

Summary
Bacterial cellulose is a strong and flexible biomaterial produced at high yields by Acetobacter species and has applications in health care, biotechnology and electronics. Naturally, bacterial cellulose grows as a large unstructured polymer network around the bacteria that produce it, and tools to enable these bacteria to respond to different locations are required to grow more complex structured materials. Here, we introduce engineered cell‐to‐cell communication into a bacterial cellulose‐producing strain of Komagataeibacter rhaeticus to enable different cells to detect their proximity within growing material and trigger differential gene expression in response. Using synthetic biology tools, we engineer Sender and Receiver strains of K. rhaeticus to produce and respond to the diffusible signalling molecule, acyl‐homoserine lactone. We demonstrate that communication can occur both within and between growing pellicles and use this in a boundary detection experiment, where spliced and joined pellicles sense and reveal their original boundary. This work sets the basis for synthetic cell‐to‐cell communication within bacterial cellulose and is an important step forward for pattern formation within engineered living materials.
Introduction
Cellulose is a simple, yet versatile glucose polymer from which biology weaves a broad array of materials. It is one of nature's most abundant polymers, and can form the basis of materials that are light and elastic, such as a loofah, or structures that are strong and stiff like bamboo (Youssefian and Rahbar, 2015; Chen et al., 2017). Cellulose is produced most abundantly by plants, however, a number of bacterial species naturally overproduce cellulose as part of their biofilm matrix, making a structure known as a pellicle (Römling and Galperin, 2015). Bacterial cellulose differentiates itself from plant cellulose in that it is typically produced as ultrapure nanocellulose fibres, free from the contaminating polymers like pectin and lignin that are coproduced by plant cells. Bacterial cellulose (BC) is well known for being strong and flexible, with a single nanofiber having the tensile strength of ~1 GPa and Young's modulus of 114 GPa; while also being highly hydrophilic, with water making ~90% of its weight in its wet state (Lee et al., 2014). These material properties have seen commercial uptake of bacterial cellulose for biomedical and cosmetic applications, such as for protective bandaging, while also offering potential as a material used in electronics, for example as a battery separator or as a matrix ingredient in organic light‐emitting diode (OLED) displays (Lee et al., 2014; Jang et al., 2017). Some BC‐producing bacteria have also been demonstrated to be amenable to genetic engineering (Florea et al., 2016), offering the possibility of using cellulose‐producing bacteria as a means to produce genetically defined materials, where the material properties and functions can be programmed by engineering at the DNA level using the modern tools of synthetic biology (Cameron et al., 2014). Programming material properties and features into biomaterials using such tools is part of the emerging new field of engineered living materials (ELMs; Nguyen et al., 2018).
In most natural systems that produce biomaterials, communication between material‐producing cells is a critical component of producing materials with structures on the microscale and macroscale. ELMs research can theoretically recapitulate this in material‐producing bacteria by leveraging the significant past work done to engineer Escherichia coli for synthetic pattern formation (Scholes and Isalan, 2017). Such engineered pattern formation work typically exploits quorum‐sensing systems from various bacteria, which are a natural method of bacterial cell‐to‐cell communication that can be re‐programmed (Scholes and Isalan, 2017). The Lux quorum‐sensing system from Vibrio fischeri is particularly well‐used in this context, utilizing the signalling molecule 3OC6‐HSL – often referred to generally as acyl‐homoserine lactone (AHL;Churchill et al., 2011). AHL is produced by the enzyme LuxI, this diffusible molecule can then bind and activate the transcription factor LuxR, which in activates the pLux promoter to drive expression of a downstream gene (Waters and Bassler, 2005). The diffusible and inducible nature of AHLs can be exploited to create a morphogen gradient – the basis for many models of patterning, such as the French Flag Model (Wolpert, 1969) and Turing patterns (Turing, 1952; Fig. 1A). In synthetic pattern formation research, the Lux system has been used to produce programmable bullseye patterns on dishes of engineered E. coli cells that produce AHL ‘Senders’, while being surrounded by a lawn of cells that express fluorescent proteins in response to different levels of AHL (‘Receivers’;Basu et al., 2005). Since then, more complex systems have been engineered using AHL signalling in E. coli, including tuneable stripe patterns with motile cells (Liu et al., 2011), hierarchical patterning with multiple AHLs (Boehm et al., 2018) and the recent creation of stochastic Turing patterns (Karig et al., 2018).
Figure 1.
