

Keywords: nerve regeneration, Parkinson's disease, mediodorsal thalamic nucleus, medial prefrontal cortex, pyramidal neurons, interneurons, discharge, neural regeneration
Abstract
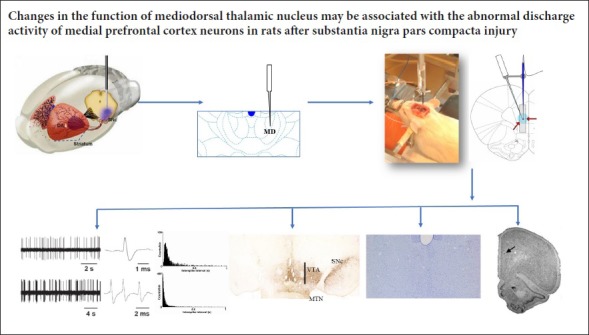
The dysfunction of the medial prefrontal cortex is associated with affective disorders and non-motor features in Parkinson’s disease. However, the exact role of the mediodorsal thalamic nucleus in the function of the prefrontal cortex remains unclear. To study the possible effects of the mediodorsal thalamic nucleus on the neurological function of the medial prefrontal cortex, a model of Parkinson’s disease was established by injecting 8 µg 6-hydroxydopamine into the substantia nigra compacta of rats. After 1 or 3 weeks, 0.3 µg ibotenic acid was injected into the mediodorsal thalamic nucleus of the midbrain. At 3 or 5 weeks after the initial injury, neuronal discharge in medial prefrontal cortex of rat brain was determined electrophysiologically. The numbers of dopamine-positive neurons and tyrosine hydroxylase immunoreactivity in substantia nigra compacta and ventral tegmental area were detected by immunohistochemical staining. Results demonstrated that after injury, the immunoreactivity of dopamine neurons and tyrosine hydroxylase decreased in the substantia nigra compacta and ventral tegmental areas of rats. Compared with normal medial prefrontal cortical neurons, at 3 and 5 weeks after substantia nigra compacta injury, the discharge frequency of pyramidal neurons increased and the discharge pattern of these neurons tended to be a burst-discharge, with an increased discharge interval. The discharge frequency of interneurons decreased and the discharge pattern also tended to be a burst-discharge, but the discharge interval was only higher at 3 weeks. At 3 weeks after the combined lesions, the discharge frequency, discharge pattern and discharge interval were restored to a normal level in pyramidal neurons and interneurons in medial prefrontal cortex. These findings have confirmed that mediodorsal thalamic nucleus is involved in regulating neuronal activities of the medial prefrontal cortex. The changes in the function of the mediodorsal thalamic nucleus may be associated with the abnormal discharge activity of the medial prefrontal cortex neurons after substantia nigra compacta injury. All experimental procedures were approved by the Institutional Animal Care and Use Committee of Xi’an Jiaotong University, China (approval No. XJTULAC2017-067) on August 26, 2017.
Chinese Library Classification No. R444; R363; R741
Introduction
Parkinson’s disease (PD) is mainly induced by the degeneration of the dopamine neurons in the midbrain substantia nigra pars compacta (SNc) (Zhou et al., 2017). This transforms the neural circuits of the basal ganglia, resulting in the impairment of control of voluntary movement in PD patients. Tremor, rigidity, slow and abnormal gait movement disorder are the main clinical symptoms of PD (Javidnia et al., 2017). In addition, there is widespread evidence of non-motor symptoms in PD patients, including cognitive impairment and emotional disorders such as depression and anxiety (Löhle et al., 2009). So far, the pathophysiological basis of non-motor symptoms in PD patients remains unclear.
The degenerative changes in the nigrostriatal pathway lead to morphological and/or functional abnormalities of many brain regions in patients and animal models of PD (Martinez and Peplow, 2018). The mediodorsal thalamic nucleus (MD) is a relay station for efferent and afferent projections between the prefrontal cortex (PFC) and subcortical regions. The circuit also involves the striatum, globus pallidum and ventral tegmental area. The MD plays an important role in emotion, cognition, learning and memory (Phillips et al., 2003; García-Cabezas et al., 2007; Mitchell et al., 2007). Clinical evidence suggests that MD damage caused by tumors, accidental injuries or degenerative diseases can lead to amnesia (Squire and Moore, 1979) or dementia (Victor et al., 1971) in humans. The cognitive impairment, sleep disturbances, depression and dysosphresia in PD patients are similar to the clinical features of MD dysfunction. Converging evidence has shown a direct relationship between the morphological and metabolic changes of the mediodorsal, or limbic, thalamus and depressive symptoms (Young et al., 2004; Li et al., 2010). Voxel-based morphometry analysis shows bilateral increased volume of the MD in patients with PD depression (Cardoso et al., 2009). Furthermore, a previous study by our research group indicated that degenerative changes in the nigrostriatal pathway induced hyperactivity of MD neurons (Li et al., 2013). These findings strongly suggest that MD dysfunction appears to be a pathophysiological and etiologic factor for non-motor symptoms such as depression, anxiety and cognitive impairment in PD patients.
The medial prefrontal cortex (mPFC) is a confined area, but functionally the most advanced for comprehensive cognitive processing. It does not directly accept fiber projection from outside the central nervous system (Riga et al., 2014). Neurons of the mPFC mainly include glutamate projection neurons (pyramidal neurons) and γ-aminobutyric acid inhibitory interneurons (Gabbott et al., 1997). Several studies have found that the MD thalamus is the core structure for processing nerve information in and out of different areas of the PFC (Cross et al., 2012). The reciprocal connections between the MD and mPFC suggest that the MD also participates in some important functions of the mPFC. For many species of animals, the mPFC plays an important role in cognition and mood regulation. The dysfunction of the mPFC is associated with affective disorders and non-motor features in PD (Cummings, 1992; Ridderinkhof et al., 2004). In spite of these early findings, the exact role that the MD plays in PFC function has yet to be revealed. It is worth noting that the functional changes of the MD may be associated with PFC dysfunction in PD patients. To investigate the possible impact of the MD on the neural function of the mPFC at different periods after the SNc was lesioned by 6-hydroxydopamine (6-OHDA), the effect of MD lesions on mPFC firing activity of SNc-lesioned rats was examined using electrophysiological techniques.
