

Abstract
Childhood and adolescent adversity are associated with a wide range of psychiatric disorders, including an increased risk for substance abuse. Despite this, the mechanisms underlying the ability of chronic stress during adolescence to alter reward signaling remains largely unexplored. Understanding how adolescent stress increases addiction-like phenotypes could inform the development of targeted interventions both before and after drug use. The current study examined how prolonged isolation stress, beginning during adolescence, affected behavioral and neuronal underpinnings to the response to cocaine in male and female mice. Adolescent-onset social isolation did not alter the ability of mice to learn an operant response for food, nor influence food self-administration or motivation for food on a progressive ratio schedule. However, male and female social isolation mice exhibited an increase in motivation for cocaine and cocaine seeking during a cue-induced reinstatement session. Additionally, we demonstrated that adolescent-onset social isolation increased cocaine-induced neuronal activation, as assessed by c-Fos expression, within the nucleus accumbens core and shell, ventral pallidum, dorsal bed nucleus of the stria terminalis, lateral septum and basolateral amygdala. Taken together, the present studies demonstrate that social isolation stress during adolescence augments the behavioral responses to cocaine during adulthood and alters the responsiveness of reward-related brain circuitry.
1. Introduction
Quality social interactions are fundamental to the mental health and well-being of an individual. A lack of social support during adolescence, either due to parental neglect, excessive bullying, or social exclusion, can increase the possibility of developing a psychiatric illness. These negative social experiences during adolescence increase the likelihood of alcohol and substance use during adolescence [1–5]. Along with increased use and abuse during adolescence, adolescent stress also increases the likelihood of substance use and addiction in adulthood [6–11]. However, it is difficult to determine the causal role of social stress in humans, as adolescent stress is often comorbid with other risk factors.
Preclinical models have shown that adolescent stress exposure can directly strengthen the effects of stimulants in adulthood. In rats, adolescent stress exposure leads to sensitized responses to acute administration of drugs, strengthened contextual drug associations, increased escalation of cocaine taking on an extended access schedule, and increased motivation for cocaine on a progressive ratio schedule [12–18]. All of these studies utilized chronic physical stressors such as social defeat, restraint, and/or forced swim. However, there is also evidence that emotional stress during adolescence can lead to alterations in cocaine addiction phenotypes. Social isolation rearing in rats leads to increased rates of acquisition of cocaine self-administration and increased motivation for cocaine [19–21]. Additionally, reintroduction of conspecific rats, in conjunction with cocaine exposure, results in greater cocaine conditioned reward [22]. That said, reintroduction to group housing after social isolation rearing could provide an additional stressor that may influence the rewarding effects of cocaine self-administration. Therefore, the aim of the current study was to examine the influence of social isolation during adolescence on cocaine taking and seeking behavior without the reintroduction of peers during adulthood.
In these studies we first established that social isolation rearing increases motivation for cocaine and reinstatement of cocaine seeking in both male and female mice. Additional studies examined how adolescent stress interacts with chronic cocaine self-administration to alter neural responses to cocaine. To achieve this, we examined neural activation patterns within the mesocorticolimbic system following an acute cocaine injection in mice that had a history of cocaine self-administration and adolescent stress exposure. Our results indicate stress-dependent alterations in drug-taking behavior, as well as associated changes in neural activation.
2. Materials and Methods
2.1. Animals.
This study used male and female c57/BL6 mice, exposed to social isolation beginning PND 21 through adulthood. Mice were randomly assigned isolation or group housing (2–5 animals per cage) at weaning (PND 21), then were all individually housed at the start of operant training (PND 60–80). All animals were housed in a temperature and humidity controlled animal care facility with a 12h light/dark cycle (lights on at 7:00 A.M.) and provided with cotton nestlets for enrichment. The Temple University Animal Care and Use Committee approved all procedures.
2.2. Drugs.
Cocaine was obtained from the National Institutes of Drug Abuse Drug Supply Program (Bethesda, MD) and dissolved in sterile 0.9% saline.
2.3. Operant Food Training.
At 8 weeks of age, mice were single-housed, food restricted to approximately 90% of their free feeding weight, and began operant food training. The animals were first trained to exhibit an operant response for sucrose pellets. They were placed in operant chambers (Med-Associates) and learned to spin a wheel manipulandum to receive a sucrose pellet. A white light, located over the active wheel, and 10s tone cue simultaneously occurred with administration of a pellet, followed by an 8s time-out with the house light off and no programmed consequences to wheel spins. Mice were food trained for 5 days on a fixed ratio schedule, where one wheel spin received a sucrose reward (FR1). After meeting criteria, they underwent 5 days of food training on a fixed ratio schedule where 5 wheel spins received a sucrose reward (FR5). During the first two sessions, the inactive wheel was immobilized. After that time, spinning on the inactive wheel has no programmed consequence. The mice were limited to a maximum of 50 pellets during the 60 min operant session. Following 10 days of food training, mice were tested for 1 day on a progressive ratio (PR) schedule, where the response requirement for each sucrose pellet increased until the subject did not fulfill the requirement. The response requirement was defined as R(i)=[5e0.2i-5] and the session ended if the animal took longer than 30 minutes to meet the requirement. The breakpoint is defined as the total number of responses or the final ratio completed.
2.4. Jugular Catheterization Surgery.
Before surgery, mice were anesthetized with 80 mg/kg ketamine and 12 mg/kg xylazine. An indwelling silastic catheter was inserted into the right jugular vein and sutured in place. The catheter was threaded subcutaneously over the shoulder blade and routed to a mesh back mount platform (Strategic Applications, Inc) to secure the placement. Following surgery, catheters were flushed daily with 0.1 mL of antibiotic (Timentin, 0.93 mg/ml) dissolved in heparinized saline and sealed with plastic obturators while not in use.
