

Abstract
There is an increasing amount of clinical evidence that hypomagnesemia (serum Mg2+ levels < 0.7 mmol/l) contributes to type 2 diabetes mellitus pathogenesis. Amongst other hypotheses, it has been suggested that Mg2+ deficiency affects insulin secretion. The aim of this study was, therefore, to investigate the acute effects of extracellular Mg2+ on glucose-stimulated insulin secretion in primary mouse islets of Langerhans and the rat insulinoma INS-1 cell line. Here we show that acute lowering of extracellular Mg2+ concentrations from 1.0 mM to 0.5 mM did not affect glucose-stimulated insulin secretion in islets or in insulin-secreting INS-1 cells. The expression of key genes in the insulin secretory pathway (e.g. Gck, Abcc8) was also unchanged in both experimental models. Knockdown of the most abundant Mg2+ channel Trpm7 by siRNAs in INS-1 cells resulted in a 3-fold increase in insulin secretion at stimulatory glucose conditions compared to mock-transfected cells. Our data suggest that insulin secretion is not affected by acute lowering of extracellular Mg2+ concentrations.
Introduction
Globally, the number of people that suffer from type-2 diabetes mellitus (T2DM) is steadily increasing, and the prevalence is predicted to pass the threshold of 500 million people by 2030 [1]. T2DM is characterized by impaired insulin secretion (i.e. insulin deficiency) and insulin sensitivity (i.e. insulin resistance), explaining the underlying pathophysiological mechanism for hyperglycemia (fasting serum blood glucose > 7 mmol/L) [2].
There is increasing evidence that hypomagnesemia (blood magnesium (Mg2+) < 0.7 mmol/L) is associated with T2DM pathogenesis and related complications [3]. Several epidemiological studies have shown that hypomagnesemia is higher in T2DM patients (14% - 48%) than in controls (2.5% - 15%) [3]. Indeed, a patient cohort of almost 400 T2DM patients showed that 30.6% suffered from hypomagnesemia with plasma Mg2+ levels below 0.7 mmol/L [4]. Furthermore, Mg2+ supplementation in T2DM patients improved insulin sensitivity and glucose metabolism [5–7].
Small scale clinical studies have demonstrated that insulin secretion may be lower in T2DM patients with hypomagnesemia [8, 9]. Supplementation of individuals without diabetes with MgCl2 significantly increased beta-cell function in a small randomized clinical trial [10]. However, experimental data concerning the role of Mg2+ in insulin secretion are conflicting, as both inhibitory and stimulatory effects have been reported in rodent models of hypomagnesemia. Isolated islets of rats on a Mg2+-deficient diet for 11 weeks showed a higher basal and glucose-stimulated insulin response [11]. In contrast, Legrand et al observed a reduced insulin response to glucose in rats on a Mg2+-deficient diet for 6 weeks compared to controls [12]. Interpretation of these contradictory results are further complicated since both short-term and long-term effects of Mg2+ may play a role.
In this study, we aimed to delineate the acute effects of extracellular Mg2+ concentrations on glucose-stimulated insulin secretion in isolated primary mouse islets of Langerhans and INS-1 cells under physiological and hyperglycemic conditions.
Materials and methods
Animal care
All procedures were conducted in compliance with the UK Animals Scientific Procedures Act (1986) and approved by the University of Oxford, Department of Physiology, Anatomy and Genetics Ethics Committee. Mice were housed in same-sex littermate groups of 2–8 animals, in a temperature and humidity-controlled room on a 12 hr light-dark cycle (lights on 7 am). Water and food (Special Diet Services, RM3) were available ad libitum. We used 10- to 16-week-old mice with a mixed (C3H, C57BL/6N, 129/sv) genetic background, as previously described [13]. Littermates consisting of wild-type mice, mice carrying Cre recombinase under the control of the rat insulin promoter (RIPII-Cre-ER mice), and mice carrying a transgene consisting of a floxed STOP codon upstream of a Kir6.2-V59M gene that was inserted into the ROSA locus (ROSA mice) (14), were used for insulin secretion experiments. The floxed Kir6.2 gene is not expressed due to the presence of the STOP codon. Likewise Cre-recombinase is not expressed as the gene is only activated when mice are injected with tamoxifen. We did not observe any differences in insulin secretion between these mice [14], or between these mice and wild-type mice on a pure C57bl/6N background.
