

Abstract Abstract
A new species of the genus Gracixalus, Gracixalusyunnanensissp. n., is described based on a series of specimens collected from southwestern and southern Yunnan, China. This species is distinguished from all other known congeners by a combination of the following characters: relatively small body size in adult males (SVL 26.0–34.2 mm); dorsal surface yellow brown or red brown; distinctive conical asperities on dorsum; males with an external subgular vocal sac and linea masculina; throat, chest, and belly nearly immaculate; venter surface orangish with yellow spots, semi-transparent; snout rounded; supratympanic fold distinct; iris bronze; lack of white patch on temporal region; tibiotarsal projection absent; sides of body nearly smooth with no black blotch; finger webbing rudimentary; and toe webbing formula I1.5–2II1.5–2.7III.5–3IV2.5–1.5V. Genetically, the new species diverges from its congeners by 2.2%–14.1% (uncorrected p-distance) and is closest to G.guangdongensis. However, the new species can morphologically be separated from G.guangdongensis by distinctive conical tubercles on dorsum (versus absent), lateral surface nearly smooth with no black blotches on ventrolateral region (versus lateral surface rough, scattered with tubercles and black blotches on ventrolateral region), snout rounded (versus triangularly pointed), iris bronze (versus iris brown), and ventral surface orangish (versus throat and chest creamy white and belly light brown).
Keywords: Gracixalusyunnanensis sp. n., taxonomy, Rhacophoridae , southwestern China
Introduction
The genus Gracixalus Delorme, Dubois, Grosjean & Ohler, 2005 is known from southern and southwestern China, Vietnam, Laos, Thailand, and Myanmar and contains 16 species including G.ananjevae (Matsui & Orlov, 2004), G.carinensis (Boulenger, 1893), G.gracilipes (Bourret, 1937), G.guangdongensis Wang, Zeng, Liu & Wang, 2018, G.jinggangensis Zeng, Zhao, Chen, Chen, Zhang & Wang, 2017, G.jinxiuensis (Hu, in Hu et al. 1978), G.lumarius Rowley, Le, Dau, Hoang & Cao, 2014, G.medogensis (Ye & Hu, 1984), G.nonggangensis Mo, Zhang, Luo, Zhou & Chen, 2013, G.quangi, Rowley, Dau, Nguyen, Cao & Nguyen, 2011, G.quyeti (Nguyen, Hendrix, Bohme, Vu & Ziegler, 2008), G.sapaensis Matsui, Ohler, Eto & Nguyen, 2017, G.seesom Matsui, Khonsue, Panha & Eto, 2015, G.supercornutus (Orlov, Ho & Nguyen, 2004), G.tianlinensis Chen, Bei, Liao, Zhou & Mo, 2018, and G.waza Nguyen, Le, Pham, Nguyen, Bonkowski & Ziegler, 2013 (Frost 2018). Of the 16 members of Gracixalus, ten were discovered in last decade (Nguyen et al. 2008, Rowley et al. 2011, Mo et al. 2013, Nguyen et al. 2013, Rowley et al. 2014, Matsui et al. 2015, Matsui et al. 2017, Zeng et al. 2017, Chen et al. 2018, Wang et al. 2018), indicating that species diversity of Gracixalus was very poorly understood in the past. Moreover, recent phylogenetic analyses (Matsui et al. 2017, Chen et al. 2018) showed that there are still several unnamed distinct lineages in the group of G.jinxiuensis, indicating that species richness of Gracixalus remains underestimated.
During recent fieldworks in Yunnan, China, we collected some specimens of a small-sized tree frog, which morphologically can be assigned to the genus Gracixalus by the presence of intercalary cartilage between terminal and penultimate phalanges of digits, tips of digits enlarged to discs bearing circummarginal grooves, vomerine teeth absent, inner (first and second) and outer (third and fourth) fingers non-opposable, and an inversed Y-shaped dark brown marking on dorsum (Fei 1999, Rowley et al. 2011, Chen et al. 2018), but morphologically and genetically can be distinguished from all recognized species of genus Gracixalus. Thus, we describe these specimens as a new species of Gracixalus.
Materials and methods
Sampling
Specimens were collected during fieldworks in Menghai County, Lancang County, and Lvchun County of Yunnan, China in 2014 to 2018 (Fig. 1). They were fixed and then stored in 80% ethanol after taking photos. Liver tissues were preserved in 99% ethanol. Specimens were deposited at the Kunming Institute of Zoology (KIZ), Chinese Academy of Sciences and Guangxi Normal University (GXNU).
Figure 1.
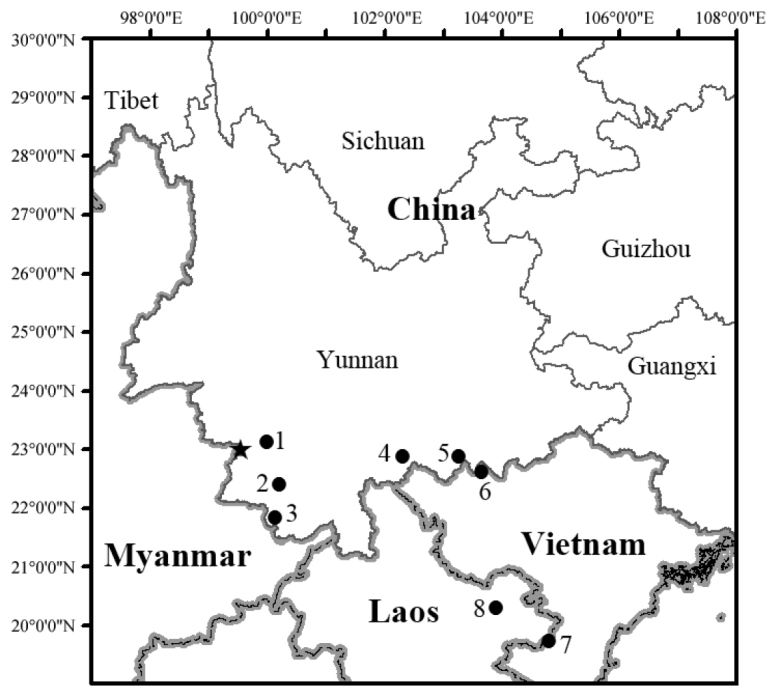
Map showing collection sites of Gracixalusyunnanensis sp. n. Star indicates the type locality (Xuelin) and circles indicate Fudong Township (1), Fazhanhe Township (2), Bada Township (3), Mt. Huanglian (4), Jinping (5), Lao Cai (6), Nghe An (7), and Houapan (8), respectively. Sequences of samples from sites 5–8 came from previous studies.