2.5. Cocaine Self-administration.
Mice were given 3–4 d to recover from surgery before the initiation of behavioral testing. The cocaine self-administration behavior was tested in 2-hour sessions (6 d per week) in the same chamber used for sucrose pellet self-administration. However, wheel responding now delivered an intravenous cocaine infusion (0.6 mg/kg/infusion), paired with the same compound cue, under the same fixed ratio (FR1) schedule as food training. After 10 days of a fixed ratio reward schedule, the mice were tested for 1 day on a progressive ratio (PR) schedule, where the response requirement for each infusion increased until the subject did not fulfill the requirement. The response requirement was defined as R(i)=[5e0.2i-5] and the session ended if the animal took longer than 30 minutes to meet the requirement. The breakpoint is defined as the total number of responses or the final ratio completed. Following PR, cocaine-seeking behavior was extinguished by replacing cocaine with 0.9% saline and removing the light and tone cues, previously paired with cocaine delivery. Daily 2 h extinction sessions occurred until animals performed <25% of their self-administration responding (average of last 3 days). Twenty-four hours after meeting this extinction criterion, animals underwent a cue-induced reinstatement session. The light and tone cues were presented non-contingently for 20 seconds, every 2 minutes during the first 10 minutes of the session. For the remainder of the session, the cues were presented contingent with operant responding, as they were during the cocaine self-administration phase. During the reinstatement session however, animals received saline infusions following responses on the active wheel. Catheter patency was tested once a week and animals were removed from the study if found to have lost patency.
2.6. Tissue Extraction.
In order to examine the effect of isolation stress on the cocaine response in the brain, a separate group was exposed to the adolescent isolation, along with group-housed controls, and at 8 weeks of age underwent operant food training, jugular catheterization, and 10 days of cocaine self-administration as described above. The cocaine self-administration data from these animals was included in the analysis of the behavioral data. Twenty-four hours after their last self-administration session, mice were either given an acute injection of cocaine (15 mg/kg) or taken from their home cage. 30 minutes following the cocaine injection, the mice received 100mg/kg pentobarbital (i.p.) before perfusion with 60 mL ice-cold PBS followed by 60 mL 4% PFA dissolved in ice-cold PBS. Brains were extracted and put in 4% PFA for 24 h before storage in 30% sucrose dissolved in PBS with 1% sodium azide. Coronal sections (40 μm) were taken using a cryostat and used for immunohistochemical staining. A separate cohort of animals that were either group housed or socially isolated at weaning were injected with saline in adulthood and underwent tissue extraction 30 minutes following the injection. Immunohistochemistry, cell counting and analysis occurred as described above.
2.7. Immunohistochemistry.
We rinsed sections in 0.5M TBS 3 times for 5 minutes each then quenched with 0.30% hydrogen peroxide for 15 minutes. Next, tissue was blocked in 10% DS-TBS for 1 h before incubation in c-Fos primary antibody (1:1000, Santa Cruz H-125) for 2 nights. On day 3, sections were rinsed again in .1%TBS-TX 3 times before a 2 h incubation in biotin donkey anti-rabbit (1:200, Jackson ImmunoResearch). Following another rinse in TBS-TX, the sections were incubated in S-HRP (Vectastain; 1:600) for 1 h then rinsed in TBS-TX 3 times. Finally, the tissue was incubated in DAB-Nickel for 5–10 minutes before the final rise in TBS until slide mounting.
2.8. Cell Counting.
c-Fos-immunolabeling was bilaterally quantified from at least 3–4 sections per mouse and averaged to determine the profile of each brain region. The experimenter quantifying was blind to group assignments. Anatomical regions were determined by the stereotaxic atlas of Franklin and Paxinos [23]. c-Fos images were quantified using automated ImageJ software.
2.9. Statistical Analysis.
All self-administration experiments were analyzed with two-way ANOVAs and post hoc comparisons as noted. The number of c-Fos immuno-labeled cells in the brain regions of interest was determined for each animal. Group differences in c-Fos immunoreactivity were analyzed using two-way ANOVAs with condition (isolation or group housing) and cocaine experience as the independent variables and number of cells as the dependent variable. Sidak’s post hoc comparisons were made when main effects or interactions were detected (p<0.05).
3. Results
3.1. Adolescent-onset social isolation does not alter the acquisition of an operant response, responding for sucrose, or the motivation for sucrose on a progressive ratio schedule.
Both male and female control and adolescent-onset social isolation mice were able to acquire an operant response for sucrose, exhibiting an increase in pellets earned and responses over the course of the 10 sessions (Fig. 1; males: effect of session, pellets= F(9,396)=2.035, p=0.034; responses= F(9,396)=6.0, p<0.0001; females: effect of session, pellets= F(9,297)=3.02, p=0.0018; responses= F(9,297)=5.2, p<0.0001). However, there was no influence of adolescent social isolation on sucrose intake or responding during the fixed ratio self-administration phase (males: effect of stress, pellets= F(1,44)=0.23, p=0.63; responses= F(1,44)=0.42, p=0.52; females: effect of stress, pellets= F(1,33)=1.02, p=0.32; responses= F(1,33)=0.97, p=.33). Furthermore, adolescent social isolation did not alter the breakpoint for sucrose on a progressive ratio schedule in either males [t(17)=0.22, p=.83] or females [t(16)=0.82, p=.43; Figure 1g,h]. Of note, there was a nonsignificant trend towards of an overall decrease in responding on PR in females (main effect of sex, F(1,33)=3.72, p=.063).
Figure 1. Adolescent isolation stress does not affect operant learning in food self-administration.
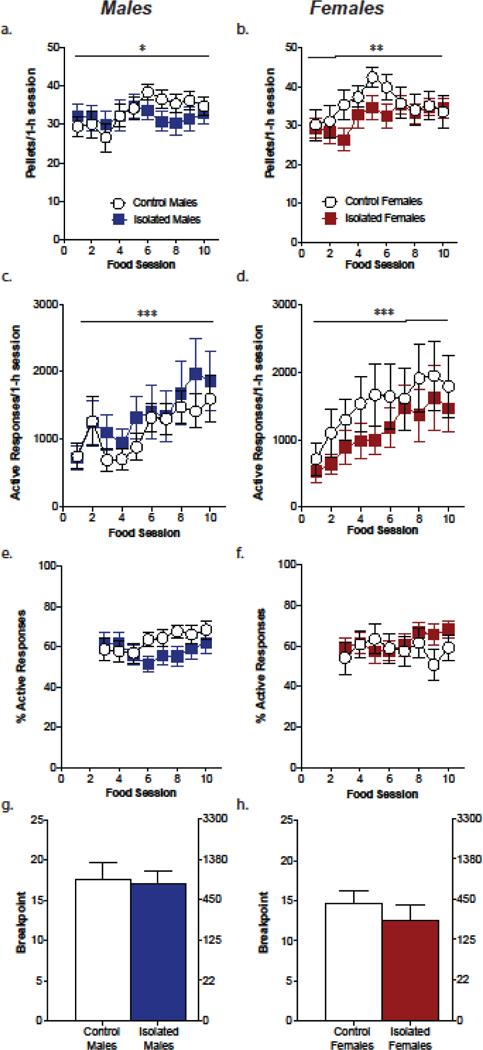
Male and female control and stress exposed mice showed consistent levels of pellets earned (a,b), a gradual increase in active responses (c,d), equal levels of percent active responding over 10 days of food training (e,f), and no differences in the responding for food on a progressive ratio schedule (g,h). *p<.05, **p<.01, **p<.001 main effect of session, n=14–20/group
3.2. Adolescent-onset social isolation in male mice increases responding for cocaine on an FR1 schedule of reinforcement.
