

Abstract
Introduction: Cancer stem cells (CSCs) are the main source of tumor resistance and recurrence. At present, the main treatment for patients with advanced or metastatic bladder cancer (BCa) is cisplatin-based combination chemotherapy. However, CSCs are not sensitive to DNA-damaging drugs due to their enhanced DNA damage response (DDR) activity.
Materials and methods: Bladder cancer stem cell-like cells (BCSLCs) were obtained by treating UMUC3 cells with cisplatin. The characteristics of the BCSLCs were identified by qPCR, flow cytometry, scratch wound-healing assays, transwell assays, tumorigenic ability experiments, Edu assays and Western blot assays in vivo. After BCSLCs were treated with norcantharidin (NCTD), the expression of Cdc6 and activation of the ATR-Chk1 pathway were detected by Western blotting. A subcutaneous tumor model in nude mice was successfully established to assess the anti-tumor efficacy of NCTD and cisplatin either alone or in combination in vivo. The tumor tissues were detected by immunohistochemistry.
Results: The derived BCSLCs showed higher expression of stemness markers, increased invasiveness, improved resistance to multiple chemotherapeutics, and higher tumorigenic capacity in vivo. The protein expression level of chromatin-binding Cdc6 was increased in BCSLCs; however, NCTD decreased the level of chromatin-binding Cdc6 and inhibited the activation of the ATR-Chk1 pathway, which ultimately led to reduction in DDR activity in BCSLCs. NCTD enhanced the killing effect of cisplatin on BCSLCs in vitro and vivo. NCTD combined with cisplatin enhanced cisplatin-induced DNA damage in BCSLCs.
Conclusion: Long-term cisplatin treatment can enrich BCSLCs. However, NCTD enhanced the killing effect of cisplatin on BCSLCs in vitro and vivo. The mechanism is inhibiting the DDR activity by reducing the expression of chromatin-binding Cdc6.
Keywords: bladder cancer stem-like cells, norcantharidin, DDR, Cdc6, cisplatin
Introduction
Bladder cancer (BCa) is one of the most common urological malignancies threatening human health, and it ranks as the 9th most frequently diagnosed cancer and the 13th cancer in terms of associated deaths worldwide.1 The most effective treatment for muscle-invasive BCa is radical total cystectomy. However, approximately 50% of patients experience recurrence and metastasis within 2 years after surgery and eventually die.2 The main treatment for patients with advanced or metastatic BCa is cisplatin-based combination chemotherapy, such as the GC (gemcitabine and cisplatin) and the MVAC (methotrexate, vinblastine, adriamycin and cisplatin) regimens.3 Traditional chemotherapeutic medicine can kill only rapidly proliferating tumor cells. However, the cancer stem cells (CSCs) in the tumor are not sensitive to traditional chemotherapy due to an enhanced DNA damage response (DDR). therefore, CSCs resist traditional chemotherapy, which results in therapy failure.4–8 In recent years, research on cancer biomarkers and/or therapeutic targets for high-grade bladder cancer has made encouraging progress in addressing treatment resistance issues.9,10 However, there are few reports on the contribution of the DDR to drug resistance in bladder cancer stem cells.
The DDR is a signaling network involving multiple signaling pathways, including the detection point pathway, DNA repair, transcriptional regulation and apoptosis. The effectiveness of DDR depends on the coordination of ataxia telangiectasia and Rad3-related kinase (ATR) involvement in the detection point pathway and various DNA repair mechanisms.11 ATR is a protein kinase that is recruited to the site of DNA damage and activated in the context of DNA damage; it plays an important role in the activation of cell cycle checkpoints.11–13
Cdc6 is a newly discovered anti-tumor target, and the main function of Cdc6 is to assemble a pre-replication complex (pre-RC) at the origin to initiate DNA replication in the G1 phase.14 Studies have found that Cdc6 has other functions: Cdc6 serves as a receptor for the ATR-ATRIP complex to bind to chromatin in yeast.15 Cdc6 interaction with ATR regulates replication-checkpoint activation in the cells of the African clawed frog, and Cdc6 silencing impairs ATR-dependent checkpoint activation.16 In gallbladder and cervical carcinoma, Cdc6 is involved in the activation of the ATR signal.17,18
Norcantharidin (NCTD) is a derivative of cantharidin and an anti-tumor drug independently developed in China.19,20 Numerous studies have shown that NCTD can inhibit DNA replication and induce apoptosis of tumor cells.19–21 Norcantharidin is currently used in clinical anti-cancer treatment.20,22–25 In our previous studies, NCTD had anti-tumor effects on various types of cancers, including bladder cancer cells. We also found that NCTD can degrade the Cdc6 protein in cancer cells.26,27 There is a high probability that NCTD affects the DDR of CSCs by inhibiting Cdc6 and enhancing the sensitivity to cisplatin.
