

Abstract
Purpose of review
In humans, only one independent immunologic correlate of reduced risk of HIV infection has been identified: a robust antibody (Ab) response to the V1V2 domain of the gp120 envelope (Env) protein. In recent years, the presence and level of V1V2-specific Abs has also been correlated with protection from SIV and SHIV infections. Here, we review the multitude of studies showing the in-vivo protective effects of V1V2 Abs and review their immunologic characteristics and antiviral functions.
Recent findings
Structural and immunologic studies have defined four epitope families in the V1V2 domain: one epitope family, V2q, which preferentially presents as a quaternary structure of the Env trimer, and another epitope family (V2qt) which requires the quaternary trimeric Env structure; these two epitope types are recognized by two families of monoclonal Abs (mAbs)–V2q-specific and V2qt-specific mAbs–which display broad and potent neutralizing activity. A third epitope family, V2i, is present as a discontinuous conformational structure that overlays the α4β7 integrin binding motif, and a fourth epitope family (V2p) exists on V2 peptides. Antibodies specific for V2i and V2p epitopes display only poor neutralizing activity but effectively mediate other antiviral activities and have been correlated with control of and/or protection from HIV, SIV and SHIV. Notably, V2q and V2qt Abs have not been induced by any vaccines, but V2p and V2i Abs have been readily induced with various vaccines in nonhuman primates and humans.
Summary
The correlation of vaccine-induced V2p and V2i Abs with protection from HIV, SIV and SHIV suggests that these Ab types are extremely important to induce with prophylactic vaccines.
Keywords: antibodies, antiviral functions, epitope, nonhuman primates, V2, vaccines
INTRODUCTION
The key to the rational design of an effective HIV vaccine depends on the identification of immune correlates of protection and immunologic mechanisms that prevent HIV acquisition. The first independent correlate of reduced risk of HIV infection in humans was identified by studies of participants in the RV144 clinical vaccine trial: a robust antibody (Ab) response to the V1V2 region of the virus gp120 envelope (Env) glycoprotein. Similar correlations were subsequently identified in studies of nonhuman primates (NHPs) between protection from SIV and SHIV and V2 Ab levels. Here, we summarize these human and NHP findings and the V2 Abs that are involved in the control of and/or protection from HIV, SIV and SHIV.
Box 1.

no caption available
STRUCTURAL AND IMMUNOLOGIC CHARACTERIZATION OF THE V1V2 DOMAIN
Studies of polyclonal sera from HIV-infected individuals have established that, over time, infection generates different ‘humoral fingerprints’ [1]. This is true for patterns of Ab specificities, subclasses and antiviral activities [2–5,6▪]. Similar findings pertain to Abs induced by vaccines targeting SIV [7▪,8] and SHIV [9–12]. Given the association between V2 Abs and protection in human and animal models, it is critical to understand the complexity of the V1V2 domain of the virus Env and the Ab response to it.
Conformational complexity of V1V2
In HIV, the V1V2 domain, like the rest of gp120, exhibits marked conformational flexibility. The V1V2 domain serves as the ‘trimer association domain’ at the apex of the closed trimeric Env, but the V1V2 domain of each of the three gp120 protomers opens out when gp120 interacts with CD4 [13–15,16▪]. The C-strand of V2, composed of amino acids (AAs) 170–176, one of the five strands composing the V1V2 β-barrel, exists in different conformations, varying between a β-strand and an α-helix [17–19], where the β-strand configuration is preferentially present in the closed, structurally constrained trimeric Env, whereas the α-helical conformation is preferred where there is less structural constraint when the Env is fully open. The preferred configuration is undoubtedly affected by the sequence of V1 and V2, substitutions at key residues, the molecular context in which the V1V2 domain is placed and the intraprotomer and interprotomer interactions of V1V2 within the Env trimer [20].
Alternative V2 conformational epitopes
As a result of this configurational complexity, there are at least four types of epitopes in the V1V2 region as shown in Fig. 1: V2q epitopes which preferentially recognize structures formed by the quaternary interaction of the three gp120 protomers and are glycan-dependent; V2q is recognized by V2q mAbs such as PG9 and PG16 [17,21–24]; V2qt epitopes which recognize quaternary, trimer-dependent V2 epitopes at the apical center of Env are recognized by several V2qt mAbs exemplified by PGT145 [25]; V2i epitopes which overlay the α4β7 integrin binding site in V2 and are recognized by V2i mAbs such as 830A and 2158 [26–28]; and V2p epitopes which are presented by V2 linear and cyclic peptides, and recognized by V2p mAbs such as CH58 and CAP228-16H [19,29▪,30▪]. The V2q, V2qt and V2i mAbs preferentially recognize their various epitopes when the C-strand of V2 is in the β-strand conformation. In contrast, V2p mAbs recognize the C-strand in its α-helical configuration [19]. These four families of mAbs that recognize the various V1V2 epitopes display distinct patterns of reactivity; an example is shown in Fig. 2 in which the patterns of reactivity of V2p and V2i mAbs are shown vs. a panel of eight V2-bearing antigens.
FIGURE 1.
