

Keywords: Alzheimer's disease, neurodegenerative disease, cognitive impairment, memory, Alzheimer's disease mouse model, mesenchymal stem cell, exosomes, neurogenesis, cognitive improvement, cell-free therapy, neural regeneration
Abstract
Studies have shown that mesenchymal stem cell-derived exosomes can enhance neural plasticity and improve cognitive impairment. The purpose of this study was to investigate the effects of mesenchymal stem cell-derived exosomes on neurogenesis and cognitive capacity in a mouse model of Alzheimer’s disease. Alzheimer’s disease mouse models were established by injection of beta amyloid 1−42 aggregates into dentate gyrus bilaterally. Morris water maze and novel object recognition tests were performed to evaluate mouse cognitive deficits at 14 and 28 days after administration. Afterwards, neurogenesis in the subventricular zone was determined by immunofluorescence using doublecortin and PSA-NCAM antibodies. Results showed that mesenchymal stem cells-derived exosomes stimulated neurogenesis in the subventricular zone and alleviated beta amyloid 1−42-induced cognitive impairment, and these effects are similar to those shown in the mesenchymal stem cells. These findings provide evidence to validate the possibility of developing cell-free therapeutic strategies for Alzheimer’s disease. All procedures and experiments were approved by Institutional Animal Care and Use Committee (CICUAL) (approval No. CICUAL 2016-011) on April 25, 2016.
Chinese Library Classification No. R456; R361; R741
Introduction
Alzheimer’s disease (AD) is the most common form of dementia, affecting brain regions that exhibit high synaptic activity and are implicated in brain functions such as memory and learning (Liang et al., 2008). As AD progresses, there is a substantial loss of neurons mainly precipitated by the extracellular deposition of beta amyloid peptides (Aβ) and the intracellular accumulation of hyperphosphorylated tau neurofibrillary tangles (Reiss et al., 2018). AD etiology is complex, and several mechanisms have been fully studied. However, evidence from many laboratories supports an Aβ accumulation as the fundamental initiator of the disease (Selkoe, 2000), but there is controversy regarding whether Aβ accumulation alone indicates inevitable progression to AD. Some investigators suggest that some other factors, such as synaptic, mitochondrial, metabolic, inflammatory, neuronal, and other age-related alterations as well as tau pathology facilitate the downstream of amyloid (Gomez-Isla et al., 1997; De Strooper and Karran 2016; Segerstrom, 2018). However, the link between these factors remains unclear.
Due to the complex pathophysiological process of this disease, there is currently no effective treatment for AD (Frozza et al., 2018). There are only five Food and Drug Administration approved drugs used for AD treatments, including tacrine, donepezil, rivastigmine, galantamine (all of them are acetylcholinesterase inhibitors), and memantine (an antagonist of NMDA receptor). Unfortunately, these drugs only have a symptomatologic effect (Santos et al., 2016). AD is a multi-factorial disease. Therefore, the treatment must be a multi-target therapy that uses molecules designed to affect different disease processes like Aβ aggregation, reduce chronic inflammation, and increase antioxidant activity (Piemontese, 2017).
Recent studies have shown that neurogenesis can take place in the adult brain, and several therapeutic strategies, mostly using therapeutic mesenchymal stem cells (MSCs), have been developed to replace the cells lost during the progression of AD (Ferreira et al., 2018). Furthermore, administration of MSCs has been shown to stimulate neurogenesis in the adult rat brain, and consequently, use of MSCs has been proposed in the treatment of neurodegenerative diseases (Yan et al., 2014; Segal-Gavish et al., 2016; Gobshtis et al., 2017; Kho et al., 2018).
MSCs are non-hematopoietic multi-potent cells that exhibit great auto-renovation and differentiation capacities. Additionally, they are capable of interacting with resident cells in the brain parenchyma orchestrating a dynamic homeostatic response that promotes tissue preservation and functional recovery (Harting et al., 2018). The interaction between MSCs and resident brain cells stimulates the production of neurotrophins such as brain-derived neurotrophic factor, vascular endothelial growth factor and nerve growth factor as well as the anti-inflammatory cytokines that can potentialize neuritic growth, neuroregeneration, and neurologic recovery (Qu and Zhang, 2017; Harris et al., 2018; Donders et al., 2018). There is evidence that administration of appropriate conditioned medium has a similar effect to administration of MSCs (Timmers et al., 2007; Mitsialis and Kourembanas, 2016). MSCs mediate their activity not through direct cell replacement, but via the paracrine activity of secretome (Nakano et al., 2016; Yang et al., 2017a, b). Further studies have revealed that one of the critical components regulating the paracrine activity of MSCs is exosomes (Drommelschmidt et al., 2017; Phinney and Pittenger, 2017).
Exosomes are 30–150 nm-sized vesicles of endocytic origin that are released into the blood stream when multivesicular bodies and cell plasma membrane fuse (Colombo et al., 2014). These nanovesicles have the ability to transfer active biomolecules such as protein, lipid, and nucleic acid between cells in response to relevant physiologic stimuli received by such cells, and accordingly exosomes will display the distinctive characteristics of their cell of origin (de Jong et al., 2012; Harting et al., 2018). Several studies have described that administration of MSCs-derived exosomes can promote neurogenesis in the subventricular zone (SVZ) and dentate gyrus (DG) of the hippocampus and reduce the cognitive impairments associated with Parkinson’s disease, stroke, and traumatic brain injury (Zhang et al., 2016; Xiong et al., 2017; Yang et al., 2017a). Furthermore, in a recent study, it was demonstrated that MSCs and MSC-derived extracellular vesicles have neuroprotective capacity against oxidative stress and synapse damage induced by Aβ toxicity. MSCs and MSC-derived extracellular vesicles have been reported to internalize and degrade Aβ oligomers, secrete antioxidant enzymes, anti-inflammatory cytokines, and neurotrophic factors to the medium (de Godoy et al., 2018). Taking all of these into consideration, exosomes can be part of a therapeutic strategy to treat AD by reducing inflammatory activity, promoting neurogenesis and improving cognitive deficits associated with the disease. The present study aims to evaluate the overall effects of MSC-derived exosomes on cognitive deficits and neurogenesis in a mouse model of AD.
Materials and Methods
Animals
Forty-eight 7–8 week-old C57BL/6 mice were purchased from Envigo RMS, Mexico. These mice were allocated in translucent polycarbonate boxes in a temperature controlled (25 ± 2°C) laboratory animal facility with light-dark cycles of 12/12 hours and ad libitum access to water and food. Animals were then randomly assigned into two groups: in the first (Aβ) group (n = 36), Aβ aggregates (Sigma-Aldrich, St. Louis, MO, USA) were administered in the dentate gyrus bilaterally, and in the normal control group (n = 8), mice were not treated. Mice in the first group were randomly assigned into three groups (n = 12). The first group was administered exosomes (exosome group), the second group was administered MSCs (MSCs group), and the third group was administered PBS solution (0.1 M, pH 7.4, AD group). All procedures and experiments were done according to national (NOM-062-ZOO-1999) and international Guide for Care and Use of Laboratory Animals with the institutional approval by the ethics committee (approval No. CICUAL 2016-011) on April 25, 2016.