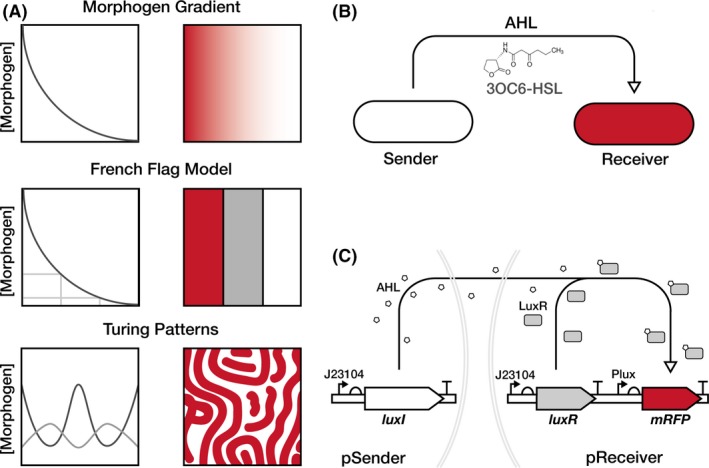
Design of a unidirectional cell‐to‐cell communication system.A. Illustration of diffusion‐driven patterning models. At the top is a morphogen gradient, a simple form of patterning that is also the basis for the French Flag model, presented immediately below, and the even more complex Turing patterns presented in the bottom row. The French Flag model requires sensing the concentration of a morphogen to differentiate the response, while Turing patterns require two morphogens that regulate each other and diffuse at different rates.B. The cell‐to‐cell communication system use here is made up of two different cells, a Sender cell producing the N‐(3‐oxohexanoyl) homoserine lactone AHL molecule and a Receiver cell that activates mRFP expression (red) in response to that AHL.C. The individual plasmid constructs used in this study. The pSender plasmid constitutively expresses the AHL synthase LuxI, and the pReceiver plasmid constitutively expresses the transcriptional activator LuxR. When LuxR is bound to AHL it upregulates expression from the PLux promoter which is this case activates expression of a monomeric red fluorescent protein (mRFP) gene. Both luxI and luxR are expressed in these constructs with constitutive promoter J23104.
Previously, we isolated a genetically tractable BC‐producing Komagataeibacter rhaeticus strain and developed a basic kit of synthetic biology parts and tools to enable its genetic engineering (Florea et al., 2016). Here, we extend our initial K. rhaeticus toolkit to now add engineered cell‐to‐cell communication to enable downstream work in patterning structural and functional properties into BC‐based ELMs. By modifying the AHL‐based systems developed previously for E. coli, we introduce and characterize cell‐to‐cell communication between engineered sender and receiver K. rhaeticus cells and demonstrate this system to be active within pellicles and able to define the boundaries between pellicle regions.
Results
Construction of Sender and Receiver plasmids
To establish synthetic cell‐to‐cell communication in K. rhaeticus, we designed two separate strains, a Sender strain that produces the diffusible AHL molecule N‐(3‐oxohexanoyl) homoserine lactone (3OC6‐HSL), and a Receiver strain that can sense AHL and respond by expressing an easy‐to‐measure reporter gene (Fig. 1B). The sender plasmid, pSender, was constructed by placing the AHL‐synthesis gene luxI downstream of the synthetic constitutive promoter J23104 (Fig. 1C). The receiver plasmid, pReceiver, used promoter J23104 for expression of the gene luxR and the LuxR‐regulated promoter, pLux for expression of the monomeric red fluorescent protein (mRFP) gene (Fig. 1C). Previous work has shown that the B0034 ribosome binding site was effective in K. rhaeticus (Florea et al., 2016), and it was therefore used upstream of luxI, luxR and mRFP coding sequences. The J23104 constitutive promoter was used in both instances as it was previously shown to have high activity in K. rhaeticus (Florea et al., 2016).
AHL production by Sender strains
We first determined whether our engineered Sender K. rhaeticus cells produced and released meaningful quantities of AHL to trigger cell‐to‐cell communication. To measure effective AHL production levels, we used E. coli transformed with pReceiver as a bioassay system. Similar to other E. coli receiver‐like plasmids (Canton et al., 2008). pReceiver gave a well‐characterized increase in reporter gene expression in response to increasing amounts of AHL (Fig. S1). We grew K. rhaeticus Sender cells, and a control K. rhaeticus containing only an empty plasmid (pEmpty), collected their spent growth media and treated the E. coli Receiver cells with this media. As HS media has a low pH, it buffered before use and experiments confirmed that it did not effect on E. coli growth (Fig. S2). Spent growth media produced by K. rhaeticus Sender cells induced mRFP production in E. coli (Fig. 2A). Interestingly, detectable levels of E. coli mRFP production were also observed when spent growth media from the K. rhaeticus pEmpty control cells were used, but not when unused HS media was added.