In this study we conducted experiments to enrich and evaluate bladder cancer stem-like cells (BCSLCs) with stemness characteristics and resistance to multiple chemotherapeutic drugs. Then, we explored the effect of norcantharidin on the Cdc6 protein and ATR activation, and verified that norcantharidin can enhance the anti-tumor effect of cisplatin in vitro and vivo by inhibiting DDR activity.
Materials and methods
Cell lines
Cells from human bladder cancer cells line UMUC3 (obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA)) were cultured through the standard procedure in RPMI-1640 (Gibco, Grand Island, NY, USA) with 10% fetal bovine serum (Gibco), 1% penicillin and 1% streptomycin (Gibco) under standard conditions (5% CO2 and 95% atmosphere, 37 °C). The drug-resistant cell line UMUC3R was obtained by administering 6.66 μM cisplatin (Selleck, Houston, TX, USA) to UMUC3 cells every other day for 90 days. Then, the cells were further treated with continuous application of 13.32 μM cisplatin for 14 days to obtain a cell subline with high expression of stemness makers. The cells in this subline are designated as bladder cancer stem cell-like cells (BCSLCs).
Identified characteristic of the BCSLCs
Expression of stemness markers
Quantitative real-time polymerase chain reaction (qPCR) was used to evaluate the expression of CD34, CD44, CD133, OCT4, ABCG2, and NANOG. Total RNA was isolated from the cells using TRIzol reagents (Takara, Japan). cDNA was synthesized using the Prime Script™ RT Reagent Kit with genomic DNA Eraser (Takara). qPCR was performed using SYBR® Premix Ex Taq™ (Takara)on a 7500 real-time PCR system (AB, Biosystems, Singapore) to detect mRNA expression. Beta-actin was used as a negative control, and normalization and fold changes were calculated using the ΔΔCt method. UMUC3 cells and BCSLCs were stained with PE-labeled anti-CD133 (eBioscience, San Diego, CA, USA). The CD133+ cells were detected by flow cytometry (FCM; Becton Dickinson, San Jose, CA)
Migration and invasion assays
Both UMUC3 cells and BCSLCs were cultured in RPMI-1640 with 10% fetal bovine serum (FBS) under standard conditions for 3 days to release them from cisplatin pressure. Then, UMUC3 cells and BCSLCs were seeded in 6-well plates with serum-free RPMI-1640 medium. Cell migration was assessed by a scratch wound-healing assay. UMUC3 cells and BCSLCs were seeded in 24-well plates with serum-free RPMI-1640 medium. Cell migration and invasion were assessed by a transwell assay.
Tumorigenic ability
Nude mice were purchased from the experimental animal center at Southern Medical University (Guangzhou, China). The protocols of animal studies were approved by the Southern Medical University Experimental Animal Ethics Committee (ethics number: L2018118). All mice received humane care in accordance with the National Institutes of Health animal use guidelines.
Nude mice (4 weeks) were maintained and treated under specific pathogen-free conditions. The tumorigenic ability of UMUC3 cells and BCSLCs was compared after subcutaneous injection of different concentrations of cells into nude mice.
Analysis of cell viability (chemotherapy resistance)
The UMUC3 cells and BCSLCs were seeded in 96-well plates and incubated for 24 hr. Next, the cells were treated with different concentrations of cisplatin (Sigma), paclitaxel (Sigma), and gemcitabine (Selleck) for another 24 hr. Cell viability was quantified by using Cell Counting Kit-8 (CCK-8) reagent (Dojindo, Kumamoto, Japan).