Cell culture and exosome collection
To collect exosomes, MSCs (ATCC, Manassas, VA, USA) were cultivated in exosome production medium centrifuged at 110,000 × g (Sorvall WX90, Thermo Scientific, Langenselbold, Germany) during 12–16 hours at 4°C. The supernatant was subsequently filtered with a 0.22 μm filter. MSCs were cultivated at a cellular density of 5 × 106 cells at 37°C in a 5% CO2 atmosphere. Medium was replaced every third day, and the previous conditioned medium was collected. Exosomes were collected by ultracentrifugation (Sorvall centrifuge rotor AH-627, Thermo Scientific) as previously reported (Thery et al., 2006) with some modifications: conditioned medium was centrifuged at 2000 × g to eliminate cell debris, and aftewards centrifugated at 25,000 × g to eliminate large vesicles, filtered through a 0.22 μm pore. The filtered medium was then spun by the ultracentrifuge at 110,000 × g for 90 minutes, and all supernatant was removed. After addition of PBS solution, the mixture was spun again at 110,000 × g for an additional 90 minutes. After removal of supernatant, exosomes were re-suspended in 50 µL of PBS. Centrifugation and ultracentrifugation were always performed at 4°C. Exosomes were quantified by the DC protein assay kit (BioRad, Hercules, CA, USA) according to manufacturer’s instructions and then stored at –80°C until further use.
Western blot assay
Western blot assay was performed to confirm the presence of exosomes in the pellets obtained through ultracentrifugation as previously reported (Thery et al., 2006). Exosomes were separated through 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) in non-reductive conditions with total protein obtained from MSCs as reference. Proteins were then transferred into a polyvinylidene fluoride (PVDF) membrane (Millipore, Darmstadt, Germany) and incubated overnight at 4°C with the exosomal marker CD81 (0.5 µg/mL, Abcam, Cambridge, MA, USA) and beta actin (Santa Cruz Biotechnology, Dallas, TX, USA). After three TBS-T washes, membranes were incubated with horseradish peroxidase (HRP)-conjugated anti-mouse secondary antibody (1:5000; Vector Laboratories, Burlingame, CA, USA) for 2 hours at room temperature. Membrane exposure was performed by chemiluminescence using Luminata Forte (MerckMillipore, Darmstadt, Germany) and the ChemiDoc™ XRS system (BioRad, Hercules, CA, USA) and Image Lab 6.0.1 software (BioRad).
SDS-PAGE with Tris-Tricine (BioRad) was performed to verify the existence of Aβ aggregates. The proteins obtained were transferred to a PVDF membrane; and the membrane was fixed in glutaraldehyde (Sigma-Aldrich) at 0.5% for 10 minutes. After three TBS-T washes, the membranes were incubated overnight at 4°C with rabbit Aβ antibody (D54D2) XP® (0.5 μg/mL, Cell Signaling, Danvers, MA, USA), then incubated with HRP-conjugated anti-rabbit secondary antibody (1:5000, Vector Laboratories, Burlingame, CA, USA) for 2 hours at room temperature as previously described.
Induction of AD and treatments
AD models were established as previously reported by Eslamizade et al. (2015), Aβ1–42 (Sigma-Aldrich) protein was incubated at 37°C 4 days prior to its administration to induce the formation of aggregates. Animals were then subsequently anesthetized with a combination of xylazine (5–16 mg/kg, PiSA Agropecuaria, Hidalgo, Mexico) and ketamine (80–120 mg/kg, PiSA Agropecuaria) administered by intraperitoneal injection. 3 µL of Aβ1–42 (10 ng/µL) aggregates were administered by bilateral stereotaxic (Leica Microsystems, Wetzlar, Germany) surgery at a flow rate of 0.5 µL/min in the DG of the hippocampus at the specified coordinates: anteroposterior –2.8 mm, mediolateral +/– 1.02 mm, and dorsoventral DV –2.03 mm according to the stereotaxic apparatus’s software Angle Two (version 3.0, Leica Microsystems). Within 14 days of administration of Aβ1–42 aggregates, cognitive abilities of mice were evaluated using the Morris Water Maze (MWM) and Novel Object Recognition (NOR) tests.
Once the AD model was established, mice were sedated with xylazine and ketamine as previously described, and different treatments were administered by stereotaxic surgery in the same coordinates used when establishing the model. Mice in the exosome group received 10 µg exosomes in 2 µL of PBS, those in the MSCs group received 1 × 106 cells in 2 µL of PBS, and those in the AD group received 2 µL of PBS. At 14 and 28 days after intervention, the MWM and NOR tests were performed again as previously described to evaluate cognitive ability improvement in mice.
Morris Water Maze test
The MWM test is widely used to assess and validate neurocognitive deficits in rodent models of neurological diseases (Vorhees and Williams 2006). This test was performed in a round pool (150 cm in diameter and 50 cm in depth) with a black bottom and filled with water at 25 ± 2°C as previously reported by Reza-Zaldivar et al. (2017); the maze was virtually divided into four quadrants. A polycarbonate square 10 × 10 cm deck was placed at the center of each quadrant about 1 cm below the surface of the water. Mice received a training period consisting of 4 daily sessions in which they were permitted to swim freely for 60 seconds, entering the water in each of four different quadrants. Animals that were not capable of reaching the platform in this time period were guided and placed on the deck for 20 seconds, a 15-minute time lapse was allowed upon entering each quadrant. All assessments were video recorded for further analyses. Endpoints evaluated were the time to reach the platform and total distance traveled.
Novel Object Recognition test
The NOR test is widely used to assess memory disorders in animal models, and it can be modified to evaluate work memory, attention, anxiety, and preference for novelty in rodents (Bevins and Besheer, 2006). There were three phases in the NOR test: habituation phase: in which the animals were placed in the exploration space without any objects during 5 minutes so they could recognize the environment 24 hours prior to the test; familiarization phase: the animals were placed in the exploration space with 2 identical objects inside (plastic bolts and screws) during 5 minutes after which they were placed for 15 minutes to rest in their respective cages; and finally the test phase at which animals were again placed in the exploration space where one of the objects was replaced by a different one, allowing the mice to interact freely with the objects for 5 minutes. The exploration space was thoroughly cleaned with 70% ethanol between every test to avoid preferences caused by odors to interfere with the test. Discrimination index (DI) between objects was calculated with the following formula: %DI = TN/ (TF + TN) × 100, where TN represents interaction with the novel object, and TF represents interaction with the familiar object.
Immunofluorescence for DCX and PSA-NCAM markers
After 14 and 28 days of behavioral tests, all mice were anesthetized with a lethal dose of sodium pentobarbital (50 mg/kg, PiSA agropecuaria) administered by intraperitoneal injection. A cardiac perfusion with PBS and 4% paraformaldehyde was performed on all mice. Brains were removed, fixed with 4% paraformaldehyde overnight at 4°C, and subsequently immersed in 10%, 20% and 30% sucrose solutions (Hycel, Jal, Mexico) with sodium azide (Sigma-Aldrich) for 24 hours at 4°C. The brains were then embedded in tissue-Tek optimum cutting temperature compound (Sakura Finetek, Torrance, CA, USA) and cut into 10 µm-thick sections using a cryostat (Leica Biosystems, Wetzlar, Germany).