Similar to what was seen for the sucrose self-administration phase, adolescent-onset social isolation did not alter cocaine intake, or number of infusions, in male or female mice during the FR1 cocaine self-administration phase (Fig. 2a,b; males: effect of stress, F(9,288)=1.0, p=0.33; females: effect of stress, F(9,189)=0.99, p=0.33). As a result of adolescent-onset social isolation, male mice exhibited an increase in responding on the active wheel for cocaine on an FR1 schedule, but female mice did not (Fig. 2c,d; males: effect of stress, F(9,297)=4.21, p=0.048; females: effect of stress, F(9,189)=0.63, p=0.44).
Figure 2. Adolescent isolation stress increases responding for cocaine on an FR1 schedule in males but not females.
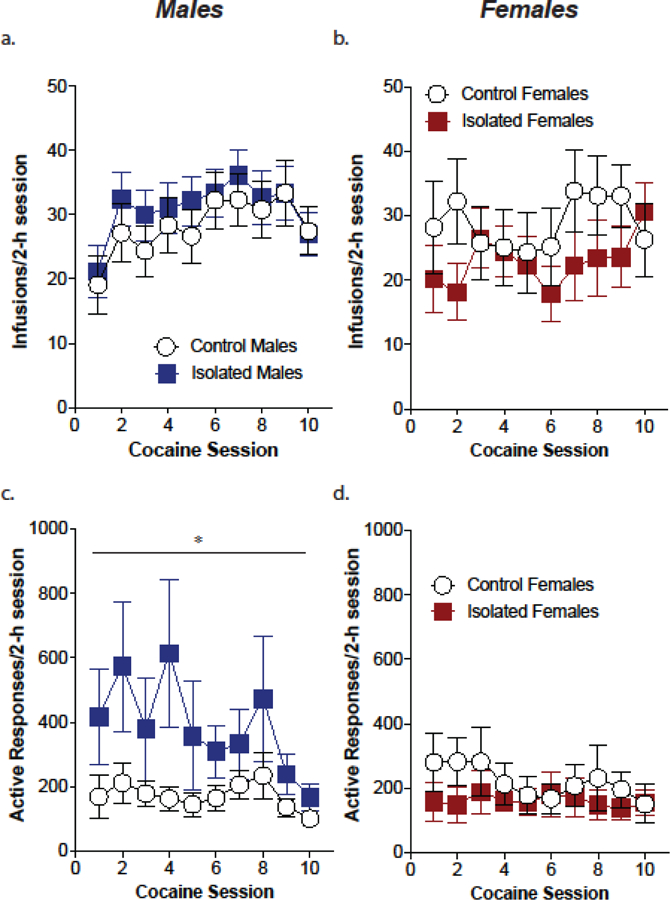
Male and female control and stress exposed mice earned equal numbers of infusions over the course of 10 days of cocaine self-administration (a,b; n=16–19/group). However, male mice exposed to adolescent-onset social isolation exhibit higher rates of responding for cocaine compared to male controls (c). Female mice isolated during adolescence do not exhibit any differences in responding compared to controls (d; n=11–13/group). **p<.01 main effect of session; *p<.05 main effect of stress
3.3. Adolescent-onset social isolation increases motivation for cocaine and potentiates cue-induced reinstatement of drug seeking in both male and female mice.
However, isolation stress led to an increase in responding on a progressive ratio schedule for cocaine in both male and female mice (Fig. 3a; effect of stress, F(1,50)=11.62, p=0.0013; effect of sex, F(1,50)=0.29, p=0.59). Isolation stress did not alter extinction learning, as male and female stressed and control mice exhibited similar rates of extinction (Fig. 3b; effect of stress, F(1,37)=0.086, p=0.77; effect of sex, F(1,37)=0.054, p=0.82) and similar levels of responding on the final day of extinction (Fig. 3c; effect of stress, F(1,37)=2.47, p=0.13; effect of sex, F(1,37)=2.5, p=0.12). Additionally there were no effects of adolescent isolation on responding on day 1 of extinction (F(1,37)=1.2, p=0.28; effect of sex, F(1,37)=0.35, p=0.56) During the cue-induced reinstatement test, both stress and control groups showed significant reinstatement (main effect of test, extinction vs. R1, when collapsed across sex, F(1,37)=13.7, p=.0007), however, the exposure of isolation stress during adolescence caused a potentiated reinstatement of drug seeking in both male and female mice (Fig. 3d; effect of stress, F(1,37)=5.46, p=0.025; effect of sex, F(1,37)=1.52, p=0.23).
Figure 3. Isolation stress during adolescence increases motivation for cocaine and cue-induced cocaine seeking in adulthood.

Male and female adolescent stress exposed mice respond more on a progressive ratio schedule of reinforcement, exhibiting a higher breakpoint (a; n=10–18/group). No differences were seen between the groups in the rate of extinction (b) or the responding on the final day of extinction (c). Twenty-four hours after the final extinction session, during the cue-induced reinstatement session, male and female mice exposed to adolescent social isolation exhibited higher rates of responding compared to controls (d; n=8–12/group). *p<.05 main effect of stress
3.4. Cocaine-induced c-Fos expression is increased following adolescent-onset social isolation stress and chronic cocaine self-administration
To better understand the neurobiological mechanisms underlying the behavioral differences observed following adolescent-onset social isolation, we looked at cocaine-evoked c-Fos expression in several brain regions following chronic cocaine exposure and adolescent stress. Due to the similar trends seen in the motivation for cocaine and the reinstatement of cocaine seeking along with the similar levels of Fos activation between isolated males and females in the cocaine group (Table 1), male and female mice were grouped together for these studies. Adolescent-onset isolation coupled with chronic cocaine exposure causes a significant increase in cocaine-induced c-Fos expression in the nucleus accumbens core and shell, dorsal bed nucleus of the stria terminalis (BNST), lateral septum, basolateral amygdala and a trend towards an increase in the ventral pallidum (Fig. 4). Additionally, we saw a main effect of acute cocaine injection without any interaction with adolescent stress exposure in the prelimbic cortex, infralimbic cortex, dorsomedial and dorsolateral striatum, and ventral BNST (Table 2).