Pancreatic islet isolation
Mice were sacrificed by cervical dislocation, and pancreatic islets were isolated by collagenase digestion. In brief, the distal end of the common bile duct was tied near the anatomical transition to the duodenum, and the pancreas was distended by injecting HANKS (in mM: 137 NaCl, 2.5 CaCl2, 1.0 NaH2PO4, 5.6 KCl, 4.2 NaHCO3, 2.5 MgSO4 and 10 HEPES (pH 7.4 with NaOH)) containing Liberase (Roche, Mannheim, Germany) (0.33 mg/ml) into the common bile duct. After digestion at 37°C for 16 min, islets were washed in the same buffer, supplemented with 0.2% Bovine Serum Albumin (BSA) (Sigma, St. Louis, USA) and 0.05% D-glucose (Fisher Scientific, Leicestershire, UK). Islets were handpicked using a dissecting microscope into RPMI 1640 medium (Gibco, Darmstadt, Germany) containing 10% FBS (Gibco, Paisley, Scotland), 1% Penicillin-Streptomycin (Gibco, Paisley, Scotland) and 11 mM D-Glucose. They were either left for 24 hrs in this medium at 37°C in a humidified atmosphere with 5% CO2, or (as indicated) cultured for 24 hrs in home-made RPMI (S1 Table) with either normal (1.0 mM) or low (0.1 mM) extracellular magnesium. Islets from different mice were pooled in all experiments.
Islet static insulin secretion
Groups of size-matched islets were handpicked and washed in Krebs-Ringer-bicarbonate (KRB) buffer: (in mM: 118.5 NaCl, 2.5 CaCl2, 1.2 KH2PO4, 4.7 KCl, 25 NaHCO3,1.2 MgCl2 and 10 HEPES (pH 7.4 with NaOH)), followed by a 0.5 hr pre-incubation in KRB without glucose (non-stimulant condition) at 37°C, 5% CO2 infusion. Static insulin secretion was assessed by challenging the islets with final concentrations of glucose and MgCl2 as indicated for 1 hr at 37°C, 5% CO2 infusion. The supernatant was collected to measure insulin secretion using a mouse insulin ELISA (Mercodia, Uppsala, Sweden). Islets were lysed in RIPA buffer (65 mM Tris-HCL, 150 mM NaCl, 5 mM EDTA, 1% NP-40, 0.5% Na-deoxycholate, 0.1% SDS, and 10% glycerol (pH 7.4 with HCl)) for 15 min on ice to extract total insulin content.
Quantitative real time PCR
Islets were lysed directly after isolation or after culture for 24 hrs in home-made RPMI (S1 Table) with glucose and MgCl2, as indicated. Islets were lysed by Qiazol (Qiagen, Hilden, Germany) using the TissueLyser II, and chloroform (Sigma, St. Louis, USA) phase separation was used to extract total RNA. Purification was performed according to the manufacturer’s protocol, including an on-column DNase (Qiagen, Hilden, Germany) digestion step to eliminate genomic DNA. RNA was eluted in two steps using pre-heated RNA-free water and the concentration was checked using a Nano-drop ND-1000 spectrophotometer (Thermo Scientific, Wilmington, USA). Equal amounts of total RNA (0.1–1.5 μg) were reversed transcribed using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems, Lithuania) according to the manufacturer’s protocol. The cDNA was subsequently used to determine the mRNA levels of Trpm6, Trpm7, Slc41a1, Slc41a2, Slc41a3, Cnnm1, Cnnm2, Cnnm3, Cnnm4, Gck and Abcc8 in islets and Trpm7, Cacna1c, Cacna1d, Ins1, Kcnj11, Abcc8, and Gck in INS-1 cells using commercially available Taqman probes by the StepOnePlus Real-Time PCR System (Applied Biosystems) and normalized to Actb expression (Table 1). Data were analyzed using the Livak (2-ΔΔCT) method.
Table 1. Taqman gene expression assays used for RT-PCR analyses.
| Target Gene | Taqman Gene Expression Assays |
|---|---|
| Primary mouse islets of Langerhans | |
| Actb | Mm00607939_s1 |
| Trpm6 | Mm00463112_m1 |
| Trpm7 | Mm00457998_m1 |
| Slc41a1 | Mm00715604_m1 |
| Slc41a2 | Mm01250930_m1 |
| Slc41a3 | Mm01182529_m1 |
| Cnnm1 | Mm00518996_m1 |
| Cnnm2 | Mm01205090_m1 |
| Cnnm3 | Mm01227346_m1 |
| Cnnm4 | Mm01227316_m1 |
| Gck | Rn00561265_m1 |
| Abcc8 | Rn01476317_m1 |
| INS-1 cells | |
| Trpm7 | Rn01328216_m1 |
| Kcnj11 | Rn01764077_s1 |
| Abcc8 | Rn01476317_m1 |
| Cacna1c | Rn00709287_m1 |
| Cacna1d | Rn01453395_m1 |
| Ins1 | Rn02121433_g1 |
| Gck | Rn00561265_m1 |
All Taqman Assays used for experiments in primary mouse islets of Langerhans (top) and INS1 cells (bottom) were commercially available (Applied Biosystems). Actb was used as control in all RT-PCRs.