Figure 2.
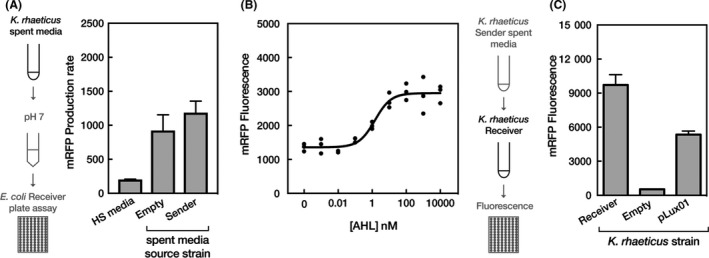
Production and effect of AHL produced by K. rhaeticus Sender strains (A) Testing the ability of K. rhaeticus Sender to produce AHL. The left illustration demonstrates the process of testing the AHL concentration in K. rhaeticus spent media. The right chart shows the mean mRFP production rate response from an E. coli Receiver assay after the addition of fresh HS media and spent media from K. rhaeticus Empty and K. rhaeticus Sender. Error bars denote the standard deviation from three repeats.B. Dose–response of K. rhaeticus Receiver 2 h post‐treatment with increasing concentrations of AHL. The response is measured in fluorescence units and is normalized with a media blank on the same plate. n = 3 for all concentrations except for 0.1 nM AHL where n = 2.C. The mean mRFP fluorescence response of liquid K. rhaeticus Receiver 1‐h postaddition of spent media from K. rhaeticus Sender. The left illustration details the process of adding spent media to K. rhaeticus and the method used to measure the fluorescence response. Treated strains included K. rhaeticus Receiver, the original pLux01 vector and K. rhaeticus with pEmpty. Error bars denote the standard deviation from three repeats.
Next, the engineered K. rhaeticus Receiver cells were assessed for their ability to respond to AHL and express the mRFP reporter gene. In order to confirm the new K. rhaeticus Receiver strain responded to AHL, a dose–response curve using standards of synthetic AHL was first generated (Fig. 2B). Thereafter, spent media from K. rhaeticus Sender cells was applied to K. rhaeticus Receiver liquid cultures (and control cultures), and mRFP levels were determined 1 h after induction (Fig. 2C). K. rhaeticus Receiver strains produced significant levels of mRFP in response to K. rhaeticus Sender spent media, giving approximately 10‐fold more red fluorescence than background levels (by comparing pEmpty control strains against K. rhaeticus Receiver). The engineered pReceiver plasmids, using the J23104 promoter for luxR expression, also showed improvement in the readout of the AHL‐response system in K. rhaeticus, producing more mRFP from the same input when compared to the previously described pLux01 construct (Florea et al., 2016) that uses a weaker promoter for luxR expression (Fig. 2B).
Cell‐to‐cell communication between Sender and Receiver strains
Having determined that Sender and Receiver K. rhaeticus could produce and respond to appropriate levels of AHL in isolation, we next examined direct cell‐to‐cell communication between mixed cultures of the two strains. To investigate interactions at the single‐cell level, we first examined our co‐cultures via fluorescence microscopy. This was enabled using a microfluidic plate set‐up that provides continual slow perfusion of new growth media. The set‐up allowed K. rhaeticus to grow in a single focal plane and yielded single‐cell resolution images of the cultures. Sender and Receiver K. rhaeticus strains were mixed at low density in a 1:1 ratio and incubated at 30°C in the microfluidic plate system for time‐lapse imaging. After 48 h, images of the co‐culture were taken and these revealed strong expression of mRFP in clusters of cells surrounded by those not showing any red signal (Fig. 3A, left). These expressing clusters are presumably Receiver cells that have grown from an ancestor while surrounded by the non‐fluorescent Sender cells. To verify this, we imaged Receiver cells grown mixed with pEmpty control cells and exposed them to 100 nM of synthetic AHL and saw the same pattern. Cells of just the K. rhaeticus Receiver strain were also grown in the set‐up and showed only very low‐level background red fluorescence (Fig. 3A, right). However, when Receiver cells were grown in liquid phase with 100 nM of synthetic AHL and treated with cellulase they should uniform activation of mRFP expression as determined by flow cytometry (Fig. S3). These controls confirm that engineered expression of AHL from Sender strains triggers significant expression of the reporter gene in Receiver K. rhaeticus cells when strains are co‐cultured together.
Figure 3.