To evaluate neurogenesis, doublecortin (DCX) and PSA-NCAM are widely used markers to identify immature neurons and neuroblasts respectively. For the immunofluorescence, six coronal sections obtained from the control, exosome, MSC and AD groups (n = 4 mice/group) were blocked in a mixture of 5% horse serum and 0.1% Triton-PBS solution for 2 hours. Subsequently, the coronal sections were incubated with rabbit anti-DCX (Abcam, Cambridge, MA, USA) and mouse anti-PSA-NCAM (1:250; Millipore Darmstadt, Germany) overnight at 4°C to evaluate neurogenesis and later on incubated for 2 hours at room temperature with secondary antibodies Alexa 488 anti-mouse (1:500; Vector Laboratories, Burlingame, CA, USA) and Alexa 594 anti-rabbit (1:500; Vector Laboratories) protected from light at all times. Vectashield with DAPI was used to stain the nucleus (0.5 μg/mL; Vector Laboratories). Finally, coronal sections were observed in a Leica DM4 DF7000T fluorescence microscope (Leica Biosystems, Wetzlar, Germany). Leica Application Suite (LAS) X version 3.1.1.15751 (Leica Biosystems) software and Leica DFC7000T camera (Leica Biosystems) were used for image acquisition with a 40× objective for all the antibodies that were used and quantified. The positive cells were counted according to standard methods (Encinas and Enikolopov, 2008) in a blinded fashion.
Statistical analysis
Data obtained from the NOR test and the number of DCX/PSA-NCAM-positive cells were analyzed using one-way analysis of variance whereas MWM data were analyzed using two-way analysis of variance. The Tukey’s post hoc test was used for comparison between groups. Statistical analyses were performed using GraphPad Prism software version 7.0 (GraphPad Software, San Diego, CA, USA). Differences between groups were considered statistically significant when P ≤ 0.05; and results are presented as the mean ± SE.
Results
Administration of Aβ1–42 aggregates induces memory and learning impairments
Administration of Aβ1–42 aggregates in the DG of hippocampus resulted in a significantly increased swimming time and inability to reach the deck (P > 0.05) compared to control animals in the Morris water maze test (Figure 1A). The control group showed a reduction in swimming time in different sessions (P < 0.05), confirming that their ability to retain spatial orientation and memory were not compromised, unlike animals in which Aβ1–42 damage was induced.
Figure 1.
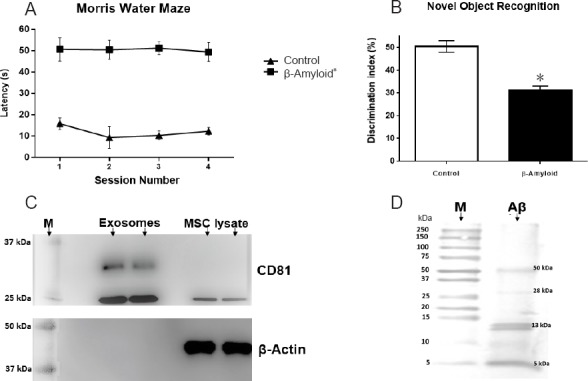
Effects of Aβ1–42 administration on mouse learning and memory and exosomal marker immunodetection.
(A) Morris water maze training phase was performed four times per day for 3 days, and test session was carried out 24 hours after the last training session. Mice of the beta-amyloid group exhibited significantly shorter latency to find the platform than mice of the normal control group (no surgery). Graph data represent time to reach platform (seconds). (B) The Novel Object Recognition test shows mouse’s ability to discriminate a familiar object from a new one. Mice of the beta-amyloid group have a lower discrimination ratio than mice of the normal control group. Data graph is interaction percentage with a new object. Both Morris water maze and Novel Object Recognition tests demonstrate the model establishment. (C) Representative western blot assay for exosomes isolation verification. CD81 marker is expressed in both ultracentrifugation pellet and MSC lysate; β-actin expression was detected only in MSC lysate. Gels were run in duplicate under no reducing conditions. (D) Representative western blot assay for Aβ aggregation. Aβ1–42 peptides can aggregate in different forms with a molecular weight from 13 to 50 kDa. However, fibrillar forms did not appear. All data are expressed as the mean ± SE; *P < 0.05, vs. normal control (control) group; n = 36 for β-amyloid group and n = 8 for normal control group. MSC: Mesenchymal stem cell; M: protein molecular weight marker; s: second.
Animals that were administered Aβ1–42 peptide aggregates demonstrated a lower DI in the NOR test (Figure 1B) compared with the animals in the normal control group (30% vs. 50% respectively; P < 0.05). Animals in the normal control group (DI 50%) showed a preference for the novel object. On the contrary, the animals that were administered Aβ1–42 (DI 30%) did not seem to have the ability to differentiate between the familiar and the novel objects.
Administration of exosomes increases learning abilities and memory loss
As reported by Thery et al. (2006), an average of 10 µg exosomes were obtained for 1 × 107 cells. Western blot results (Figure 1C) revealed the presence of exosomal marker CD81 (approximately at 25 kDa) in the pellets obtained by ultracentrifugation and in MSCs lysate. Beta actin (40 kDa) was only found in cell lysates since exosomes did not suffer lysis.
The MWM tests performed at 14 and 28 days after the administration of exosomes (Figure 2A) revealed that both exosomes and MSCs improved the performance of animals that were administered Aβ1–42 aggregates. Cells and nanovesicles alike optimized the animal’s capacity to remember the localization of the deck, which resulted in significantly shorter swimming periods.
Figure 2.

Effects of exosome administration on cognitive impairment of a mouse model of Alzheimer’s disease.
(A) Swimming time of mice during different sessions of Morris Water Maze test. Test sessions were carried out 14 and 28 days (sessions five and six respectively) after treatments. At session five of Morris water maze test, mice in the exosomes and MSC groups had better performance than mice in the AD group. Dotted line represents treatments. At session six of Morris Water Maze test, there was no difference between control, exosomes and MSC groups (*). (B) Novel Object Recognition test was performed after 14- and 28-day treatments. After 14 days, improvement in cognitive impairment, as indicated by discrimination ratio, was only found in the MSC group compared with the exosomes and AD groups. After 28-day treatment, there was no difference in discrimination ratio between exosomes, MSC, and normal control (control ) groups. These results suggest that exosome and MSC administration reduced cognitive impairment. Data are shown as the mean ± SE. #P < 0.05, vs. AD group (n = 4). MSC: Mesenchymal stem cell.
At 14 days after the administration of MSCs (5th session), mouse swimming time was shortened compared to that in the normal control group (P > 0.05). Swimming time was, but not significantly, shorter in the MSCs group than in the exosome group (P > 0.05). There was no significant difference in the swimming time between normal control and exosome groups (P > 0.05). However, at 28 days after treatment (6th session), the MSCs and exosome groups presented similar results to those of the control group (P < 0.05), while AD group did not show improvement in any session (P > 0.05) compared with the MSCs and exosome groups.