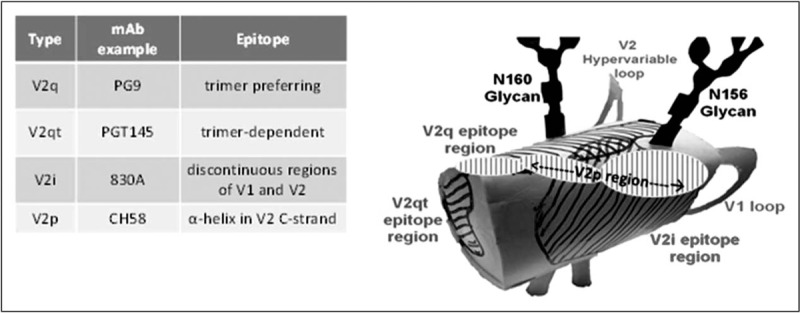
Diagram of the position of the four epitope classes in the V1V2 region (V2p, V2i, V2q and V2qt), and examples of mAbs that target each of these epitopes.
FIGURE 2.
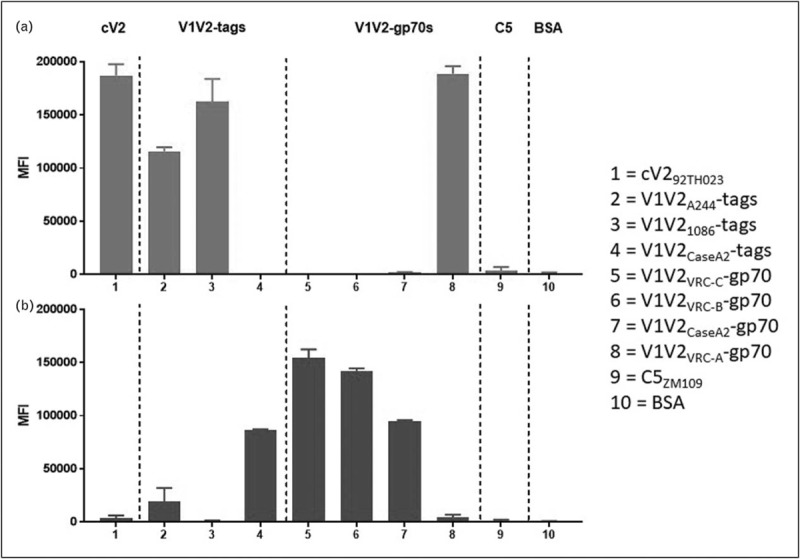
Luminex reactivity of V2p mAb 8490 (a) and V2i mAb 697-30D (b) vs. cyclic V2 peptide (column 1), V1V2-tags (columns 2–4), V1V2-gp70 fusion proteins (columns 5–8), C5 peptide (column 9) and BSA (column 10).
The correlates of reduced risk in RV144
RV144 is the only clinical vaccine trial to date which provided a marginal but statistically significant reduced rate of infection [31] as well as an independent correlate of risk (COR) of HIV infection: an inverse relationship between the incidence of infection and the level of Abs binding to a V1V2CaseA2-gp70 fusion protein [32–34]. The V1V2CaseA2-gp70 fusion protein preferentially reacts with V2i mAbs, and indeed, several V2i mAbs have been selected from the cells of HIV-infected individuals using this reagent, and these V2i mAbs are highly cross-clade reactive [35,36]. In this context, it is noteworthy that while V1V2CaseA2-gp70 carries the V1V2 sequence from a clade B strain [37], infections occurring in the RV144 participants were primarily because of clade AE (CRF01_AE), the predominant circulating strain in Thailand, and this supports the hypothesis that the V2 Abs implicated in the inverse COR were cross-clade reactive.
Notably, in human vaccine studies other than RV144, the induction of highly reactive and functional V2-specific Abs has not been strong, for example in studies such as VAX003, VAX004 and HVTN100 [5,38,39]. It is hypothesized that the gp120 of the clade AE A244 strain used in RV144 is unusual in its ability to efficiently induce V2 Abs. Thus, for example, the immunogens used in HVTN100 [ALVAC-HIV (vCP2438) and bivalent Subtype C gp120 s (1086 and TV1)], which was the precursor of the ongoing phase III HVTN702 study in South Africa, induced a markedly poorer V2 response than that attained in RV144 [38]. These findings are of particular interest in the context of recent studies in which robust V2i and V2p Ab responses have been elicited using V2-targeting vaccine constructs in rabbits [40,41] and NHPs [42]. Notably, in the latter study, the use of a trimeric V1V2A244-scaffold fusion protein as part of an immunogen cocktail appeared to be particularly effective in inducing broadly reactive and functional V2 Abs.
In addition to the inverse COR with V2i Abs, a similar role for V2p Abs has been documented. Thus, studies with plasma from RV144 vaccinees demonstrated an inverse COR in terms of the Ab response to linear V2 peptides tested by microarray [43], and the correlations of Abs cross-reactive with V2 peptides representing different HIV clades were at least as significant as the correlation seen with the primary variable generated using the V1V2CaseA2-gp70 fusion protein. As noted, linear and cyclic V2 (cV2) peptides preferentially assume a structure when complexed with specific V2p mAbs in which the C-strand is in an α-helical configuration [19,29▪,30▪], and two such V2p mAbs were isolated from circulating cells of an individual receiving the RV144 vaccine regimen [19]. Thus, we know from polyclonal and mAb studies emanating from RV144 that V2p Abs recognizing the α-helix in the C-strand of V2 were induced by the RV144 vaccine and that they constitute an inverse COR ([43] and Table 1) [32,43–48,49▪,50,51,52▪]. Additional V2p mAbs have recently been isolated from individuals infected with clade C [29▪,30▪]. All of these V2p mAbs have been crystallized and reveal the targeted epitope in the V2 C-strand as an α-helix or helix-loop, and, like the plasma V2p polyclonal Abs in RV144 vaccinees [33], these mAbs are cross-clade reactive (Fig. 2A) [37].