Morphology
Morphometric data were taken using digital calipers to the nearest 0.1 mm. Morphological terminologies follow Matsui et al. (2017) and Wang et al. (2018). Measurements include:
SVL snout-vent length (from tip of snout to vent);
HL head length (from tip of snout to rear of jaws);
HW head width (width of head at its widest point);
SL snout length (from tip of snout to anterior border of eye);
IND internarial distance (distance between nares);
IOD interorbital distance (minimum distance between upper eyelids);
UEW upper eyelid width (maximum width of upper eyelid);
ED eye diameter (diameter of exposed portion of eyeball);
TD tympanum diameter;
FHL forearm and hand length (from elbow to tip of third finger);
THL thigh length (from vent to knee);
TL tibia length (distance from knee to heel);
FL foot length (from proximal end of inner metatarsal tubercle to tip of fourth toe);
TFL length of foot and tarsus (from tibiotarsal joint to tip of fourth toe).
Comparative morphological data of other Gracixalus species were taken from their original descriptions or re-descriptions (Boulenger 1893, Hu et al. 1981, Ye and Hu 1984, Matsui and Orlov 2004, Orlov et al. 2004, Nguyen et al. 2008, Rowley et al. 2011, Mo et al. 2013, Nguyen et al. 2013, Rowley et al. 2014, Matsui et al. 2015, Matsui et al. 2017, Zeng et al. 2017, Chen et al. 2018, Wang et al. 2018).
Molecular analyses
Total genomic DNA was extracted from liver tissues. Tissue samples were digested using proteinase K, and subsequently purified following a standard phenol/chloroform isolation and ethanol precipitation. A fragment encoding partial 16S rRNA gene was amplified and sequenced following Yu et al. (2010). All new sequences have been deposited in GenBank under Accession Nos. MK234876–MK234883 (Table 1). Available homologous sequences of Gracixalus were obtained from GenBank. Rhacophorusborneensis Matsui, Shimada & Sudin, 2013 and Kurixalusidiootocus (Kuramoto & Wang, 1987) were selected as outgroups according to Matsui et al. (2017) and sequences of them were also downloaded from GenBank.
Table 1.
Species used in phylogenetic analysis of this study.
| Species | Locality | Voucher no. | GenBank no. |
|---|---|---|---|
| Rhacophorus borneensis | Sabah, Malaysia | BORN 22410 | AB781693 |
| Kurixalus idiootocus | Taiwan, China | KUHE 12979 | AB933306 |
| Gracixalus ananjevae | Nghe An, Vietnam | VNMN 03012 | JN862546 |
| Gracixalus gracilipes | Ha Giang, Vietnam | AMNH A163897 | DQ283051 |
| Pingbian, Yunnan, China | 060821196Rao | GQ285668 | |
| Lao Cai, Vietnam | AMS R 177672 | KT374014 | |
| Gracixalus guangdongensis | Hunan, China | CIB HN201108200 | LC011936 |
| Guangdong, China | SYS a004902 | MG520193 | |
| Guangdong, China | SYS a005750 | MG520197 | |
| Gracixalus jinggangensis | Mt. Jinggang, Jiangxi | SYS a003186 | KY624587 |
| Gracixalus jinxiuensis | Jinxiu, Guangxi, China | SYS a002182 | KY624584 |
| Jinxiu, Guangxi, China | SYS a002183 | KY624585 | |
| Jinxiu, Guangxi, China | KIZ 060821013 | EF564524 | |
| Jinxiu, Guangxi, China | KIZ 061210YP | EU215525 | |
| Gracixalus lumarius | Kon Tum, Vietnam | AMS R 176202 | KF918412 |
| Gracixalus nonggangensis | Guangxi, China | NHMG 200910010 | JX841318 |
| Gracixalus quyeti | Cha Noi, Vietnam | VNUH 160706 | EU871428 |
| Gracixalus quangi | Nghe An, Vietnam | AMS R173417 | JN862539 |
| Gracixalus sapaensis | Lao Cai, Vietnam | MNHN 1999.5961 | AY880503 |
| Lai Chau, Vietnam | IEBR 2351 | EU871425 | |
| Lao Cai, Vietnam | CIB XM-439 | GQ285670 | |
| Lao Cai, Vietnam | KUHE 46401 | LC011938 | |
| Lao Cai, Vietnam | KUHE 46402 | LC011939 | |
| Lao Cai, Vietnam | MNHN 1999.5966 | LC140970 | |
| Lao Cai, Vietnam | VNMN 4211 | LC140971 | |
| Lao Cai, Vietnam | VNMN 4212 | LC140972 | |
| Lao Cai, Vietnam | VNMN 4358 | LC140973 | |
| Gracixalus seesom | Kanchanaburi, Thailand | KUHE 35084 | LC011932 |
| Gracixalus supercornutus | Kon Tum, Vietnam | AMS R173887 | JN862545 |
| Gia Lai, Vietnam | AMS R176287 | KT374016 | |
| Gracixalus tianlinensis | Guangxi, China | NHMG 1705015 | MH117960 |
| Guangxi, China | NHMG 1705016 | MH117961 | |
| Gracixalus waza | Cao Bang, Vietnam | IEBR A.2012.2 | JX896681 |
| Cao Bang, Vietnam | VNMN A.2012.2 | JX896684 | |
| Gracixalus sp. | Wenshan, Yunnan, China | 03320Rao | GQ285669 |
| Gracixalusyunnanensis sp. n. | Houapan, Laos | KUHE 32453 | LC011937 |
| Lao Cai, Vietnam | VNMN 4355 | LC140985 | |
| Lao Cai, Vietnam | VNMN 4357 | LC140986 | |
| Lao Cai, Vietnam | VNMN 4371 | LC140987 | |
| Nghe An, Vietnam | AMS R173454 | JN862547 | |
| Jinping, Yunnan, China | KIZ 060821126 | EF564525 | |
| Lvchuan, Yunnan, China | GXNU YU000060 | MK234876 | |
| Bada, Menghai, Yunnan, China | KIZ 20160216 | MK234877 | |
| Xuelin, Lancang, Yunnan, China | KIZ 20160222 | MK234878 | |
| Xuelin, Lancang, Yunnan, China | KIZ 20160223 | MK234879 | |
| Fudong, Lancang, Yunnan, China | KIZ 20160226 | MK234880 | |
| Fazhanhe, Lancang, Yunnan, China | KIZ 20160228 | MK234881 | |
| Fazhanhe, Lancang, Yunnan, China | KIZ 20160229 | MK234882 | |
| Fazhanhe, Lancang, Yunnan, China | KIZ 20160230 | MK234883 |
Sequences were aligned using MUSCLE with the default parameters in MEGA version 7 (Kumar et al. 2016). Uncorrected pairwise distances between species were calculated in MEGA version 7. The best substitution model was selected using the corrected Akaike Information Criterion (AICc) in jMODELTEST version 2.1.10 (Darriba et al. 2012). Three methods were used to construct phylogeny of the genus Gracixalus. Firstly, Bayesian inference (BI) was performed in MRBAYES version 3.2.6 (Ronquist et al. 2012) based on the selected substitution model (TIM2 + I + G). Two runs were performed simultaneously with four Markov chains starting from random tree. The chains were run for 5,000,000 generations and sampled every 100 generations. Convergence and burn-in were checked using the program Tracer version 1.6. (Rambaut et al. 2014) and plot of the generation versus the log likelihood values. The first 25% of the sampled trees were discarded as burn-in and the remaining trees were used to create a consensus tree and to estimate Bayesian posterior probabilities (BPPs). Secondly, maximum likelihood (ML) analysis was conducted in RAXML-HPC version 8.2.10 (Stamatakis 2014) with 1000 rapid bootstrap replicates. Finally, a neighbor-joining (NJ) tree was constructed using PAUP* version 4.0b10 (Swofford 2002) and nodal supports were assessed by 1000 bootstrap replicates.