At 14 days after treatment, the MSCs group showed a greater improvement in the NOR test (Figure 2B) and the MWM test. Animals that were administered cells significantly increased their percentage of interaction with the novel object (40% vs. 60%, P < 0.05). Administration of exosomes did not present a significant improvement when compared to the AD group (42% vs. 40% respectively; P > 0.05). However, at 28 days after treatment, the exosome group showed a similar improvement to the MSCs group (P > 0.05). The exosome group increased the percentage of interaction time with the novel object. At 28 days (6th session) after treatment, both exosome and MSCs groups exhibited increased percentage of interaction time compared to the AD group (60%, 58%, and 40% respectively). There was no significant difference when compared to the normal control group (P > 0.05).
Administration of exosomes enhances neurogenesis in the SVZ
PSA-NCAM is a plasma cell membrane protein that grants the cells the ability to move, more evidently in neuroblasts. DCX is a protein associated with microtubules that enables proper neurite growth and development and is more abundant in cells in a more advanced developmental stage than that of neuroblasts. To evaluate if treatment with MSCs derived exosomes increases neurogenesis in the SVZ, 10 µm-thick brain sections were immuno-stained with anti PSA-NCAM and DCX antibodies (Figure 3A) and analyzed according to Encinas and Enikolopov (2008).
Figure 3.

Expression of neural precursor markers DCX and polysialylated neuronal cell adhesion molecule in subventricular zone.
(A) At 14 and 28 days after treatments, neurogenesis evaluation in the subventricular zone was performed with PSA-NCAM and DCX markers for neural precursors. Positive immunofluorescence staining for DCX and PSA-NCAM in the normal control, AD, exosome and MSC groups. Yellow arrows indicate DCX/PSA-NCAM double labeled cells in merge images. Scale bars: 25 μm. (B) Quantification of PSA-NCAM/DCX double labeled cells. After exosomes and MSC administration, the number of PSA-NCAM/DCX positive cells increased. These results suggest that exosomes and MSC administration increase neurogenesis in the subventricular zone. Original magnification 40×. Data are represented as the mean ± SE. *P < 0.05, vs. control group; †P < 0.05, vs. AD group at 14 days; §P < 0.05, vs. AD group at 28 days. DCX: Doublecortin; MSC: mesenchymal stem cell; AD: Alzheimer’s disease.
Results of immunofluorescences analysis are shown in Figure 3B. The number of neural precursors in the SVZ was grossly evident. The number of immunoreactive cells in the AD group was smaller than that in the normal control, exosome, and MSCs groups. At 14 days, both MSCs and exosome groups did not show significant differences in the number of immunoreactive cells compared to the normal control group (P > 0.05). The number of immunoreactive cells in the control, exosome, and MSCs groups was significantly increased than that in the AD group (P < 0.05). At 28 days, exosome and MSCs groups presented an increment of more than 50% in the number of immunoreactive cells compared to 14 days after administration of both treatments (P < 0.05). At 28 days, both exosome and MSCs groups displayed a statistically significant increase in the number of immunoreactive cells compared with the normal control group (P < 0.05). The number of immunoreactive cells increased approximately 30–40% respectively compared to the normal control group (P < 0.05). At 28 days after treatment, the number of immunoreactive cells in the control, exosome and MSCs groups was significantly increased than that in the AD group (P < 0.05). There was no significant difference in the number of immunoreactive cells between exosome and MSCs groups at 14 or 28 days after treatment (P > 0.05).
Discussion
Cognitive decline in animal models of AD has been attributed to aggregation of Aβ peptides that triggers events such as inflammation and oxidative stress and the events that negatively affect overall neuron survival and proper synaptic transmission that effectively leads to cognitive deficit and memory loss (Frozza et al., 2018). Our results showed that administration of Aβ1–42 peptide aggregates leads to neurological alterations mainly associated with learning and memory, as evaluated by the MWM and NOR tests. Results of the behavioral test showed an increase in swimming time and a limited ability to differentiate novel objects in the animals that were administered Aβ. These results are similar to those previously reported and are indicative of the proper implementation of the disease model (Nitta et al., 1994; Sipos et al., 2007; Takeda et al., 2009; Balducci et al., 2010; Kim et al., 2016).
The animal model of AD by administration of Aβ peptides was first described by Nitta et al. in 1994, in that time they described the cholinergic pyramidal neurons as the cells most affected by the toxicity of these peptides. Additionally, it has been demonstrated that Aβ peptides inhibit LTP response (the main mechanism responsible for the generation and preservation of memory) through hyperpolarization of neuron cell membrane and thus the opening of channels that allow the entrance of Ca2+ and K+ (Small et al., 2001; LaFerla and Green, 2012; Eslamizade et al., 2015). This model emulates the relevant characteristics of AD, such as memory loss and cognitive deficits; and it is at the present time a useful model when studying AD (Morgese et al., 2015; Sharma et al., 2016; Kasza et al., 2017). Taking all our results into consideration, we conclude that the administration of Aβ1–42 aggregates induces memory loss and cognitive deterioration. Currently, there is no effective treatment for AD.
That is why it is important to halt, or at least mitigate the neurological deterioration associated with the disease. Cell therapy, specifically with MSCs has emerged as a therapeutic strategy for the treatment of AD (Kim et al., 2013; Cui et al., 2017; Gobshtis et al., 2017). Studies have shown that MSCs exert their therapeutic activity mainly by secreting neurotrophic factors such as brain-derived neurotrophic factor, vascular endothelial growth factor, and basic fibroblast growth factor as well as by triggering the production of anti-inflammatory cytokines such as interleukin-10 and transforming growth factor-β (Hasan et al., 2017; Matthay et al., 2017; Kim et al., 2018). Recently, MSCs have been reported to stimulate the production of miRNA that have the ability to interact with neurogenic niches giving rise to the proliferation and differentiation of neural precursors (Lee et al., 2014; Gong et al., 2017). By means of this secretory activity, MSCs can protect neurons against apoptosis, mitigate the inflammatory response initiated by activating the damaged astrocytes and microglia that have an overall local anti-inflammatory effect, and promote the proliferation of neural precursors in the neurogenic niches that could originate new neurons or glial cells (Volkman and Offen, 2017; Reza-Zaldivar et al., 2018). In addition, MSCs have been reported to treat neurological disorders in different aninal models of the disease (Yan et al., 2014; Schwerk et al., 2015; Guo et al., 2017; Cunningham et al., 2018). There is also evidence that the therapeutic effect elicited by MSCs comes not only from a cell replacement but also cell ability to secrete substances such as exosomes (Nakano et al., 2016). Exosomes have been shown to be one of the most important components of the secretory activity of MSCs, and indeed, they display similar effects to those seen in MSCs, however, the mechanisms by which exosomes exert their therapeutic effects have not been completely described (Yun et al., 2013; Zhang et al., 2016; Cao et al., 2017; Yang et al., 2017a, b).