Table 1. Statistical analysis of sex differences in c-Fos profiles following cocaine.
This table summarizes the number of c-Fos positive cells in male and female mice that underwent adolescent social stress. As no statistically significant differences were found between males and females, these data were grouped together for future analyses.
| Brain Region | Fos+ Cells Stressed Males |
Fos+ Cells Stressed Females |
Effect of Sex t(6), p |
|---|---|---|---|
| NAc Core | 930 | 1038 | t=0.26, p=.81 |
| NAc Shell | 687 | 577 | t=0.43, p=.68 |
|
Ventral Pallidum |
243 | 238 | t=0.03, p=.98 |
| dBNST | 464 | 512 | t=0.26, p=.80 |
|
Lateral Septum |
381 | 353 | t=0.33, p=.76 |
| BLA | 525 | 640 | t=1.1, p=.32 |
Figure 4. Chronic cocaine exposure after adolescent stress causes an increased c-Fos response in the nucleus accumbens core and shell, basolateral amygdala, and lateral septum.
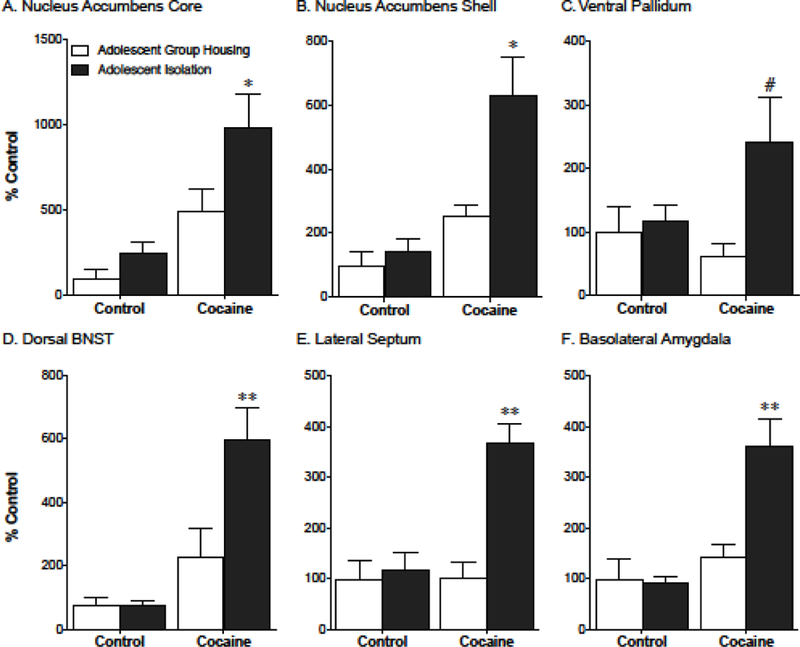
Stress exposed mice show greater c-Fos activation (relative to the naïve group housed control group which is at 100%) in the nucleus accumbens core (a) and shell (b), ventral pallidum (c), dorsal BNST (d), lateral septum (e), and basolateral amygdala (f) after chronic cocaine compared to group housed controls. #p=.05, *p<.05, **p<.01 post-hoc control cocaine vs. isolation cocaine; n=5–9/group
Table 2. Statistical analysis of c-Fos profiles following cocaine.
This table summarizes the statistical analyses performed to examine the effect of adolescent isolation stress on the Fos expression following acute cocaine exposure in mice with a history of cocaine self-administration.
| Brain Region | Fos+ Cells Group Control |
Fos+ Cells Isolation Stress |
Effect of Stress F(1,22), p |
Effect of Cocaine F(1,22), p |
Interaction F(1,22), p |
Sidak’s Post-hoc Cocaine Control vs. Iso |
|---|---|---|---|---|---|---|
| PrL | 216.2 | 326.6 | F=2.68, p=.11 | F=6.59, *p=.018 | F=.17, p=.68 | – |
| IL | 168.7 | 263 | F=3.71, p=.068 | F=9.21, **p=.006 | F=0.89, p=.36 | – |
| NAc Core | 490.8 | 983.8 | F=4.99, *p=.04 | F=15.9, **p=.0006 | F=1.53, p=.23 | *p=.047 |
| NAc Shell | 253.5 | 632.3 | F=5.80, *p=.02 | F=13.4, *p=.002 | F=3.65, p=.069 | *p=.016 |
| dmStr | 478 | 1031 | F=1.12, p=.30 | F=5.69, *p=.028 | F=0.97, p=.34 | – |
| dlStr | 532.6 | 888 | F=0.52, p=.48 | F=6.97, *p=.016 | F=0.63, p=.44 | – |
|
Ventral Pallidum |
62.4 | 240.9 | F=3.46, p=.06 | F=0.63, p=.43 | F=2.27, *p=.149 | #p=.05 |
| dBNST | 228.1 | 595.9 | F=6.61, *p=.02 | F=21.7, ***p=.0006 | F=6.50, *p=.026 | **p=.006 |
| vBNST | 250.8 | 397.1 | F=1.11, p=.30 | F=9.28, **p=.006 | F=1.01, p=.33 | – |
|
Lateral Septum |
102 | 367.2 | F=14.0, **p=.001 | F=11.0, **p=.004 | F=10.6, *p=.004 | ***p=.0004 |
| BLA | 219.4 | 594.4 | F=33.0, *p<.0001 | F=12.7, **p=.004 | F=12.2, *p=.004 | ***p=.0003 |
| CeA | 195.9 | 269.4 | F=0.89, p=.35 | F=3.43, p=.07 | F=0.07, p=.79 | – |
3.5. Saline-induced c-Fos expression is not altered by adolescent-onset social isolation stress
As the stress of the injection itself may have contributed to our findings above, a separate cohort of male and female mice that were either group housed or socially isolated at weaning were injected with saline and c-Fos expression was examined. We focused on the brain regions where we saw differences following cocaine and there was no effect of adolescent social isolation on saline-induced c-Fos expression within the NAc Core, NAc shell, ventral pallidum, dBNST, Lateral Septum, or BLA (Table 3).