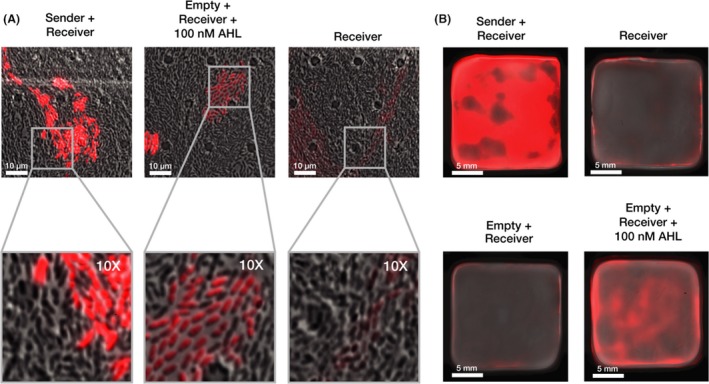
Cell‐to‐cell signalling functions within co‐cultured pellicles.A. Fluorescence microscopy of co‐cultures. Images detail K. rhaeticus growth after 48 h within three separate chambers on the same microfluidics plate. K. rhaeticus co‐cultures were produced from a 1:1 inoculation of each strain. Images were taken with a 60× oil emulsion lens. The bottom column displays a digitally zoomed in region of the images in the top. All microscopy images were taken with the same settings for brightfield and mRFP fluorescence channels. Brightness and contrast for the brightfield channel were adjusted to improve clarity, while the red fluorescence channel was left unadjusted (B) Pellicle co‐cultures. The top left pellicle is a 1:1 mix of Sender and Receiver culture and the top right a pure Receiver pellicle. Both the bottom pellicles are a 1:1 mix of Empty and Receiver, where the bottom left was left to grow without AHL while the bottom right had 100 nM AHL added to it on second day of growth.
The results so far showed that our engineered K. rhaeticus behave similarly to E. coli engineered with similar constructs in past work, however, unlike E. coli, K. rhaeticus, naturally produces and grows within a macroscale material structure. While giving many benefits in terms of downstream applications, the growth of the bacteria in a pellicle may alter the way that gene circuits operate, for example by having the cells switch to stationary phase or making cell conditions anaerobic and thus changing the cell state. Therefore, to determine if the engineered communication system can function in this environment, we next switched to pellicle growth experiments using co‐cultures of our Sender and Receiver strains. A 1:1 inoculation ratio of Sender and Receiver K. rhaeticus was set‐up and grown alongside equivalent growth of the same control combinations used in the microscopy experiment (Fig. 3B). After 4 days, this gave pellicles with the thickness of approximately 5 mm, and when scanned using a fluorescence imager, significant mRFP expression was observed across the pellicle in the Sender+Receiver combination. No fluorescence was observed with just Receiver cells or in the control pellicle where pEmpty was present in cells mixed with the K. rhaeticus Receiver. Fluorescence was observed when synthetic AHL was added exogenously at 100 nM AHL to this control pellicle (Fig. 3B), although the signal was not as strong as for the engineered co‐culture, presumably because local AHL concentrations are higher and continually produced when it is being produced within the material.
Cell‐to‐cell communication between Sender and Receiver pellicles
Past work in E. coli synthetic biology has established that there are many downstream applications once cells within a co‐culture can be engineered to communicate (Basu et al., 2005; Boehm et al., 2018; Karig et al., 2018). Given the unique situation of having now established such a system in a material‐producing microbe, we decided to investigate if it could be used to mediate communication between whole pieces of grown materials. In engineered living materials this could be used, for example, to determine the edges of grown areas and trigger gene expression at section boundaries. To demonstrate this, Sender and Receiver K. rhaeticus pellicles were grown separately, before being placed adjacent to one another in a plate with HS media for 24 h. Control experiments were also done in parallel, where pellicles from cells containing pEmpty were placed with Receiver pellicles and coincubated for 24 h with and without exogenous synthetic AHL. As before pellicles were imaged using a fluorescent scanner, and the images of the pellicle pairs show that the Receiver pellicle only activates mRFP expression when coincubated with the Sender pellicle or when exogenous AHL was added (Fig. 4A). The signal is strongest at the pellicle edges, representing the areas that see new growth of cells and have the maximum exposure to diffusing AHL coming from the Sender pellicles.
Figure 4.

Cell‐to‐cell signalling can function between separate Sender and Receiver pellicles. All pellicle images have two colour channels, a pseudo‐brightfield and red fluorescence channel.
A. Receiver pellicle induction by Sender pellicle. Left images show two Receiver pellicles after 24 h incubation in 5 ml HS media, the top pellicle without any added AHL and the bottom pellicle with 100 nM AHL. The right images show Receiver‐Sender and Empty‐Sender pellicle pairs after 24 h co‐incubation in 5 ml HS media.