This study demonstrated that MSCs-derived exosomes, in a similar way as seen with MSCs have the capacity to improve the cognitive deficits associated with administration of Aβ1–42 aggregates as well as potentiate neurogenesis in the SVZ. Behavioral tests performed at 14 days after treatments showed that both MSCs and exosomes improved the performance of mice with AD. In MWM, administration of MSCs and MSC-derived exosomes substantially improved the ability of AD mice to find the deck, possibly by increasing its capacity to remember the localization of the deck, which resulted in significantly shorter swimming periods, while AD group had the longest swimming time in all sessions. In NOR test, both MSCs and MSC-derived exosome treatments increased the mice’s ability to differentiate a novel object from a familiar one, unlike animals from the AD group, which spend the same time to identify both objects. In this study, at 14 days after MSCs and MSC-derived exosome treatments, a better outcome was observed in mice receiving MSC treatment than those receiving MSC-derived exosome. This is attributable to the environmental stress that modifies the secretion capacity of MSCs, leading to a better outcome. The secretion of exosomes is considered as the mechanism of cellular adaptation. Exosome composition and secretion will depend on the environment in which the cell is located. An example of the mechanism of cellular adaptation was reported by Harting et al. (2018) who reported an MSC response to an inflammatory stimulus with exosomes production with a high anti-inflammatory capacity in MSCs co-cultured with extracts of ischemic brain tissue.
However, all these results are in accordance with previous studies that describe the improvement of cognitive functional capacities in different animal models of brain damage (Yun et al., 2013; Schwerk et al., 2015; Otero-Ortega et al., 2018). In a study performed by Li et al. (2017) in a traumatic brain injury model, it was found that MSC-derived exosomes disrupted the polarization of M1 microglia and triggered their transition to M2 phenotype, which dramatically reduced inflammation by enhancing the levels of anti-inflammatory factors that induced the functional recovery. Nakano et al. (2016) showed that treatment with exosomes displayed a neuroprotective effect against oxidative stress, in addition to an expansion in neuritic density in neurons as well as improvement of neurological capacities altered by administration of streptozotocin. Additionally, in this study, we also evaluated the effect of administration of exosomes and MSCs in the promotion of neurogenesis in the subventricular zone. In agreement with the improvement in performance seen in the behavioral test, mice treated with MSCs and MSC-derived exosomes increased the numbers of PSA-NCAM and DCX positive cells in the subventricular zone in both treatment groups compared to the AD group. At 14 days after treatment, MSCs and exosome groups showed similar numbers of PSA-NCAM and DCX positive cells to those in the normal control group. MSCs and exosome groups had a similar number of positive cells and there was no significant difference between these groups. At 28 days after treatment, exosome and MSCs groups exhibited significantly increased number of positive cells than normal control group. Both treatment groups had statistically significant increases in positive cells compared to the AD group. The AD group showed fewer positive staining cells than the control, exosome, and MSCs groups.
MSCs administration led to higher numbers of PSA-NCAM and DCX positive cells than MSC-derived exosome administration. This could be the reason of adaptation of MSCs to the stress environment as described by Harting et al. (2018). Our results show that MSCs and MSC-derived exosomes can effectively promote neurogenesis in the subventricular zone in this model of Alzheimer’s disease. Recent studies have reported that administration of exosomes increases the number of new neurons in the neurogenic niches (in the subventricular zone and dentate gyrus), however, the precise molecular and cellular mechanisms remain to be described (Doeppner et al., 2015; Xin et al., 2017b; Han et al., 2018). Some studies provide evidence that one of the mechanisms by which exosomes interact with neurogenic niches is by the transference of miRNAs to precursor neural precursor cells, triggering neural remodeling events, neurogenesis, angiogenesis and synaptogenesis (Cheng et al., 2018). Yang et al. (2017a) showed that exosomes modified to contain and deliver miRNAs are capable of interacting with resident cells in damaged areas and promote neurogenesis. miR-133b enriched exosomes triggered the release of exosomes by astrocytes, potentiated neutritic growth, and promoted neurovascular plasticity (Xin et al., 2013, 2017).
Taking all into consideration, these results confirm the therapeutic benefit conferred by MSCs derived exosomes in promoting subventricular zone neurogenesis, in addition to regaining functional cognitive abilities affected by administration of Aβ1–42 peptide aggregates. Full understanding of neuroplastic events and their impact in neurological recovery can be effectively translated into therapeutic strategies with positive clinical effects in neurodegenerative disorders.
Conclusions
This study proved that MSC-derived exosomes can enhance neurogenesis and restore cognitive function lost by administration of Aβ1–42 aggregates. These findings further support the role of exosomes as a possible therapeutic strategy for the treatment of chronic degenerative neurologic diseases.
Notwithstanding the positive evidence obtained, it is necessary to fully understand the precise mechanisms by which exosomes contribute to the activation of neurogenic niches, and how potentiating neurogenesis confers recovery of cognitive capacities that become compromised in neurological diseases, alongside of the main exosomal components responsible for these effects.
Additional file: Open peer review report 1 (113.2KB, pdf) .
Footnotes
Conflicts of interest: None declared.
Financial support: The present work was sponsored by CONACYT scholarship #487713 and by Fondo Mixto de Ciencia y Tecnología del Estado de Jalisco grant JAL-2014-0-250508.
Institutional review board statement: All procedures and experiments were done according to national (NOM-062-ZOO-1999) and international Guide for Care and Use of Laboratory Animals with the institutional approval by the ethics committee (approval No. CICUAL 2016-011) on April 25, 2016.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Baruh Polis, Bar-Ilan University, Medical School, Israel.
Funding: The present work was sponsored by CONACYT scholarship #487713 and by Fondo Mixto de Ciencia y Tecnología del Estado de Jalisco grant JAL-2014-0-250508.