INS-1 cells
Culture
INS-1 832–13 cells were cultured in RPMI 1640 GlutaMaxTM (Gibco, Darmstadt, Germany) supplemented with 10% FBS, 50 μM β-mercaptoethanol (Sigma, St. Louis, USA), 1 mM sodium pyruvate (Sigma, St. Louis, USA), 1% HEPES (Gibco, Paisley, Scotland), and 1% pen/strep in a humidified atmosphere (37°C, 5% CO2). For mRNA expression analysis, INS-1 cells were seeded in 12-well plates (3 x 105 cells per well) to reach confluence in 72 hrs. For the last 48 hrs, INS-1 cells were transfected with siTrpm7 (siGENOME SMARTpool siRNA, Dharmacon, Lafayette, USA) (final concentration 20 nM) and Lipofectamine® RNAiMAX reagent (Invitrogen, Carlsbad, USA) (1 ng/ml) in Opti-MEM (Gibco, Paisley, Scotland).
INS-1 static insulin secretion
INS-1 cells were cultured in 24-well plates (3 x 105 cells per well) prior to the static secretion assay. INS-1 cells were washed in HBSS buffer (in mM: 114 NaCl, 4.7 KCl, 25.5 NaHCO3, 1.2 KH2PO4, 1.16 MgCl2, 20 HEPES, 2.5 CaCl2 (pH 7.4 with NaOH)), supplemented with 0.07% BSA. After pre-incubation for 2 hrs in HBSS without D-glucose (non-stimulant condition), INS-1 cells were challenged for 0.5 hr with final concentrations of D-glucose and MgCl2, as indicated. At the end of the incubation period, both the supernatant and the total insulin content (obtained by lysing cells in RIPA buffer) were collected, centrifuged for 5 min, at 14,000 rpm, 4°C, and directly stored at -20°C. Insulin secretion and total insulin content were determined using a mouse insulin ELISA kit.
Statistics
In all experiments, all data are expressed as the mean ± SEM. All statistical analyses were performed using GraphPad (Prism 7) software. Statistical comparisons were analyzed by an unpaired Student’s t-test or by a two-way ANOVA with a Tukey’s multiple comparison test. p < 0.05 was regarded as statistically significant.
Results
Acute lowering of Mg2+ does not affect insulin secretion in pancreatic β cells under normal- and hyperglycemic conditions
To investigate whether Mg2+ fulfils a functional role in pancreatic β cells in a normal and hyperglycemic environment, acute effects of extracellular Mg2+ on insulin secretion were examined in primary mouse islets of Langerhans (Fig 1). Insulin secretion at normal (1.0 mM) extracellular Mg2+ for 1hr increased 6-fold in response to elevation of glucose from 2 to 20 mM (Fig 1A) and increased 3-fold when expressed as a percentage of insulin content (Fig 1B). Reduction of extracellular Mg2+ from 1.0 to 0.5 mM for 1 hr had no effect on either basal insulin secretion or that stimulated by 20 mM glucose (Fig 1A and 1B). We next assessed the effects of low Mg2+ in combination with hyperglycemia. Culture at 25 mM glucose (1.2 mM Mg2+) for 24 hrs enhanced basal insulin secretion but was without effect on GSIS. Subsequent exposure to 0.5 or 1.0 mM Mg2+ for 1 hr did not alter basal insulin secretion or GSIS (Fig 1C and 1D). Likewise, increasing the time of culture to 48 hrs at normal (11 mM) or high (25 mM) glucose before acutely lowering extracellular Mg2+ did not alter the results (S1 Fig). Thus, acute exposure to low extracellular Mg2+ does not affect insulin secretion.
Fig 1. Acute lowering of Mg2+ does not affect insulin secretion in pancreatic β cells under normal- and hyperglycemic conditions.

(A-B) Insulin secretion from isolated islets (n = 10 replicates, islets from 16 mice, 8 islets per replicate) challenged by 2 mM and 20 mM glucose at 0.5 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr, after 24 hrs of culture at 11 mM glucose (1.2 mM Mg2+). (C-D) Insulin secretion from mouse pancreatic islets (n = 3, islets from 3 mice, 8 islets per replicate) stimulated by 2 mM and 20 mM glucose with 0.5 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr after 24 hrs of culture at 25 mM glucose (1.2 mM Mg2+). Insulin secretion is presented as ng/islet/hr (A, C) and as normalized to total insulin content (B, D). Statistical significance was determined by two-way ANOVA. *, p < 0.05, 20 mM glucose vs. 2 mM glucose.
Effects of prolonged exposure to low extracellular Mg2+ on insulin secretion
To determine whether prolonged exposure to low extracellular Mg2+ affects GSIS, primary mouse islets of Langerhans were cultured for 24 hrs at either low (0.1 mM) or physiological (1.0 mM) extracellular Mg2+ and under both normal (11 mM) or hyperglycemic (25 mM) glucose concentrations. Insulin secretion was then measured in response to 2 and 20 mM glucose (Fig 2A–2D). In islets cultured at 11 mM glucose, elevation of glucose from 2 to 20 mM increased insulin secretion significantly at both low and normal Mg2+ (16-fold and 30-fold, respectively; Fig 2A and 2B). In islets cultured at 25 mM glucose, there was no difference in basal insulin secretion (2mM glucose) at low and normal Mg2+ (Fig 2C and 2D). However, insulin secretion at 20 mM glucose was significantly higher in islets exposed to 0.1 mM Mg2+ compared to 1.0 mM Mg2+ (Fig 2C and 2D). Thus, in islets cultured under hyperglycemic, low Mg2+ conditions, insulin secretion in response to 20 mM glucose is potentiated.