DNA replication and expression of Cdc6 in BCSLCs
Edu assays
The UMUC3 cells and BCSLCs were seeded in 6-well plates, and DNA replication was measured using a Cell-Light™ Edu Apollo® 567 in vitro imaging kit (RiboBio, Guangzhou, China) according to the instructions for the kit. The cells were observed under an inverted microscope (Nikon TE2000-S, Tokyo, Japan) and photographed.
Chromatin binding assays
The UMUC3 cells and BCSLCs were treated with the same concentration cisplatin (19.98 μM). After 48 hr, the cells were harvested and resuspended in tubes with EB buffer (100 mM KCl, 50 mM HEPES-KOH pH 7.5, 2.5 mM MgCl2, 50 mM Na4P2O7, 0.1 mM NaVO3, 0.5% Triton X-100) with protease inhibitors and then incubated for 10 min on ice. The tubes were flicked every 2–3 min to mix the solution during incubation. Subsequently, 30% ice-cold sucrose with protease inhibitors was added to the bottom of the tubes. The tubes were spun at 15k rpm for10 min at 4 °C, and the supernatants were transferred to new tubes. The pellets were washed with EB buffer and vibrated briefly for resuspension, followed by spinning in a microfuge at 15k rpm for 5 min at 4 °C. The supernatants from the two steps were combined (the non-chromosomal fraction). The pellets (the chromatin-binding fraction) were resuspended in EB buffer and analyzed by Western blotting.
Western blot analysis
After the protein concentration was measured by using a BCA protein assay kit, equal amounts of protein from the samples were separated by electrophoresis on a 10% SDS-PAGE gel and transferred onto polyvinylidene fluoride membranes. After blocking with 5% nonfat milk solution for 1 hr, the membrane was incubated at 4 °C with a primary monoclonal antibody against Cdc6 (CST, MA, USA), beta-actin (CST) or histone-H3 (CST) overnight. After washing with TBST, the membrane was incubated with secondary antibodies against goat HRP-conjugated IgG at room temperature for 1 hr. The membrane was washed, and the proteins were detected by an ECL system (Millipore, Billerica, MA, USA). Western blot bands were analyzed by imageJ.
The effect of NCTD on BCSLCs in vitro
The effect of NCTD on the expression of Cdc6-c in BCSLCs
BCSLCs were treated with norcantharidin (NCTD) at different concentrations (0 μM, 50 μM, 100 μM and 200 μM) for 48 hr. Chromatin-binding proteins were extracted, and the protein expression of Cdc6-C was analyzed by Western blotting. Histone-H3 was used as an internal control.
The effect of NCTD on the expression of ATR-C, p-Chk1 and p-Cdc25c in BCSLCS and UMUC3 cells
NCTD (50 μM) was continuously applied to BCSLCs and UMUC3 cells for 48 hr. The chromatin-binding protein was extracted, and the expression of ATR-C was analyzed by Western blotting (6% SDS-PAGE). Histone-H3 was used as an internal control. Total protein was extracted by using RIPA Lysis Buffer (Beyotime, Shanghai, China). The expression of phosphorylated chk1 (p-Chk1), p-Cdc25C was analyzed by Western blotting (10% SDS-PAGE), and beta-actin was used as an internal control.
The cell viability was analyzed by the cck-8 assay
BCSLCs were divided into four groups: NCTD+ cisplatin, NCTD, cisplatin, and control. The corresponding doses of NCTD (50 μM) and/or Cisplatin (19.98 μM) were added to each group. After 48 hr, the cell viability was analyzed by the cck-8 assay.
Effect of NCTD combined with cisplatin on BCSLCs in vivo
Subcutaneous tumor formation was established by injecting 1×106 BCSLCs subcutaneously into the right flank of nude mice. The nude mice with subcutaneous tumors were divided into four groups (5 mice in each group, with an average body weight of approximately 19.22±1.16 g): cisplatin, NCTD, and NCTD + cisplatin(combo) and Control. When the tumor diameter grew to 6 mm, drugs were administered every 3 days as follows: cisplatin group: 0.2 ml cisplatin solution (0.5 mg/kg), NCTD group: 0.2 ml norcantharidin solution (20 mg/kg), combo group: 0.1 ml cisplatin solution (0.5 mg/kg) combined with 0.1 ml norcantharidin solution (20 mg/kg), and control group: 0.2 ml of normal saline (0.9% NS). Mice with subcutaneous tumor undergoing drug treatment were observed for 21 days, and the volume of the tumor was measured every 3 days. After 21 days, the tumor tissues were extracted for immunohistochemical determination with Cdc6, ATR and γ-H2AX (Abcam) antibodies.