Results
The obtained alignment of 16S rRNA sequences is 543 bp in length after cutting off both ragged sides. The newly collected samples from Bada, Xuelin, Fudong, Fazhanhe, and Lvchun of Yunnan, China form a distinct lineage together with samples from Houapan of Laos (KUHE 32453), Nghe An (AMS R173454) and Lao Cai (VNMN 4355, 4357, 4371) of Vietnam, and Jinping of Yunnan (KIZ 060821126) that were sequenced by previous studies (Yu et al. 2008, Rowley et al. 2011, Matsui et al 2015, Matsui et al. 2017) (Figs 2, 3). Both Bayesian inference and Maximum likelihood analyses recovered this lineage as the sister to the clade consisting of G.ananjevae and Gracixalus sp. (GQ285669) with weak support (Fig. 2), whereas the NJ analysis revealed that it is closest to G.guangdongensis with weak support (Fig. 3). Average uncorrected pairwise distances (p-distance) between the new species and other species ranged from 2.2% (G.guangdongensis) to 14.1% (G.lumarius) (Table 2).
Figure 2.

Bayesian phylogram of Gracixalus inferred from 543 bp of 16S rRNA gene. Numbers above and below branches are Bayesian posterior probabilities and ML bootstrap values (only values above 50% are shown), respectively.
Figure 3.

Neighbor-joining (NJ) tree of Gracixalus inferred from 543 bp of 16S rRNA gene. Numbers above branches are bootstrap values (only values above 50% are shown).
Table 2.
Uncorrected p-distances (%) between Gracixalus species estimated from 16S rRNA sequences.
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Gracixalusyunnanensis sp. n. | – | |||||||||||||||
| 2 | G. ananjevae | 3.9 | – | ||||||||||||||
| 3 | Gracixalus sp. (GQ285669) | 5.1 | 2.3 | – | |||||||||||||
| 4 | G. sapaensis | 4.7 | 6.0 | 6.6 | – | ||||||||||||
| 5 | G. quangi | 8.0 | 9.6 | 9.5 | 9.1 | – | |||||||||||
| 6 | G. supercornutus | 9.2 | 10.9 | 10.7 | 10.5 | 3.0 | – | ||||||||||
| 7 | G. nonggangensis | 7.1 | 8.8 | 8.9 | 7.9 | 10.5 | 11.5 | – | |||||||||
| 8 | G. waza | 6.3 | 8.7 | 9.3 | 7.3 | 11.6 | 12.8 | 2.5 | – | ||||||||
| 9 | G. lumarius | 14.1 | 14.5 | 14.6 | 15.9 | 14.3 | 15.3 | 16.0 | 16.0 | – | |||||||
| 10 | G. gracilipes | 10.3 | 11.0 | 11.3 | 10.4 | 5.2 | 6.2 | 12.4 | 13.3 | 15.0 | – | ||||||
| 11 | G. jinxiuensis | 6.0 | 7.3 | 7.5 | 7.2 | 9.6 | 10.9 | 7.7 | 7.9 | 16.1 | 11.4 | – | |||||
| 12 | G. jinggangensis | 5.0 | 7.3 | 7.7 | 6.3 | 8.0 | 9.5 | 7.5 | 7.5 | 14.5 | 11.3 | 7.0 | – | ||||
| 13 | G. seesom | 8.6 | 10.4 | 9.7 | 8.8 | 6.0 | 6.7 | 10.2 | 10.1 | 16.0 | 6.1 | 9.6 | 9.9 | – | |||
| 14 | G. quyeti | 10.5 | 11.4 | 11.0 | 10.9 | 6.5 | 6.1 | 11.2 | 12.2 | 14.0 | 7.3 | 10.2 | 11.1 | 8.1 | – | ||
| 15 | G. tianlinensis | 4.3 | 6.3 | 6.5 | 3.0 | 9.4 | 10.5 | 7.2 | 6.4 | 14.8 | 10.3 | 5.9 | 6.5 | 7.8 | 10.0 | – | |
| 16 | G. guangdongensis | 2.2 | 4.6 | 5.4 | 5.0 | 7.8 | 9.5 | 7.2 | 6.7 | 14.2 | 10.1 | 5.6 | 5.3 | 8.3 | 10.6 | 4.4 | – |
Morphologically, these newly collected specimens can be distinguished from G.guangdongensis by a series of characters, including distinctive conical tubercles on dorsum (versus absent), lateral surfaces nearly smooth with no black blotches on ventrolateral region (versus lateral surfaces rough, scattered with tubercles and black blotches on ventrolateral region), snout rounded (versus triangularly pointed), iris bronze (versus iris brown), and ventral surface orangish (versus throat and chest creamy white and belly light brown). These specimens also differ from other members of Gracixalus in a series of characters. Herein we describe these specimens as a new species.
Gracixalus yunnanensis sp. n.
http://zoobank.org/1D19A62E-B4B2-4EDA-975D-4DCFD58DEDAD
Figure 4.

Dorsolateral (a) and ventral (b) views of the holotype of Gracixalusyunnanensis sp. n. in life and dorsal (c) and ventral (d) views of the holotype of Gracixalusyunnanensis sp. n. in preservative. Linea masculina is pointed by arrow.
Figure 5.

Ventral views of hand (a) and foot (b) of the holotype of Gracixalusyunnanensis sp. n. in preservative.