Fig 2. Effects of prolonged exposure to low extracellular Mg2+ on insulin secretion and gene expression.

(A-B) Insulin secretion from isolated islets (n = 5 replicates, islets from 4 mice, 8 islets per replicate) challenged by 2 mM and 20 mM glucose at 0.1 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr, after 24 hrs of culture at 11 mM glucose and either 0.1 mM or physiological (1.0 mM) extracellular Mg2+. (C-D) Insulin secretion from mouse pancreatic islets (n = 5 replicates, islets from 4 mice, 8 islets per replicate) stimulated by 2 mM and 20 mM glucose at 0.1 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr after 24 hrs of culture at 25 mM glucose, and either 0.1 mM or 1.0 mM extracellular Mg2+. Insulin secretion is presented as ng/islet/hr (A, C) and as normalized to total insulin content (B, D). *, p < 0.05 (2 mM vs. 20 mM glucose); #, p < 0.05 (0.1 mM Mg2+, 20 mM glucose vs. 1.0 mM Mg2+, 20 mM glucose). Two-way ANOVA with a Tukey’s multiple comparison test. (E-L) The mRNA transcript levels of Gck (E), Abcc8 (F), Trpm6 (G), Trpm7 (H), Slc41a1 (I), Slc41a3 (J), Cnnm2 (K), Cnnm4 (L) were measured in islets (n = 3 replicates, islets from 15 mice, 8 islets per replicate) after 24 hrs of culture at 11 mM glucose (solid bar), or 25 mM glucose (open bar) and either 0.1 mM Mg2+ or 1.0 mM Mg2+. mRNA expression levels were determined by quantitative RT-qPCR and normalized to Actb expression. Data are expressed relative to 1.0 mM Mg2+, 11 mM glucose.
Extracellular Mg2+ does not regulate mRNA levels of selected genes
The mRNA expression levels of selected genes were examined by RT-qPCR in islets cultured at 11 mM or 25 mM glucose and either 0.1 mM or 1.0 mM Mg2+. There were no significant effects on mRNA transcript levels of key Mg2+ channels and exchangers; Trpm6, Trpm7, Slc41a1, Slc41a3, Cnnm2, and Cnnm4 (Fig 2G–2L). There were also no significant differences in mRNA expression levels of Gck and Abcc8 between low Mg2+ and physiological Mg2+ concentrations and between normal and hyperglycemic glucose conditions (Fig 2E and 2F).
Extracellular Mg2+ does not affect GSIS in INS-1 cells
To verify our results from the experiments on primary mouse islets, we assessed the effects of low Mg2+ on GSIS in the rat insulinoma INS-1 cell line (Fig 3). Acute reduction of extracellular Mg2+ from 1.0 to 0.5 mM for 0.5 hr had no effect on either basal insulin secretion or that stimulated by 20 mM glucose (Fig 3A and 3B). Additionally, prolonged culture of 48 hrs under low Mg2+ conditions did not affect the transcript levels of Gck, Kcnj11, Abcc8, Cacna1c, and Cacna1d nor Trpm7 and Slc41a1 (Fig 3C–3I).
Fig 3. Extracellular Mg2+ does also not affect GSIS in INS-1 cells.

(A-B) INS1 cells were cultured for 48 hrs at 11 mM glucose (1.2 mM Mg2+) followed by 24 hrs at 5 mM glucose (1.2 mM Mg2+) prior to the experiment. Insulin secretion from INS-1 cells stimulated by 2 mM and 20 mM glucose with 0.5 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 0.5 hr (representative experiment from three independent experiments). Insulin secretion is presented as ng/3*105 cells per hr (A) and normalized to total cell content (B). *, p < 0.05 (2mM vs. 20mM glucose) using two-way ANOVA. (C-I) The mRNA transcript levels of Gck (C), Kcnj11 (D), Abcc8 (E), Cacna1c (F), Cacna1d (G), Trpm7 (H), and Slc41a1 (I) were measured from INS-1 cells after 72 hrs of culture at standard glucose (11 mM) of which the last 48 hrs in low Mg2+ (0.1 mM) (solid bar) or physiological Mg2+ (1.0 mM) (open bar) culture conditions. mRNA expression levels were determined by quantitative RT-qPCR and normalized for Actb expression. Data are expressed relative to 1.0 mM Mg2+.