Table 1. Studies showing statistically significant correlations of V2-specific antibodies that correlate with protection from and/or control of HIV, SIV and SHIV.
| Year | Protection and/or control of | Correlation with Abs specific for | Immunization | Reference |
| 2012 | HIV SIVmac251 | V2i V2p | Active Active | Haynes et al. [32] Barouch et al. [44] |
| 2013 | HIV SIVmac251 | V2p V2p | Active Active | Gottardo et al. [43] Pegu et al. [45] |
| 2014 | SIVmac251 | V2p | Active | Gordon et al. [46] |
| 2015 | SIVE660 | V2p | Active | Roederer et al. [47] |
| 2016 | SIVmac251 | V2p | Active | Vaccari et al. [48] |
| 2018 | SHIVBaL SIVsmE66 SHIVBaL SHIVBaL | V2i V2p V2 V2p | Passive Active Active Active | Hessell et al. [49▪] Singh et al. [50] Hessell et al. [51] Weiss et al. [52▪] |
THE ROLE OF V2 ABS IN THE CONTROL OF AND PROTECTION FROM SIV AND SHIV INFECTIONS IN NONHUMAN PRIMATES
The original observation of an inverse COR in RV144 has been supported by many subsidiary studies of the RV144 data [3,4,43,53,54]. Nonetheless, there are critics who remain skeptical of the RV144 correlates analyses [55]. This skepticism is now tempered by both active and passive immunization studies from many laboratories showing correlates of protection from SIV and SHIV infection with the presence of V2 Abs (Table 1).
V2p-specific Abs protect against SIV infection
In an NHP vaccine study using vaccine regimens consisting of Ad26 and/or modified vaccinia Ankara vector-based vaccines expressing SIVSME543 gag, pol and Env antigens with subsequent intrarectal (i.r.) challenges with SIVmac251, there was at least 80% reduction in per-exposure probability of infection [44]. The strength of Ab binding to a biotinylated cyclic V2 peptide from SIVSME543 correlated positively with the number of challenges required to establish infection (P < 0.0001).
Another NHP study utilized priming with gp160 DNA from SIVmac239 and boosting with recombinant Ad5SIVmac239 with subsequent multiple mucosal challenges with SIVE660[47]. Among the significant humoral response correlates identified was the strength of binding of plasma IgG Abs to a V1V2SIVmac239 linear peptide with time to infection (P = 0.009). In this study, a sieving effect was also noted at a glycosylation site in V1V2 that conferred neutralization resistance. This is similar to the sieving signature identified in the RV144 human trial [53].
Additional studies in NHPs also support the role of V2-specific Abs in protection from or control of SIV. With sera from animals immunized with a regimen similar to that used in RV144, Ab assays revealed that the reduced risk of SIVmac251 acquisition correlated with the presence of mucosal IgG to cyclic V2 (P = 0.0018) [48,45]. In yet another study, where animals were immunized with gp120 protein and human papilloma pseudoviruses expressing SIVmac251 genes +/– ALVAC-SIVmac251, a significant correlation was found between the number of challenges to achieve persistent infection with SIVmac251 and the avidity index for V1V2 Abs in blood (P = 0.014) [46]. And most recently, in NHPs immunized with SIVmac251-derived env plasmids and monomeric M766 gp120 protein followed by challenge with SIVE660, inverse correlations were identified for plasma and mucosal V1V2 responses with peak viral load (P = 0.05 and P = 0.01, respectively), responses to cyclic V2 peptides with postpeak and chronic viremia (P = 0.01 for each) and V2-specific responses with delayed virus acquisition and postinfection control [50]. Each of these experiments suggests that Abs that hampered SIV infection were of the V2p type which recognize the α-helical V2 C-strand configuration.
V2p Abs protect against SHIV infection
Most recently, an RV144-like vaccine regimen was tested in NHPs that were challenged with SHIVBaL[51]. All three unimmunized animals were infected after two i.r. challenges, but in five of the nine immunized macaques, tight control of viremia was noted as reflected by only transient and low plasma viral load (PVL) measurements, with no measurable virus in tissues at necropsy 13 weeks after challenge. Luminex studies of the plasma from these animals showed a correlate of protection from SHIVBaL with Abs of the V2p type that were reactive with V1V21086-tags [19], a reagent in which the V2 C-strand preferentially adopts an α-helical conformation as shown by circular dichroism [52▪].