Figure 6.

Dorsal view of paratype KIZ 20160226 (a) and ventral view of paratype GXNU YU000060 (b).
Type material.
Holotype.KIZ 20160222, an adult male, collected at 21:05 on 1 June 2017 by Hong Hui from Xuelin Township, Lancang County, Yunnan Province, China (23°0'39.4"N, 99°31'54"E, 1864 m elevation).
Paratypes. Seven adult males: KIZ 20160223 collected at 21:05 on 1 June 2017 by Hong Hui from the type locality; KIZ 20160216 collected at 21:00 on 7 June 2014 by Hong Hui from Bada Township, Menghai County, Yunnan Province, China (21°50'8.9"N, 100°6'57.8"E, 1870 m elevation); KIZ 20160226 collected at 21:50 on 27 May 2017 by Hong Hui from Fudong Township, Lancang County, Yunnan Province, China (23°7'13.6"N, 99°58'33.9"E, 2166 m elevation); KIZ 20160228–20160230 collected at 21:40–22:15 on 10 June 2017 by Hong Hui from Fazhanhe Township, Lancang County, Yunnan Province, China (22°24'3.4"N, 100°12'4.2"E, 1822 m elevation); and GXNU YU000060 collected at 21:00 on 7 June 2018 by Jian Wang from Mt. Huanglian, Lvchun County, Yunnan Province, China (22°53'N, 102°18'E, 1918 m elevation).
Etymology.
The specific epithet yunnanensis refers to the distribution of this species in China, Yunnan Province.
Diagnosis.
The new species is assigned to genus Gracixalus based upon molecular data and the following morphological characters: the presence of intercalary cartilage between terminal and penultimate phalanges of digits, tips of digits enlarged to discs bearing circummarginal grooves, vomerine teeth absent, inner (first and second) and outer (third and fourth) fingers non-opposable, and an inversed Y-shaped dark brown marking on dorsum (Fei 1999, Rowley et al. 2011, Chen et al. 2018). The new species is distinguished from its congeners by a combination of 1) SVL 26.0–34.2 mm in males; 2) dorsal surface yellow brown or red brown; 3) distinctive conical tubercles on dorsum; 4) males with an external subgular vocal sac; 5) throat granular; 6) finger webbing rudimentary; 7) linea masculina, a band of connective tissue between the rectus abdominus muscle and oblique abdominus muscle, present in males; 8) tibiotarsal articulation reaching eye; 9) snout rounded; 10) white patch absent on temporal region; 11) tibiotarsal projection absent; 12) supratympanic fold distinct; 13) ventral surface orangish, nearly immaculate, and semi-transparent; 14) nuptial pads present on finger I; 15) heels overlapping when legs at right angle to body; 16) iris bronze; and 17) body sides nearly smooth with no black blotch.
Description of holotype.
Adult male (SVL 29.7 mm); head wider (HW 10.9 mm) than long (HL 9.5 mm); snout rounded, slightly projecting beyond margin of lower jaw in ventral view, rounded in profile; canthus rostralis rounded; loreal region oblique, slightly concave; nostril oval, protuberant, closer to tip of snout than eye; IND (2.9 mm) slightly narrower than IOD (3.0 mm) and wider than UEW (2.5 mm); eye large, horizontal diameter (ED 4.2 mm) equal to snout length (SL 4.2 mm); pupil oval, horizontal; pineal ocellus absent; tympanum distinct, diameter (TD 1.5 mm) smaller than half of ED; supratympanic fold distinct, extending from posterior corner of eye to above insertion of arm; vomerine teeth absent; tongue notched posteriorly; a pair of vocal sac slits on floor of mouth at both corners; an external subgular vocal sac.
Forelimb relatively robust; length of forearm and hand (FHL 14.1 mm) 47% of SVL; relative length of fingers I < II < IV < III; tips of all fingers expanded into discs with circummarginal grooves; disc of third finger large, slightly wider than tympanum; nuptial pads present on base of finger I; webbing between fingers rudimentary; subarticular tubercles prominent, rounded, single, formula 1, 1, 2, 2; supernumerary tubercles present; an inner metacarpal tubercle, oval; one outer metacarpal tubercle, rounded.
Heels overlapping when legs at right angle to body; tibiotarsal articulation reaching to middle of eye when hindlimb adpressed to body; relative length of toes I < II < III < V < IV; tips of toes expanded into discs with circummarginal grooves; discs of toes smaller than those of fingers; toes webbed, webbing formula I1.5–2II1.5–2.7III.5–3IV2.5–1.5V following Savage (1975); subarticular tubercles distinct and rounded, formula 1, 1, 2, 3, 2; supernumerary tubercles present; inner metatarsal tubercle oval; outer metatarsal tubercle absent.
Dorsal surface scattered with many small conical tubercles on head, upper eyelids, and dorsum; flanks of body and dorsal surface of limbs smooth, few small conical tubercles on hindlimbs and forearms; throat, chest, belly, and venter of thigh granulated; few small conical tubercles scattered on venter of thigh, tibia, and forearm.
Coloration of holotype.
In life, iris bronze; dorsal surface yellow brown with a dark brownish Y-shaped marking across back, covering interorbital region and posterior eyelids, bifurcating into two branches on the shoulder, and reaching the posterior of the back; limbs dorsally brown with dark brown bars; sides of head faint brown; flanks yellow brown, mottled with faint pink on lower part; minute dark spots densely scattered on lower part of flanks, temporal region, and upper jaw; skin of ventral surface semi-transparent, orangish with yellow spots; nuptial pads and discs faint yellow; linea masculina visible, white (Fig. 4b).
In preservative, color faded, pattern same as in life. Dorsal surface grayish brown, with a darker brown Y-shaped marking; dorsal side of limbs barred with dark brown; ventral surface of throat, chest, belly, forelimbs, and hindlimbs faded to whitish.
Morphological variation.
Measurements are shown in Table 3. Because the holotype and paratypes of the new species are all male, sexual dimorphism could not be determined. IOD is slightly wider than IND in holotype and most paratypes with the exception of KIZ 20160228, and TL is longer than FL in holotype and most paratypes with exceptions of KIZ 20160226 and KIZ 20160229.
Table 3.
