

Abstract
Recently, we showed that imidazole dipeptide such as carnosine contained abundantly in chicken breast meat improves brain function in a double-blind randomized controlled trial. However, the underlying molecular mechanisms remain unknown. Here, we investigated whether carnosine activates intestinal epithelial cells and induces the secretion of factors that activate brain function. We focused on exosomes derived from intestinal epithelial cells as mediators of brain-gut interaction. Results showed that exosomes derived from Caco-2 cells treated with carnosine significantly induced neurite growth in SH-SY5Y cells. To clarify the molecular basis of this finding, we performed integrated analysis of microRNAs (miRNAs) with altered expression in exosomes in response to carnosine treatment and mRNAs with altered expression in target cells in response to exosome treatment to identify related miRNAs and their target genes. The combination of miR-6769-5p and its target gene ATXN1 was found to be involved in the exosome-induced activation of neuronal cells.
Introduction
Carnosine, an imidazole dipeptide, has been reported to exert anti-fatigue, anti-oxidant, and exercise performance-enhancing effects [1,2]. Recently, we reported that carnosine and its related peptide anserine improved brain function, as evidenced by experiments using a mouse model of Alzheimer’s disease [3] and a double-blind randomized controlled trial [4]. The expression of inflammatory cytokine CCL24 was suppressed in peripheral blood mononuclear cells from elderly individuals who had ingested carnosine/anserine, suggesting that the latter is a key molecule responsible for the observed effect [5]. Notably, we further demonstrate that anserine/carnosine supplementation preserves brain structure and function in elderly individuals carrying APOE4 [6].
Although carnosine significantly improves brain function, the underlying molecular mechanisms remain unknown. As carnosine is degraded into β-alanine and l-histidine by carnosinase in serum [7], carnosine itself is not thought to be directly transported to the brain. Accordingly, we tested the alternative possibility that carnosine activates intestinal epithelial cells and induces the secretion of factors that penetrate the blood-brain barrier and activate neuronal cells. We have previously shown that carnosine activates intestinal epithelial cells [8], and suggested that this carnosine-induced activation of intestinal epithelial cells results in brain-gut interaction via the secretion of brain-derived neurotrophic factor [9]. In the present study, we targeted exosomes derived from intestinal epithelial cells as mediators that activate the brain-gut interaction. Further, we sought to clarify the involvement of exosomes in the carnosine-induced activation of the brain-gut interaction.
Materials and methods
Cell culture and reagents
We used a human colon cancer cell line, Caco-2, and a human neuroblastoma cell line, SH-SY5Y. Both cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Nissui, Tokyo, Japan) supplemented with 10% fetal bovine serum (FBS; Life Technologies, Gaithersburg, MD, USA) at 37°C in 5% CO2. Primary mouse cortex neurons (Thermo Fisher Scientific K.K., Tokyo, Japan) were cultured in complete neurobasal medium supplemented with GlutaMax-I and B-27 (Thermo Fisher Scientific K.K.) according to the instruction. Carnosine was purchased from Wako (Osaka, Japan).
Exosome isolation
ExoQuick Solution (System Biosciences Inc., Mountain View, CA, USA) and MagCapture Exosome Isolation Kit PS (Wako) were used to isolate exosomes from supernatant of Caco-2 cells treated with 1 or 10 mM carnosine, according to the manufacturer’s instructions [10]. The concentration of carnosine to treat Caco-2 cells was determined by referring to the previous reports [11].
Exosome ELISA
Purification of exosomes was confirmed by PS Capture Exosome ELISA Kit (Wako) according to the manufacturer’s instructions. Antibodies used as capture reagents were as follows: anti-CD63 contained in the above kit, and anti-CD81, anti-CD9 and anti-Hsp70 contained in the EXOAB-KIT-1 (System Biosciences). For detection of the latter three antigens, we used a goat anti-rabbit IgG HRP-conjugated secondary antibody.
Incorporation of exosome into SH-SY5Y cells
Exosomes (200 ng) were labeled with 2 μM PKH26 using the PKH26 Red Fluorescence Cell Linker Kit (Sigma-Aldrich, St. Louis, MO, USA) for 1 hr at 37°C. After labeling exosomes with PKH26, free PKH26 was removed by ultrafiltration using Vivacon 500 (100,000 MWCO; Sartorius Stedim Biotech GmbH, Goettingen, Germany). Labeled exosomes (200 mg/well) were added to SH-SY5Y cells (2 × 104 cells/well) in a 96-well plate (96 well SensoPlate Plus Microplate, Glass bottom, Greiner Bio-one, Tokyo, Japan), and observed under a confocal microscope (TCS SP8, Leica Microsystems K.K., Tokyo, Japan).
Quantitative evaluation of neurite growth
After seeding onto a μClear Fluorescence Black Plate (Greiner-Bio One), SH-SY5Y cells were treated with exosomes for 24 h. SH-SY5Y cells were fixed with 4% paraformaldehyde for 15 min, blocked with blocking buffer (1 × PBS, 5% goat serum, 0.3% Triton X-100) for 1 h, and treated with Neuro-Chrom Pan Neuronal Marker (Merck Millipore, Billerica, MA, USA) at 25°C overnight. After washing with PBS, cells were stained with Alexa Fluor 555 F(ab′)2 fragment of goat antibody IgG (H+L) (Thermo Fisher Scientific, Waltham, MA, USA) for 1 h. After washing with PBS, cells were further stained with Hoechst 33342 (Dojindo, Kumamoto, Japan) for 15 min, and neurite length was measured using the IN Cell Analyzer 1000 (GE Healthcare, Amersham Place, UK).