Trpm7 is the main Mg2+ channel in primary mouse islets
To understand which Mg2+ channels and transporters/exchangers play a pivotal role in pancreatic β cells, their mRNA expression levels were analyzed in primary mouse islets of Langerhans using RT-qPCR (Fig 4). RT-qPCR analysis demonstrated that the Mg2+ channel Trpm7 and the Mg2+ exchanger Cnnm4 were the highest expressed among those tested in primary mouse islets. Notably, Trpm7 expression was 8-fold higher than its close homologue Trpm6, both of which are crucial for epithelial Mg2+ transport (Fig 4A).
Fig 4. Trpm7 is the main Mg2+ channel in primary mouse islets.
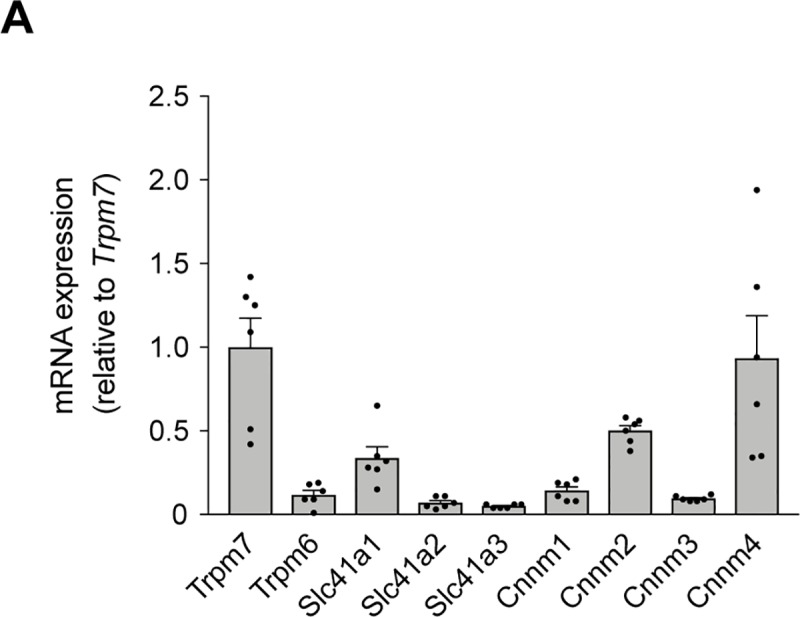
(A) The mRNA transcript levels of Trpm7, Trpm6, Slc41a1, Slc41a2, Slc41a3, Cnnm1, Cnnm2, Cnnm3, and Cnnm4 in islets (n = 6 replicates, islets from 6 mice). mRNA expression levels were determined by quantitative RT-qPCR and normalized to Actb expression. Data are expressed relative to Trpm7 expression.
Knockdown of Trpm7 results in increased insulin secretion in INS-1 cells
Given the high expression levels of the Mg2+ channel Trpm7, we hypothesized that knockdown of Trpm7 might reduce intracellular Mg2+ levels and thereby influence GSIS. Trpm7 was knocked down using siRNA in INS-1 cells (Fig 5). Trpm7 expression was 60% reduced by siRNA transfection (S2 Fig). Basal insulin secretion in Trpm7 knockdown cells was similar to that in control cells. Secretion was significantly increased (by 16-fold) in response to 20 mM glucose in Trpm7 knockdown cells (p = 0.006). At 20 mM glucose, there was significantly more secretion in Trpm7 knockdown cells than control cells (Fig 5A and 5B). Trpm7 knockdown did not affect the mRNA expression levels of key players in GSIS including Gck, Ins1, Kcnj11, Abcc8, Cacna1c, and Cacna1d (Fig 5C–5H).
Fig 5. Knockdown of Trpm7 results in increased insulin secretion in INS-1 cells.

(A-B) Insulin secretion from INS-1 cells (n = 3 experiments, 3 replicates each) stimulated by 2 mM and 20 mM glucose (1.0 mM Mg2+). Cells were cultured for 24 hrs at 11 mM glucose (1.2 mM Mg2+) followed by a further 48 hrs culture at 11 mM glucose (1.2 mM Mg2+) after transfection with siNON-targeting (siNT) (solid bar) or siTrpm7 (open bar). Insulin secretion is presented as ng/3*105 cells per hour (A) and normalized to total cell content (B). *, p < 0.05 (2 mM vs. 20 mM glucose); #, p < 0.05 (siTrpm7, 20 mM glucose vs. siNT, 20 mM glucose). Two-way ANOVA. (C-H) The mRNA transcript levels of Gck (C), Ins1 (p = 0.069) (D), Kcnj11 (p = 0.069) (E), Abcc8 (p = 0.058) (F), Cacna1c (G), Cacna1d (H) measured in INS-1 cells after 24 hrs of culture at 11 mM glucose (1.2 mM Mg2+) followed by a further 48 hrs culture at 11 mM glucose (1.2 mM Mg2+) after transfection with siNON-targeting (siNT) (solid bar) or siTrpm7 (open bar) (n = 3 experiments, 3 replicates each). mRNA expression levels were determined by quantitative RT-qPCR and normalized to Actb expression. Data are expressed relative to siNT levels.
