulate gyrus. Nat Rev Neurosci 2005; 6: 533–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt BA, Finch DM, Olson CR. Functional heterogeneity in cingulate cortex: the anterior executive and posterior evaluative regions. Cereb Cortex 1992; 2: 435–43. [DOI] [PubMed] [Google Scholar]
- Wagner U, Fischer S, Born J. Changes in emotional responses to aversive pictures across periods rich in slow-wave sleep versus rapid eye movement sleep. Psychosom Med 2002; 64: 627–34. [DOI] [PubMed] [Google Scholar]
- Wagner U, Hallschmid M, Rasch B, Born J. Brief sleep after learning keeps emotional memories alive for years. Biol Psychiatry 2006; 60: 788–90. [DOI] [PubMed] [Google Scholar]
- Wagner U, N’Diaye K, Ethofer T, Vuilleumier P. Guilt-specific processing in the prefrontal cortex. Cereb Cortex 2011; 21: 2461–70. [DOI] [PubMed] [Google Scholar]
- Wassing R, Benjamins JS, Dekker K, Moens S, Spiegelhalder K, Feige B et al. . Slow dissolving of emotional distress contributes to hyperarousal. Proc Natl Acad Sci USA 2016; 113: 2538–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wassing R, Benjamins J, Schalkwijk F, Van Someren EJW. Overnight worsening of emotional distress indicates maladaptive sleep in insomnia. Sleep 2019; 42: zsy268. [DOI] [PubMed] [Google Scholar]
- Whalley MG, Kroes MC, Huntley Z, Rugg MD, Davis SW, Brewin CR. An fMRI investigation of posttraumatic flashbacks. Brain Cogn 2013; 81: 151–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wittchen HU, Jacobi F, Rehm J, Gustavsson A, Svensson M, Jonsson B et al. . The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur Neuropsychopharmacol 2011; 21: 655–79. [DOI] [PubMed] [Google Scholar]
- Woolrich MW, Behrens TE, Beckmann CF, Jenkinson M, Smith SM. Multilevel linear modelling for FMRI group analysis using Bayesian inference. Neuroimage 2004; 21: 1732–47. [DOI] [PubMed] [Google Scholar]
- Woolrich MW, Ripley BD, Brady M, Smith SM. Temporal autocorrelation in univariate linear modeling of FMRI data. Neuroimage 2001; 14: 1370–86. [DOI] [PubMed] [Google Scholar]
- Yoo SS, Gujar N, Hu P, Jolesz FA, Walker MP. The human emotional brain without sleep—a prefrontal amygdala disconnect. Curr Biol 2007; 17: R877–8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data and software/scripts used in this study are available on reasonable request from the corresponding author.