Quantitative reverse transcription polymerase chain reaction
RNA was prepared using the High Pure RNA Isolation kit (Roche Diagnostics GmbH, Mannheim, Germany), and cDNA was prepared using the SuperScript IV Reverse Transcriptase (Thermo Fisher Scientific), as described previously [12]. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed using the Thunderbird SYBR qPCR mix (Toyobo, Osaka, Japan) and Thermal Cycler Dice Real Time System TP-800 (Takara, Shiga, Japan). Samples were analyzed in triplicate, and gene expression levels were normalized to the corresponding glyceraldehyde-3-phosphate dehydrogenase (GAPDH) level. PCR primer sequences are shown in S1 Table.
miRNA microarray assay
Microarray assay of miRNA contained in exosomes derived from non-treated or carnosine-treated Caco-2 cells was performed. Total RNA was isolated from exosomes using TRIzol Reagent (Thermo Fisher Scientific) and purified using miRNeasy Mini Kit (Qiagen, Valencia, CA). Total RNA (100 ng) of each sample was labeled using FlashTag Biotin HSR RNA Labeling Kit (Affymetrix, Santa Clara, CA, USA), and hybridized to a Affimetrix GeneChip miRNA 4.0 Array. Relative hybridization intensities and background hybridization values were calculated using the Affymetrix Expression Console. We processed the raw CEL files for gene-level analysis with median polish summarization and quantile normalization by Affymetrix Transcriptome Analysis Console Software, and obtained normalized intensity values. To identify up- or down-regulated genes, we calculated ratios (non-log scaled fold-change) from the normalized intensities of each gene for comparisons between control and experimental samples. Then, we established criteria for regulated genes: (up-regulated genes) ratio ≥ 1.3-fold, (down-regulated genes) ratio ≤ 0.77 [13]. Target genes for miRNA were predicted using TargetScan (http://www.targetscan.org/vert_71/). To determine significantly over-represented gene ontology (GO) categories and significantly enriched pathways, we used tools and data provided by the Database for Annotation, Visualization, and Integrated Discovery (DAVID, http://david.abcc.ncifcrf.gov) [14,15].
mRNA microarray assay
Total RNA was isolated from SH-SY5Y cells treated with exosomes derived from non-treated or carnosine-treated Caco-2 cells using Isogen II (Nippon Gene, Tokyo, Japan). The RNA quality was tested as described previously [9]. cRNA was labeled with Low Input Quick Amp Labeling (Agilent Technologies, Santa Clara, CA, USA), and hybridized to DNA microarray (SurePrint G3 Human Gene Expression 8x60K v3, Agilent). Relative hybridization intensities and background hybridization values were calculated using Feature Extraction Software (Agilent).
We identified genes with altered expression, as described previously [9]. We then established criteria for significantly up- or down-regulated genes: up-regulated genes, Z-score ≥ 2.0 and ratio ≥ 1.5-fold; down-regulated genes: Z-score ≤ -2.0 and ratio ≤ 0.66-fold. Significantly over-represented GO categories and enriched pathways were determined as described above. Heat map was generated using MeV software [16], and a hierarchical clustering method was used to sort the genes. Color indicates the distance from the median of each row.
Pathway analysis
We performed pathway analysis by subjecting selected genes to Ingenuity Pathway Analysis (IPA, Qiagen, Redwood City, CA, USA). Using the Ingenuity Knowledge Base, known functional pathways were tested for enrichment with selected genes. Selected genes were assigned to known functional networks based on canonical pathways, relationship to upstream regulators, molecular and cellular functional grouping, and associated network functions. In IPA, an assignment of nodes in gene network uses published observations stored in the Ingenuity Pathway Knowledge Base.
To obtain additional insight into gene functions, selected genes were examined for their potential role in neuronal cells using NCBI gene (www.ncbi.nlm.nih.gov/gene).
Expression analysis of miRNA
Exosomes were prepared from supernatant of Caco-2 cells treated with carnosine using the MagCapture Exosome Isolation Kit PS, as described above. Total RNA in the exosomes was isolated using a miRNeasy Serum/Plasma Kit (Qiagen) according to the manufacturer’s protocol. Cellular total RNA was isolated as described above. cDNA against miRNA was synthesized using the miScript II Kit (Qiagen). Primer sequences to amplify miRNA (miR-6769b-5p, 5′-UGGUGGGUGGGGAGGAGAAGUGC-3′; miR-24-3p, 5′-UGGCUCAGUUCAGCAGGAACAG-3′; miR-937-5p, 5′-GUGAGUCAGGGUGGGGCUGG-3′) were designed by Qiagen. qRT-PCR was performed using the miScript SYBR Green PCR Kit (Qiagen) according to the manufacturer’s protocol. The determination of miRNA levels in each sample was performed by the 2–ΔΔCt method [17].
Transfection of miR mimic and siRNA
The miR-6769b-5p mimic (UGGUGGGUGGGGAGGAGAAGUGC, MSY0027620, Qiagen) and siRNA against ATXN1 (siATXN1_8: CCCTTCTACCCTCAACGACAA, SI04317964, Qiagen; siATXN1_9: AAGCAACGACCTGAAGATCGA, SI04332027, Qiagen) were transfected into SH-SY5Y cells using the HyPerFect Transfection Reagent (Qiagen). Transfection conditions were determined using AllStar Hs Cell Death Control siRNA (Qiagen). After transfection, the expression of miR6769b-5p and ATXN1 mRNA was determined by qRT-PCR, and the neurite length was quantitatively evaluated as described above.