Measurements (mm) of Gracixalusyunnanensis sp. n. Abbreviations defined in text.
| Voucher no. | Sex | SVL | HL | HW | SL | IND | IOD | UEW | ED | TD | DNE | DNS | FHL | THL | TL | TFL | FL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KIZ 20160216 | m | 30.0 | 9.5 | 11.4 | 4.0 | 3.4 | 3.5 | 2.5 | 4.4 | 1.9 | 2.4 | 1.8 | 15.4 | 13.1 | 13.8 | 20.2 | 13.3 |
| KIZ 20160222 | m | 29.7 | 9.5 | 10.9 | 4.2 | 2.9 | 3.0 | 2.5 | 4.2 | 1.5 | 2.4 | 1.7 | 14.1 | 12.3 | 13.2 | 18.8 | 12.9 |
| KIZ 20160223 | m | 28.5 | 9.4 | 10.5 | 4.0 | 2.9 | 3.0 | 2.5 | 4.0 | 1.8 | 2.2 | 1.6 | 13.9 | 12.6 | 13.0 | 18.7 | 12.5 |
| KIZ 20160226 | m | 34.2 | 10.1 | 12.1 | 4.7 | 3.3 | 3.8 | 2.8 | 4.4 | 1.9 | 2.5 | 2.1 | 15.6 | 13.3 | 14.1 | 21.1 | 14.2 |
| KIZ 20160228 | m | 28.7 | 9.4 | 11.0 | 4.2 | 3.0 | 3.0 | 2.5 | 4.1 | 1.6 | 2.4 | 1.7 | 14.6 | 12.6 | 13.3 | 19.2 | 12.8 |
| KIZ 20160229 | m | 26.0 | 9.0 | 9.4 | 3.5 | 2.6 | 2.8 | 2.3 | 3.5 | 1.3 | 2.0 | 1.3 | 12.8 | 11.0 | 11.6 | 17.0 | 11.6 |
| KIZ 20160230 | m | 26.4 | 8.3 | 10.0 | 3.8 | 2.9 | 3.3 | 2.5 | 3.8 | 1.6 | 2.2 | 1.7 | 13.4 | 12.0 | 12.7 | 18.3 | 11.8 |
| GXNU YU000060 | m | 27.3 | 8.7 | 9.5 | 4.1 | 2.7 | 2.8 | 2.5 | 3.7 | 1.7 | 2.3 | 1.7 | 13.4 | 11.9 | 12.7 | 18.8 | 12.3 |
Color of dorsal and ventral surfaces varied among individuals. Dorsal ground color of the holotype and four paratypes (KIZ 20160216, KIZ 20160223, KIZ 20160228, and KIZ 20160230) is yellow brown, and dorsal ground color of remaining paratypes (KIZ 20160226, KIZ 20160229, and GXNU YU000060) is red brown. Ventral surface of all specimens is nearly immaculate with the exception of paratype GXNU YU000060, which has dark marbling on throat, chest, and belly (Fig. 6). Conical tubercles on dorsum of specimens with red brown ground color are more distinct visually (Fig. 6).
Distribution.
In China, the new species is known from Yunnan (Lancang County, Menghai County, Lvchun County, and Jinping County). In addition, the new species also occurs in Laos (Houapan) and Vietnam (Lao Cai and Nghe An) because our molecular analyses revealed that samples from Houapan (KUHE 32453), Lao Cai (VNMN 4355, 4357, 4371), and Nghe An (AMS R173454) that were sequenced by previous studies also belong to the new species (Figs 2, 3). In Yunnan, specimens were found sitting on leaves of herbaceous plants (e.g., Amomumtsaoko and Eupatoriumadenophorum). No eggs and tadpoles were found.
Comparisons.
A summary of morphological comparisons presents in Table 4. The new species can be distinguished from G.ananjevae by having distinctive conical tubercles on dorsum (versus absent), sides of body smooth (versus coarsely granular), skin of throat granular (versus plain), and snout rounded (versus slightly pointed); from G.carinensis by having smaller body size in males (SVL 26.0–34.2 mm versus 30.2–38.1 mm), having distinctive conical tubercles on dorsum (versus absent), having an external vocal sac in males (versus an internal vocal sac), ventral surface orangish (versus white), and less developed toe webbing (Fig. 7); from G.gracilipes by having bigger body size in males (SVL 26.0–34.2 mm versus 20–24 mm), distinctive conical tubercles present on dorsum (versus absent), dorsal surface yellow brown or red brown (versus greenish), males with an external vocal sac (versus internal), throat granular (versus smooth), finger webbing rudimentary (versus absent), tibiotarsal articulation reaching to eye (versus reaching to between eye and nostril), snout rounded (versus triangular pointed), white patch absent on temporal region (versus present), tibiotarsal projection absent (versus present), and iris bronze (versus brown); and from G.guangdongensis by having distinctive conical tubercles on dorsum (versus absent), dorsal surface yellow brown or red brown (versus brown), flanks nearly smooth with no black blotches on ventrolateral region (versus flanks rough, scattered with tubercles and black blotches on ventrolateral region), snout rounded (versus triangularly pointed), ventral surface orangish (versus throat and chest creamy white and belly light brown), and iris bronze (versus iris brown).
Table 4.
Morphological characters for comparisons among Gracixalus species. “?” = not known or not clearly defined in the literature.