P-Reviewer: Polis B; C-Editor: Zhao M; S-Editor: Li CH; L-Editor: Song LP; T-Editor: Liu XL
References
- 1.Balducci C, Beeg M, Stravalaci M, Bastone A, Sclip A, Biasini E, Tapella L, Colombo L, Manzoni C, Borsello T, Chiesa R, Gobbi M, Salmona M, Forloni G. Synthetic amyloid-beta oligomers impair long-term memory independently of cellular prion protein. Proc Natl Acad Sci U S A. 2010;107:2295–2300. doi: 10.1073/pnas.0911829107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bevins RA, Besheer J. Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat Protoc. 2006;1:1306–1311. doi: 10.1038/nprot.2006.205. [DOI] [PubMed] [Google Scholar]
- 3.Cao N, Liao T, Liu J, Fan Z, Zeng Q, Zhou J, Pei H, Xi J, He L, Chen L, Nan X, Jia Y, Yue W, Pei X. Clinical-grade human umbilical cord-derived mesenchymal stem cells reverse cognitive aging via improving synaptic plasticity and endogenous neurogenesis. Cell Death Dis. 2017;8:e2996. doi: 10.1038/cddis.2017.316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cheng X, Zhang G, Zhang L, Hu Y, Zhang K, Sun X, Zhao C, Li H, Li YM, Zhao J. Mesenchymal stem cells deliver exogenous miR-21 via exosomes to inhibit nucleus pulposus cell apoptosis and reduce intervertebral disc degeneration. J Cell Mol Med. 2018;22:261–276. doi: 10.1111/jcmm.13316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–289. doi: 10.1146/annurev-cellbio-101512-122326. [DOI] [PubMed] [Google Scholar]
- 6.Cui Y, Ma S, Zhang C, Cao W, Liu M, Li D, Lv P, Xing Q, Qu R, Yao N, Yang B, Guan F. Human umbilical cord mesenchymal stem cells transplantation improves cognitive function in Alzheimer’s disease mice by decreasing oxidative stress and promoting hippocampal neurogenesis. Behav Brain Res. 2017;320:291–301. doi: 10.1016/j.bbr.2016.12.021. [DOI] [PubMed] [Google Scholar]
- 7.Cunningham CJ, Redondo-Castro E, Allan SM. The therapeutic potential of the mesenchymal stem cell secretome in ischaemic stroke. J Cereb Blood Flow Metab. 2018;38:1276–1292. doi: 10.1177/0271678X18776802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.de Godoy MA, Saraiva LM, de Carvalho LRP, Vasconcelos-Dos-Santos A, Beiral HJV, Ramos AB, Silva LRP, Leal RB, Monteiro VHS, Braga CV, de Araujo-Silva CA, Sinis LC, Bodart-Santos V, Kasai-Brunswick TH, Alcantara CL, Lima APCA, da Cunha-E Silva NL, Galina A, Vieyra A, De Felice FG, Mendez-Otero R, Ferreira ST. Mesenchymal stem cells and cell-derived extracellular vesicles protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-β oligomers. J Biol Chem. 2018;293:1957–1975. doi: 10.1074/jbc.M117.807180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.de Jong OG, Verhaar MC, Chen Y, Vader P, Gremmels H, Posthuma G, Schiffelers RM, Gucek M, van Balkom BW. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J Extracell Vesicles. 2012;1 doi: 10.3402/jev.v1i0.18396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.De Strooper B, Karran E. The cellular phase of Alzheimer’s disease. Cell. 2016;164:603–615. doi: 10.1016/j.cell.2015.12.056. [DOI] [PubMed] [Google Scholar]
- 11.Doeppner TR, Herz J, Görgens A, Schlechter J, Ludwig AK, Radtke S, de Miroschedji K, Horn PA, Giebel B, Hermann DM. Extracellular vesicles improve post-stroke neuroregeneration and prevent postischemic immunosuppression. Stem Cells Transl Med. 2015;4:1131–1143. doi: 10.5966/sctm.2015-0078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Donders R, Bogie JFJ, Ravanidis S, Gervois P, Vanheusden M, Marée R, Schrynemackers M, Smeets HJM, Pinxteren J, Gijbels K, Walbers S, Mays RW, Deans R, Van Den Bosch L, Stinissen P, Lambrichts I, Gyselaers W, Hellings N. Human Wharton’s Jelly-derived stem cells display a distinct immunomodulatory and proregenerative transcriptional signature compared to bone marrow-derived stem cells. Stem Cells Dev. 2018;27:65–84. doi: 10.1089/scd.2017.0029. [DOI] [PubMed] [Google Scholar]
- 13.Drommelschmidt K, Serdar M, Bendix I, Herz J, Bertling F, Prager S, Keller M, Ludwig AK, Duhan V, Radtke S, de Miroschedji K, Horn PA, van de Looij Y, Giebel B, Felderhoff-Müser U. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain Behav Immun. 2017;60:220–232. doi: 10.1016/j.bbi.2016.11.011. [DOI] [PubMed] [Google Scholar]
- 14.Encinas JM, Enikolopov G. Identifying and quantitating neural stem and progenitor cells in the adult brain. Methods Cell Biol. 2008;85:243–272. doi: 10.1016/S0091-679X(08)85011-X. [DOI] [PubMed] [Google Scholar]
- 15.Eslamizade MJ, Saffarzadeh F, Mousavi SM, Meftahi GH, Hosseinmardi N, Mehdizadeh M, Janahmadi M. Alterations in CA1 pyramidal neuronal intrinsic excitability mediated by Ih channel currents in a rat model of amyloid beta pathology. Neuroscience. 2015;305:279–292. doi: 10.1016/j.neuroscience.2015.07.087. [DOI] [PubMed] [Google Scholar]
- 16.Ferreira AC, Sousa N, Bessa JM, Sousa JC, Marques F. Metabolism and adult neurogenesis: Towards an understanding of the role of lipocalin-2 and iron-related oxidative stress. Neurosci Biobehav Rev. 2018;95:73–84. doi: 10.1016/j.neubiorev.2018.09.014. [DOI] [PubMed] [Google Scholar]
- 17.Frozza RL, Lourenco MV, De Felice FG. Challenges for Alzheimer’s disease therapy: insights from novel mechanisms beyond memory defects. Front Neurosci. 2018;12:37. doi: 10.3389/fnins.2018.00037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gobshtis N, Tfilin M, Wolfson M, Fraifeld VE, Turgeman G. Transplantation of mesenchymal stem cells reverses behavioural deficits and impaired neurogenesis caused by prenatal exposure to valproic acid. Oncotarget. 2017;8:17443–17452. doi: 10.18632/oncotarget.15245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gómez-Isla T, Hollister R, West H, Mui S, Growdon JH, Petersen RC, Parisi JE, Hyman BT. Neuronal loss correlates with but exceeds neurofibrillary tangles in Alzheimer’s disease. Ann Neurol. 1997;41:17–24. doi: 10.1002/ana.410410106. [DOI] [PubMed] [Google Scholar]
- 20.Gong M, Yu B, Wang J, Wang Y, Liu M, Paul C, Millard RW, Xiao D-S, Ashraf M, Xu M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget. 2017;8:45200–45212. doi: 10.18632/oncotarget.16778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Guo S, Zhen Y, Wang A. Transplantation of bone mesenchymal stem cells promotes angiogenesis and improves neurological function after traumatic brain injury in mouse. Neuropsychiatr Dis Treat. 2017;13:2757–2765. doi: 10.2147/NDT.S141534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Han Y, Seyfried D, Meng Y, Yang D, Schultz L, Chopp M, Seyfried D. Multipotent mesenchymal stromal cell-derived exosomes improve functional recovery after experimental intracerebral hemorrhage in the rat. J Neurosurg. 2018:1–11. doi: 10.3171/2018.2.JNS171475. [DOI] [PubMed] [Google Scholar]