Animal experiments
C57/BL6J mice (6 weeks old, male, n = 3) were orally administered carnosine (2 g/L) in drinking water for 7 days. Exosome was prepared from serum by using MagCapture Exosome Isolation Kit PS as described previously. All mouse experiments and protocols were in accordance with the Guide for the Care and Use of Laboratory Animals and were approved by the Ethics Committees on Animal Experimentation (Kyushu University; approved number: A30-244-0).
Statistical analysis
All experiments were performed at least 3 times, and the corresponding data are shown. The results are presented as mean ± standard deviation. Statistical significance was determined using a two-sided Student’s t-test. Statistical significance was defied as P<0.05 (*P<0.05; **P<0.01; ***P<0.001).
Results
Isolation of exosomes from the supernatant of carnosine-treated Caco-2 cells
Exosomes were isolated from the supernatant of carnosine-treated Caco-2 cells using the MagCapture Exosome Isolation Kit PS. We determined the particle size of isolated exosomes by dynamic light scattering using ELSZ-0S (Otsuka Electronics, Osaka, Japan). Results showed that Caco-2 cells produced exosomes of about 50 nm in diameter, regardless of carnosine treatment (Fig 1A–1C). Furthermore, we tested for the expression of exosome marker proteins using the PS Capture Exosome ELISA Kit. Results showed that all exosomes, regardless of carnosine treatment, expressed exosome marker proteins at almost the same level, although the expression levels of CD63 and CD9 were relatively low (Fig 1D). These results demonstrate that Caco-2 cells produce exosomes of the same size that express marker proteins at almost the same level, regardless of carnosine treatment.
Fig 1. Exosomes isolated from supernatant of Caco-2 cells.

Particle sizes of isolated exosomes derived from non-treated Caco-2 cells (A) and those of Caco-2 cells treated with 1 mM carnosine (B) and 10 mM carnosine (C) were determined by dynamic light scattering using ELSZ-0S (Otsuka Electronics). D, Expression of marker proteins of exosome was determined using the PS Capture Exosome ELISA kit (Wako).
Incorporation of exosomes into SH-SY5Y cells
In the present study, we speculated that carnosine activates the brain-gut interaction via exosomes produced from carnosine-treated intestinal cells. Firstly, we investigated the incorporation of Caco-2-derived exosomes into SH-SY5Y cells. As shown in S1 Fig, Caco-2-derived exosomes labeled with PKH26 were incorporated into SH-SY5Y cells. No significant difference in the incorporation efficiency of exosomes was observed between Caco-2 cell treatments.
Activation of SH-SY5Y cells by treatment with Caco-2-derived exosomes
As shown above, Caco-2-derived exosomes could be incorporated into SH-SY5Y cells. Here, we attempted to evaluate whether Caco-2-derived exosomes induce neurite growth in SH-SY5Y cells. Firstly, we optimized the conditions for neurite growth in SH-SY5Y cells induced by exosomes isolated from the supernatant of carnosine-treated Caco-2 cells. Then, we treated SH-SY5Y cells with exosomes equivalent to 90 ng protein for 1 day, and quantitatively evaluated neurite growth using the IN Cell Analyzer 1000. As shown in Fig 2A, exosomes derived from the supernatant of Caco-2 cells treated with 10 mM carnosine (Exo-Car10) significantly induced neurite growth in SH-SY5Y cells compared with exosomes derived from the supernatant of non-treated Caco-2 cells (Exo-Ctrl). The length and diameter of neurites in SH-SY5Y cells were significantly augmented by treatment with Exo-Car10; this effect was equivalent to that observed when SH-SY5Y cells were treated with retinoic acid (Fig 2A and 2B).