B. Boundary detection. Spliced Sender‐Receiver pellicle was grown for 24 h and then removed from soft agar surface and imaged. A digitally zoomed in region of the boundary between the two pellicles is featured.
Having verified that the Sender pellicle can produce enough AHL to diffuse out and induce gene expression in Receiver pellicles in just 24 h, we next moved to boundary detection. In this case, cut Sender and Receiver pellicles were placed adjacent to one another and left to continue to grow in media so that they fuse together to make a larger piece. This technique of fusing pellicles via growth can aid in growing large material products. After only 24 h incubation of closely adjacent pellicles, fusion had occurred along most of the boundary, now creating a single pellicle. This was imaged as before and revealed a clear line of strong mRFP expression along the edge of the previous Receiver pellicle. This line of expression reveals where the border of the fusion has occurred and in future engineered strains could be the location of the expression of other proteins, such as those that provide stronger material bonding (Fig. 4B).
Discussion
Here, we constructed a unidirectional cell‐to‐cell communication system in the cellulose‐producing bacteria K. rhaeticus using a system of two separate engineered strains: Sender and Receiver. We utilized the Lux quorum‐sensing system, commonly used in artificial pattern formation experiments, and showed it to be functional in K. rhaeticus, even within growing bacterial cellulose pellicles. Previous research on synthetic patterning of bacterial cells has entirely focused on patterns formed by colonies on agar surfaces, cells in microfluidic chambers or multiwell plates, whereas here we demonstrate this system working as the engineered cells grow in and around their own macromaterial structure.
The work presented here offers a route towards the development of more intricate gene regulation and signalling circuits within growing pellicles made by K. rhaeticus, and will be enabled in the future by the addition of further signalling and regulatory components. Increasing the complexity of cell‐to‐cell communication, for example by bringing in a second diffusible signalling molecule, would allow the realization of more elaborate patterns, such as Turing patterns, and also enable bidirectional communication between engineered cells.
Interestingly, our initial characterization with E. coli Receiver strains showed that unengineered K. rhaeticus produces other molecules that partially trigger the Lux system response in E. coli (Fig. 2A). This hints at an unknown native quorum system in K. rhaeticus which could be further investigated and potentially exploited as a second communication route. While this triggered some expression in Receiver E. coli, it notably did not cause a response in Receiver K. rhaeticus (Fig. 3), suggesting that it is not the exact same AHL molecule. A genomic search of K. rhaeticus only identifies an orphaned luxR homologue and no obvious AHL synthases (Florea et al., 2016); however, recent work has shown that a closely related Acetobacter strain does produce quorum molecules (Liu et al., 2018).
A consideration for future work with our system is how activated gene expression in our engineered strains impairs cell growth. As cellulose production by K. rhaeticus prohibits accurate measurement of cell count by optical density, it is a challenge to measure how growth rate is altered by the induced expression of genes such as mRFP. It is notable that after 2 days of growth and imaging, Sender cells visually outnumbered Receiver cells in the microfluidic device despite starting at a 1:1 ratio (Fig. 3A). This suggests that continual mRFP expression (and the imaging of this) is a burden to the Receiver cells. Downstream users of the cell‐to‐cell signalling modules described here may wish to consider how expression burden could affect the performance of engineered consortia over time.
A direct potential utility of the boundary detection shown here (Fig. 4), could be to create a self‐repairing material that re‐joins cuts with stronger attachment than just natural pellicle fusion. This could be achieved by replacing the mRFP gene in the pReceiver construct by genes that express proteins that promote stronger attachment, for example by the surface display of adhesive catecholamines (Park et al., 2014) or via production of bacterial adhesive fibres such as Curli (Nguyen et al., 2014). Our work could also be further expanded by combining our signalling with genetic logic gates that link cell‐to‐cell communication with decision‐making and the response to combinations of external and internal stimuli (Brophy and Voigt, 2014; Scott and Hasty, 2016; Liu et al., 2018). Overall, we believe that a living bacterial cellulose material capable of programmable cell‐to‐cell communication could be adapted for a variety of purposes, and further engineered to create patterned materials with unique macrostructure‐driven material properties and novel functions.