Discussion
This study demonstrates that reduced extracellular Mg2+ does not impair insulin secretion from pancreatic β-cells. This conclusion is based on the following results: i) Acute exposure to low extracellular Mg2+ has no effect on insulin secretion from either isolated mouse islets of Langerhans or INS-1 cells; ii) In islets cultured at 25 mM glucose, to mimic the hyperglycemic state found in T2DM, insulin secretion was actually enhanced by reducing extracellular Mg2+; iii) The mRNA expression levels of Mg2+ channels/transporters and key players in GSIS, in both islets and INS1 cells, were unaffected by extracellular Mg2+.
Previous experimental approaches have focused on the long-term effects of Mg2+ on insulin secretion in mouse and rat models. The results are conflicting. Reis et al showed a higher insulin response to basal and 8.3 mmol/L glucose concentrations in isolated islets of rats fed a Mg2+-deficient diet for 11 weeks [11]. In contrast, Legrand et al. observed a reduced insulin release [12]. The difficulty with long term Mg2+ deprivation is that both the extracellular and intracellular Mg2+ concentration may be reduced. In our approach, we aimed to separate the effects of extracellular Mg2+ and the effects of intracellular Mg2+. Interestingly, an acute reduction of extracellular Mg2+ did not affect insulin secretion in either primary islets or INS-1 cells. Similar findings were reported by an early study in perfused rat pancreas [15]. In contrast, knockdown of Trpm7, which is permeable to Mg2+, significantly increased glucose-stimulated insulin release. This leads us speculate that changes in intracellular Mg2+ might modulate insulin release. Indeed, data from RIN m5F cells showed a stimulatory effect on insulin secretion when intracellular Mg2+ levels were reduced [16, 17].
Our results indicate that transient hypomagnesemia does not affect insulin secretion. However, patients with hypomagnesemia have chronic hypomagnesemia. Long term hypomagnesemia might reduce intracellular Mg2+ levels and thereby modify GSIS. Indeed, our results show that culturing islets for 24 hrs in a hyperglycemic environment under low Mg2+ conditions stimulated GSIS. This is in line with the negative correlation between magnesium levels and HOMA-B in a Canadian T2DM cohort [9].
Interestingly, our data show the importance of Trpm7 in insulin secretion from pancreatic islets. Trpm7 is the highest expressed Mg2+ channel. Although a major limitation of our study is the lack of measurement of intracellular Mg2+ levels, others have extensively reported that targeting Trpm7 severely reduces intracellular Mg2+ levels [18–20]. Additionally, as TRPM7 can also function as a Ca2+ channel, we cannot exclude that TRPM7-mediated Ca2+ uptake explains the increased insulin secretion [21].
In recent years, several studies have shown that hypomagnesemia is a risk factor for progression of T2DM [3, 22]. Several hypotheses that could explain the mechanisms of this association have been proposed [23, 24]. Given the conflicting literature and our data showing that acute changes in extracellular Mg2+ do not inhibit insulin secretion in both pancreatic islets and INS-1 cells, we postulate that hypomagnesemia does not exert its effect primarily on insulin secretion. Based on recent large studies in animals and patients, other factors such as lipid metabolism may be more important in determining the interaction between hypomagnesemia and progression of type 2 diabetes [4, 25, 26].
In conclusion, we showed in different models that changes in extracellular Mg2+ do not impair glucose-stimulated insulin secretion. However, further research is needed to elucidate whether regulation of the intracellular Mg2+ concentration by Trpm7 may influence insulin release.
Supporting information
(DOCX)
(A-B) Insulin secretion from isolated islets (n = 3 replicates, 9 mice, 8 islets per replicate) challenged with 2 mM and 20 mM glucose and 0.5 mM Mg2+ (solid bar) or 1.0 mM Mg2+ (open bar) for 1 hr after 48 hrs of culture at 11 mM glucose (1.2 mM Mg2+). (C-D) Insulin secretion from mouse pancreatic islets (n = 3 replicates, 9 mice, 8 islets per replicate) stimulated by 2 mM and 20 mM glucose with 0.5 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr after 48 hrs of culture at 25 mM glucose (1.2 mM Mg2+). Insulin secretion is presented as ng/islet/hr (A, C) and normalized to total insulin content (B, D). *, p < 0.05 (2 mM vs. 20 mM glucose); Two-way ANOVA.
(DOCX)
(A) RT-qPCR of Trpm7 mRNA in INS-1 cells (n = 3 experiments, 3 replicates each) following transfection with siNON-targeting (siNT) (solid bar) or siTrpm7 (open bar). mRNA expression levels were determined by quantitative RT-qPCR and normalized to Actb expression. Data are expressed relative to siNT. *, p < 0.05, Student’s t-test (two-tailed).