Fig 2. Activation of neuronal cells by Caco-2-derived exosome.
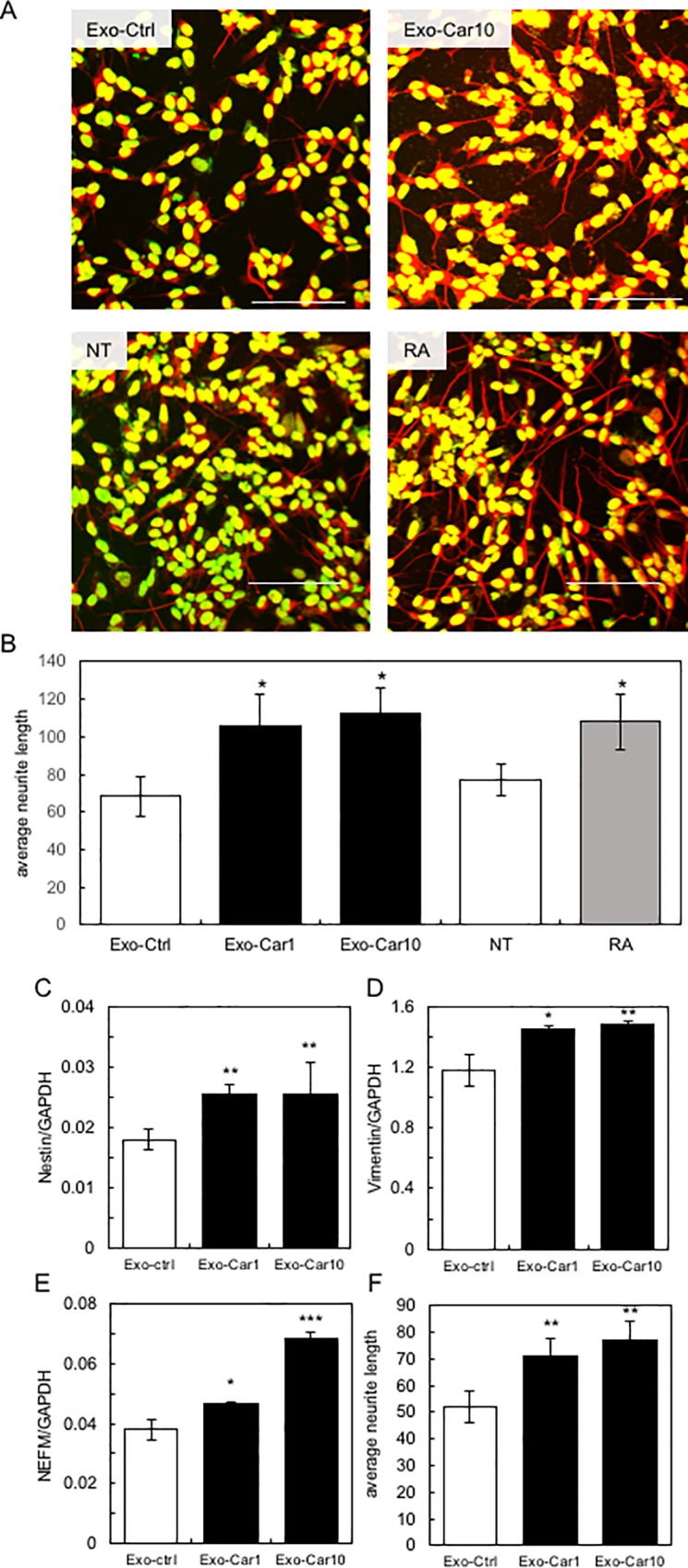
A, Neurite growth in SH-SY5Y cells treated with exosomes derived from non-treated Caco-2 cells (Exo-ctrl), from Caco-2 cells treated with 10 mM carnosine (Exo-Car10), and neurite growth in non-treated SH-SY5Y cells (NT) and cells treated with retinoic acid (RA) were shown. B, Neurite growth in SH-SY5Y cells was quantitatively determined using the IN Cell Analyzer 1000. C-E, Expression of neuronal cell marker genes of nestin (C), vimentin (D) and neurofilament medium (NEFM; E) was quantitatively determined by qRT-PCR. Neurite growth in primary mouse cortex neurons treated with exosomes (Exo-ctrl, Exo-Car1 and Exo-Car10) was quantitatively determined using the IN Cell Analyzer 1000 as described above (F).
Next, we tested for the expression of neuronal cell marker genes of nestin, vimentin, and neurofilament medium (NEFM). Results showed that Exo-Car1 and Exo-Car10 significantly augmented the expression of these marker genes (Fig 2C–2E). These results indicate that Exo-Car activates and induces neurite growth in SH-SY5Y cells. Furthermore, Exo-Car induced neurite growth in primary mouse cortex neurons as well as neuronal cell line, SH-SY5Y (Fig 2F), suggesting that Exo-Car activates neurons.
Expression profile analysis of exosome miRNA
We next tried to clarify the molecular mechanisms of Exo-Car to activate SH-SY5Y cells, namely miRNA in exosome and its target genes. Firstly, we searched for miRNA with altered expression in Exo-Car and its target genes. After Caco-2 cells were treated with 1 mM or 10 mM carnosine, exosomes were isolated from the supernatant using ExoQuick Solution, as described above. Changes in the expression profile of exosome miRNA in response to carnosine treatment were analyzed using the Affimetrix GeneChip miRNA 4.0 array. The expression of numerous miRNAs was altered in exosomes via treatment with 1 mM or 10 mM carnosine (Fig 3). The expression of 27 miRNAs and 19 miRNAs were altered in Caco-2 cells treated with 1 mM and 10 mM carnosine, respectively. Among these miRNAs, 7 miRNAs (miR-103a-3p, miR-194-5p, miR-619-5p, miR-6732-5p, miR-6769b-5p, miR-6771-5p, and miR-6775-5p) were similarly regulated in Caco-2 cells treated with 1 mM and 10 mM carnosine.
Fig 3. Expression profile analysis of exosome miRNA.

After Caco-2 cells were treated with 1 mM or 10 mM carnosine, isolated exosomes were used for miRNA microarray analysis with an Affimetrix GeneChip miRNA 4.0 array. miRNAs with altered expression in response to carnosine treatment are shown.
Next, we predicted target genes for these miRNAs using TargetScan, and determined significantly over-represented GO categories and significantly enriched pathways using DAVID. Here, we performed cluster analysis of these target genes to predict the function of miRNA. Results showed that many target genes of the selected miRNA were involved in the development and differentiation of neuronal cells, axon guidance, and neurotrophin signaling pathway, suggesting that these miRNAs contained in the carnosine-treated exosomes activate neuronal cells via their target genes (S2 Table).