Experimental procedures
Strains, plasmids and culturing conditions
Escherichia coli was cultured in at 37°C shaking at 250 rpm in Lysogeny broth (LB; 10 g l−1 Tryptone, 5 g l−1 Yeast Extract and 5 g l−1 NaCl) or statically at 37°C in LB agar with 34 μg ml−1 chloramphenicol when appropriate. Cultures of K. rhaeticus iGEM were grown at 30°C in Hestrin–Schramm media (HS; 20 g l−1 glucose, 10 g l−1 yeast extract, 10 g l−1 peptone, 2.7 g l−1 Na2HPO4 and 1.3 g l−1 citric acid, pH 5.6–5.8) or on HS agar plates (1,5% agar) and supplemented with 2% cellulase, when appropriate 34 μg ml−1 chloramphenicol was added. Electroporation of K. rhaeticus strains was performed as detailed previously (Florea et al., 2016).
For K. rhaeticus pellicle growth, parental pellicles were first generated by inoculating strains from glycerol stocks in 5 ml HS media and incubated at 30°C for 7 days. In order to maintain plasmids, 340 ng μl−1 chloramphenicol was added. Liquid beneath the parent pellicle was then used, in 1:50 ratio, to inoculate parallel experimental pellicles. Experimental pellicles were harvested after 3–4 days of growth at 30°C. To generate co‐cultured pellicles, the equal volumes of media from beneath the parent pellicle of both strains was removed and mixed, this mixture was then used to inoculate fresh HS media in a deep‐well 24 well plate (Axygen, Union City, CA, USA).
Plasmids used in this study are listed in Table 1. The vector backbones were based on previous BioBrick compatible vectors (for detailed cloning description see (Florea et al., 2016)) and J23104 promoter was adapted using Phusion polymerase (NEB) and PCR mutagenesis (primers listed in Table 2).
Table 1.
Plasmids used in this study
| Plasmids used in this study | Details | Source |
|---|---|---|
| pEmpty (previously named pSEVA331Bb) | E. coli‐K. rhaeticus expression vector; ori‐pBRR1 origin of replication; Chloramphenicol resistant; pSEVA based plasmid with Biobrick modifications | Florea et al. (2016) |
| pLux01 | pSEVA331Bb derivative carrying mRFP gene under the control of pLux and luxR gene under control of pLac promoter | Florea et al. (2016) |
| pReceiver | pLux01 derivative carrying mRFP gene under the control of pLux and luxR gene under control of J23104 promoter | This study |
| pSender | pSEVA331Bb derivative carrying LuxI gene under the control of J23104 promoter | This study |
Table 2.
Primers used in this study
| Name | Sequence |
|---|---|
| VJG_LuxReverse(P) | TCCTGTGTGAAATTGTTATC |
| VJG_J23104SendFrd | TTGACAGCTAGCTCAGTCCTAGGTATTGTGCTAGCAAAGAGGAGAAATACTAGATGA |
| VJG_J23104RecFrd | TTGACAGCTAGCTCAGTCCTAGGTATTGTGCTAGCAAAGAGGAGAAATACTAGATGAA |
| Frd_pSEVA | AGGGCGGCGGATTTGTCC |
Spent media and AHL induction E. coli bioassay
Spent media was made by growing K. rhaeticus strains to late exponential/early stationary phase, cells were then pelleted and removed, and supernatant filtered before use. For E. coli bioassays, spent media or synthetic AHL (Sigma‐Aldrich K3255) was added to equal aliquots of exponentially growing E. coli Receiver cultures in 96‐well plates, with all experimental points in triplicate. These were measured for growth (OD600) and mRFP production (ex., 590 nm; em., 645 nm) over time in a Synergy HT Microplate Reader (BioTek, Winooski, VT, USA). mRFP production rate was calculated as the difference in fluorescence between successive time points normalized by OD600. When using spent media with E. coli, 10 μl of spent HS media was added to 90 μl of E. coli Receiver cultures. This spent media was buffered to pH 7 using Tris pH 8.0 immediately before adding to the E. coli.
Spent media and AHL induction K. rhaeticus bioassay
Komagataeibacter rhaeticus strains were grown to late‐exponential phase in 25 ml tubes, with vigorous shaking and addition of 2% cellulase to prevent clumping and film formation. Cell density was measured by sampling from these cultures into 1 ml cuvettes with a spectrophotometer (Eppendorf Biospectometer) used to determine OD600. For mRFP measurements, 90 μl aliquots of freshly vortexed K. rhaeticus cultures diluted to OD600 1.0 were plated into 96 well plates and 10 μl volumes of either spent media or diluted synthetic AHL were added. After 1 h at 30°C, fluorescent measurements were taken at ex., 590 nm; em., 645 nm subtracting the autofluorescence of the media determined from media blank wells on the same plate. The mRFP measurements were not normalized for cell density, as OD600 values cannot be accurately determined for K. rhaeticus cells growing cellulose in microwell plates.