THE ROLE OF V2I ABS IN SHIV VIRAL CONTROL
To date, only indirect, correlative data have linked V2 Abs with protection in human and NHP experiments. The first direct in-vivo test of the hypothesis emanating from RV144 — that V2 Abs could reduce the risk of infection — was reported recently in experiments investigating the role of a passively administered V2i mAb in protection from, and/or control of infection in NHPs challenged i.r. with SHIVBaL[49▪]. NHPs received 3 weekly doses at 10 mg/kg of the IgG1 isoform of the V2i mAb 830A (n = 12), whereas control animals (n = 12) received a dengue-specific mAb (DEN3) at the same times and doses. Animals were challenged with SHIVBaL twice during each week that they received the passively transferred mAb. Blood specimens were drawn at regular intervals at the time of and after challenge, and animals were sacrificed and necropsied 6 weeks after infection. On the basis of SIVgag RNA copies/ml in plasma, 11/12 control animals were infected by the sixth and final SHIVBaL challenge dose, and while the 12th animal had no detectable PVL, SHIV was detected in its liver at necropsy. Of the 12 animals receiving V2i mAb 830A, three had no detectable PVL at necropsy and no SIVgag DNA was detected in their tissues. Of these latter three macaques, one was plasma aviremic after all challenge doses and two had only low and transient positive PVLs at a single time point. Compared in a grouped analysis, the PVL in the 830A recipients was significantly lower than that in the DEN3 controls (P = 0.031). The cell-associated viral load (CAVL) DNA in peripheral blood mononuclear cells was assessed as virus copies collected during the course of the experiment, and a statistical analysis revealed a significant difference in CAVLs between controls and the IgG1 830A group (P = 0.038). In addition, copy numbers of viral DNA in 13 different tissues were measured at the time of necropsy and revealed significant differences in the viral DNA loads between viremic 830A-treated and control macaques (P = 0.015). Copy numbers of SIVgag DNA associated with each tissue sampled from the 830A-treated macaques were compared individually to the corresponding tissue from animals in the control group and again significant differences in copy numbers were found in iliosacral, axillary and inguinal lymph nodes and from mixed tissues from the reproductive tract. Thus, while too few animals remained uninfected in the treated group to achieve statistical significance in terms of the risk of infection, the data demonstrate that the presence of the passively administered V2i 830A mAb had significant effects against SHIV challenge in macaques by reducing the viral infectious titer so that animals were either not infected or experienced lower level virus production in blood and tissues, reduced plasma virus load, and decreased viral DNA in lymphoid tissues. This is the first direct demonstration showing the ability of V2i Abs to impede SHIV infection [49▪].
The biologic functions of V2 antibodies
Neutralizing Abs were not an inverse COR in the RV144 vaccine trial, suggesting that non-neutralizing Ab effects were critical. These effects could be mediated by either the Fab portion of the Ab which binds to antigens on the surface of the virus or virus-infected cell and/or by the Fc portion of the Ab which binds to Fc receptors (FcRs) after the Fab fragment binds to its antigen.
Antiviral activities mediated by the Fab fragment of antibodies
Neutralization of virus infectivity is the most frequently measured antiviral activity, resulting as a function of the attachment of Abs to virions. However, several other phenomena belong to this category of antiviral functions and appear to play a critical role in vivo[56]. These include:
-
(1)
Virus aggregation: Abs can bind to virions; when they do, they can aggregate viruses, enhancing their destruction via phagocytosis. It has been shown that there is a clear association between virus aggregation and virus phagocytosis [57], and this may play a role in reducing the virus inoculum upon exposure to the virus, resulting in a reduced risk of infection. Similarly, this mechanism may assist in the clearance of circulating virus, which, perhaps as a result, has a half-life in circulation of only approximately 0.3 days [58]. Monoclonal IgG and dimeric IgA Abs, as well as polyclonal purified serum IgA from RV144 participants, have been shown to aggregate HIV virions, and the specificities of the Abs used to do so include mAbs to various epitopes of both gp120 and gp41 [59].
-
(2)
Virus capture: This in-vitro assay has been used to demonstrate that the RV144 vaccine-elicited gp120-specific Abs of multiple specificities (V3, V2 and C1) can bind to virions [60]. These studies were extended to show that C1-specific and V2-specific mAbs derived from vaccinees acted in synergy to capture virions [54].
-
(3)
Inhibition of virus binding to cell-surface α4β7: It was shown in 2008 that a tripeptide in the V2 loop of gp120 (at AA 179–181) constitutes a motif that mimics structures presented by the natural ligands of the α4β7 integrin, a gut-homing receptor, and that the HIV-1 envelope protein gp120 bound to an activated form of α4β7 [61]. In a recent follow-up to the initial findings, the V2 domain of gp120 was shown to preferentially engage extended forms of α4β7 in a cation-sensitive manner. A 15-mer V2-derived peptide (AA 168–181) can bind to α4β7, and V2p-specific mAbs derived from vaccinated and infected individuals recognize this peptide and block the V2/α4β7 interaction [62▪]. Other laboratories have reproduced and extended these findings [63] and have shown that purified IgG from some uninfected RV144 vaccinees can also inhibit the V2/α4β7 interaction. These data suggest that anti-V2 Abs may play a role in vivo, blocking the gp120/α4β7 interaction and thus preventing HIV acquisition and/or controlling early targeting of the gut lymphoid tissues by the virus.
Fc-dependent effector functions
Many Abs Mediate antiviral effects because of their ability to bind to infected cells and/or virions, leading to conformational changes in the Fc fragment which allow it to bind to FcRs on the surface of various cell types such as T cells, monocyte/macrophages, polymorphonuclear granulocytes, dendritic cells, and so on. These Abs include both bnAbs and Abs that are poor or non-neutralizers, can be specific for various regions of the gp120 and gp41 Env proteins (Fig. 3 and [19,30▪,40,54,64–70]), and can bind to the Env trimer in its different states (closed, partially open or fully open [16▪]). The Fc/FcR interaction initiates antiviral activities that include Ab-dependent cellular phagocytosis (ADCP) and Ab-dependent cellular cytotoxicity [2,4,54,71–73] as well as complement-mediated virolysis [39,74–76]. Many of these mechanisms have been associated with reduced risk of infection. For example, ADCP has been associated with reduced HIV infection risk in humans [39,77] and with SIV infection in NHPs [12], and Ab-dependent complement activation and deposition [78,79▪] as well as Ab-dependent cell-mediated viral inhibition have been shown to contribute to control of SIV and SHIV [80–83].
FIGURE 3.