Expression profile analysis of mRNA in the Exo-Car-treated SH-SY5Y cells
Next, in order to identify target genes of Exo-Car in SH-SY5Y cells, we analyzed the expression profile of mRNA in SH-SY5Y cells treated with exosomes. After Caco-2 cells were treated with 1 mM and 10 mM carnosine, exosomes (Exo-Car1 and Exo-Car10) were isolated as described previously. Total RNA was prepared from SH-SY5Y cells treated with these exosomes and with RA as a positive control, and used for DNA microarray analysis. Compared with SH-SY5Y cells treated with exosomes isolated from non-treated Caco-2 cells (Exo-Ctrl), 585 genes and 579 genes showed altered expression in SH-SY5Y cells treated with Exo-Car1 and Exo-Car10, respectively. Furthermore, as shown in the heat map (S2 Fig), many genes showed similar expression profiles between SH-SY5Y cells treated with Exo-Car1, Exo-Car10, and RA. We speculate that genes with altered expressions in Exo-Car-treated SH-SY5Y cells are under the control of miRNA with altered expression in Exo-Car, and involved in the activation of SH-SY5Y cells.
Integrated analysis
We then performed integrated analysis of miRNAs with altered expression in exosomes in response to carnosine treatment and of mRNAs with altered expression in SH-SY5Y cells in response to Exo-Car treatment. Further, we attempted to identify miRNAs and their target genes involved in the activation of SH-SY5Y cells.
Firstly, we compared target genes of miRNA with altered expression in Exo-Car to mRNAs with altered expression in Exo-Car-treated SH-SY5Y, and identified 137 and 77 typical genes for 1 mM and 10 mM carnosine treatment, respectively. We then selected 14 genes that have been reported to activate and/or maintain neuronal cells using the NCBI database (https://www.ncbi.nlm.nih.gov/pubmed/) (Table 1). Some of other genes were related to the activation of immune cells and maintenance of adipose tissue. Next, we analyzed the interaction network of these genes by in silico analysis (Ingenuity Pathway Analysis (IPA)) to select possible genes of significance and their regulating miRNAs (Table 2, S3 Fig).
Table 1. Neuronal gene and its regulating miRNA selected by integration analysis.
| Gene name | Exo-Car1 (Z score) |
Exo-Car10 (Z score) |
Regulatory miRNA | Function |
|---|---|---|---|---|
| ATXN1 | -16.404 | -15.972 | miR-6769b-5p | neurodegenerative disorders |
| miR-6794-5p | ||||
| SLITRK5 | -13.012 | -12.765 | miR-6769b-5p | neurite-modulating activity |
| miR-3196 | ||||
| miR-4507 | ||||
| ATCAY | 1.609 | – | miR-24-3p | neuron-restricted protein |
| miR-619-5p | ||||
| miR-937-5p | ||||
| SNCAIP | 6.362 | – | miR-24-3p | interacts with α-synuclein in neuronal tissue |
| CPLX2 | 5.243 | – | miR-194-5p | function in synaptic vesicle exocytosis |
| miR-619-5p | ||||
| NRSN1 | 2.599 | – | miR-4668 | transduction of nerve signals |
| KIRREL3 | -8.776 | – | miR-6769b-5p | expressed in adult brain |
| KCNG4 | 12.3 | – | miR-24-3p | regulating neurotransmitter release |
| miR-4668 | ||||
| miR-619-5p | ||||
| NOTCH1 | 1.66 | – | miR-24-3p | receptor for neuronal DNER |
| NRXN1 | 2.062 | – | miR-3162 | the neurexin family |
| PHOX2A | 1.857 | – | miR-3162 | role in development of nervous system |
| AVP | -1.62 | – | miR-3196 | transported axonally to the nerve endings |
| LHX3 | 1.626 | – | miR-4532 | development and motor neuron specification |
| VAMP4 | – | 1.663 | miR-103a-3p | support distinct forms of neurotransmission |
Table 2. Selected miRNA and target gene.
| miRNA | Target gene |
|---|---|
| miR-6769b-5p | ATXN1, KIRREL3, SLITRK5 |
| miR-619-5p | ATCAY, CPLX2, KCNG4 |
| miR-24-3p | ATCAY, KCNG4, NOTCH1, SNCAIP |
| miR-937-5p | ATCAY |
Expression profile of miRNA
Firstly, we tested for changes in expression of the selected miRNA (miR-6769b-5p, miR-24-3p, miR-937-5p, and miR-619-5p) in Caco-2 cells treated with carnosine. The expression of miR-6769b-5p was up-regulated, while that of miR-24-3p and miR-937-5p was down-regulated in Caco-2 cell treated with carnosine; however, that of miR-619-5p remained unchanged (Fig 4A, 4D and 4G).
Fig 4. Expression profile of miRNA.

The changes in the expression of the selected miRNA (miR-6769b-5p, miR-24-3p, and miR-937-5p) in Caco-2 cells treated with carnosine are shown in A, D, and G, respectively. The changes in the expression of miRNA in cells treated with Exo-Car are shown in B (miR-6769b-5p), E (miR-24-3p) and H (miR-937-5p). The changes in the expression of miRNA (C, miR-6769b-5p; F, miR-24-3p; I, miR-937-5p) in SH-SY5Y cells treated with Exo-Car are shown.
Next, we tested for changes in expression of these miRNAs in Exo-Car-treated cells. After preparing the Exo-Car, we quantitatively determined the expression level of miRNAs. Consistent with the results above (Fig 4A, 4D and 4G), the expression of miR-6769b-5p was up-regulated, and that of miR-24-3p and miR-937-5p was down-regulated in Exo-Car (Fig 4B, 4E and 4H). The change in the expression of miR-619-5p in Exo-Car was not significant (S4 Fig).