Table 3. Statistical analysis of c-Fos profiles following saline.
This table summarizes the statistical analyses performed to examine whether adolescent isolation stress influences Fos expression following a single saline injection.
| Brain Region | Fos+ Cells Group Control |
Fos+ Cells Isolation Stress |
Effect of Stress t(10), p |
|---|---|---|---|
| NAc Core | 121 | 131 | t=0.56, p=0.59 |
| NAc Shell | 64.6 | 68.9 | t=0.33, p=0.75 |
|
Ventral Pallidum |
9.10 | 7.62 | t=0.69, p=0.51 |
| dBNST | 55.1 | 47.9 | t=0.90, p=0.39 |
|
Lateral Septum |
110 | 132 | t=1.37, p=0.20 |
| BLA | 56.8 | 46.1 | t=1.28, p=0.23 |
4. Discussion
Here we present evidence that social isolation beginning in adolescence leads to increased seeking and motivation for cocaine in adulthood. Additionally, we present a novel observation of an increase in cocaine-induced c-Fos expression within regions implicated in reward processing as well as regions that mediate stress responsivity. We suggest that this increased behavioral responsivity to cocaine may be mediated by heightened glutamate activity within reward and stress related circuitry.
4.1. Adolescent social isolation promotes vulnerability to drug-associated behaviors in adulthood in both male and female mice.
Early life stress is a risk factor for the development of addiction in humans [5–7]. Social stressors are highly prevalent and among the most powerful stressful stimuli experienced by humans [24]. Modeling this increased risk in animals can provide us with a means to identify neural mechanisms that underlie increased vulnerability in humans. Rodents who are socially isolated at weaning exhibit enhanced alcohol consumption [25, 26], enhanced cocaine intake [27], enhanced locomotor response to amphetamine [28], and enhanced cocaine-evoked dopamine release [29]. In the present study, social isolation at weaning led to an increase in cocaine responding during self-administration. Notably, the number of cocaine infusions did not increase, supporting the idea that adolescent stress exposure is modulating motivation for the drug (Fig. 2). Further, this increased motivation is seen on a progressive ratio schedule as well as increased drug seeking during cue-induced reinstatement (Fig. 3). This effect of adolescent-onset social isolation was not observed in food responding, which suggests a unique effect on drug-related behavior.
While the pathologically relevant time period for isolation is adolescence, the socially isolated animals in the present study were not reintroduced to group housing conditions before beginning the operant training paradigm. Instead, animals in the control group were isolated at this time due to issues with group housing animals with indwelling catheters. While it is possible that the duration of the social isolation influenced the current findings, the control animals had been socially isolated for 3 weeks at start of the cocaine self-administration phase, the enhanced drug-associated behaviors were unique to animals isolated during adolescence. Moreover, the effects of adolescent social isolation were specific to cocaine as no differences in food intake or motivation for food were seen.
These effects are consistent with what has been seen following other forms of adolescent stress. In particular, numerous studies have demonstrated that adolescent social defeat increases drug-associated behaviors in male rodents. Specifically, increased conditioned place preference to cocaine and MDMA is seen following adolescent social defeat stress [30, 31]. Similarly, both adolescent social defeat and social isolation increase cocaine self-administration in male rats [12, 32]. However, post-weaning social isolation served to protect rats from this increase in cocaine taking seen following social defeat in pair-housed animals [12]. In contrast to the increase in addictive phenotypes seen following adolescent social stress, stress earlier in life may be protective. Maternal separation stress leads to decreased cocaine self-administration, particularly in males [33–35]. This suggests that perhaps adolescence is a critical period for stress exposure to predispose individuals to develop addiction.
4.2. The influence of sex on the effects of adolescent social isolation on drug-associated behaviors in adulthood.
The majority of models examining social stress have utilized only male animals due to male’s innate territorial aggression toward other males, a behavior that is absent in females under normal conditions [36, 37]. The current study is the first to demonstrate that adolescent social stress can increase drug-associated behaviors in adult females. As human females are more likely to develop stress-related psychiatric disorders [38], developing a model to examine the influence of adolescent social stress on drug-associated behaviors in females is critical.
Although we did not see sex differences in the ability of adolescent social isolation to increase the motivation for cocaine or cue-induced cocaine seeking during reinstatement, we did find that male mice exposed to social isolation during adolescence exhibit higher rates of responding during cocaine self-administration (Fig. 2). This increase in responding was not accompanied by an increase in cocaine intake, indicating that these mice exhibited more responses during the time out period. This may suggest that male mice have increased habitual responding or an increase in impulsivity and future studies could tease apart what is responsible for this pattern of responding. Of note, the mice were trained on an operant response for sucrose prior to the cocaine self-administration phase so it is impossible to rule out the influence of sucrose extinction on the responding during the beginning of cocaine self-administration phase. However, the specific effect of adolescent social isolation on motivation for cocaine vs. sucrose supports our hypothesis that the effects on seeking are specific.
4.3. Adolescent social isolation increases the neuronal response to cocaine in adulthood.
As the brain is still developing throughout childhood and adolescence, stress during this period of life may alter the development of reward circuitry. Human functional imaging studies indicate that childhood trauma leads to dysfunction in reward processing in adulthood [39, 40]. Furthermore, PET binding studies suggest that adults with a history of childhood trauma exhibit increased dopamine response to stimulants [41]. However, it is not clear from these studies whether these alterations in reward processing are a consequence of stress exposure or represent some common vulnerability to stress and addiction that precedes trauma. The current study determined that adolescent social isolation increases cocaine-induced activation of several brain regions previously implicated in stress and reward-related behavior in mice [Fig. 4; 42, 43–46]. In group-housed, male and female mice with a history of cocaine self-administration, acute cocaine exposure led to an increase in c-Fos expression in the nucleus accumbens core and shell and dorsal bed nucleus of the stria terminalis. However, in socially isolated mice with a history of cocaine self-administration, acute cocaine increased c-Fos to a greater extent in these three regions as well as engaging additional brain regions: ventral pallidum (VP), lateral septum (LS), and the basolateral amygdala (BLA).