(DOCX)
Acknowledgments
The authors thank Raúl Terron Exposito, Maria Rohm and Melissa Brereton for their technical support.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by grants from the Netherlands Organization for Scientific Research (NWO Rubicon 825.14.021, NWO VENI 016.186.012) and the Wellcome Trust (884655, 089795). Jeroen de Baaij is supported by the Dutch Diabetes Research Foundation (2017.81.014). Frances Ashcroft holds a European Research Council Advanced Investigatorship and a Royal Society Research Wolfson Merit Award. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Olokoba AB, Obateru OA, Olokoba LB. Type 2 diabetes mellitus: a review of current trends. Oman Med J. 2012;27(4):269–73. 10.5001/omj.2012.68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cantley J, Ashcroft FM. Q&A: insulin secretion and type 2 diabetes: why do beta-cells fail? BMC Biol. 2015;13:33 Epub 2015/05/20. 10.1186/s12915-015-0140-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pham PC, Pham PM, Pham SV, Miller JM, Pham PT. Hypomagnesemia in patients with type 2 diabetes. Clinical journal of the American Society of Nephrology: CJASN. 2007;2(2):366–73. Epub 2007/08/21. 10.2215/CJN.02960906 . [DOI] [PubMed] [Google Scholar]
- 4.Kurstjens S, de Baaij JH, Bouras H, Bindels RJ, Tack CJ, Hoenderop JG. Determinants of hypomagnesemia in patients with type 2 diabetes mellitus. European journal of endocrinology. 2017;176(1):11–9. Epub 2016/10/22. 10.1530/EJE-16-0517 . [DOI] [PubMed] [Google Scholar]
- 5.Guerrero-Romero F, Rodriguez-Moran M. [Oral magnesium supplementation: an adjuvant alternative to facing the worldwide challenge of type 2 diabetes?]. Cir Cir. 2014;82(3):282–9. Epub 2014/09/23. . [PubMed] [Google Scholar]
- 6.Paolisso G, Passariello N, Pizza G, Marrazzo G, Giunta R, Sgambato S, et al. Dietary magnesium supplements improve B-cell response to glucose and arginine in elderly non-insulin dependent diabetic subjects. Acta endocrinologica. 1989;121(1):16–20. Epub 1989/07/01. . [DOI] [PubMed] [Google Scholar]
- 7.Paolisso G, Sgambato S, Pizza G, Passariello N, Varricchio M, D'Onofrio F. Improved insulin response and action by chronic magnesium administration in aged NIDDM subjects. Diabetes care. 1989;12(4):265–9. Epub 1989/04/01. 10.2337/diacare.12.4.265 . [DOI] [PubMed] [Google Scholar]
- 8.Rodriguez-Moran M, Guerrero-Romero F. Insulin secretion is decreased in non-diabetic individuals with hypomagnesaemia. Diabetes Metab Res Rev. 2011;27(6):590–6. Epub 2011/04/14. 10.1002/dmrr.1206 . [DOI] [PubMed] [Google Scholar]
- 9.Randell EW, Mathews M, Gadag V, Zhang H, Sun G. Relationship between serum magnesium values, lipids and anthropometric risk factors. Atherosclerosis. 2008;196(1):413–9. Epub 2006/12/13. 10.1016/j.atherosclerosis.2006.11.024 . [DOI] [PubMed] [Google Scholar]
- 10.Guerrero-Romero F, Rodriguez-Moran M. Magnesium improves the beta-cell function to compensate variation of insulin sensitivity: double-blind, randomized clinical trial. Eur J Clin Invest. 2011;41(4):405–10. Epub 2011/01/19. 10.1111/j.1365-2362.2010.02422.x . [DOI] [PubMed] [Google Scholar]
- 11.Reis MA, Latorraca MQ, Carneiro EM, Boschero AC, Saad MJ, Velloso LA, et al. Magnesium deficiency improves glucose homeostasis in the rat: studies in vivo and in isolated islets in vitro. The British journal of nutrition. 2001;85(5):549–52. Epub 2001/05/12. . [DOI] [PubMed] [Google Scholar]
- 12.Legrand C, Okitolonda W, Pottier AM, Lederer J, Henquin JC. Glucose homeostasis in magnesium-deficient rats. Metabolism: clinical and experimental. 1987;36(2):160–4. Epub 1987/02/01. . [DOI] [PubMed] [Google Scholar]
- 13.Brereton MF, Iberl M, Shimomura K, Zhang Q, Adriaenssens AE, Proks P, et al. Reversible changes in pancreatic islet structure and function produced by elevated blood glucose. Nat Commun. 2014;5:4639 Epub 2014/08/26. 10.1038/ncomms5639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Girard CA, Wunderlich FT, Shimomura K, Collins S, Kaizik S, Proks P, et al. Expression of an activating mutation in the gene encoding the KATP channel subunit Kir6.2 in mouse pancreatic beta cells recapitulates neonatal diabetes. The Journal of clinical investigation. 2009;119(1):80–90. Epub 2008/12/10. 10.1172/JCI35772 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Atwater I, Frankel BJ, Rojas E, Grodsky GM. Beta cell membrane potential and insulin release; role of calcium and calcium:magnesium ratio. Q J Exp Physiol. 1983;68(2):233–45. Epub 1983/04/01. . [DOI] [PubMed] [Google Scholar]