Materials and Methods
Animals
Ninety-seven healthy clean male Sprague-Dawley rats aged 9–11 weeks and weighing 260–350 g were provided by the Experimental Animal Center of Shaanxi Province, Xi’an, China (license No. SCXK [Shaan] 2018-001). The experiment was conducted on the following six groups of rats: (1) normal rats (n = 15); (2) rats 3 weeks after unilateral SNc lesion (n = 17); (3) rats 5 weeks after unilateral SNc lesion (n = 15); (4) rats with unilateral MD lesion (n = 15); (5) rats with combined MD and SNc lesions after 3 weeks (n = 18); and (6) rats with combined MD and SNc lesions after 5 weeks (n = 17). All experiment procedures followed the Guidance of the National Institute of Health on the Care and Use of Laboratory Animals (NIH publication No. 80-23, revised 1996) and the guidelines of the Institutional Animal Care and Use Committee of Xi’an Jiaotong University, China (approval No. XJTULAC2017-067) on August 26, 2017. Every effort was made to minimize the number and suffering of animals used in experiments.
Unilateral injection of 6-OHDA damages SNc
The method to cause unilateral SNc damage was the same as described previously (Fan et al., 2011). In brief, the rats were anesthetized with 4% chloral hydrate (400 mg/kg, intraperitoneally) and pretreated with desipramine (Sigma-Aldrich, St. Louis, MO, USA), dissolved in normal saline, 25 mg/kg, intraperitoneally, to protect norepinephrine neurons from 6-OHDA toxicity. Subsequently, the rats were fixed on a stereotaxic frame (SN-2N, Narishige, Tokyo, Japan). 6-OHDA (Sigma-Aldrich; dissolved in saline containing 0.01% ascorbic acid, 8 µg/4 µL) was injected into the right SNc (anteroposterior – 5.0–5.2 mm, lateral 1.9–2.0 mm, dorsal 7.1–7.2 mm; Paxinos and Watson, 2005). The injection was made at a rate of 0.5 µL/min using a glass micropipette connected to a 10 µL microsyringe. At 1 week after the operation, the rats were injected subcutaneously with apomorphine 0.05 mg/kg body weight (the apomorphine, Sigma-Aldrich, was dissolved in saline containing 0.01% ascorbic acid). Only rats that rotated to the unlesioned side more than 20 times every 5 minutes were selected for the subsequent experiment.
Unilateral injection of ibotenic acid damages MD
The rats were anesthetized with 4% chloral hydrate (400 mg/kg, intraperitoneally) and fixed on the stereotaxic frame (SN-2N, Narishige, Tokyo, Japan). The three-dimensional coordinates of MD were determined: anteroposterior −2.8 mm, lateral 0.5−0.6 mm, dorsal 5.3 mm (Paxinos and Watson, 2005). The 1 µL microsyringe (Hamilton, Reno, NV, USA), closely connected with the glass microelectrode, was used to slowly position the needle at the determined coordinates. Ibotenic acid (Sigma-Aldrich) was dissolved in PBS (10 µg/µL) and 0.3 µL was injected into MD at a rate of 0.1 µL/min. All rats were given 80 thousand units of penicillin, subcutaneously, before and after operations to prevent infection. For the groups with combined SNc and MD lesions, one group had ibotenic acid injected into the MD 1 week after SNc lesions, the other group 3 weeks after (Figure 1). A previous report showed that mechanical injury during drug injection had no effect on neuronal discharge activity (Wang et al., 2009).
Figure 1.
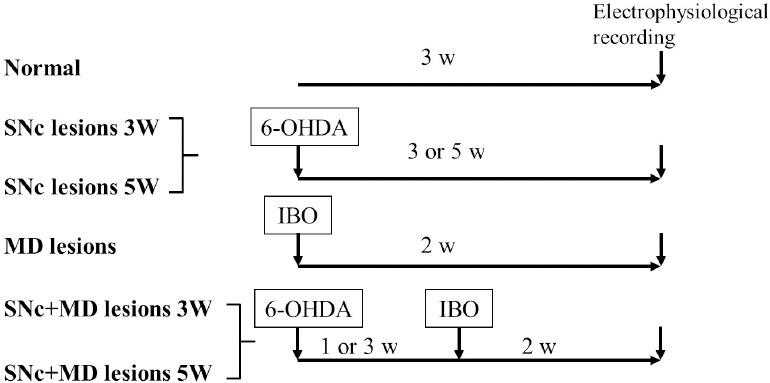
Experimental procedures for each group.
6-OHDA: 6-Hydroxydopamine; IBO: ibotenic acid; MD: mediodorsal thalamic nucleus; SNc: substantia nigra pars compacta; w: weeks.
Electrophysiological recordings
The electrophysiological recording of mPFC neurons was conducted at 3 and 5 weeks after SNc lesions (SNc-lesioned rats) and 2 weeks after MD lesions (MD-lesioned rats and double lesion rats). After 4% chloral hydrate anesthesia (400 mg/kg intraperitoneally), rats were fixed on a stereotaxic instrument. A heating pad was used to keep the rectal temperature at 37 ± 0.5°C during the whole electrocardiogram monitoring. A glass microelectrode (World Precision Instruments, Sarasota, FL, USA) (8–12 MΩ) filled with 2% pontamine blue was directed stereotaxically to the mPFC (anteroposterior 2.8–3.4, lateral 0.6–1.0, dorsal 1.5–3.9). The neuronal discharge was displayed on an oscilloscope (VC-11, Nihon Kohden, Tokyo, Japan) via a microelectrode amplifier and fed into a computer equipped with Spike 2 analysis software (Cambridge Electronic Design, Cambridge, UK) for online or offline analysis. The recorded mPFC pyramidal neurons displayed wide action potentials (> 1 ms) and irregular discharge patterns with burst activity (Tseng et al., 2006). The interneurons exhibited narrow action potentials (< 0.85 ms) (Constantinidis and Goldman-Rakic, 2002).
The discharge pattern was judged by comparing the inter-spike interval histogram (bin width = 4 ms) with the visual observation of the discharge sequence. According to the inter-spike interval histogram and discharge sequence, discharge patterns were divided into the following types: (1) regular, or slightly irregular, discharge, where the inter-spike interval histogram is approximately normally distributed; (2) irregular discharge, the inter-spike interval histogram is asymmetrically distributed; (3) burst discharge, the inter-spike interval histogram presents a significant positive skewed distribution. The coefficient of variation of the mean inter-spike interval (ISI) was also calculated (Breit et al., 2001). The determination of the discharge patterns and the calculation of all parameters required recording at least 500 continuous spikes.