Finally, we tested for changes in the expression of these miRNAs in SH-SY5Y cells treated with Exo-Car. After preparing Exo-Car from the supernatant of carnosine-treated Caco-2 cells, SH-SY5Y cells were treated with the Exo-Car and changes in the expression of these miRNA in SH-SY5Y cells were quantitatively determined by qRT-PCR. Results demonstrated that the expression of miR-6769b-5p was up-regulated, and that of miR-24-3p and miR-937-5p was down-regulated in SH-SY5Y cells treated with Exo-Car (Fig 4C, 4F and 4I). The expression of miR-619-5p remained unchanged in SH-SY5Y cells even after treatment with Exo-Car.
Expression profile of target genes in Exo-Car-treated SH-SY5Y cells
Next, we selected several genes among target genes of miRNA (Tables 1 & 2) and analyzed the expression profiles of these genes in Exo-Car-treated SH-SY5Y cells (Fig 5). Among these target genes, ATXN1 and SLITRK5 were significantly down-regulated in Exo-Car-treated SH-SY5Y cells (Fig 5A and 5B). The expression of ATCAY and SNCAIP was up-regulated only in Exo-Car1-treated cells (Fig 5C and 5D). Thus, we focused on miR-6769b-5p, which regulates the expression of ATXN1 and SLITRK5.
Fig 5. Expression profile of target genes in Exo-Car-treated SH-SY5Y cells.

The expression profile of target genes of miRNA in Exo-Car-treated SH-SY5Y cells, as determined by qRT-PCR, is shown (A, ATXN1; B, SLITRK5; C, ATCAY; D, SNCAIP).
The involvement of miR-6769b-6-5p and ATXN1 in neurite growth in SH-SY5Y cells
To evaluate the role of miR-6769b-5p in the regulation of ATXN1 expression and neurite growth in SH-SY5Y cells, we used miRNA mimics with similar functions to miR-6769b-5p. As shown in Fig 6A–6C, the miR-6769b-5p mimic was efficiently transduced into the SH-SY5Y cells, resulting in the significant down-regulation of the expression of ATXN1 and induction of neurite growth in SH-SY5Y cells. These results suggest that the decreased expression of ATXN1 induces neurite growth in SH-SY5Y cells. To confirm this, we used siRNA targeting ATXN1 (siATXN1_8 and siATXN1-_9). Results clearly showed that siATXN1s efficiently down-regulated the expression of ATXN1 in SH-SY5Y cells (Fig 6D), which significantly induced neurite growth in SH-SY5Y cells (Fig 6E). These results suggest that decreased expression of ATXN1 induced by miR-6769b-5p contained in Exo-Car triggered neurite growth in SH-SY5Y cells.
Fig 6. Involvement of miR-6769b-5p and ATXN1 in neurite growth in SH-SY5Y cells.

The expression of miR-6769b-5p (A) and ATXN1 (B), and neurite growth (C) in SH-SY5Y cells transduced with miR-6769b-5p mimic was determined by qRT-PCR and by using the IN Cell Analyzer 1000, respectively. D, The expression of ATXN1 in SH-SY5Y cells transduced with siRNA targeting ATXN1 (siATXN1_8 and siATXN1_9) were determined by qRT-PCR. E, Neurite growth in SH-SY5Y cells transduced with siATXN1_8 and siATXN1_9 was determined using the IN Cell Analyzer 1000.
Finally, we tested whether serum exosome in mice orally administered carnosine can activate neuronal cells. As shown in Fig 7, serum exosome derived from mice orally administered carnosine significantly extended neurite in SH-SY5Y cells, suggesting that carnosine intake induces exosome that activates neuronal cells in serum. We would like to clarify the molecular mechanisms for generation and functionalities of the carnosine-induced serum exosome in the future study.
Fig 7. Serum exosome of mice orally administered with carnosine activates neuronal cells.

Exosome was prepared from mice orally administered drinking water containing carnosine (B) or not (A) by using MagCapture Exosome Isolation Kit PS as decribed previously. Neurite growth in SH-SY5Y cells treated with these exosomes was quantitatively determined using the IN Cell Analyzer 1000.
Discussion
Numerous food components have been reported to possess the ability to affect cognitive function and emotion; these include omega-3 fatty acids, curcumin, flavonoids, and resveratrol [18]. Several gut hormones and regulators of synaptic plasticity, such as brain-derived neurotrophic factor, have been reported to affect brain function. In this study, we presented a novel mechanism of action of such a “brain food”, namely carnosine. Until now, we showed that carnosine induces the secretion of several cytokines including BDNF, which would result in the activation of brain-gut interaction and its consequent brain function [9]. Here, we demonstrated that carnosine induced the secretion of exosomes from intestinal epithelial cells, and that the resulting exosomes and its miRNA activated neuronal cells, resulting in the activation of brain function.
Exosomes have received much interest since their key roles in intercellular/intertissue communication has been emerged. Numerous studies have focused on the exosome as a transporter of distinct bioactive molecules such as miRNA, mRNA, and protein, as well as on its therapeutic applications. In particular, exosomes derived from food are of interest owing to their ability to modulate cellular functions. Exosomes are known to be contained in numerous foods including fruits, milk, nuts, seeds, and roots [19]. However, there are a limited number of reports describing the functionality of exosomes whose secretion is induced by food factors. Curcumin has been reported to ameliorate endolysosomal cholesterol accumulation by shuttling cholesterol out of the cells via exosome secretion [20]. However, the involvement of exosomes in the food-induced activation of brain-gut interactions, which is speculated to represent an important molecular mechanism underlying the effects of diet on brain function, has not been reported to date.