- 23.Harris VK, Stark J, Vyshkina T, Blackshear L, Joo G, Stefanova V, Sara G, Sadiq SA. Phase I trial of intrathecal mesenchymal stem cell-derived neural progenitors in progressive multiple sclerosis. EBioMedicine. 2018;29:23–30. doi: 10.1016/j.ebiom.2018.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Harting MT, Srivastava AK, Zhaorigetu S, Bair H, Prabhakara KS, Toledano Furman NE, Vykoukal JV, Ruppert KA, Cox CS, Jr, Olson SD. Inflammation-stimulated mesenchymal stromal cell-derived extracellular vesicles attenuate inflammation. Stem Cells. 2018;36:79–90. doi: 10.1002/stem.2730. [DOI] [PubMed] [Google Scholar]
- 25.Hasan A, Deeb G, Rahal R, Atwi K, Mondello S, Marei HE, Gali A, Sleiman E. Mesenchymal stem cells in the treatment of traumatic brain injury. Front Neurol. 2017;8:28. doi: 10.3389/fneur.2017.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kasza Á, Penke B, Frank Z, Bozsó Z, Szegedi V, Hunya Á, Németh K, Kozma G, Fülöp L. Studies for improving a rat model of Alzheimer’s disease: icv administration of well-characterized β-amyloid 1-42 oligomers induce dysfunction in spatial memory. Molecules. 2017;22 doi: 10.3390/molecules22112007. pii: E2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kho AR, Kim OJ, Jeong JH, Yu JM, Kim HS, Choi BY, Suh SW, Chung TN. Administration of placenta-derived mesenchymal stem cells counteracts a delayed anergic state following a transient induction of endogenous neurogenesis activity after global cerebral ischemia. Brain Res. 2018;1689:63–74. doi: 10.1016/j.brainres.2018.03.033. [DOI] [PubMed] [Google Scholar]
- 28.Kim HO, Choi SM, Kim HS. Mesenchymal stem cell-derived secretome and microvesicles as a cell-free therapeutics for neurodegenerative disorders. Tissue Eng Regen Med. 2013;10:93–101. [Google Scholar]
- 29.Kim HY, Lee DK, Chung BR, Kim HV, Kim Y. Intracerebroventricular injection of amyloid-β peptides in normal mice to acutely induce Alzheimer-like cognitive deficits. J Vis Exp. 2016 doi: 10.3791/53308. doi: 10.3791/53308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kim YR, Ahn SM, Pak ME, Lee HJ, Jung DH, Shin YI, Shin HK, Choi BT. Potential benefits of mesenchymal stem cells and electroacupuncture on the trophic factors associated with neurogenesis in mice with ischemic stroke. Sci Rep. 2018;8:2044. doi: 10.1038/s41598-018-20481-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.LaFerla FM, Green KN. Animal models of Alzheimer disease. Cold Spring Harb Perspect Med. 2012;2 doi: 10.1101/cshperspect.a006320. pii: a006320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee HK, Finniss S, Cazacu S, Xiang C, Brodie C. Mesenchymal stem cells deliver exogenous miRNAs to neural cells and induce their differentiation and glutamate transporter expression. Stem Cells Dev. 2014;23:2851–2861. doi: 10.1089/scd.2014.0146. [DOI] [PubMed] [Google Scholar]
- 33.Li Y, Yang YY, Ren JL, Xu F, Chen FM, Li A. Exosomes secreted by stem cells from human exfoliated deciduous teeth contribute to functional recovery after traumatic brain injury by shifting microglia M1/M2 polarization in rats. Stem Cell Res Ther. 2017;8:198. doi: 10.1186/s13287-017-0648-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liang WS, Dunckley T, Beach TG, Grover A, Mastroeni D, Ramsey K, Caselli RJ, Kukull WA, McKeel D, Morris JC, Hulette CM, Schmechel D, Reiman EM, Rogers J, Stephan DA. Altered neuronal gene expression in brain regions differentially affected by Alzheimer’s disease: a reference data set. Physiol Genomics. 2008;33:240–256. doi: 10.1152/physiolgenomics.00242.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Matthay MA, Pati S, Lee JW. Concise review: mesenchymal stem (stromal) cells: biology and preclinical evidence for therapeutic potential for organ dysfunction following trauma or sepsis. Stem Cells. 2017;35:316–324. doi: 10.1002/stem.2551. [DOI] [PubMed] [Google Scholar]
- 36.Mitsialis SA, Kourembanas S. Stem cell-based therapies for the newborn lung and brain: Possibilities and challenges. Semin Perinatol. 2016;40:138–151. doi: 10.1053/j.semperi.2015.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Morgese MG, Colaianna M, Mhillaj E, Zotti M, Schiavone S, D’Antonio P, Harkin A, Gigliucci V, Campolongo P, Trezza V, De Stradis A, Tucci P, Cuomo V, Trabace L. Soluble beta amyloid evokes alteration in brain norepinephrine levels: role of nitric oxide and interleukin-1. Front Neurosci. 2015;9:428. doi: 10.3389/fnins.2015.00428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nakano M, Nagaishi K, Konari N, Saito Y, Chikenji T, Mizue Y, Fujimiya M. Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into damaged neurons and astrocytes. Sci Rep. 2016;6:24805. doi: 10.1038/srep24805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nitta A, Itoh A, Hasegawa T, Nabeshima T. Beta-amyloid protein-induced Alzheimer disease animal model. Neurosci Lett. 1994;170:63–66. doi: 10.1016/0304-3940(94)90239-9. [DOI] [PubMed] [Google Scholar]
- 40.Otero-Ortega L, Gomez de Frutos MC, Laso-Garcia F, Rodriguez-Frutos B, Medina-Gutierrez E, Lopez JA, Vazquez J, Diez-Tejedor E, Gutierrez-Fernandez M. Exosomes promote restoration after an experimental animal model of intracerebral hemorrhage. J Cereb Blood Flow Metab. 2018;38:767–779. doi: 10.1177/0271678X17708917. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 41.Phinney DG, Pittenger MF. Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells. 2017;35:851–858. doi: 10.1002/stem.2575. [DOI] [PubMed] [Google Scholar]
- 42.Piemontese L. New approaches for prevention and treatment of Alzheimer’s disease: a fascinating challenge. Neural Regen Res. 2017;12:405–406. doi: 10.4103/1673-5374.202942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Qu J, Zhang H. Roles of mesenchymal stem cells in spinal cord injury. Stem Cells Int. 2017;2017:5251313. doi: 10.1155/2017/5251313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Reiss AB, Arain HA, Stecker MM, Siegart NM, Kasselman LJ. Amyloid toxicity in Alzheimer’s disease. Rev Neurosci. 2018;29:613–627. doi: 10.1515/revneuro-2017-0063. [DOI] [PubMed] [Google Scholar]