| Species | Adult male SVL (mm) | Conical tubercles on dorsum | Dorsal color in life | Vocal sac | Skin of body sides | Skin of throat | Finger webbing | Linea masculina | Tibiotarsal articulation | Snout | White patch on temporal region | Tibiotarsal projection | Supratympanic fold | venter | Nuptial pads | heels | iris |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G.yunnanensis sp. n. | 26.0–34.2 | present, small | yellow brown or red brown | external | smooth, no black blotches | granular | rudimentary | present, | reaching eye | rounded | absent | absent | distinct | orangish with yellow spots, immaculate, semi-transparent | on finger I | overlapping | bronze |
| G. ananjevae | 32 | absent | ? | ? | coarsely granular | plain | rudimentary | ? | reaching eye | slightly pointed | absent | absent | distinct | immaculate | on finger I | overlapping | ? |
| G. carinensis | 30.2–38.1 | absent | purplish, reddish, or greyish brown | internal | ? | granular | rudimentary | ? | reaching eye | rounded | absent | absent | distinct | immaculate white | ? | ? | ? |
| G. gracilipes | 20–24 | absent | greenish | internal | smooth with white stripe | smooth | rudimentary | ? | reaching eye | triangularly pointed | absent | absent | distinct | yellowish white | on fingers I and II | overlapping | brown |
| G. guangdongensis | 26.1–34.7 | absent | brown | ? | rough, black blotches | granular | absent | present | reaching between eye and nostril | triangularly pointed | present | present | distinct | throat and chest creamy white, belly light brown, semi-transparent | on finger I | overlapping | brown |
| G. jinggangensis | 27.9–33.8 | absent | brown to beige | ? | rough with tubercles | granular | rudimentary | ? | reaching eye | triangularly pointed | absent | absent | distinct | Throat and chest dirty white with dark specks, belly white anteriorly with dark marking and posteriorly yellowish, semi-transparent | on fingers I and II | just meeting | golden |
| G. jinxiuensis | 23.5–26.3 | ? | brown | internal | rough with tubercles | granular | rudimentary | absent | reaching eye | rounded | absent | absent | distinct | gray-brown with dark marbling | on finger I | just meeting | ? |
| G. lumarius | 38.9–41.6 | present | yellow | external | ? | granular | rudimentary | ? | ? | rounded | absent | absent | indistinct | opaque pink | on finger I | ? | dark gold |
| G. medogensis | 26.5 | absent | grass green | internal | ? | granular | absent | present | reaching eye | rounded | absent | absent | distinct | pale green | on finger I | overlapping | ? |
| G. nonggangensis | 29.9–35.3 | absent | yellowish-olive with dark-green mark | internal | rough with tubercles | granular | absent | absent | reaching tip of snout | rounded | absent | absent | distinct | white with dark marbling, semi-transparent | absent | overlapping | olive |
| G. quangi | < 25 | present, small | olive-green | external | with black blotches | smooth | absent | ? | ? | triangularly pointed | present | present | distinct | opaque white with translucent pale green margins | on finger I | ? | bronze |
| G. quyeti | ? | present | brownish to moss-green | ? | rough with sharp tubercles | smooth | rudimentary | ? | reaching to snout | rounded | absent | absent | indistinct | belly immaculate white | ? | overlapping | ? |
| G. sapaensis | 21–37 | absent | Golden ochre | ? | coarsely scattered with large tubercles | ? | rudimentary | ? | reaching eye | rounded | absent | absent | distinct | throat, chest, and belly light yellow, with dark marking | on finger I | overlapping | golden |
| G. seesom | 21.6–23.0 | absent | tan | ? | with large tubercles and white blotches | smooth | rudimentary | ? | reaching between eye and nostril | triangularly pointed | absent | absent | distinct | anterior belly opaque white and posterior belly translucent | absent | overlapping | golden |
| G. supercornutus | 22.0–24.1 | present, bigger horn-like | green with brown spots | ? | ? | granular | ? | ? | ? | pointed | present | present | distinct | light with white spots | ? | ? | ? |
| G. tianlinensis | 30.3–35.9 | absent | brown to beige | external | ? | granular | absent | ? | ? | rounded | absent | absent | distinct | throat and chest gray with dark specks, belly creamy white, opaque | on fingers I and II | ? | bronze |
| G. waza | 27.1–32.9 | absent | greyish-green to moss-green | ? | with small granulars | smooth | absent | ? | ? | rounded | absent | absent | distinct | Throat and chest white with dark marbling, belly immaculate white, semi-transparent | on finger I | overlapping | ? |
Figure 7.

Ventral views of foot of the holotype of Gracixalusyunnanensis sp. n. (a) and lectotype of Gracixaluscarinensis (b; reproduced from Matsui et al. 2017).
Gracixalusyunnanensis sp. n. differs from G.jinggangensis by having distinctive conical tubercles on dorsum (versus absent), flanks nearly smooth (versus rough with tubercles), snout rounded (versus triangularly pointed), ventral surface orangish and immaculate (versus throat and chest dirty white with dark specks, belly white anteriorly with dark marking and posteriorly yellowish), nuptial pads present only on finger I (versus nuptial pads present on fingers I and II), heels overlapping when hindlimbs held at right angles to the body (versus just meeting), and iris bronze (versus iris golden); from G.jinxiuensis by larger body size in males (SVL 26.0–34.2 mm versus 23.5–26.3 mm), males with an external vocal sac (versus vocal sac internal), flanks nearly smooth (versus rough with tubercles), linea masculina present (versus absent), ventral surface orangish and immaculate (versus ventral surface gray-brown with dark marbling), and sole of feet and palms smooth (versus rough with dense large tubercles); and from G.lumarius by smaller body size in males (SVL 26.0–34.2 mm versus 38.9–41.6 mm), dorsal surface yellow brown or red brown (versus yellow), and venter orangish and semi-transparent (versus venter opaque pink), supratympanic fold distinct (versus indistinct), and iris bronze (versus dark gold).
Gracixalusyunnanensis sp. n. can be distinguished from G.medogensis by having distinctive conical tubercles on dorsum (versus absent), dorsal surface yellow brown or red brown (versus grass green), males with an external vocal sac (versus an internal vocal sac), finger webbing rudimentary (versus absent), and venter orangish (versus pale green); from G.nonggangensis by having conical tubercles on dorsum (versus absent), dorsum yellow brown or red brown with a dark brown marking (versus yellowish-olive with a dark-green marking), males with an external vocal sac (versus internal), flanks smooth (versus rough with tubercles), finger webbing rudimentary (versus absent), linea masculina present in males (versus absent), tibiotarsal articulation reaching to eye (versus reaching to tip of snout), ventral surface immaculate (versus throat, chest, and belly white with dark marbling), nuptial pads present on finger I (versus absent), and iris bronze (versus olive); from G.quangi by having bigger body size in males (SVL 26.0–34.2 mm versus < 25 mm), dorsal surface yellow brown or red brown (versus olive-green), black spots absent on flanks and ventral surface of thighs (versus present), throat granular (versus smooth), finger webbing rudimentary (versus absent), snout rounded (versus triangular pointed), white patch absent on temporal region (versus present), tibiotarsal projection absent (versus present), and ventral surface orangish (versus opaque white with translucent pale green margins); and from G.quyeti by dorsal surface yellow brown or red brown (versus brownish to moss-green), flanks nearly smooth (versus rough with sharp tubercles), throat granular (versus smooth), tibiotarsal articulation reaching to eye (versus reaching to snout), supratympanic fold distinct (versus indistinct), and throat and chest immaculate (versus throat, margin of throat, and chest yellow-white with brown marbling).
Gracixalusyunnanensis sp. n. differs from G.sapaensis by having distinctive conical tubercles on dorsum (versus absent) and sides of body nearly smooth (versus coarsely scattered with large round tubercles); from G.seesom by bigger body size in males (SVL 26.0–34.2 mm versus 21.6–23.0 mm), conical tubercles present on dorsum (versus absent), dorsal surface yellow brown or red brown (versus tan), flanks nearly smooth with no white blotches (versus flanks with large tubercles and white blotches), throat granular (versus smooth), snout rounded (versus triangular pointed), nuptial pads present on finger I (versus absent), and iris bronze (versus golden); and from G.supercornutus by bigger body size in males (SVL 26.0–34.2 mm versus 22.0–24.1 mm), conical tubercles on dorsum small (versus considerable bigger horn-like projections in supraorbital area, around cloaca, and on dorsal surface, forelimbs and hindlimbs), dorsal surface yellow brown or red brown (versus greenish), snout rounded (versus triangular pointed), white patch absent on temporal region (versus present), and tibiotarsal projection absent (versus present).