In the present study, we sought to elucidate the molecular basis of exosome-mediated activation of neuronal cells by integrated analysis of target genes of differentially expressed miRNAs in exosomes derived from carnosine-treated Caco-2 cells and of differentially expressed mRNAs in cells treated with these exosomes. This integrated analysis, subsequent database screening for genes that activate and maintain neuronal cells, and pathway analysis using IPA enabled the successful identification of target genes and their regulatory miRNAs. Enerly et al. described roles for miRNAs in primary breast tumors by miRNA-mRNA integrated analysis in a previous study [21]; however, to our knowledge, the present study is the first to identify both miRNAs in exosomes and target mRNAs in recipient cells of exosomes via integrated analyses.
To date, studies have revealed that the miR-17-92 cluster and miR-124 function to elongate neurites [22,23]. Here, we identified a novel miRNA, miR-6769-5p, that activates and elongates neurites in SH-SY5Y cells. Furthermore, we identified ATXN1 as one of target genes of miR-6769-5p. ATXN1 has been reported to be involved in the pathophysiology of cerebellar seizures [24,25], and further that the silencing of ATXN1 mRNA provides therapeutic benefits in a mouse model of polyglutamine expansion diseases [26]. In this study, we found that miR-6769-5p-mediated reduction of ATXN1 expression led to the extension of neurites in neuronal cells and possibly to the activation of brain-gut interaction mediated by carnosine-induced exosomes. MiR-6967-5p and ATXN1 may therefore represent key factors involved in the activation of brain-gut interaction mediated by specific food components. Next, we need to clarify whether other brain foods activate brain-gut interaction via miR-6769-5p and ATXN1. Furthermore, it would become evident in the future study that various intercellular/intertissue interactions elicited by food components are mediated by functional exosomes.
Supporting information
The incorporation of exosomes labeled with PKH26 (red arrow) was observed under a confocal microscope (TCS SP8, Leica Microsystems).
(TIFF)
Total RNA from SH-SY5Y cells treated with exosomes derived from non-treated Caco-2 cells or Caco-2 cells treated with 1 mM or 10 mM carnosine were used for DNA microarray analysis with the SurePrint G3 Human Gene Expression 8x60K v3 array. The heat map was shown.
(TIFF)
Representative interaction network of target genes of miRNA with altered expression in Exo-Car and mRNAs with altered expression in Exo-Car-treated SH-SY5Y revealed by in silico analysis.
(TIFF)
The changes in the expression of miR-619-5p in Caco-2 cells treated with carnosine (A). The changes in the expression of miR-619-5p in cells treated with Exo-Car are shown in B. The changes in the expression of miR-619-5p in SH-SY5Y cells treated with Exo-Car are shown in C.
(TIFF)
(PDF)
(PDF)
Acknowledgments
The authors would like to thank N. Oshima (GE Healthcare) for her expert assistance with the IN Cell Analyzer 1000, Y. Okugawa (Center for Advanced Instrumental and Educational Supports, Faculty of Agriculture, Kyushu University) for her assistance with TCS SP8 (Leica Microsystems), and K. Yasuda (Cell Innovator, Fukuoka, Japan) for her assistance with microarray analysis.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
YK awarded JSPS KAKENHI (Japan Society for the Promotion of Science, https://www.jsps.go.jp/, Grant Number: 16K14929). YK received the collaborative research cost from NH Foods Ltd. NH Foods Ltd provided support in the form of salaries for authors [MS, TH], but did not have any additional role in the study design, data collection, and analysis, decision to publish, or preparation of the manuscript. The specific roles of these authors are articulated in the 'author contribution' section.
References
- 1.Boldyrev AA, Aldini G, Derave W. Physiology and pathophysiology of carnosine. Physiological Reviews. 2013;93: 1803–1845. 10.1152/physrev.00039.2012 [DOI] [PubMed] [Google Scholar]
- 2.Sale C, Saunders B, Harris RC. Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids. 2010;39: 321–333. 10.1007/s00726-009-0443-4 [DOI] [PubMed] [Google Scholar]
- 3.Herculano B, Tamura M, Ohba A, Shimatani M, Kutsuna N, Hisatsune T. β-alanyl-L-histidine rescues cognitive deficits caused by feeding a high fat diet in a transgenic mouse model of Alzheimer's disease. J Alzheimers Dis. 2013;33: 983–997. 10.3233/JAD-2012-121324 [DOI] [PubMed] [Google Scholar]
- 4.Hisatsune T, Kaneko J, Kurashige H, Cao Y, Satsu H, Totsuka M, et al. Effect of anserine/carnosine supplementation on verbal episodic memory in elderly people. J Alzheimers Dis. 2015;50: 149–159. 10.3233/JAD-150767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Katakura Y, Totsuka M, Imabayashi E, Matsuda H, Hisatsune T. Anserine/carnosine supplementation suppresses the expression of the inflammatory chemokine CCL24 in peripheral blood mononuclear cells from elderly people. Nutrients. 2017;9: 1199 10.3390/nu9111199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ding Q, Tanigawa K, Kaneko J, Totsuka M, Katakura Y, Imabayashi E, et al. Anserine/carnosine supplementation preserves blood flow in the prefrontal brain of elderly people carrying APOE e4. Aging and Disease. 2018;9: 334–345. 10.14336/AD.2017.0809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bellia F, Vecchio G, Rizzarelli E. Carnosinases, their substrates and diseases. Molecules. 2014;19: 2299–2329. 10.3390/molecules19022299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fujii K, Abe K, Kadooka K, Matsumoto T, Katakura Y. Carnosine activates the CREB pathway in Caco-2 cells. Cytotechnology. 2017;69: 523–527. 10.1007/s10616-017-0089-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kadooka K, Fujii K, Matsumoto T, Sato M, Morimatsu F, Tashiro K, et al. Mechanisms and consequences of carnosine-induced activation of intestinal epithelial cells. Journal of Functional Foods. 2015;13: 32–37. 10.1016/j.jff.2014.12.024 [DOI] [Google Scholar]