The ventral pallidum receives input from the nucleus accumbens and serves to integrate information from multiple cell populations. Chronic cocaine exposure leads to potentiated output in accumbal MSNs [47]. Therefore, the increase in c-Fos activation within the accumbens and VP could represent a further potentiation of this response by adolescent stress exposure. Future work could examine the ability of stress to influence plasticity within this circuit. Cocaine also led to activation of stress-related brain circuits in animals that received adolescent social stress. The BLA projections to both the nucleus accumbens and the orbitofrontal cortex play key roles in cue-induced cocaine seeking [48–50]. Furthermore, early life stress leads to increased spine density and synaptic excitability within the BLA [51]. Coupled with our findings, this supports that increased activation within the BLA could mediate the increase in cue-induced cocaine seeking seen following adolescent social isolation. Recently, the LS has also been implicated in cocaine seeking behavior. Inputs from the LS to the ventral tegmental area are activated during cue-induced reinstatement [52] and projections from the dorsal hippocampus to the LS are activated during context-induced reinstatement [53]. Further examination of the influence of adolescent stress on inputs both to and from the LS could provide insight into the behavioral differences seen in reinstatement behavior in the current study.
Our results are consistent with previous work demonstrating that isolation rearing increases the ability of an acute cocaine injection to induce c-Fos expression within the nucleus accumbens core and shell [54]. The current study expands this to demonstrate cocaine-induced changes in c-Fos expression in animals with a history of cocaine self-administration. We see more widespread increases in regions implicated in stress along with addiction. Previous studies have demonstrated increases in c-Fos expression following acute cocaine in regions where we did not see increases in the group-housed animals. This includes studies done in drug naïve mice demonstrating increased c-Fos expression the lateral septum [55] and studies examining c-Fos expression immediately following self-administration that have seen increased c-Fos in the BLA and VP [56]. There may be something unique about the combination of self-administration and experimenter administered cocaine or these differences may be due to across species differences as all these studies were done in rats. Additionally, a large body of work has examined c-Fos expression in cocaine-experienced mice following periods of abstinence or extinction. These paradigms lead to robust increases in c-Fos expression throughout brain regions implicated in reward, including the nucleus accumbens and regions implicated in stress reactivity, including the BLA [57–59]. These data, coupled with the reinstatement data in the current study, suggest that we might see a different pattern of c-Fos expression if we examined animals after reinstatement.
4.3. Conclusion
Taken together, the current results indicate that adolescent-onset social isolation leads to an increase in responding during fixed-ratio cocaine self-administration, motivation for cocaine, and cocaine seeking during cue-induced reinstatement. These increases are specific to drug reward, as social isolation did not impact sucrose self-administration or motivation for sucrose. Accompanying these increases in behavioral response to cocaine is an increase in the neuronal response to cocaine as evidenced by increased c-Fos expression. These results highlight the role of adolescent stress in addiction and suggest that individual vulnerabilities to addiction may be mediated by alterations in AMPA transmission in the nucleus accumbens.
Acknowledgements
This work was supported by National Institute on Drug Abuse (NIDA) Grant R00 DA033372 (L.A.B.) and a Brain & Behavior Research Foundation NARSAD award (L.A.B.).
We thank Dr. Debra Bangasser for providing the feedback on the manuscript.
Role of the funding source
None of the funding sources had any role in the study design, data collection, data analysis, writing of the manuscript or the decision to submit the manuscript for publication.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of Interest
The authors declare no conflict of interest.
References
- [1].Topper LR, Castellanos-Ryan N, Mackie C, Conrod PJ, Adolescent bullying victimisation and alcohol-related problem behaviour mediated by coping drinking motives over a 12 month period, Addict Behav 36(1–2) (2011) 6–13. [DOI] [PubMed] [Google Scholar]
- [2].Tharp-Taylor S, Haviland A, D’Amico EJ, Victimization from mental and physical bullying and substance use in early adolescence, Addict Behav 34(6–7) (2009) 561–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Somaini L, Donnini C, Manfredini M, Raggi MA, Saracino MA, Gerra ML, Amore M, Leonardi C, Serpelloni G, Gerra G, Adverse childhood experiences (ACEs), genetic polymorphisms and neurochemical correlates in experimentation with psychotropic drugs among adolescents, Neurosci Biobehav Rev 35(8) (2011) 1771–8. [DOI] [PubMed] [Google Scholar]
- [4].Nelson DE, Higginson GK, Grant-Worley JA, Physical abuse among high school students. Prevalence and correlation with other health behaviors, Arch Pediatr Adolesc Med 149(11) (1995) 1254–8. [DOI] [PubMed] [Google Scholar]
- [5].Dube SR, Miller JW, Brown DW, Giles WH, Felitti VJ, Dong M, Anda RF, Adverse childhood experiences and the association with ever using alcohol and initiating alcohol use during adolescence, J Adolesc Health 38(4) (2006) 444 e1–10. [DOI] [PubMed] [Google Scholar]
- [6].Dube SR, Felitti VJ, Dong M, Chapman DP, Giles WH, Anda RF, Childhood abuse, neglect, and household dysfunction and the risk of illicit drug use: the adverse childhood experiences study, Pediatrics 111(3) (2003) 564–72. [DOI] [PubMed] [Google Scholar]
- [7].Brents LK, Tripathi SP, Young J, James GA, Kilts CD, The role of childhood maltreatment in the altered trait and global expression of personality in cocaine addiction, J Psychiatr Res 64 (2015) 23–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Kaltiala-Heino R, Rimpela M, Rantanen P, Rimpela A, Bullying at school--an indicator of adolescents at risk for mental disorders, J Adolesc 23(6) (2000) 661–74. [DOI] [PubMed] [Google Scholar]