Histological and immunohistochemical staining
Nissl staining of the mPFC and MD sections determines the location of the recording point and the extent of the MD lesion. After electrophysiological recording, the last recording site was marked by electrophoresis of pontamine blue (−20 µA, 15 minutes). Under an overdose of anesthesia, the rats were perfused with 200 mL saline through the heart, and then fixed with 4% paraformaldehyde 200 mL. The brain was taken and fixed for 4 hours in the same fixative, followed by 25% sucrose solution overnight. All samples were frozen and cut into 40 µm thick coronal sections using a cryostat. Briefly, sections were placed in xylene and gradient alcohol for dewaxing and dehydration, stained with 0.1% cresyl violet for 30 seconds, washed with distilled water, dehydrated and sealed.
To observe the degree of degeneration of dopamine neurons after 6-OHDA injection, tyrosine hydroxylase (TH) immunohistochemical staining was performed on frozen sections of the SNc. Briefly, sections were incubated with anti-TH polyclonal antibody (rabbit anti-rat, 1:1000, AB 152; Chemicon, San Diego, CA, USA) at 4°C for 48 hours, biotinylated goat anti-rabbit IgG (1:200; Chemicon) for 2 hours and avidin-biotin-peroxidase complex (1:100; Vector Laboratories, Burlingame, CA, USA) for 2 hours at room temperature. Next, the sections were exposed at room temperature for 5−10 minutes to 0.05% 3,3′-diaminobemidine (Sigma-Aldrich) solution used as the chromogen in subsequent chromogenic reactions. Finally, these sections were rinsed and mounted on gelatin slides. After dehydration, the slides were cleared in xylene and sealed. The staining was quantified under a microscope (BX51; Olympus, Tokyo, Japan). The count of the dopamine neuron bodies in the SNc and ventral tegmental area was carried out on two representative sections per rat. Only rats with a complete absence of TH-positive neurons of SNc were included in the statistical analysis.
Statistical analysis
All data are represented by the mean ± SEM or number (percent). SPSS 16.0 (SPSS, Chicago, IL, USA) was used for statistical analysis. One-way analysis of variance was used to compare the discharge frequency of each group. The comparison of mean ISI variation coefficient was performed by Mann-Whitney U test. The comparison of the discharge patterns distribution of neurons in different groups was performed by the chi-square test. The number of positive neurons of the SNc and ventral tegmental area in the lesioned and unlesioned sides of the SNc-lesioned rats was assessed using the Student’s paired t-test. P < 0.05 was considered statistically significant.
Results
Location and electrophysiological feature of the recording neurons
All electrophysiological recording sites were labeled by the glass microelectrode electrophoresis pontamine blue and confirmed by histological staining to be located in the mPFC (Figure 2A). The characteristics of all mPFC pyramidal neurons and interneurons recorded were consistent with previous results (Constantinidis and Goldman-Rakic, 2002; Tseng et al., 2006) (Figure 2B).
Figure 2.
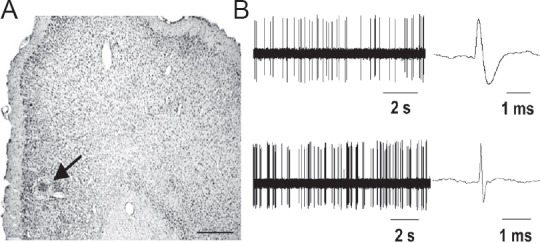
Location and electrophysiological characteristics of recorded neurons in the mPFC of rats.
(A) Electrophysiological recording sites (arrow) labeled by iontophoretically injected pontamine blue were shown by Nissl staining of mPFC. Scale bar: 400 µm. (B) Spontaneous electrical activity and action potential waveform of representative mPFC pyramidal neurons (upper) and interneurons (under) in normal rats. mPFC: Medial prefrontal cortex; s: seconds.
Feature of TH-immunoreactive neuron loss in the SNc and ventral tegmental area and the extent of MD lesion
6-OHDA-lesioned rats rotated to the unlesioned side more than 20 times every 5 minutes (53 ± 3 turns/5 min) after subcutaneous injection of apomorphine and completely lost TH immunoreactivity in the SNc (Figure 3A and B). TH-immunoreactive neurons in ventral tegmental area were also reduced by 41 ± 2% on the lesioned side (n = 12; P < 0.001; Student’s paired t test; Figure 3A and B). Nissl staining showed the location and range of ibotenic acid-induced lesions within the MD (Figure 3C and D).
Figure 3.
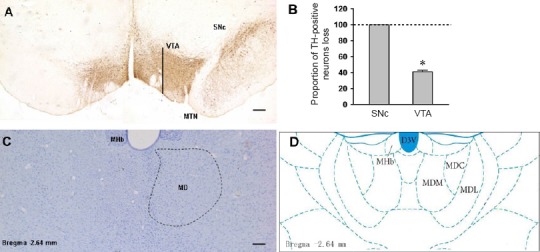
TH immunohistochemical staining of the SNc/VTA and Nissl staining of the lesion of the MD in rats.
(A) TH immunohistochemical staining images showed the comparison of dopamine neurons between the injected side (left) and non-injected side (right) in the 6-OHDA-lesioned rat. (B) Proportion of TH-immunoreactive neurons loss in SNc and VTA of the 6-OHDA-lesioned rat. (C) Nissl staining showed contrast with unlesioned side (right), damage effect of ibotenic acid injection on MD (left), and the location map of MD (D). Scale bars: 200 µm. D3V: Dorsal 3rd ventricle; MD: mediodorsal thalamic nucleus; MDM, MDC, MDL: mediodorsal thalamic nucleus, medial part, central part, lateral part; MHb: medial habenular nucleus; MTN: medial terminal nucleus; SNc: substantia nigra pars compacta; TH: tyrosine hydroxylase; VTA: ventral tegmental area.
mPFC neuronal activity in normal rats
In the normal group, the mean discharge frequency of pyramidal neurons was 1.26 ± 0.12 spikes/s (n = 59; Table 1). Most pyramidal neurons (75%) discharged irregularly, and only 25% were in bursts (Figure 4). The mean coefficient of variation of ISI was 1.2 ± 0.07 (Table 1). The mean discharge frequency of interneurons was 9.35 ± 1.16 spikes/s (n = 39; Table 1). The discharge patterns of interneurons were 51% irregular and 49% burst types (Figure 4). The mean coefficient of variation of ISI was 1.49 ± 0.07 (Table 1).
Table 1.