The new species can be distinguished from G.tianlinensis by smaller body size in males (SVL 26.0–34.2 mm versus 30.3–35.9 mm), distinctive conical tubercles present on dorsum (versus absent), dorsal surface yellow brown or red brown (versus brown to beige), finger webbing rudimentary (versus absent), ventral surface orangish, immaculate, and semi-transparent (versus throat and chest gray with dark specks and belly creamy white, opaque), and nuptial pads present on finger I (versus on fingers I and II); and from G.waza by having distinctive conical tubercles on dorsum (versus absent), dorsal surface yellow brown or red brown (versus greyish-green to moss-green), throat granular (versus smooth), finger webbing rudimentary (versus absent), and ventral surface immaculate (versus throat and chest with dark marbling).
Discussion
Although G.yunnanensis sp. n. only diverges from G.guangdongensis by a distance of 2.2%, it can be morphologically separated from G.guangdongensis by a series of characters including distinctive conical tubercles on dorsum (versus absent), dorsal surface yellow brown or red brown (versus brown), flanks nearly smooth with no black blotches on ventrolateral region (versus flanks rough, scattered with tubercles and black blotches on ventrolateral region), snout rounded (versus triangularly pointed), ventral surface orangish (versus throat and chest creamy white and belly light brown), and iris bronze (versus iris brown) (Table 4). In addition, the new species has linea masculina (Fig. 4b), whereas G.guangdongensis likely lacks linea masculina (Fig. 8), although it was not described in Wang et al. (2018). Moreover, the new species and G.guangdongensis were recovered as reciprocally monophyletic and the new species is not directly related to G.guangdongensis or other known congeners with strong support (Figs 2, 3). Therefore, we think that G.yunnanensis sp. n. should be diagnosed as an independent species. It has been revealed that low interspecific genetic distance seems to be very common in frogs from Southeast Asia (e.g., 2.2%−21.2% in Megophryidae, 1.8%−16.0% in Ranidae, and 1.5%−19.8% in Rhacophoridae; Grosjean et al. 2015).
Figure 8.

Ventral view of male holotype of G.guangdongensis (SYS a005724) in life (reproduced from Wang et al. 2018).
Historically, G.yunnanensis sp. n. was once confused with G.jinxiuensis in that the Jinping specimen (KIZ 060821126), Houapan specimen (KUHE 32453), Nghe An specimen (AMS R173454), and Lao Cai specimens (VNMN 4355, 4357, 4371) were originally identified as G.jinxiuensis by Yu et al. (2008), Matsui et al. (2015), Rowley et al. (2011), and Matsui et al. (2017), respectively. However, the new species can easily be distinguished from G.jinxiuensis by having bigger body size, an external vocal sac, and linea masculina in males (Table 4).
Gracixalus now contains a total of 17 species and our phylogenetic analyses revealed that this genus consists of three major clades, one consisting of G.lumarius (Clade I), one consisting of G.seesom, G.quyeti, G.quangi, G.supercornutus, and G.gracilipes (Clade II), and one consisting of all other species (Clade III) (Figs 2, 3). This result is consistent with Zeng et al. (2017), Chen et al. (2018), and Wang et al. (2018). However, like these previous studies, the present study did not achieve a complete resolution of phylogenetic relationships among these three clades and phylogenetic relationships within clades II and III. Thus, more studies will be needed to resolve the phylogenetic relationships among this genus. Additionally, taxonomic confusions still exist in Gracixalus: Matsui et al. (2015) and Wang et al. (2018) considered that G.nonggangensis should be synonymized with G.waza because of low genetic distance between them. However, morphologically, males of G.waza have developed nuptial pads on finger I according to Nguyen et al. (2013), whereas males of G.nonggongensis lack nuptial pad according to Mo et al. (2013). If indeed this is the case, we would prefer that G.nonggangensis and G.waza represent two different species. Furthermore, cryptic species might exist in G.nonggangensis because its monophyly was not supported in Matsui et al. (2015) and Wang et al. (2018). In addition, studies will be necessary to confirm whether the specimen from Wenshan, Yunnan, China (voucher number: 03320Rao; GenBank accession no.: GQ285669) belongs to G.ananjevae or not. We found that they are sister to each other with strong support values (Figs 2, 3), which is consistent with Mo et al. (2013); the genetic distance between them is moderate (2.3%; Table 2).
Supplementary Material
Acknowledgements
We thank Tan He and Wenjing Jiang for their assistances with fieldworks. Thanks also to the three reviewers for their helpful comments on the manuscript. This work was supported by the National Natural Science Foundation of China (Nos. 31872212 and 31301870) and CAS “Light of West China” Program to Guohua Yu.
Citation
Yu G, Hui H, Wang J, Rao D, Wu Z, Yang J (2019) A new species of Gracixalus (Anura, Rhacophoridae) from Yunnan, China. ZooKeys 851: 91–111. https://doi.org/10.3897/zookeys.851.32157
Footnotes
These authors contributed equally to this work
Contributor Information
Dingqi Rao, Email: raodq@mail.kiz.ac.cn.
Zhengjun Wu, Email: wu_zhengjun@aliyun.com.
Junxing Yang, Email: yangjx@mail.kiz.ac.cn.