- [9].Hoffmann JP, Cerbone FG, Su SS, A growth curve analysis of stress and adolescent drug use, Substance use & misuse 35(5) (2000) 687–716. [DOI] [PubMed] [Google Scholar]
- [10].Dube SR, Anda RF, Felitti VJ, Edwards VJ, Croft JB, Adverse childhood experiences and personal alcohol abuse as an adult, Addict Behav 27(5) (2002) 713–25. [DOI] [PubMed] [Google Scholar]
- [11].Anda RF, Whitfield CL, Felitti VJ, Chapman D, Edwards VJ, Dube SR, Williamson DF, Adverse childhood experiences, alcoholic parents, and later risk of alcoholism and depression, Psychiatric services (Washington, D.C 53(8) (2002) 1001–9. [DOI] [PubMed] [Google Scholar]
- [12].Burke AR, Miczek KA, Escalation of cocaine self-administration in adulthood after social defeat of adolescent rats: role of social experience and adaptive coping behavior, Psychopharmacology (Berl) 232(16) (2015) 3067–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Peleg-Raibstein D, Feldon J, Differential effects of post-weaning juvenile stress on the behaviour of C57BL/6 mice in adolescence and adulthood, Psychopharmacology (Berl) 214(1) (2011) 339–51. [DOI] [PubMed] [Google Scholar]
- [14].Lepsch LB, Gonzalo LA, Magro FJ, Delucia R, Scavone C, Planeta CS, Exposure to chronic stress increases the locomotor response to cocaine and the basal levels of corticosterone in adolescent rats, Addict Biol 10(3) (2005) 251–6. [DOI] [PubMed] [Google Scholar]
- [15].Cruz FC, Marin MT, Leao RM, Planeta CS, Stress-induced cross-sensitization to amphetamine is related to changes in the dopaminergic system, J Neural Transm (Vienna) 119(4) (2012) 415–24. [DOI] [PubMed] [Google Scholar]
- [16].Burke AR, Forster GL, Novick AM, Roberts CL, Watt MJ, Effects of adolescent social defeat on adult amphetamine-induced locomotion and corticoaccumbal dopamine release in male rats, Neuropharmacology 67 (2013) 359–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Burke AR, Watt MJ, Forster GL, Adolescent social defeat increases adult amphetamine conditioned place preference and alters D2 dopamine receptor expression, Neuroscience 197 (2011) 269–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Rodriguez-Arias M, Montagud-Romero S, Rubio-Araiz A, Aguilar MA, Martin-Garcia E, Cabrera R, Maldonado R, Porcu F, Colado MI, Minarro J, Effects of repeated social defeat on adolescent mice on cocaine-induced CPP and self-administration in adulthood: integrity of the blood-brain barrier, Addict Biol (2015). [DOI] [PubMed]
- [19].Baarendse PJ, Limpens JH, Vanderschuren LJ, Disrupted social development enhances the motivation for cocaine in rats, Psychopharmacology (Berl) 231(8) (2014) 1695–704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Bozarth MA, Murray A, Wise RA, Influence of housing conditions on the acquisition of intravenous heroin and cocaine self-administration in rats, Pharmacol Biochem Behav 33(4) (1989) 903–7. [DOI] [PubMed] [Google Scholar]
- [21].Schenk S, Lacelle G, Gorman K, Amit Z, Cocaine self-administration in rats influenced by environmental conditions: implications for the etiology of drug abuse, Neurosci Lett 81(1–2) (1987) 227–31. [DOI] [PubMed] [Google Scholar]
- [22].Thiel KJ, Okun AC, Neisewander JL, Social reward-conditioned place preference: a model revealing an interaction between cocaine and social context rewards in rats, Drug Alcohol Depend 96(3) (2008) 202–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Franklin KBJ, Paxinos G, Paxinos and Franklin’s The mouse brain in stereotaxic coordinates, Fourth edition. ed., Academic Press, an imprint of Elsevier, Amsterdam, 2013. [Google Scholar]
- [24].Backstrom T, Winberg S, Central corticotropin releasing factor and social stress, Front Neurosci 7 (2013) 117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].McCool BA, Chappell AM, Early social isolation in male Long-Evans rats alters both appetitive and consummatory behaviors expressed during operant ethanol self-administration, Alcoholism, clinical and experimental research 33(2) (2009) 273–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Chappell AM, Carter E, McCool BA, Weiner JL, Adolescent rearing conditions influence the relationship between initial anxiety-like behavior and ethanol drinking in male Long Evans rats, Alcoholism, clinical and experimental research 37 Suppl 1 (2013) E394–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Ding Y, Kang L, Li B, Ma L, Enhanced cocaine self-administration in adult rats with adolescent isolation experience, Pharmacol Biochem Behav 82(4) (2005) 673–7. [DOI] [PubMed] [Google Scholar]
- [28].Herrmann AP, Benvenutti R, Pilz LK, Elisabetsky E, N-acetylcysteine prevents increased amphetamine sensitivity in social isolation-reared mice, Schizophrenia research 155(1–3) (2014) 109–11. [DOI] [PubMed] [Google Scholar]
- [29].Yorgason JT, Calipari ES, Ferris MJ, Karkhanis AN, Fordahl SC, Weiner JL, Jones SR, Social isolation rearing increases dopamine uptake and psychostimulant potency in the striatum, Neuropharmacology 101 (2016) 471–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Montagud-Romero S, Nunez C, Blanco-Gandia MC, Martinez-Laorden E, Aguilar MA, Navarro-Zaragoza J, Almela P, Milanes MV, Laorden ML, Minarro J, Rodriguez-Arias M, Repeated social defeat and the rewarding effects of cocaine in adult and adolescent mice: dopamine transcription factors, proBDNF signaling pathways, and the TrkB receptor in the mesolimbic system, Psychopharmacology (Berl) 234(13) (2017) 2063–2075. [DOI] [PubMed] [Google Scholar]
- [31].Garcia-Pardo MP, Blanco-Gandia MC, Valiente-Lluch M, Rodriguez-Arias M, Minarro J, Aguilar MA, Long-term effects of repeated social stress on the conditioned place preference induced by MDMA in mice, Prog Neuropsychopharmacol Biol Psychiatry 63 (2015) 98–109. [DOI] [PubMed] [Google Scholar]
- [32].Yajie D, Lin K, Baoming L, Lan M, Enhanced cocaine self-administration in adult rats with adolescent isolation experience, Pharmacol Biochem Behav 82(4) (2005) 673–7. [DOI] [PubMed] [Google Scholar]
- [33].Matthews K, Robbins TW, Everitt BJ, Caine SB, Repeated neonatal maternal separation alters intravenous cocaine self-administration in adult rats, Psychopharmacology (Berl) 141(2) (1999) 123–34. [DOI] [PubMed] [Google Scholar]
- [34].Martini M, Valverde O, A single episode of maternal deprivation impairs the motivation for cocaine in adolescent mice, Psychopharmacology (Berl) 219(1) (2012) 149–58. [DOI] [PubMed] [Google Scholar]