Comparision of discharge frequency and coefficients of variation of ISI of mPFC pyramidal neurons or interneurons between groups
| Type of neurons | Group | Number of neurons (number of rats) | Discharge frequency (spikes/s) | Coefficient of variation of ISI |
|---|---|---|---|---|
| Pyramidal cells | ||||
| Normal | 59 (6) | 1.26±0.12 | 1.20±0.07 | |
| SNc lesions 3 W | 47 (6) | 2.12±0.25* | 1.59±0.07** | |
| SNc lesions 5 W | 46 (5) | 2.81±0.41** | 1.32±0.04 | |
| MD lesions | 56 (6) | 2.36±0.27** | 1.74±0.11** | |
| SNc+MD lesions 3 W | 57 (6) | 1.58±0.18 | 1.59±0.06** | |
| SNc+MD lesions 5 W | 61 (7) | 2.69±0.33** | 1.72±0.06** | |
| Interneurons | ||||
| Normal | 39 (9) | 9.35±1.16 | 1.49±0.07 | |
| SNc lesions 3 W | 45 (11) | 6.25±0.64* | 1.63±0.04* | |
| SNc lesions 5 W | 36 (10) | 6.55±0.81* | 1.56±0.07 | |
| MD lesions | 44 (9) | 6.55±0.68* | 2.02±0.08** | |
| SNc+MD lesions 3 W | 43 (12) | 9.05±1.04 | 1.82±0.08** | |
| SNc+MD lesions 5 W | 37 (10) | 6.47±0.94* | 1.59±0.05 |
Data in discharge frequency and coefficient of variation of ISI are expressed as the mean ± SEM. *P < 0.05, **P < 0.01, vs. normal rats. One-way analysis of variance was used to compare the discharge frequency of each group. The comparison of coefficient of variation was performed by Mann-Whitney U test. mPFC: Medial prefrontal cortex; ISI: inter-spike interval; SNc: substantia nigra pars compacta; MD: mediodorsal thalamic nucleus; W: weeks.
Figure 4.
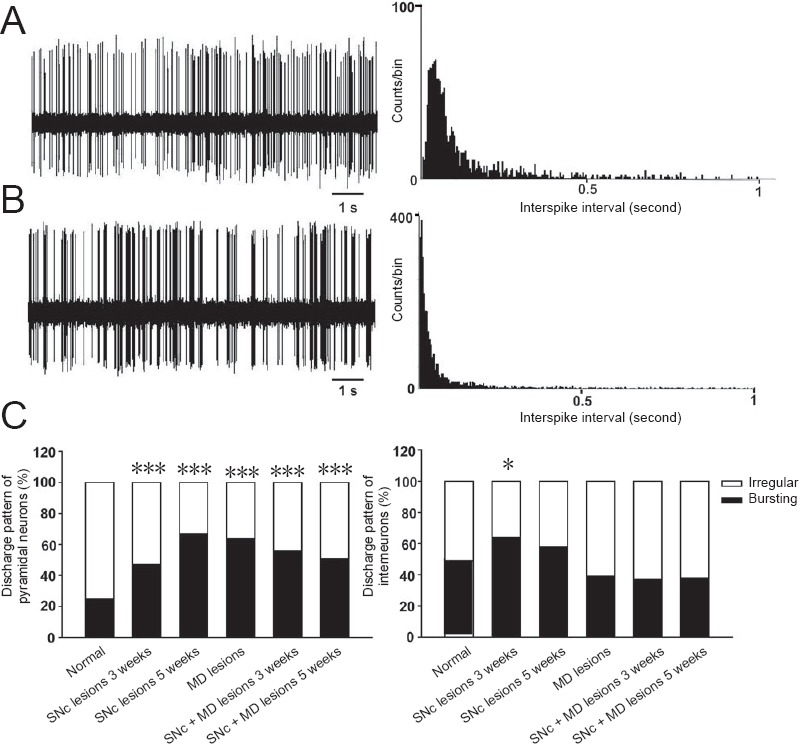
Discharge patterns of neurons recorded in the mPFC of rats.
Representative sample of spontaneous activity of medial prefrontal cortex neurons showed discharge pattern of neurons and corresponding inter-spike interval histogram (bin width = 4 ms) (A, irregular discharge; B, bursting discharge). (C) Distribution of discharge pattern of mPFC pyramidal neurons or interneurons in each group. *P < 0.05, ***P ≤ 0.001, vs. normal group (one-way analysis of variance). mPFC: Medial prefrontal cortex; SNc: substantia nigra pars compacta; MD: mediodorsal thalamic nucleus.
mPFC neuronal activity in SNc-lesioned rats
At 3 weeks after SNc lesions, the mean discharge frequency of mPFC pyramidal neurons significantly increased in the SNc-lesioned group (2.12 ± 0.25 spike/s) compared with normal group (n = 47, P < 0.05; one-way analysis of variance; Table 1). There was an increase in neurons exhibiting the burst-discharge pattern, with 53% of pyramidal neurons exhibiting irregular discharge and 47% exhibiting burst-discharge pattern (χ2 = 10.5, df = 1, P = 0.001; chi-square test; Figure 4). The mean coefficient of variation of ISI increased to 1.59 ± 0.07 (P < 0.001, Mann-Whitney U test; Table 1). However, the mean discharge frequency of interneurons decreased (6.25 ± 0.64 spike/s) compared with normal rats. Discharge patterns were more likely to be burst-discharge, with 36% of the interneurons exhibiting irregular discharge and 64% showing burst-discharge pattern (χ2 = 4.6, df = 1, P < 0.05; chi-square test; Figure 4). The mean coefficient of variation of ISI (1.63 ± 0.04) was significantly higher than that of normal group (P < 0.05, Mann-Whitney U test; Table 1), another indication that the interneurons of SNc-lesioned rats tended to exhibit a more burst-discharge pattern.
At 5 weeks after SNc lesions by 6-OHDA, the mean discharge frequency of pyramidal neurons (2.82 ± 0.41 spike/s) was significantly higher than that of the normal group (n = 46, P < 0.01; one-way analysis of variance; Table 1). Compared with normal rats, the discharge pattern was more prone to burst-discharge, 33% of pyramidal neurons showed irregular discharge, and 67% showed burst-discharge (χ2 = 35.5, df = 1, P < 0.001; chi-square test; Figure 4). The mean coefficient of variation of ISI (1.32 ± 0.04) was slightly increased compared with the normal group.