References
- Boulenger GA. (1893) Concluding report on the reptiles and batrachians obtained in Burma by Signor L. Fea dealing with the collection made in Pegu and the Karin Hills in 1887−88. Annali del Museo Civico di Storia Naturale di Genov 13: 304–347. 10.5962/bhl.part.9543 [DOI] [Google Scholar]
- Bourret R. (1937) Notes herpétologiques sur l’Indochine française. XIV. Les batraciens de la collection du Laboratoire des Sciences Naturelles de l’Université. Descriptions de quinze espèces ou variétés nouvelles. Annexe au Bulletin Général de l’Instruction Publique 1937: 5–56. 10.5962/bhl.part.22065 [DOI] [Google Scholar]
- Chen W, Bei Y, Liao X, Zhou S, Mo Y. (2018) A new species of Gracixalus (Anura: Rhacophoridae) from West Guangxi, China. Asian Herpetological Research 9: 74–84. 10.11646/zootaxa.3616.1.5 [DOI] [Google Scholar]
- Darriba D, Taboada GL, Doallo R, Posada D. (2012) jModelTest 2: more models, new heuristics and parallel computing. Nature Methods 9: 772–772. 10.1038/nmeth.2109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delorme M, Dubois A, Grosjean S, Ohler A. (2005) Une nouvelle classification générique et subgénérique de la tribu des Philautini (Amphibia, Anura, Rhacophorinae). Bulletin Mensuel de la Société Linnéenne de Lyon 74: 165–171. 10.3406/linly.2005.13595 [DOI] [Google Scholar]
- Fei L. (1999) Atlas of Amphibians of China. Henan Publishing House of Science and Technology, Zhengzhou, China, 432 pp. [Google Scholar]
- Frost DR. (2018) Amphibian Species of the World: and Online Reference. Version 6.0. American Museum of Natural History, New York, USA. http://research.amnh.org/herpetology/amphibia/index.html/ [Accessed 27 November 2018]
- Grosjean S, Ohler A, Chuaynkern Y, Cruaud C, Hassanin A. (2015) Improving biodiversity assessment of anuran amphibians using DNA barcoding of tadpoles. Case studies from Southeast Asia. Comptes Rendus Biologies 338: 351–361. 10.1016/j.crvi.2015.03.015 [DOI] [PubMed] [Google Scholar]
- Hu S, Fei L, Ye C. (1978) Three new amphibian species in China. Materials for Herpetological Research 4: 20.
- Hu S, Tian W, Wu G. (1981) Three new species of Amphibians from Guangxi. Acta Herpetological Sinica 17: 111–120. [Google Scholar]
- Kumar S, Stecher G, Tamura K. (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870–1874. 10.1093/molbev/msw054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuramoto M, Wang C. (1987) A new rhacophorid treefrog from Taiwan, with comparisons to Chirixaluseiffingeri (Anura, Rhacophoridae). Copeia 1987: 931–942. 10.2307/1445556 [DOI] [Google Scholar]
- Matsui M, Khonosue W, Panha S, Eto K. (2015) A new tree frog of the genus Gracixalus from Thailand (Amphibia: Rhacophoridae). Zoological Science 32: 204–210. 10.2108/zs140238 [DOI] [PubMed] [Google Scholar]
- Matsui M, Ohler A, Eto K, Nguyen TT. (2017) Distinction of Gracixaluscarinensis from Vietnam and Myanmar, with description of a new species. Alytes 33: 25–37. [Google Scholar]
- Matsui M, Orlov N. (2004) A new species of Chirixalus from Vietnam (Anura: Rhacophoridae). Zoological Science 21: 671–676. 10.2108/zsj.21.671 [DOI] [PubMed] [Google Scholar]
- Matsui M, Shimada T, Sudin A. (2013) A new gliding frog of the genus Rhacophorus from Borneo. Current Herpetology 32: 112–124. 10.5358/hsj.32.112 [DOI] [Google Scholar]
- Mo Y, Zhang W, Luo Y, Zhou S, Chen W. (2013) A new species of the genus Gracixalus (Amphibia: Anura: Rhacophoridae) from Southern Guangxi, China. Zootaxa 3616: 61–72. 10.11646/zootaxa.3616.1.5 [DOI] [PubMed] [Google Scholar]
- Nguyen TQ, Hendrix R, Böhme W, Vu TN, Ziegler T. (2008) A new species of the genus Philautus (Amphibia: Anura: Rhacophoridae) from the Truong Son Range, Quang Binh Province, central Vietnam. Zootaxa 1925: 1–13. [Google Scholar]
- Nguyen TQ, Le MD, Pham CT, Nguyen TT, Bonkowski M, Ziegler T. (2013) A new species of Gracixalus (Amphibia: Anura: Rhacophoridae) from northern Vietnam. Organisms Diversity & Evolution 13: 203–214. 10.1007/s13127-012-0116-0 [DOI] [Google Scholar]
- Orlov NL, Ho TC, Nguyen QT. (2004) A new species of the genus Philautus from central Vietnam (Anura: Rhacophoridae). Russian Journal of Herpetology 11: 51–64. [Google Scholar]
- Rambaut A, Suchard MA, Xie D, Drummond AJ. (2014) Tracer v1.6. http://tree.bio.ed.ac.uk/softeare/tracer
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. 10.1093/sysbio/sys029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowley JJL, Dau QV, Nguyen TT, Cao TT, Nguyen SN. (2011) A new species of Gracixalus (Anura: Rhacophoridae) with a hyperextended vocal repertoire from Vietnam. Zootaxa 3125: 22–38. 10.1163/15685381-00003007 [DOI] [Google Scholar]
- Rowley JJL, Le DTT, Dau VQ, Hoang HD, Cao TT. (2014) A striking new species of phytotelm-breeding tree frog (Anura: Rhacophoridae) from central Vietnam. Zootaxa 3785: 25–37. 10.11646/zootaxa.3785.1.2 [DOI] [PubMed] [Google Scholar]
- Savage JM. (1975) Systematics and distribution of the Mexican and Central American stream frogs related to Eleutherodactylus rugulosus. Copeia 1975: 254–306. 10.2307/1442883 [DOI] [Google Scholar]
- Stamatakis A. (2014) RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30: 1312–1313. 10.1093/bioinformatics/btu033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swofford DL. (2002) PAUP: Phylogenetic Analysis Using Parsimony (*and other Methods), Version 4.0. Sinauer Associates, Sunderland, MA.
- Wang J, Zeng Z, Lyu Z, Liu Z, Wang Y. (2018) Description of a new species of Gracixalus (Amphibia: Anura: Rhacophoridae) from Guangdong Province, southeastern China. Zootxa 4420: 251–269. 10.11646/zootaxa.4420.2.7 [DOI] [PubMed] [Google Scholar]
- Ye C, Hu S. (1984) A new species of Philautus (Anura: Rhacophoridae) from Xizang Autonomous Region. Acta Herpetologica Sinica 3: 67–69. [Google Scholar]
- Yu G, Zhang M, Yang J. (2010) A species boundary within the Chinese Kurixalusodontotarsus species group (Anura: Rhacophoridae): New insights from molecular evidence. Molecular Phylogenetics and Evolution 56: 942–950. 10.1016/j.ympev.2010.05.008 [DOI] [PubMed] [Google Scholar]
- Zeng Z, Zhao J, Chen C, Chen G, Zhang Z, Wang Y. (2017) A new species of the genus Gracixalus (Amphibia: Anura: Rhacophoridae) from Mount Jinggang, southeastern China. Zootaxa 4250: 171–185. 10.11646/zootaxa.4250.2.3 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.