- [35].O’Connor RM, Moloney RD, Glennon J, Vlachou S, Cryan JF, Enhancing glutamatergic transmission during adolescence reverses early-life stress-induced deficits in the rewarding effects of cocaine in rats, Neuropharmacology 99 (2015) 168–76. [DOI] [PubMed] [Google Scholar]
- [36].Haller J, Fuchs E, Halasz J, Makara GB, Defeat is a major stressor in males while social instability is stressful mainly in females: towards the development of a social stress model in female rats, Brain Res Bull 50(1) (1999) 33–9. [DOI] [PubMed] [Google Scholar]
- [37].Takahashi A, Chung JR, Zhang S, Zhang H, Grossman Y, Aleyasin H, Flanigan ME, Pfau ML, Menard C, Dumitriu D, Hodes GE, McEwen BS, Nestler EJ, Han MH, Russo SJ, Establishment of a repeated social defeat stress model in female mice, Sci Rep 7(1) (2017) 12838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Bangasser DA, Valentino RJ, Sex differences in molecular and cellular substrates of stress, Cellular and molecular neurobiology 32(5) (2012) 709–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Mehta MA, Gore-Langton E, Golembo N, Colvert E, Williams SC, Sonuga-Barke E, Hyporesponsive reward anticipation in the basal ganglia following severe institutional deprivation early in life, J Cogn Neurosci 22(10) (2010) 2316–25. [DOI] [PubMed] [Google Scholar]
- [40].Dillon DG, Holmes AJ, Birk JL, Brooks N, Lyons-Ruth K, Pizzagalli DA, Childhood adversity is associated with left basal ganglia dysfunction during reward anticipation in adulthood, Biol Psychiatry 66(3) (2009) 206–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Oswald LM, Wand GS, Kuwabara H, Wong DF, Zhu S, Brasic JR, History of childhood adversity is positively associated with ventral striatal dopamine responses to amphetamine, Psychopharmacology (Berl) 231(12) (2014) 2417–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Kalivas PW, Addiction as a pathology in prefrontal cortical regulation of corticostriatal habit circuitry, Neurotox Res 14(2–3) (2008) 185–9. [DOI] [PubMed] [Google Scholar]
- [43].Koob GF, Le Moal M, Review. Neurobiological mechanisms for opponent motivational processes in addiction, Philos Trans R Soc Lond B Biol Sci 363(1507) (2008) 3113–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Shalev U, Grimm JW, Shaham Y, Neurobiology of relapse to heroin and cocaine seeking: a review, Pharmacological reviews 54(1) (2002) 1–42. [DOI] [PubMed] [Google Scholar]
- [45].Thiel KJ, Pentkowski NS, Peartree NA, Painter MR, Neisewander JL, Environmental living conditions introduced during forced abstinence alter cocaine-seeking behavior and Fos protein expression, Neuroscience 171(4) (2010) 1187–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Vanderschuren LJ, Di Ciano P, Everitt BJ, Involvement of the dorsal striatum in cue-controlled cocaine seeking, J Neurosci 25(38) (2005) 8665–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Creed M, Ntamati NR, Chandra R, Lobo MK, Luscher C, Convergence of Reinforcing and Anhedonic Cocaine Effects in the Ventral Pallidum, Neuron 92(1) (2016) 214–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Lasseter HC, Wells AM, Xie X, Fuchs RA, Interaction of the basolateral amygdala and orbitofrontal cortex is critical for drug context-induced reinstatement of cocaine-seeking behavior in rats, Neuropsychopharmacology 36(3) (2011) 711–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Arguello AA, Richardson BD, Hall JL, Wang R, Hodges MA, Mitchell MP, Stuber GD, Rossi DJ, Fuchs RA, Role of a Lateral Orbital Frontal Cortex-Basolateral Amygdala Circuit in Cue-Induced Cocaine-Seeking Behavior, Neuropsychopharmacology 42(3) (2017) 727–735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Stefanik MT, Kalivas PW, Optogenetic dissection of basolateral amygdala projections during cue-induced reinstatement of cocaine seeking, Frontiers in behavioral neuroscience 7 (2013) 213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Guadagno A, Wong TP, Walker CD, Morphological and functional changes in the preweaning basolateral amygdala induced by early chronic stress associate with anxiety and fear behavior in adult male, but not female rats, Prog Neuropsychopharmacol Biol Psychiatry 81 (2018) 25–37. [DOI] [PubMed] [Google Scholar]
- [52].Mahler SV, Aston-Jones GS, Fos activation of selective afferents to ventral tegmental area during cue-induced reinstatement of cocaine seeking in rats, J Neurosci 32(38) (2012) 13309–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].McGlinchey EM, Aston-Jones G, Dorsal Hippocampus Drives Context-Induced Cocaine Seeking via Inputs to Lateral Septum, Neuropsychopharmacology (2017). [DOI] [PMC free article] [PubMed]
- [54].Howes SR, Dalley JW, Morrison CH, Robbins TW, Everitt BJ, Leftward shift in the acquisition of cocaine self-administration in isolation-reared rats: relationship to extracellular levels of dopamine, serotonin and glutamate in the nucleus accumbens and amygdala-striatal FOS expression, Psychopharmacology (Berl) 151(1) (2000) 55–63. [DOI] [PubMed] [Google Scholar]
- [55].Briand LA, Vassoler FM, Pierce RC, Valentino RJ, Blendy JA, Ventral tegmental afferents in stress-induced reinstatement: the role of cAMP response element-binding protein, J Neurosci 30(48) (2010) 16149–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Zahm DS, Becker ML, Freiman AJ, Strauch S, Degarmo B, Geisler S, Meredith GE, Marinelli M, Fos after single and repeated self-administration of cocaine and saline in the rat: emphasis on the Basal forebrain and recalibration of expression, Neuropsychopharmacology 35(2) (2010) 445–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Hearing MC, Miller SW, See RE, McGinty JF, Relapse to cocaine seeking increases activity-regulated gene expression differentially in the prefrontal cortex of abstinent rats, Psychopharmacology (Berl) 198(1) (2008) 77–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Zavala AR, Biswas S, Harlan RE, Neisewander JL, Fos and glutamate AMPA receptor subunit coexpression associated with cue-elicited cocaine-seeking behavior in abstinent rats, Neuroscience 145(2) (2007) 438–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Zavala AR, Browning JR, Dickey ED, Biswas S, Neisewander JL, Region-specific involvement of AMPA/Kainate receptors in Fos protein expression induced by cocaine-conditioned cues, Eur Neuropsychopharmacol 18(8) (2008) 600–11. [DOI] [PMC free article] [PubMed] [Google Scholar]