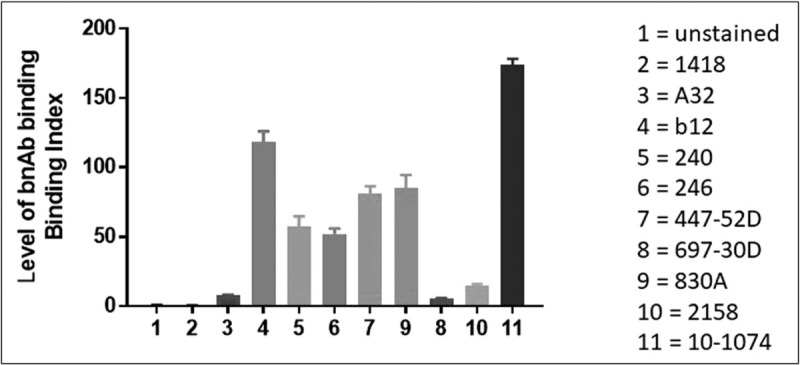
Level of binding of mAbs to native-like Env. Results shown are for binding of mAbs to tetherinhi Jurkat cells nucleofected with an mCherry+ NL4-3 reporter construct as described by Alvarez et al.[70]. Monoclonal Abs used (and the epitope for which they are specific) include: human antiparvovirus mAb (1418), A32 (anti-C1), b12 (CD4bs), 240 and 246 (gp41, cluster I), 447-52D (V3 crown), 697-30D (V2i), 830A (V2i), 2158 (V2i) and 10-1074 (V3-glycan).
CONCLUSION
The first and only independent correlate of reduced risk of HIV infection in humans was identified by studies of participants in the RV144 clinical vaccine trial: a robust Ab response to the V1V2 region of the virus gp120 Env glycoprotein. Subsequent to this observation, several active and passive immunization studies in NHPs identified the presence and level of V2 Abs as correlates of protection from SIV and SHIV infections. Currently, 11 vaccine studies in humans and NHPs (summarized in Table 1) support the role of V1V2-specific Abs in protection. In each case, the Abs involved displayed little or no neutralizing activity but mediated other antiviral activities. Protection was documented against viruses heterologous to the strains used in the vaccines. These studies suggest a new paradigm for vaccine development: protection from and/or control of infection can be achieved with Abs that are induced by existing vaccine constructs, are effective against heterologous viruses, do not display broad and potent neutralizing activity and mediate a variety of non-neutralizing Fab-mediated and Fc-mediated antiviral activities.
Acknowledgements
The authors thank Drs. David Easterhof and Barton Haynes for providing V2p mAb 8490.
Financial support and sponsorship
Funding received for this work came from the U.S. National Institutes of Allergy and Infectious Diseases (NIH grant P01 AI100151) and funds from the Division of Infectious Diseases, Department of Medicine, Icahn School of Medicine at Mount Sinai, New York, NY 10029, USA.
Conflicts of interest
There are no conflicts of interest.
REFERENCES AND RECOMMENDED READING
Papers of particular interest, published within the annual period of review, have been highlighted as:
▪ of special interest
▪▪ of outstanding interest
REFERENCES
- 1.Liu P, Overman RG, Yates NL, et al. Dynamic antibody specificities and virion concentrations in circulating immune complexes in acute to chronic HIV-1 infection. J Virol 2011; 85:11196–11207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chung AW, Kumar MP, Arnold KB, et al. Dissecting polyclonal vaccine-induced humoral immunity against HIV using systems serology. Cell 2015; 163:988–998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yates NL, Liao HX, Fong Y, et al. Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Sci Transl Med 2014; 6:228ra39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chung AW, Ghebremichael M, Robinson H, et al. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci Transl Med 2014; 6:228ra38. [DOI] [PubMed] [Google Scholar]
- 5.Balasubramanian P, Williams C, Shapiro MB, et al. Functional antibody response against V1V2 and V3 of HIV gp120 in the VAX003 and VAX004 vaccine trials. Sci Rep 2018; 8:542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6▪.Yates NL, deCamp AC, Korber BT, et al. HIV-1 envelope glycoproteins from diverse clades differentiate antibody responses and durability among vaccinees. J Virol 2018; 92: e01843-17. [DOI] [PMC free article] [PubMed] [Google Scholar]; Analysis of vaccine-elicited V1V2 binding antibody in longitudinal samples from the RV144 clinical trial revealed a striking heterogeneity among individual vaccinees in maintaining durable responses. These data support the idea that a major goal for vaccine development is to improve antibody levels, breadth and durability at the population level.
- 7▪.Ackerman ME, Das J, Pittala S, et al. Route of immunization defines multiple mechanisms of vaccine-mediated protection against SIV. Nat Med 2018; 24:1590–1598. [DOI] [PMC free article] [PubMed] [Google Scholar]; The route of immunization plays a critical role for phagocytic Fc-effector Ab activity in protection from SIV, driving Fc activity via distinct innate effector cells and antibody isotypes. The same correlates predict protection from SHIV infection. Thus, functional humoral mechanisms initiated by distinct vaccination routes and immunization strategies are pivotal for inducing multiple, potentially complementary correlates of immunity.