The mean discharge frequency of the interneurons significantly decreased (6.55 ± 0.81 spikes/s) compared with normal group (n = 36, P < 0.05; one-way analysis of variance; Table 1). The proportion of bursting neurons increased to 58% and that of irregular neurons was only 42%, but there was no significant difference from the normal group (Figure 3). The mean variation coefficient of ISI was unchanged at 1.56 ± 0.07.
mPFC neuronal activity in MD-lesioned rats
The mean discharge frequency of mPFC pyramidal neurons (2.36 ± 0.27 spikes/s) was significantly higher than that of the normal group 2 weeks after ibotenic acid-induced lesions of the MD (n = 46, P < 0.01; one-way analysis of variance; Table 1). The distribution of discharge patterns was 36% irregular and 64% bursting neurons. The proportion of bursting neurons significantly increased (χ2 = 30.8, df = 1, P < 0.001; chi-square test; Figure 4). The mean coefficient of variation of ISI significantly increased to 1.75 ± 0.11 compared with the normal group (P < 0.001, Mann-Whitney U test; Table 1).
The mean discharge frequency of interneurons (6.55 ± 0.68 spike/s) was significantly lower than that of the normal group (n = 44, P < 0.05; one-way analysis of variance; Table 1). Discharge patterns tended to be more of the irregular type, with burst-discharge pattern (39%) and irregular-discharge (61%) (Figure 4). The mean coefficient of variation of ISI was significantly higher than that of the normal group (P < 0.001; Mann-Whitney U test, Table 1).
mPFC neuronal activity in rats with SNc + MD combined lesions
Neuronal activity in the mPFC was recorded during the third and fifth weeks after combined SNc and MD lesions. At 3 weeks, the discharge frequency of mPFC pyramidal neurons slightly increased (1.58 ± 0.18 spikes/s; n = 57), but not significantly, compared with the normal group (Table 1). This suggests that the discharge frequency of the pyramidal neurons returned to normal levels. The percentage of burst-discharge neurons significantly increased to 56% (χ2 = 19.9, df = 1, P < 0.001; chi-square test; Figure 4). The mean coefficient of variation of ISI significantly increased to 1.59 ± 0.06 (P < 0.001, Mann-Whitney U test, Table 1). Furthermore, the mean discharge frequency of interneurons had no significant change compared with the normal group (Table 1). The discharge pattern, 62% irregular and 38% bursting neurons, indicated no significant difference from the normal group. The mean coefficient of variation of ISI (1.82 ± 0.08) was higher than that of the normal group (P < 0.001; Mann-Whitney U test; Table 1).
During the fifth week, the mean discharge frequency of pyramidal neurons in the mPFC significantly increased (2.69 ± 0.33 spike/s) compared with the normal group (n = 61, P < 0.01; one-way analysis of variance; Table 1). The discharge patterns of these neurons tended to show burst-discharge activities, with 49% showing irregular discharge patterns and 51% showing burst-discharge patterns (χ2 =14.3, df = 1, P < 0.001; chi-square test; Figure 4). The mean coefficient of variation of ISI (1.72 ± 0.06) was significantly increased compared with the normal group (P < 0.001, Mann-Whitney U test; Table 1).
The discharge frequency of interneurons (6.47 ± 0.94 spike/s) was significantly lower than that of the normal group (n = 37, P < 0.05; one-way analysis of variance; Table 1). The majority (62%) of interneurons showed irregular discharge patterns and 38% exhibited burst-discharge patterns (Figure 4). The mean variation coefficient of ISI was 1.59 ± 0.05, showing no significant difference compared with the normal group (Table 1).
Discussion
The main findings of this study show that: (1) During the third and fifth weeks after SNc lesion, the discharge frequency of mPFC pyramidal neurons noticeably increased, whereas the discharge frequency of the interneurons decreased. In both types of neurons there was an increase in tendency to exhibit burst-discharge pattern. (2) In rats with MD lesion, the discharge frequency of mPFC pyramidal neurons was dramatically increased, while the discharge frequency of interneurons was markedly decreased. (3) During the third week after the SNc + MD combined lesion, no significant change in the discharge frequency of mPFC pyramidal and interneurons was observed compared with normal rats. However, during the fifth week after the SNc + MD combined lesion, the discharge frequency and pattern of mPFC neurons were very similar to those in rats with SNc lesion only.
Spontaneous discharge activity of mPFC neurons after SNc lesions
The mPFC mainly receives dopamine afferent fibers from the ventral tegmental area and expresses two dopamine receptors, D1 and D2 (Walsh and Han, 2014). The electrical and chemical stimulation of the ventral tegmental area can inhibit the spontaneous discharge activity of PFC pyramidal neurons (Lewis and O’Donnell, 2000; Tseng et al., 2006). When 6-OHDA was used to destroy the medial forebrain bundle there was a reduction in the expression of D1 and D2 receptors in the PFC of rats (Wang et al., 2005). It can be inferred from the results of these studies that 6-OHDA injection into the SNc caused the degeneration and necrosis of dopamine neurons in the ventral tegmental area of rats. Simultaneously, the expression of dopamine receptors in mPFC decreased, leading to the disinhibition of pyramidal neurons.
The discharge frequency of mPFC interneurons in SNc-lesioned rats decreased, which may be due to the following reasons. First, dopamine fibers from the ventral tegmental area innervate interneurons expressing D1 and D2 receptors (Vincent et al., 1995; Bjorklund and Dunnett, 2007; Ranganath and Jacob, 2016). Electrophysiological studies have also demonstrated that chemical stimulation on the ventral tegmental area can excite fast-spiking interneurons of the PFC, activate D1 receptors, and also increase the excitability of fast-spiking interneurons (Seamans et al., 2001; Gorelova et al., 2002). We postulate that 6-OHDA toxicity results in a decreased number of dopamine neurons in the ventral tegmental area, as well as a reduction in D1 and D2 receptors expressed on the mPFC neurons. This would result in the reduced discharge frequency of interneurons. Second, the mPFC also accepts 5-hydroxytryptamine innervations from the midbrain raphe nuclei in mice (Muzerelle et al., 2016). 5-Hydroxytryptamine axons mainly form synapses with PFC interneurons in monkeys. In vitro electrophysiological experiments also demonstrated that 5-hydroxytryptamine-positive nerve fibers can depolarize most of the interneurons in the superficial layer of cortex (Foehring et al., 2002). The reduction of 5-hydroxytryptamine level in the mPFC might be another reason for the decreased discharge frequency of interneurons in the SNc-lesioned rats.
The discharge patterns of neurons are determined by their own cellular properties and neural modulation from other brain areas. Studies have shown that unilateral lesion in the nigrostriatal pathway resulted in the discharge frequency of 5-hydroxytryptamine-positive neurons increasing significantly in the dorsal raphe nucleus and median raphe nucleus of rats. There was also a tendency for neurons to burst-discharge (Wang et al., 2009). mPFC neurons receive a large number of 5-hydroxytryptamine-positive nerve fibers from the raphe nuclei, which makes mPFC neurons more prone to burst-discharge.