- 16.Ishizuka J, Bold RJ, Townsend CM Jr., Thompson JC. In vitro relationship between magnesium and insulin secretion. Magnes Res. 1994;7(1):17–22. Epub 1994/03/01. . [PubMed] [Google Scholar]
- 17.Murakami M, Ishizuka J, Sumi S, Nickols GA, Cooper CW, Townsend CM Jr., et al. Role of extracellular magnesium in insulin secretion from rat insulinoma cells. Proc Soc Exp Biol Med. 1992;200(4):490–4. Epub 1992/09/11. . [DOI] [PubMed] [Google Scholar]
- 18.Ryazanova LV, Rondon LJ, Zierler S, Hu Z, Galli J, Yamaguchi TP, et al. TRPM7 is essential for Mg(2+) homeostasis in mammals. Nat Commun. 2010;1:109 Epub 2010/11/04. 10.1038/ncomms1108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schmitz C, Perraud AL, Johnson CO, Inabe K, Smith MK, Penner R, et al. Regulation of vertebrate cellular Mg2+ homeostasis by TRPM7. Cell. 2003;114(2):191–200. Epub 2003/07/31. . [DOI] [PubMed] [Google Scholar]
- 20.Mittermeier L, Demirkhanyan L, Stadlbauer B, Breit A, Recordati C, Hilgendorff A, et al. TRPM7 is the central gatekeeper of intestinal mineral absorption essential for postnatal survival. Proceedings of the National Academy of Sciences of the United States of America. 2019. Epub 2019/02/17. 10.1073/pnas.1810633116 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Monteilh-Zoller MK, Hermosura MC, Nadler MJ, Scharenberg AM, Penner R, Fleig A. TRPM7 provides an ion channel mechanism for cellular entry of trace metal ions. The Journal of general physiology. 2003;121(1):49–60. Epub 2003/01/01. 10.1085/jgp.20028740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kieboom BCT, Ligthart S, Dehghan A, Kurstjens S, de Baaij JHF, Franco OH, et al. Serum magnesium and the risk of prediabetes: a population-based cohort study. Diabetologia. 2017;60(5):843–53. Epub 2017/02/23. 10.1007/s00125-017-4224-4 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gommers LM, Hoenderop JG, Bindels RJ, de Baaij JH. Hypomagnesemia in Type 2 Diabetes: A Vicious Circle? Diabetes. 2016;65(1):3–13. Epub 2015/12/24. 10.2337/db15-1028 . [DOI] [PubMed] [Google Scholar]
- 24.Gunther T. The biochemical function of Mg(2)+ in insulin secretion, insulin signal transduction and insulin resistance. Magnesium research. 2010;23(1):5–18. Epub 2010/03/17. 10.1684/mrh.2009.0195 . [DOI] [PubMed] [Google Scholar]
- 25.Kurstjens S, de Baaij JHF, Overmars-Bos C, van den Munckhof ICL, Garzero V, de Vries MA, et al. Increased NEFA levels reduce blood Mg(2+) in hypertriacylglycerolaemic states via direct binding of NEFA to Mg(2). Diabetologia. 2018. Epub 2018/11/15. 10.1007/s00125-018-4771-3 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kurstjens S, van Diepen JA, Overmars-Bos C, Alkema W, Bindels RJM, Ashcroft FM, et al. Magnesium deficiency prevents high-fat-diet-induced obesity in mice. Diabetologia. 2018. Epub 2018/07/11. 10.1007/s00125-018-4680-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(A-B) Insulin secretion from isolated islets (n = 3 replicates, 9 mice, 8 islets per replicate) challenged with 2 mM and 20 mM glucose and 0.5 mM Mg2+ (solid bar) or 1.0 mM Mg2+ (open bar) for 1 hr after 48 hrs of culture at 11 mM glucose (1.2 mM Mg2+). (C-D) Insulin secretion from mouse pancreatic islets (n = 3 replicates, 9 mice, 8 islets per replicate) stimulated by 2 mM and 20 mM glucose with 0.5 mM Mg2+ (solid bar) and 1.0 mM Mg2+ (open bar) for 1 hr after 48 hrs of culture at 25 mM glucose (1.2 mM Mg2+). Insulin secretion is presented as ng/islet/hr (A, C) and normalized to total insulin content (B, D). *, p < 0.05 (2 mM vs. 20 mM glucose); Two-way ANOVA.
(DOCX)
(A) RT-qPCR of Trpm7 mRNA in INS-1 cells (n = 3 experiments, 3 replicates each) following transfection with siNON-targeting (siNT) (solid bar) or siTrpm7 (open bar). mRNA expression levels were determined by quantitative RT-qPCR and normalized to Actb expression. Data are expressed relative to siNT. *, p < 0.05, Student’s t-test (two-tailed).
(DOCX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.