- 8.Miller-Novak LK, Das J, Musich TA, et al. Analysis of complement-mediated lysis of simian immunodeficiency virus (SIV) and SIV-infected cells reveals sex differences in vaccine-induced immune responses in rhesus macaques. J Virol 2018; 92: [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bradley T, Pollara J, Santra S, et al. Pentavalent HIV-1 vaccine protects against simian-human immunodeficiency virus challenge. Nat Commun 2017; 8:15711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Srivastava IK, Stamatatos L, Kan E, et al. Purification, characterization, and immunogenicity of a soluble trimeric envelope protein containing a partial deletion of the V2 loop derived from SF162, an R5-tropic human immunodeficiency virus type 1 isolate. J Virol 2003; 77:11244–11259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barnett SW, Srivastava IK, Kan E, et al. Protection of macaques against vaginal SHIV challenge by systemic or mucosal and systemic vaccinations with HIV-envelope. AIDS 2008; 22:339–348. [DOI] [PubMed] [Google Scholar]
- 12.Barouch DH, Stephenson KE, Borducchi EN, et al. Protective efficacy of a global HIV-1 mosaic vaccine against heterologous SHIV challenges in rhesus monkeys. Cell 2013; 155:531–539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kwon YD, Finzi A, Wu X, et al. Unliganded HIV-1 gp120 core structures assume the CD4-bound conformation with regulation by quaternary interactions and variable loops. Proc Natl Acad Sci U S A 2012; 109:5663–5668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mao Y, Wang L, Gu C, et al. Subunit organization of the membrane-bound HIV-1 envelope glycoprotein trimer. Nat Struct Mol Biol 2012; 19:893–899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Munro JB, Gorman J, Ma X, et al. Conformational dynamics of single HIV-1 envelope trimers on the surface of native virions. Science 2014; 346:759–763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16▪.Wang H, Barnes CO, Yang Z, et al. Partially open HIV-1 envelope structures exhibit conformational changes relevant for coreceptor binding and fusion. Cell Host Microbe 2018; 24:579–592.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]; Cryo-electron micrographic structures of SOSIP with various Abs and/or CD4 reveal the structure of the displaced V1V2 and reveal its structures.
- 17.McLellan JS, Pancera M, Carrico C, et al. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 2011; 480:336–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pan R, Gorny MK, Zolla-Pazner S, Kong XP. The V1V2 region of HIV-1 gp120 forms a five-stranded beta barrel. J Virol 2015; 89:8003–8010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liao H-X, Bonsignori M, Alam SM, et al. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 2013; 38:1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Powell RLR, Totrov M, Itri V, et al. Plasticity and epitope exposure of the HIV-1 envelope trimer. J Virol 2017; 91: e00410-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gorny MK, Stamatatos L, Volsky B, et al. Identification of a new quaternary neutralizing epitope on human immunodeficiency virus type 1 virus particles. J Virol 2005; 79:5232–5237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kimura T, Wang XH, Williams C, et al. Human monoclonal antibody 2909 binds to pseudovirions expressing trimers but not monomeric HIV-1 envelope proteins. Hum Antibodies 2009; 18 (1–2):35–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pancera M, Zhou T, Druz A, et al. Structure and immune recognition of trimeric prefusion HIV-1 Env. Nature 2014; 514:455–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Walker LM, Phogat SK, Chan-Hui PY, et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009; 326:285–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lee JH, Andrabi R, Su CY, et al. A broadly neutralizing antibody targets the dynamic HIV envelope trimer apex via a long, rigidified, and anionic beta-hairpin structure. Immunity 2017; 46:690–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mayr LM, Cohen S, Spurrier B, et al. Epitope mapping of conformational V2-specific anti-HIV human monoclonal antibodies reveals an immunodominant site in V2. PLoS One 2013; 8:e70859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gorny MK, Moore JP, Conley AJ, et al. Human anti-V2 monoclonal antibody that neutralizes primary but not laboratory isolates of HIV-1. J Virol 1994; 68:8312–8320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nyambi PN, Mbah HA, Burda S, et al. Conserved and exposed epitopes on intact, native, primary human immunodeficiency virus type 1 virions of group M. J Virol 2000; 74:7096–7107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29▪.Wibmer CK, Richardson SI, Yolitz J, et al. Common helical V1V2 conformations of HIV-1 Envelope expose the alpha4beta7 binding site on intact virions. Nat Commun 2018; 9:4489. [DOI] [PMC free article] [PubMed] [Google Scholar]; Structural studies of the epitope recognized by new V2p mAbs derived from patients infected with HIV.
- 30▪.van Eeden C, Wibmer CK, Scheepers C, et al. V2-directed vaccine-like antibodies from HIV-1 infection identify an additional K169-binding light chain motif with broad ADCC activity. Cell Rep 2018; 25:3123–3135.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]; Functional studies of new V2p mAbs derived from HIV-infected individuals.