Spontaneous discharge activity of mPFC neurons after MD lesions
The MD is strongly associated with the PFC both in structure and function and represents the primary subcortical structure projected onto the PFC. Neuroanatomical studies have shown that the interactive projection between the MD and PFC is an excitatory glutamate fiber (Cruikshank et al., 2010; Barad et al., 2012). In a series of electron microscope studies, Kuroda et al. (1998) described that the nerve fibers projected by the MD mainly formed synaptic connections with pyramidal neurons in PFC. More recently, several workers have found that the γ-aminobutyric acid-positive local circuit interneurons also receive an input from the MD (Floresco and Grace, 2003; Kuroda et al., 2004; Rotaru et al., 2005). This suggests that MD cells could not only control the firing activity of PFC pyramidal neurons, but also modulate the activity of inhibitory γ-aminobutyric acid neurons. It has been demonstrated that axons produced by MD neurons directly act on parvalbumin interneurons and excite these neurons, thereby mediating the feed-forward inhibition of mPFC third-layer pyramidal neurons (Delevich et al., 2015). A previous study reported that at 10 to 15 days after the MD was damaged by kainic acid, the number of PFC neurons excited was reduced to 18% (Ferron et al., 1984). In this study, we speculated that MD lesions, caused by ibotenic acid, impair the excitatory input from the MD on mPFC interneuron activity. As a result, the interneurons showed a decreased mean discharge frequency compared with normal rats. As described before, these interneurons have a strong inhibitory regulation effect on pyramidal neurons via a local circuit. Thus, the mean discharge frequency of mPFC pyramidal neurons noticeably increased.
Spontaneous discharge activity of mPFC neurons after combined SNc and MD lesions
One of our most striking findings is that during the third week, after the combined lesions, no significant change in the discharge frequency of mPFC pyramidal and interneurons was observed. The MD mainly sends out fibers to the PFC, while the MD also receives fibers from the amygdala, olfactory cortex, basal ganglia and hippocampal formation (Kuroda and Price, 1991a, b; Ray et al., 1992; Timbie and Barbas, 2015). A study of the effects of converging projections to the PFC indicated that the MD may play a role in modulating the inhibitory effects of dopaminergic afferents from the ventral tegmental area on PFC neurons (Ferron et al., 1984). Stimulation of the median raphe nucleus inhibits the neuronal discharge of the mPFC induced by the MD (Mantz et al., 1990). The median raphe nucleus is one of the main sources of 5-hydroxytryptamine transmitter in the brain. It appears that the 5-hydroxytryptamine neurotransmitter system is a modulator for MD regulation of the mPFC. Further studies are necessary for a better understanding of the effect of morphological and functional changes of the dopamine and 5-hydroxytryptamine transmitter systems on the mediodorsal thalamic prefrontal loop in the Parkinsonian states.
The complex fiber connection and function of the MD in the brain makes it is easy to infer that the motor and non-motor features of PD have a causal relationship with MD function disorder. Until recently, however, little was known about the relationship between the MD and PD. This preliminary study has begun to investigate the relationship between the MD and PD non-motor symptoms and related brain regions. It will be necessary to obtain detailed knowledge of the influence of functional deficits of the MD and/or the disturbance of innervation of the PFC from the MD at different stages of PD.
In conclusion, the discharge frequency and pattern of pyramidal neurons and interneurons in mPFC were significantly different at 3 or 5 weeks after unilateral SNc lesions in rats. The discharge activities of the two kinds of neurons also changed significantly after the MD was damaged. However, in group 5, where the ipsilateral MD was damaged one week after the SNc lesions, the abnormal discharge of pyramidal neurons and interneurons in mPFC was reversed. However, damage to the MD 3 weeks after the SNc lesions did not reverse the abnormal discharge activity of either type of neuron. These results suggest that the MD is involved in regulating the neural activity of the mPFC. It suggests that the functional changes of the MD may be associated with the occurrence of mPFC dysfunction in PD. The functional defect of the MD or/and the effect of MD fiber innervation on the mPFC at different stages of PD may differ. These experimental results contribute to a functional explanation for neuropsychiatric symptoms such as depression, anxiety and mood disorders in PD patients. They may provide guidance in the treatment of PD non-motor symptoms and other neuropsychiatric diseases.
Additional file: Open peer review reports 1 (104.5KB, pdf) and 2 (103.1KB, pdf) .
Footnotes
Conflicts of interest: None declared.
Financial support: This work was supported by the Key Research Project of Science and Technology of Henan Province, China, No. 14B180007 (to LLF); and the Development Project of Science and Technology of Luoyang Municipality, Henan Province, China, No. 1401087A-5 (to LLF). The funding bodies played no role in the study design, in the collection, analysis and interpretation of data, in the writing of the paper, and in the decision to submit the paper for publication.
Institutional review board statement: The experimental protocol was approved by the Institutional Animal Care and Use Committee of Xi’an Jiaotong University, China (approval No. XJTULAC2017-067) on August 26, 2017.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewers: John Mitrofanis, University of Sydney, Australia; Jadwiga Szymura, University of Physical Education in Krakow, Poland.
Funding: This work was supported by the Key Research Project of Science and Technology of Henan Province of China, No. 14B180007 (to LLF); and the Development Project of Science and Technology of Luoyang Municipality of China, No. 1401087A-5 (to LLF).