Fluorescence imaging
Fluorescence images of pellicles were taken with a Fujifilm FLA‐5000 Fluorescent Image Analyser, with excitation at 532 nm, 600 v and Cy5 filter, while Pseudo‐brightfield images were taken with excitation at 473 nm, 400 v and FITC filter. Fluorescence microscopy images were taken on a Nikon Eclipse Ti inverted microscope using a Rolera EM‐C2 camera. K. rhaeticus strains were grown within a ONIX microfluidics plate (B04A; Merck) with flowing HS media at 1 psi and incubated at 30°C. Brightfield and mRFP fluorescence images were taken after 48 h with a 60× oil emulsion objective, exposure time 1.25 s and EM gain of 675. Corresponding brightfield images were contrast enhanced with a phase filter (Ph3). Image processing and compositing were conducted in FIJI (Schindelin et al., 2012).
Pellicle‐to‐pellicles assays
The separate pellicle pairs were placed adjacent to each other within the same petri dish with 5 ml HS media and incubated at 30°C with gentle shaking of 60 RPM. After 24 h the pellicle pairs were removed, and fluorescence imagery was taken. The Boundary detection assay used a larger pellicle, grown for 2 days in a 10 × 10 cm square petri dish with 50 ml HS media. To produce a uniform pellicle at this larger surface area, a 1:10 ration of the liquid from under the first batch of pellicles was used as inoculum. Sender and receiver pellicles were removed from cultures, excess media was removed, and the pellicles were sliced in half along the diagonal. To facilitate diffusion yet prevent sinking, the two pellicles were placed in proximity on soft 0.6% HS agar and incubated at 30°C for 24 h before fluorescence imagery was taken.
Conflict of interest
None declared.
Author contributions
V.J.G., K.T.W. and T.E. conceived and designed the experiments, K.T.W., A.D. and A.G. performed the experiments, K.T.W., V.J.G. and A.D. analysed the experimental data, V.J.G., K.T.W. and T.E. prepared the manuscript.
Supporting information
Fig. S1. Dose response of E. coli Receiver cells induced with increasing concentrations of synthetic AHL.Fig. S2. Growth curve showing that buffered HS media (grey), and spent media sourced from K. rhaeticus strains with the Sender (red) or empty (black) plasmids do not alter E. coli growth.Fig. S3. Flow cytometry data of the induced and uninduced K. rhaeticus Receiver strains.
Acknowledgements
The authors wish to thank Dr Benjamin Reeve and Dr Carlos Bricio‐Garberi for initial help with this project and acknowledge the UK Engineering and Physical Sciences Research Council (EPSRC) for the funding of this work.
Microbial Biotechnology (2019) 12(4), 611–619
Funding Information
UK Engineering and Physical Sciences Research Council (EPSRC).
References
- Basu, S. , Gerchman, Y. , Collins, C.H. , Arnold, F.H. , and Weiss, R. (2005) A synthetic multicellular system for programmed pattern formation. Nature 434: 1130–1134. [DOI] [PubMed] [Google Scholar]
- Boehm, C.R. , Grant, P.K. , and Haseloff, J. (2018) Programmed hierarchical patterning of bacterial populations. Nat Commun 9: 776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brophy, J.A.N. , and Voigt, C.A. (2014) Principles of genetic circuit design. Nat Methods 11: 508–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron, D.E. , Bashor, C.J. , and Collins, J.J. (2014) A brief history of synthetic biology. Nat Rev Microbiol 12: 381–390. [DOI] [PubMed] [Google Scholar]
- Canton, B. , Labno, A. , and Endy, D. (2008) Refinement and standardization of synthetic biological parts and devices. Nat Biotechnol 26: 787–793. [DOI] [PubMed] [Google Scholar]
- Chen, Y. , Su, N. , Zhang, K. , Zhu, S. , Zhao, L. , Fang, F. , Ren, L. , and Guo, Y. (2017) In‐depth analysis of the structure and properties of two varieties of natural luffa sponge fibers. Materials (Basel) 10: 479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churchill, M.E.A. , Sibhatu, H.M. , and Uhlson, C.L. (2011) Defining the structure and function of acyl‐homoserine lactone autoinducers. Methods Mol Biol 692: 159–171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Florea, M. , Hagemann, H. , Santosa, G. , Abbott, J. , Micklem, C.N. , Spencer‐Milnes, X. , de Arroyo Garcia, L. , Paschou, D. , Lazenbatt, C. , Kong, D. , Chughtai, H. , Jensen, K. , Freemont, P.S. , Kitney, R. , Reeve, B. , and