- 31.Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med 2009; 361:2209–2220. [DOI] [PubMed] [Google Scholar]
- 32.Haynes BF, Gilbert PB, McElrath MJ, et al. Immune correlates analysis of the ALVAC-AIDSVAX HIV-1 vaccine efficacy trial. N Engl J Med 2012; 366:1275–1286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zolla-Pazner S, deCamp AC, Cardozo T, et al. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PLos One 2013; 8:e53629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zolla-Pazner S, DeCamp AC, Gilbert PB, et al. Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLos One 2014; 9:e87572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gorny MK, Pan R, Williams C, et al. Functional and immunochemical cross-reactivity of V2-specific monoclonal antibodies from human immunodeficiency virus type 1-infected individuals. Virology 2012; 427:198–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pinter A, Honnen WJ, He Y, et al. The V1/V2 domain of gp120 is a global regulator of sensitivity of primary human immunodeficiency virus type 1 isolates to neutralization by antibodies commonly induced upon infection. J Virol 2004; 78:5205–5215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pinter A, Honnen WJ, Kayman SC, et al. Potent neutralization of primary HIV-1 isolates by antibodies directed against epitopes present in the V1/V2 domain of HIV-1 gp120. Vaccine 1998; 16:1803–1811. [DOI] [PubMed] [Google Scholar]
- 38.Shen X, Moodie Z, McMillan A, et al. V1V2 IgG and Antibody Fc Effector Functions in a Subtype C ALVAC-HIV and Bivalent Subtype C gp120/MF59 HIV-1 Vaccine Trial in South Africa. HIV Research for Prevention. Madrid. 2018. Abstract OA02.03. [Google Scholar]
- 39.Perez LG, Martinez DR, deCamp AC, et al. V1V2-specific complement activating serum IgG as a correlate of reduced HIV-1 infection risk in RV144. PLoS One 2017; 12:e0180720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zolla-Pazner S, Powell R, Yahyaei S, et al. Rationally-designed vaccines targeting the V2 region of HIV-1 gp120 induce a focused, cross clade-reactive biologically functional antibody response. J Virol 2016; 90:10993–11006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jiang X, Totrov M, Li W, et al. Rationally designed immunogens targeting HIV-1 gp120 V1V2 induce distinct conformation-specific antibody responses in rabbits. J Virol 2016; 90:11007–11019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hessell AJ, Powell R, Jaing X, et al. Multimeric epitope-scafold HIV vaccines target V1V2 and differentially tune polyfunctional antibody responses. Cell Rep 2019; in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gottardo R, Bailer RT, Korber BT, et al. Plasma IgG to linear epitopes in the V2 and V3 regions of HIV-1 gp120 as correlates of infection risk in the RV144 vaccine efficacy trial. PLoS One 2013; 8:e75665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Barouch DH, Liu J, Li H, et al. Vaccine protection against acquisition of neutralization-resistant SIV challenges in rhesus monkeys. Nature 2012; 482:89–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pegu P, Vaccari M, Gordon S, et al. Antibodies with high avidity to the gp120 envelope protein in protection from simian immunodeficiency virus SIV(mac251) acquisition in an immunization regimen that mimics the RV-144 Thai trial. J Virol 2013; 87:1708–1719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gordon SN, Doster MN, Kines RC, et al. Antibody to the gp120 V1/V2 loops and CD4+ and CD8+ T cell responses in protection from SIVmac251 vaginal acquisition and persistent viremia. J Immunol 2014; 193:6172–6183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Roederer M, Keele BF, Schmidt SD, et al. Immunological and virological mechanisms of vaccine-mediated protection against SIV and HIV. Nature 2014; 505:502–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Vaccari M, Gordon SN, Fourati S, et al. Adjuvant-dependent innate and adaptive immune signatures of risk of SIV acquisition. Nat Med 2016; 22:762–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49▪.Hessell AJ, Shapiro MB, Powell R, et al. Reduced cell-associated DNA and improved viral control in macaques following passive transfer of a single anti-V2 monoclonal antibody and repeated SHIV challenges. J Virol 2018; 92: e02198-17. [DOI] [PMC free article] [PubMed] [Google Scholar]; Efficacy of V2i mAbs passively administered to NHPs and challenged with SHIVBaL.
- 50.Singh S, Ramirez-Salazar EG, Doueiri R, et al. Control of heterologous simian immunodeficiency virus SIVsmE660 infection by DNA and protein coimmunization regimens combined with different toll-like-receptor-4-based adjuvants in macaques. J Virol 2018; 92: e00281-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hessell AJ DM, Philip B, Shilpi P, et al. Tight Control of SHIV BaL.P4 Challenge in Rhesus Macaques Co-immunized With DNA and Protein HIV Vaccine Regimen. HIV Research for Prevention; Madrid. 2018. Abstract OA09.4. [Google Scholar]
- 52▪.Weiss S, Vincenza I, Ruimin P, et al. Tight Control of SHIVBaL in Rhesus Macaques Immunized With gp160 DNA + gp120 Proteins (Clades E and B) Correlates With V2p Antibodies. HIV Research for Prevention; Madrid. 2018. Abstract OA14.06LB. [Google Scholar]; First description of a correlate of protection from SHIV infection with the presence of V2p Abs.
- 53.Rolland M, Edlefsen PT, Larsen BB, et al. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature 2012; 490:417–420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pollara J, Bonsignori M, Moody MA, et al. HIV-1 vaccine-induced C1 and V2 Env-specific antibodies synergize for increased antiviral activities. J Virol 2014; 88:7715–7726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Desrosiers RC. Protection against HIV acquisition in the RV144 trial. J Virol 2017; 91: e00905-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mayr LM, Su B, Moog C. Non-neutralizing antibodies directed against HIV and their functions. Front Immunol 2017; 8:1590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gach JS, Bouzin M, Wong MP, et al. Human immunodeficiency virus type-1 (HIV-1) evades antibody-dependent phagocytosis. PLoS Pathog 2017; 13:e1006793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Perelson AS, Neumann AU, Markowitz M, et al. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 1996; 271:1582–1586. [DOI] [PubMed] [Google Scholar]
- 59.Alexander MR, Sanders RW, Moore JP, Klasse PJ. Short Communication: virion aggregation by neutralizing and nonneutralizing antibodies to the HIV-1 envelope glycoprotein. AIDS Res Hum Retroviruses 2015; 31:1160–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Liu P, Yates NL, Shen X, et al. Infectious virion capture by HIV-1 gp120-specific IgG from RV144 vaccinees. J Virol 2013; 87:7828–7836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Arthos J, Cicala C, Martinelli E, et al. HIV-1 envelope protein binds to and signals through integrin alpha4beta7, the gut mucosal homing receptor for peripheral T cells. Nat Immunol 2008; 9:301–309. [DOI] [PubMed] [Google Scholar]
- 62▪.Lertjuthaporn S, Cicala C, Van Ryk D, et al. Select gp120 V2 domain specific antibodies derived from HIV and SIV infection and vaccination inhibit gp120 binding to alpha4beta7. PLoS Pathog 2018; 14:e1007278. [DOI] [PMC free article] [PubMed] [Google Scholar]; HIV and SIV V2 mAbs elicited by both vaccination and infection block the interaction of V2 and α4β7.