P-Reviewers: Mitrofanis J, Szymura J; C-Editor: Zhao M; S-Editors: Yu J, Li CH; L-Editors: Dawes EA, Stow A, Qiu Y, Song LP; T-Editor: Liu XL
References
- 1.Barad Z, Shevtsova O, Arbuthnott GW, Leitch B. Selective loss of AMPA receptors at corticothalamic synapses in the epileptic stargazer mouse. Neuroscience. 2012;217:19–31. doi: 10.1016/j.neuroscience.2012.05.011. [DOI] [PubMed] [Google Scholar]
- 2.Bjorklund A, Dunnett SB. Dopamine neuron systems in the brain: an update. Trends Neurosci. 2007;30:194–202. doi: 10.1016/j.tins.2007.03.006. [DOI] [PubMed] [Google Scholar]
- 3.Breit S, Bouali-Benazzouz R, Benabid AL, Benazzouz A. Unilateral lesion of the nigrostriatal pathway induces an increase of neuronal activity of the pedunculopontine nucleus, which is reversed by the lesion of the subthalamic nucleus in the rat. Eur J Neurosci. 2001;14:1833–1842. doi: 10.1046/j.0953-816x.2001.01800.x. [DOI] [PubMed] [Google Scholar]
- 4.Cardoso EF, Maia FM, Fregni F, Myczkowski ML, Melo LM, Sato JR, Marcolin MA, Rigonatti SP, Cruz AC, Jr, Barbosa ER, Amaro E., Jr Depression in Parkinson’s disease: convergence from voxel-based morphometry and functional magnetic resonance imaging in the limbic thalamus. Neuroimage. 2009;47:467–472. doi: 10.1016/j.neuroimage.2009.04.059. [DOI] [PubMed] [Google Scholar]
- 5.Constantinidis C, Goldman-Rakic PS. Correlated discharges among putative pyramidal neurons and interneurons in the primate prefrontal cortex. J Neurophysiol. 2002;88:3487–3497. doi: 10.1152/jn.00188.2002. [DOI] [PubMed] [Google Scholar]
- 6.Cross L, Brown MW, Aggleton JP, Warburton EC. The medial dorsal thalamic nucleus and the medial prefrontal cortex of the rat function together to support associative recognition and recency but not item recognition. Learn Mem. 2012;20:41–50. doi: 10.1101/lm.028266.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cruikshank SJ, Urabe H, Nurmikko AV, Connors BW. Pathway-specific feedforward circuits between thalamus and neocortex revealed by selective optical stimulation of axons. Neuron. 2010;65:230–245. doi: 10.1016/j.neuron.2009.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cummings JL. Depression and Parkinson’s disease: a review. Am J Psychiatry. 1992;149:443–454. doi: 10.1176/ajp.149.4.443. [DOI] [PubMed] [Google Scholar]
- 9.Delevich K, Tucciarone J, Huang ZJ, Li B. The mediodorsal thalamus drives feedforward inhibition in the anterior cingulate cortex via parvalbumin interneurons. J Neurosci. 2015;35:5743–5753. doi: 10.1523/JNEUROSCI.4565-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fan LL, Zhang QJ, Liu J, Feng J, Gui ZH, Ali U, Zhang L, Hou C, Wang T, Hui YP, Sun YN, Wu ZH. In vivo effect of 5-HT7 receptor agonist on pyramidal neurons in medial frontal cortex of normal and 6-hydroxydopamine-lesioned rats: an electrophysiological study. Neuroscience. 2011;190:328–338. doi: 10.1016/j.neuroscience.2011.06.011. [DOI] [PubMed] [Google Scholar]
- 11.Ferron A, Thierry AM, Le Douarin C, Glowinski J. Inhibitory influence of the mesocortical dopaminergic system on spontaneous activity or excitatory response induced from the thalamic mediodorsal nucleus in the rat medial prefrontal cortex. Brain Res. 1984;302:257–265. doi: 10.1016/0006-8993(84)90238-5. [DOI] [PubMed] [Google Scholar]
- 12.Floresco SB, Grace AA. Gating of hippocampal-evoked activity in prefrontal cortical neurons by inputs from the mediodorsal thalamus and ventral tegmental area. J Neurosci. 2003;23:3930–3943. doi: 10.1523/JNEUROSCI.23-09-03930.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Foehring RC, van Brederode JF, Kinney GA, Spain WJ. Serotonergic modulation of supragranular neurons in rat sensorimotor cortex. J Neurosci. 2002;22:8238–8250. doi: 10.1523/JNEUROSCI.22-18-08238.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gabbott PL, Dickie BG, Vaid RR, Headlam AJ, Bacon SJ. Local-circuit neurones in the medial prefrontal cortex (areas 25, 32 and 24b) in the rat: morphology and quantitative distribution. J Comp Neurol. 1997;377:465–499. doi: 10.1002/(sici)1096-9861(19970127)377:4<465::aid-cne1>3.0.co;2-0. [DOI] [PubMed] [Google Scholar]
- 15.García-Cabezas MA, Rico B, Sánchez-González MA, Cavada C. Distribution of the dopamine innervation in the macaque and human thalamus. Neuroimage. 2007;34:965–984. doi: 10.1016/j.neuroimage.2006.07.032. [DOI] [PubMed] [Google Scholar]
- 16.Gorelova N, Seamans JK, Yang CR. Mechanisms of dopamine activation of fast-spiking interneurons that exert inhibition in rat prefrontal cortex. J Neurophysiol. 2002;88:3150–3166. doi: 10.1152/jn.00335.2002. [DOI] [PubMed] [Google Scholar]
- 17.Javidnia M, Kurd-Misto BT, Moussa CE. An update on clinical trials targeting human tauopathies. Clin Trials Degener Dis. 2017;2:66–76. [Google Scholar]
- 18.Kuroda M, Price JL. Synaptic organization of projections from basal forebrain structures to the mediodorsal thalamic nucleus of the rat. J Comp Neurol. 1991a;303:513–533. doi: 10.1002/cne.903030402. [DOI] [PubMed] [Google Scholar]
- 19.Kuroda M, Price JL. Ultrastructure and synaptic organization of axon terminals from brainstem structures to the mediodorsal thalamic nucleus of the rat. J Comp Neurol. 1991b;313:539–552. doi: 10.1002/cne.903130313. [DOI] [PubMed] [Google Scholar]
- 20.Kuroda M, Yokofujita J, Oda S, Price JL. Synaptic relationships between axon terminals from the mediodorsal thalamic nucleus and gamma-aminobutyric acidergic cortical cells in the prelimbic cortex of the rat. J Comp Neurol. 2004;477:220–234. doi: 10.1002/cne.20249. [DOI] [PubMed] [Google Scholar]
- 21.Lewis BL, O’Donnell P. Ventral tegmental area afferents to the prefrontal cortex maintain membrane potential ‘up’ states in pyramidal neurons via D(1) dopamine receptors. Cereb Cortex. 2000;10:1168–1175. doi: 10.1093/cercor/10.12.1168. [DOI] [PubMed] [Google Scholar]