Ellis, T. (2016) Engineering control of bacterial cellulose production using a genetic toolkit and a new cellulose‐producing strain. Proc Natl Acad Sci USA 113: E3431–E3440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang, W.D. , Hwang, J.H. , Kim, H.U. , Ryu, J.Y. , and Lee, S.Y. (2017) Bacterial cellulose as an example product for sustainable production and consumption. Microb Biotechnol 10: 1181–1185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karig, D. , Martini, K.M. , Lu, T. , DeLateur, N.A. , Goldenfeld, N. , and Weiss, R. (2018) Stochastic Turing patterns in a synthetic bacterial population. Proc Natl Acad Sci USA 115: 6572–6577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, K.Y. , Buldum, G. , Mantalaris, A. , and Bismarck, A. (2014) More than meets the eye in bacterial cellulose: biosynthesis, bioprocessing, and applications in advanced fiber composites. Macromol Biosci 14: 10–32. [DOI] [PubMed] [Google Scholar]
- Liu, C. , Fu, X. , Liu, L. , Ren, X. , Chau, C.K.L. , Li, S. , Xiang, L. , Zeng, H. , Chen, G. , Tang, L.H. , Lenz, P. , Cui, X. , Huang, W. , Hwa, T. , and Huang, J.D. (2011) Sequential establishment of stripe patterns in an expanding cell population. Science 334: 238–241. [DOI] [PubMed] [Google Scholar]
- Liu, M. , Liu, L. , Jia, S. , Li, S. , Zou, Y. , and Zhong, C. (2018) Complete genome analysis of Gluconacetobacter xylinus CGMCC 2955 for elucidating bacterial cellulose biosynthesis and metabolic regulation. Sci Rep 8: 6266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen, P.Q. , Botyanszki, Z. , Tay, P.K.R. , and Joshi, N.S. (2014) Programmable biofilm‐based materials from engineered curli nanofibres. Nat Commun 5: 4945. [DOI] [PubMed] [Google Scholar]
- Nguyen, P.Q. , Courchesne, N.M.D. , Duraj‐Thatte, A. , Praveschotinunt, P. , and Joshi, N.S. (2018) Engineered living materials: prospects and challenges for using biological systems to direct the assembly of smart materials. Adv Mater 30: 1704847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, J.P. , Choi, M.J. , Kim, S.H. , Lee, S.H. , and Lee, H. (2014) Preparation of sticky Escherichia coli through surface display of an adhesive catecholamine moiety. Appl Environ Microbiol 80: 43–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Römling, U. , and Galperin, M.Y. (2015) Bacterial cellulose biosynthesis: diversity of operons, subunits, products, and functions. Trends Microbiol 23: 545–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schindelin, J. , Arganda‐Carreras, I. , Frise, E. , Kaynig, V. , Longair, M. , Pietzsch, T. , Preibisch, S. , Rueden, C. , Saalfeld, S. , Schmid, B. , Tinevez, J.Y. , White, D.J. , Hartenstein, V. , Eliceiri, K. , Tomancak, P. , and Cardona, A. (2012) Fiji: an open‐source platform for biological‐image analysis. Nat Methods 9: 676–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholes, N.S. , and Isalan, M. (2017) A three‐step framework for programming pattern formation. Curr Opin Chem Biol 40: 1–7. [DOI] [PubMed] [Google Scholar]
- Scott, S.R. , and Hasty, J. (2016) Quorum sensing communication modules for microbial consortia. ACS Synth Biol 5: 969–977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turing, A.M. (1952) The chemical basis of morphogenesis. Philos Trans R Soc B Biol Sci 237: 37–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waters, C.M. , and Bassler, B.L. (2005) Quorum sensing: cell‐to‐cell communication in bacteria. Annu Rev Cell Dev Biol 21: 319–346. [DOI] [PubMed] [Google Scholar]
- Wolpert, L. (1969) Positional information and the spatial pattern of cellular differentiation. J Theor Biol 25: 1–47. [DOI] [PubMed] [Google Scholar]
- Youssefian, S. , and Rahbar, N. (2015) Molecular origin of strength and stiffness in bamboo fibrils. Sci Rep 5: 11116. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Dose response of E. coli Receiver cells induced with increasing concentrations of synthetic AHL.Fig. S2. Growth curve showing that buffered HS media (grey), and spent media sourced from K. rhaeticus strains with the Sender (red) or empty (black) plasmids do not alter E. coli growth.Fig. S3. Flow cytometry data of the induced and uninduced K. rhaeticus Receiver strains.