- 63.Peachman Kristina K, Karasavvas N, Chenine A-L, et al. Identification of new regions in HIV-1 gp120 variable 2 and 3 loops that bind to α4β7 integrin receptor. PLoS One 2015; 10:e0143895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Alvarez RA, Hamlin RE, Monroe A, et al. HIV-1 Vpu antagonism of tetherin inhibits antibody-dependent cellular cytotoxic responses by natural killer cells. J Virol 2014; 88:6031–6046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Holl V, Peressin M, Decoville T, et al. Nonneutralizing antibodies are able to inhibit human immunodeficiency virus type 1 replication in macrophages and immature dendritic cells. J Virol 2006; 80:6177–6181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Musich T, Li L, Liu L, et al. Monoclonal antibodies specific for the V2, V3, CD4-binding site, and gp41 of HIV-1 mediate phagocytosis in a dose-dependent manner. J Virol 2017; 91: e02325-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mayr LM, Decoville T, Schmidt S, et al. Nonneutralizing antibodies targeting the V1V2 domain of HIV exhibit strong antibody-dependent cell-mediated cytotoxic activity. Sci Rep 2017; 7:12655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Chung AW, Crispin M, Pritchard L, et al. Identification of antibody glycosylation structures that predict monoclonal antibody Fc-effector function. AIDS 2014; 28:2523–2530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Burton DR, Hessell AJ, Keele BF, et al. Limited or no protection by weakly or nonneutralizing antibodies against vaginal SHIV challenge of macaques compared with a strongly neutralizing antibody. Proc Natl Acad Sci U S A 2011; 108:11181–11186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Alvarez RA, Maestre AM, Law K, et al. Enhanced FCGR2A and FCGR3A signaling by HIV viremic controller IgG. JCI Insight 2017; 2:e88226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bonsignori M, Pollara J, Moody MA, et al. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J Virol 2012; 86:11521–11532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Gomez-Roman VR, Patterson LJ, Venzon D, et al. Vaccine-elicited antibodies mediate antibody-dependent cellular cytotoxicity correlated with significantly reduced acute viremia in rhesus macaques challenged with SIVmac251. J Immunol 2005; 174:2185–2189. [DOI] [PubMed] [Google Scholar]
- 73.Excler JL, Ake J, Robb ML, et al. Nonneutralizing functional antibodies: a new ‘old’ paradigm for HIV vaccines. Clin Vaccine Immunol 2014; 21:1023–1036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Aasa-Chapman MM, Holuigue S, Aubin K, et al. Detection of antibody-dependent complement-mediated inactivation of both autologous and heterologous virus in primary human immunodeficiency virus type 1 infection. J Virol 2005; 79:2823–2830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Spear GT, Landay AL, Sullivan BL, et al. Activation of complement on the surface of cells infected by human immunodeficiency virus. J Immunol 1990; 144:1490–1496. [PubMed] [Google Scholar]
- 76.Spear GT, Sullivan BL, Landay AL, Lint TF. Neutralization of HIV-1 by complement occurs by viral lysis. J Virol 1990; 64:5869–5873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Sips M, Krykbaeva M, Diefenbach TJ, et al. Fc receptor-mediated phagocytosis in tissues as a potent mechanism for preventive and therapeutic HIV vaccine strategies. Mucosal Immunol 2016; 9:1584–1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ackerman ME, Mikhailova A, Brown EP, et al. Polyfunctional HIV-specific antibody responses are associated with spontaneous HIV control. PLoS Pathog 2016; 12:e1005315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79▪.Alter G, Dowell KG, Brown EP, et al. High-resolution definition of humoral immune response correlates of effective immunity against HIV. Mol Syst Biol 2018; 14:e7881. [DOI] [PMC free article] [PubMed] [Google Scholar]; Systemic serological analysis of a cohort of HIV-infected individuals with varying degrees of viral control revealed multifaceted and coordinated contributions of polyclonal antibodies to diverse antiviral responses and suggests key biophysical features predictive of viral control.
- 80.Forthal DN, Gilbert PB, Landucci G, Phan T. Recombinant gp120 vaccine-induced antibodies inhibit clinical strains of HIV-1 in the presence of fc receptor-bearing effector cells and correlate inversely with HIV infection rate. J Immunol 2007; 178:6596–6603. [DOI] [PubMed] [Google Scholar]
- 81.Hidajat R, Xiao P, Zhou Q, et al. Correlation of vaccine-elicited systemic and mucosal nonneutralizing antibody activities with reduced acute viremia following intrarectal simian immunodeficiency virus SIVmac251 challenge of rhesus macaques. J Virol 2009; 83:791–801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Florese RH, Demberg T, Xiao P, et al. Contribution of nonneutralizing vaccine-elicited antibody activities to improved protective efficacy in rhesus macaques immunized with Tat/Env compared with multigenic vaccines. J Immunol 2009; 182:3718–3727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Xiao P, Zhao J, Patterson LJ, et al. Multiple vaccine-elicited nonneutralizing antienvelope antibody activities contribute to protective efficacy by reducing both acute and chronic viremia following simian/human immunodeficiency virus SHIV89.6P challenge in rhesus macaques. J Virol 2010; 84:7161–7173. [DOI] [PMC free article] [PubMed] [Google Scholar]