- 22.Li W, Liu J, Skidmore F, Liu Y, Tian J, Li K. White matter microstructure changes in the thalamus in Parkinson disease with depression: A diffusion tensor MR imaging study. AJNR Am J Neuroradiol. 2010;31:1861–1866. doi: 10.3174/ajnr.A2195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li WJ, Xi Y, Zhang QQ, Hui YP, Xue L, Zhang H, Zhang LN, Yuan HF. Neuronal firing activity changes in mediodorsal thalamus nucleus in rats with Parkinson’s disease. Xi’an Jiaotong Daxue Xuebao: Yixue Ban. 2013;34:614–618. [Google Scholar]
- 24.Löhle M, Storch A, Reichmann H. Beyond tremor and rigidity: non-motor features of Parkinson’s disease. J Neural Transm (Vienna) 2009;116:1483–1492. doi: 10.1007/s00702-009-0274-1. [DOI] [PubMed] [Google Scholar]
- 25.Mantz J, Godbout R, Tassin JP, Glowinski J, Thierry AM. Inhibition of spontaneous and evoked unit activity in the rat medial prefrontal cortex by mesencephalic raphe nuclei. Brain Res. 1990;524:22–30. doi: 10.1016/0006-8993(90)90487-v. [DOI] [PubMed] [Google Scholar]
- 26.Martinez B, Peplow PV. Neuroprotection by immunomodulatory agents in animal models of Parkinson’s disease. Neural Regen Res. 2018;13:1493–1506. doi: 10.4103/1673-5374.237108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mitchell AS, Baxter MG, Gaffan D. Dissociable performance on scene learning and strategy implementation after lesions to magnocellular mediodorsal thalamic nucleus. J Neurosci. 2007;27:11888–11895. doi: 10.1523/JNEUROSCI.1835-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Muzerelle A, Scotto-Lomassese S, Bernard JF, Soiza-Reilly M, Gaspar P. Conditional anterograde tracing reveals distinct targeting of individual serotonin cell groups (B5-B9) to the forebrain and brainstem. Brain Struct Funct. 2016;221:535–561. doi: 10.1007/s00429-014-0924-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Phillips ML, Drevets WC, Rauch SL, Lane R. Neurobiology of emotion perception I: The neural basis of normal emotion perception. Biol Psychiatry. 2003;54:504–514. doi: 10.1016/s0006-3223(03)00168-9. [DOI] [PubMed] [Google Scholar]
- 30.Ranganath A, Jacob SN. Doping the Mind: Dopaminergic Modulation of Prefrontal Cortical Cognition. Neuroscientist. 2016;22:593–603. doi: 10.1177/1073858415602850. [DOI] [PubMed] [Google Scholar]
- 31.Ray JP, Russchen FT, Fuller TA, Price JL. Sources of presumptive glutamatergic/aspartatergic afferents to the mediodorsal nucleus of the thalamus in the rat. J Comp Neurol. 1992;320:435–456. doi: 10.1002/cne.903200403. [DOI] [PubMed] [Google Scholar]
- 32.Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004;306:443–447. doi: 10.1126/science.1100301. [DOI] [PubMed] [Google Scholar]
- 33.Riga D, Matos MR, Glas A, Smit AB, Spijker S, Van den Oever MC. Optogenetic dissection of medial prefrontal cortex circuitry. Front Syst Neurosci. 2014;8:230. doi: 10.3389/fnsys.2014.00230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rotaru DC, Barrionuevo G, Sesack SR. Mediodorsal thalamic afferents to layer III of the rat prefrontal cortex: synaptic relationships to subclasses of interneurons. J Comp Neurol. 2005;490:220–238. doi: 10.1002/cne.20661. [DOI] [PubMed] [Google Scholar]
- 35.Seamans JK, Gorelova N, Durstewitz D, Yang CR. Bidirectional dopamine modulation of GABAergic inhibition in prefrontal cortical pyramidal neurons. J Neurosci. 2001;21:3628–3638. doi: 10.1523/JNEUROSCI.21-10-03628.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Squire LR, Moore RY. Dorsal thalamic lesion in a noted case of human memory dysfunction. Ann Neurol. 1979;6:503–506. doi: 10.1002/ana.410060607. [DOI] [PubMed] [Google Scholar]
- 37.Timbie C, Barbas H. Pathways for emotions: specializations in the amygdalar, mediodorsal thalamic, and posterior orbitofrontal network. J Neurosci. 2015;35:11976–11987. doi: 10.1523/JNEUROSCI.2157-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tseng KY, Mallet N, Toreson KL, Le Moine C, Gonon F, O’Donnell P. Excitatory response of prefrontal cortical fast-spiking interneurons to ventral tegmental area stimulation in vivo. Synapse. 2006;59:412–417. doi: 10.1002/syn.20255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Victor M, Adams RD, Collins GH. The Wernicke-Korsakoff syndrome. A clinical and pathological study of 245 patients, 82 with post-mortem examinations. Contemp Neurol Ser. 1971;7:1–206. [PubMed] [Google Scholar]
- 40.Vincent SL, Khan Y, Benes FM. Cellular colocalization of dopamine D1 and D2 receptors in rat medial prefrontal cortex. Synapse. 1995;19:112–120. doi: 10.1002/syn.890190207. [DOI] [PubMed] [Google Scholar]
- 41.Walsh JJ, Han MH. The heterogeneity of ventral tegmental area neurons: Projection functions in a mood-related context. Neuroscience. 2014;282:101–108. doi: 10.1016/j.neuroscience.2014.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang Q, Wang PH, McLachlan C, Wong PT. Simvastatin reverses the downregulation of dopamine D1 and D2 receptor expression in the prefrontal cortex of 6-hydroxydopamine-induced Parkinsonian rats. Brain Res. 2005;1045:229–233. doi: 10.1016/j.brainres.2005.03.016. [DOI] [PubMed] [Google Scholar]
- 43.Wang S, Zhang QJ, Liu J, Wu ZH, Wang T, Gui ZH, Chen L, Wang Y. Unilateral lesion of the nigrostriatal pathway induces an increase of neuronal firing of the midbrain raphe nuclei 5-HT neurons and a decrease of their response to 5-HT(1A) receptor stimulation in the rat. Neuroscience. 2009;159:850–861. doi: 10.1016/j.neuroscience.2008.12.051. [DOI] [PubMed] [Google Scholar]
- 44.Young KA, Holcomb LA, Yazdani U, Hicks PB, German DC. Elevated neuron number in the limbic thalamus in major depression. Am J Psychiatry. 2004;161:1270–1277. doi: 10.1176/appi.ajp.161.7.1270. [DOI] [PubMed] [Google Scholar]
- 45.Zhou J, Bao LW, Liang J. Behavior improvement and inflammatory regulation in Parkinson’s disease rats after neural stem cell transplantation. Zhongguo Zuzhi Gongcheng Yanjiu. 2017;21:5299–5304. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.