- 10.Nakai W, Yoshida T, Diez D, Miyatake Y, Nishibu T, Imawaka N, et al. A novel affinity-based method for the isolation of highly purified extracellular vesicles. Sci Rep. 2016;6: 33935 10.1038/srep33935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Son DO, Satsu H, Kiso Y, Totsuka M, Shimizu M. Inhibitory effect of carnosine on interleukin-8 production in intestinal epithelial cells through translation regulation. Cytokine. 2008; 42: 265–276. 10.1016/j.cyto.2008.02.011 [DOI] [PubMed] [Google Scholar]
- 12.Yamashita S, Ogawa K, Ikei T, Fujiki T, Katakura Y. FOXO3a potentiates hTERT gene expression by activating c-MYC and extends the replicative life-span of human fibroblast. PLoS ONE. 2014;9: e101864 10.1371/journal.pone.0101864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bolstad BM, Irizarry RA, Åstrand M, Speed TP. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics. 2003;19: 185–193. 10.1093/bioinformatics/19.2.185 [DOI] [PubMed] [Google Scholar]
- 14.Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucl Acids Res. 2009;37: 1–13. 10.1093/nar/gkn923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4: 44–57. 10.1038/nprot.2008.211 [DOI] [PubMed] [Google Scholar]
- 16.Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, et al. TM4: a free, open-source system for microarray data management and analysis. BioTechniques. 2003;34: 374–378. 10.2144/03342mt01 [DOI] [PubMed] [Google Scholar]
- 17.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25: 402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 18.Gómez-Pinilla F. Brain foods: the effects of nutrients on brain function. Nature Reviews Neuroscience. 2008;9: 568–578. 10.1038/nrn2421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pérez-Bermúdez P, Blesa J, Soriano JM, Marcilla A. Extracellular vesicles in food: Experimental evidence of their secretion in grape fruits. Eur J Pharm Sci. 2017;98: 40–50. 10.1016/j.ejps.2016.09.022 [DOI] [PubMed] [Google Scholar]
- 20.Canfrán Duque A, Pastor Ó, Quintana Portillo R, Lerma M, Peña G, Martín Hidalgo A, et al. Curcumin promotes exosomes/microvesicles secretion that attenuates lysosomal cholesterol traffic impairment. Mol Nutr Food Res. 2014;58: 687–697. 10.1002/mnfr.201300350 [DOI] [PubMed] [Google Scholar]
- 21.Enerly E, Steinfeld I, Kleivi K, Leivonen S-K, Aure MR, Russnes HG, et al. miRNA-mRNA integrated analysis reveals roles for miRNAs in primary breast tumors. PLoS ONE. 2011;6: e16915 10.1371/journal.pone.0016915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang Y, Ueno Y, Liu XS, Buller B, Wang X, Chopp M, et al. The MicroRNA-17-92 cluster enhances axonal outgrowth in embryonic cortical neurons. J Neurosci. 2013;33: 6885–6894. 10.1523/JNEUROSCI.5180-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yu J-Y, Chung K-H, Deo M, Thompson RC, Turner DL. MicroRNA miR-124 regulates neurite outgrowth during neuronal differentiation. Exp Cell Res. 2008;314: 2618–2633. 10.1016/j.yexcr.2008.06.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ingram M, Wozniak EAL, Duvick L, Yang R, Bergmann P, Carson R, et al. Cerebellar transcriptome profiles of ATXN1 transgenic mice reveal SCA1 disease progression and protection pathways. Neuron. 2016;89: 1194–1207. 10.1016/j.neuron.2016.02.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Asher M, Johnson A, Zecevic B, Pease D, Cvetanovic M. Ataxin-1 regulates proliferation of hippocampal neural precursors. Neuroscience. 2016;322: 54–65. 10.1016/j.neuroscience.2016.02.011 [DOI] [PubMed] [Google Scholar]
- 26.Keiser MS, Kordower JH, Gonzalez-Alegre P, Davidson BL. Broad distribution of ataxin 1 silencing in rhesus cerebella for spinocerebellar ataxia type 1 therapy. Brain. 2015;138: 3555–3566. 10.1093/brain/awv292 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The incorporation of exosomes labeled with PKH26 (red arrow) was observed under a confocal microscope (TCS SP8, Leica Microsystems).
(TIFF)
Total RNA from SH-SY5Y cells treated with exosomes derived from non-treated Caco-2 cells or Caco-2 cells treated with 1 mM or 10 mM carnosine were used for DNA microarray analysis with the SurePrint G3 Human Gene Expression 8x60K v3 array. The heat map was shown.
(TIFF)
Representative interaction network of target genes of miRNA with altered expression in Exo-Car and mRNAs with altered expression in Exo-Car-treated SH-SY5Y revealed by in silico analysis.
(TIFF)
The changes in the expression of miR-619-5p in Caco-2 cells treated with carnosine (A). The changes in the expression of miR-619-5p in cells treated with Exo-Car are shown in B. The changes in the expression of miR-619-5p in SH-SY5Y cells treated with Exo-Car are shown in C.
(TIFF)
(PDF)
(PDF)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.