- 45.Reza-Zaldivar EE, Hernandez-Sapiens MA, Minjarez B, Gutierrez-Mercado YK, Marquez-Aguirre AL, Canales-Aguirre AA. Potential effects of MSC-derived exosomes in neuroplasticity in Alzheimer’s disease. Front Cell Neurosci. 2018;12:317. doi: 10.3389/fncel.2018.00317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Reza-Zaldívar EE, Sandoval-Avila S, Gutiérrez-Mercado YK, Vázquez-Méndez E, Canales-Aguirre AA, Esquivel-Solís H, Gómez-Pinedo U, Márquez-Aguirre AL. Human recombinant erythropoietin reduces sensorimotor dysfunction and cognitive impairment in rat models of chronic kidney disease. Neurologia. 2017 doi: 10.1016/j.nrl.2017.07.016. pii: S0213-4853(17)30276-1. [DOI] [PubMed] [Google Scholar]
- 47.Santos MA, Chand K, Chaves S. Recent progress in repositioning Alzheimer’s disease drugs based on a multitarget strategy. Future Med Chem. 2016;8:2113–2142. doi: 10.4155/fmc-2016-0103. [DOI] [PubMed] [Google Scholar]
- 48.Schwerk A, Altschuler J, Roch M, Gossen M, Winter C, Berg J, Kurtz A, Akyuz L, Steiner B. Adipose-derived human mesenchymal stem cells induce long-term neurogenic and anti-inflammatory effects and improve cognitive but not motor performance in a rat model of Parkinson’s disease. Regen Med. 2015;10:431–446. doi: 10.2217/rme.15.17. [DOI] [PubMed] [Google Scholar]
- 49.Segal-Gavish H, Karvat G, Barak N, Barzilay R, Ganz J, Edry L, Aharony I, Offen D, Kimchi T. Mesenchymal stem cell transplantation promotes neurogenesis and ameliorates autism related behaviors in BTBR mice. Autism Res. 2016;9:17–32. doi: 10.1002/aur.1530. [DOI] [PubMed] [Google Scholar]
- 50.Segerstrom SC. Personality and incident Alzheimer’s disease: theory, evidence, and future directions. J Gerontol B Psychol Sci Soc Sci. 2018 doi: 10.1093/geronb/gby063. doi: 10.1093/geronb/gby063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Selkoe DJ. Toward a comprehensive theory for Alzheimer’s disease. Hypothesis: Alzheimer’s disease is caused by the cerebral accumulation and cytotoxicity of amyloid beta-protein. Ann N Y Acad Sci. 2000;924:17–25. doi: 10.1111/j.1749-6632.2000.tb05554.x. [DOI] [PubMed] [Google Scholar]
- 52.Sharma S, Verma S, Kapoor M, Saini A, Nehru B. Alzheimer’s disease like pathology induced six weeks after aggregated amyloid-beta injection in rats: increased oxidative stress and impaired long-term memory with anxiety-like behavior. Neurol Res. 2016;38:838–850. doi: 10.1080/01616412.2016.1209337. [DOI] [PubMed] [Google Scholar]
- 53.Sipos E, Kurunczi A, Kasza A, Horváth J, Felszeghy K, Laroche S, Toldi J, Párducz A, Penke B, Penke Z. Beta-amyloid pathology in the entorhinal cortex of rats induces memory deficits: implications for Alzheimer’s disease. Neuroscience. 2007;147:28–36. doi: 10.1016/j.neuroscience.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 54.Small DH, Mok SS, Bornstein JC. Alzheimer’s disease and Abeta toxicity: from top to bottom. Nat Rev Neurosci. 2001;2:595–598. doi: 10.1038/35086072. [DOI] [PubMed] [Google Scholar]
- 55.Takeda S, Sato N, Niisato K, Takeuchi D, Kurinami H, Shinohara M, Rakugi H, Kano M, Morishita R. Validation of Abeta1-40 administration into mouse cerebroventricles as an animal model for Alzheimer disease. Brain Res. 2009;1280:137–147. doi: 10.1016/j.brainres.2009.05.035. [DOI] [PubMed] [Google Scholar]
- 56.Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol Chapter 3:Unit 322. 2006 doi: 10.1002/0471143030.cb0322s30. [DOI] [PubMed] [Google Scholar]
- 57.Timmers L, Lim SK, Arslan F, Armstrong JS, Hoefer IE, Doevendans PA, Piek JJ, El Oakley RM, Choo A, Lee CN, Pasterkamp G, de Kleijn DP. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res. 2007;1:129–137. doi: 10.1016/j.scr.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 58.Volkman R, Offen D. Concise review: mesenchymal stem cells in neurodegenerative diseases. Stem Cells. 2017;35:1867–1880. doi: 10.1002/stem.2651. [DOI] [PubMed] [Google Scholar]
- 59.Vorhees CV, Williams MT. Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat Protoc. 2006;1:848–858. doi: 10.1038/nprot.2006.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Xin H, Li Y, Liu Z, Wang X, Shang X, Cui Y, Zhang ZG, Chopp M. MiR-133b promotes neural plasticity and functional recovery after treatment of stroke with multipotent mesenchymal stromal cells in rats via transfer of exosome-enriched extracellular particles. Stem Cells. 2013;31:2737–2746. doi: 10.1002/stem.1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Xin H, Wang F, Li Y, Lu QE, Cheung WL, Zhang Y, Zhang ZG, Chopp M. Secondary release of exosomes from astrocytes contributes to the increase in neural plasticity and improvement of functional recovery after stroke in rats treated with exosomes harvested from microRNA 133b-overexpressing multipotent mesenchymal stromal cells. Cell Transplant. 2017a;26:243–257. doi: 10.3727/096368916X693031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Xin H, Katakowski M, Wang F, Qian JY, Liu XS, Ali MM, Buller B, Zhang ZG, Chopp M. MicroRNA cluster miR-17-92 cluster in exosomes enhance neuroplasticity and functional recovery after stroke in rats. Stroke. 2017b;48:747–753. doi: 10.1161/STROKEAHA.116.015204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Xiong Y, Mahmood A, Chopp M. Emerging potential of exosomes for treatment of traumatic brain injury. Neural Regen Res. 2017;12:19–22. doi: 10.4103/1673-5374.198966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Yan Y, Ma T, Gong K, Ao Q, Zhang X, Gong Y. Adipose-derived mesenchymal stem cell transplantation promotes adult neurogenesis in the brains of Alzheimer’s disease mice. Neural Regen Res. 2014;9:798–805. doi: 10.4103/1673-5374.131596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yang J, Zhang X, Chen X, Wang L, Yang G. Exosome mediated delivery of miR-124 promotes neurogenesis after ischemia. Mol Ther Nucleic Acids. 2017a;7:278–287. doi: 10.1016/j.omtn.2017.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yang Y, Ye Y, Su X, He J, Bai W, He X. MSCs-derived exosomes and neuroinflammation, neurogenesis and therapy of traumatic brain injury. Front Cell Neurosci. 2017b;11:55. doi: 10.3389/fncel.2017.00055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Yun HM, Kim HS, Park KR, Shin JM, Kang AR, il Lee K, Song S, Kim YB, Han SB, Chung HM, Hong JT. Placenta-derived mesenchymal stem cells improve memory dysfunction in an Aβ1-42-infused mouse model of Alzheimer’s disease. Cell Death Dis. 2013;4:e958. doi: 10.1038/cddis.2013.490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhang Y, Chopp M, Zhang ZG, Katakowski M, Xin H, Qu C, Ali M, Mahmood A, Xiong Y. Systemic administration of cell-free exosomes generated by human bone marrow derived mesenchymal stem cells cultured under 2D and 3D conditions improves functional recovery in rats after traumatic brain injury. Neurochem Int. 2016;111:69–81. doi: 10.1016/j.neuint.2016.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.