Statistics
The experiments in vivo were repeated 3 times. All results are presented as the means ± SD. Statistical analysis of qPCR, FACS, Edu, scratch wound-healing, transwell, Western blot data was performed using an independent sample t-test. Data cck-8 assay for cell viability and immunohistochemistry detection were analyzed using one-way ANOVA. The difference in tumor volume between the two groups was compared by using repeated measures ANOVA. All data were analyzed using SPSS (version 22.0), and P<0.05 was considered indicative of statistical significance.
Results
Characterization of BCSLCs
Compared with that in UMUC3 cells, the expression of CD34, CD133, ABCG2, OCT4 and NANOG in BCSLCs was significantly increased (Figure 1A and B). The rate of CD133+ cells in BCSLCs was significantly higher than that in UMUC3 cells (Figure 1C and D). The cell migration capability of BCSLCs was significantly greater than that of UMUC3 cells, according to the scratch wound-healing assay (Figure 1E and F) and transwell migration assay (Figure 1G and H). The cell invasion capability of BCSLCs was significantly greater than that of UMUC3 cells, according to the transwell invasion assay (Figure 1G and H). Different concentrations of UMUC3 cells and BCSLCs were injected subcutaneously into nude mice, which were then observed for 4 weeks. The results showed that the tumorigenic ability of BCSLCs was significantly higher than that of UMUC3 cells; BCSLCs can form subcutaneous tumors after injection of 5×104 tumor cells, while UMUC3 cells require 1×106 tumor cells (Table 1). Injection of 5×106 BCSLCs and UMUC3 cells can induce the formation of subcutaneous tumors in all mice. However, the subcutaneous tumors formed by the BCSLCs were significantly larger than those formed by the UMUC3 cells, and the difference was statistically significant (Figure 1I and J, Table 2). BCSLCs show greater chemotherapy resistance than UMUC3 (parental) cells (Table 3).
Figure 1.

Characterization of BCSLCs. (A, B) Expression of CD34, CD44, ABCG2, CD133, OCT4 and NANOG in UMUC3 cells and BCSLCs according to qPCR assays. (C) Flow cytometry was used to detect CD133+ cells in UMUC3 cells and BCSLCs. (D) Proportion of CD133+ cells (%) is presented. (E) Cell gaps were photographed at 0, 24, 48, and 72 hr after scratching. (F) Distance between cells in the scratch zone (% of basal) is shown. (G) Cell migration and invasion as determined by the transwell assay were photographed. (H) Cells per field were counted. (I) Subcutaneous tumor formation was established under the skin on the side of the back in nude mice with 5×106 cells UMUC3 cells and BCSLCs after 4 weeks (Left/UMUC3 cells, Right/BCSLCs). (J) The tumors were extracted from the mice after day 28 (Upper/BCSLCs, Lower/UMUC3 cells). There were significant differences in size. (*P<0.05).
Abbreviation: BCSLCs, bladder cancer stem-like cells.
Table 1.
Tumor formation rate from cells injected at different concentrations
| Cell Type | Number of Cells | |||
|---|---|---|---|---|
| 5×104 | 1×105 | 1×106 | 5×106 | |
| BCSLCs | 1/5 | 3/5 | 5/5 | 5/5 |
| UMUC3 | 0/5 | 0/5 | 2/5 | 5/5 |
Abbreviation: BCSLCs, bladder cancer stem-like cells.
Table 2.
Volume of subcutaneous tumors in the UMUC3 and BCSLCs groups of mice (mean ± SD) (n=5) (*P<0.05)
| Cell Type | Volume (mm3) |
|---|---|
| BCSLCs | 3354.00±83.35* |
| UMUC3 | 178.63±19.45 |
Abbreviation: BCSLCs, bladder cancer stem-like cells.
Table 3.
The half maximal inhibitory concentration (IC50) of chemotherapy drugs to UMUC3 and BCSLCs (mean ± SD) (n=3) (*P<0.05)
| Cell Type | Cisplatin(μM) | Paclitaxel (nM) | NCTD(μM) | Gemcitabine(nM) |
|---|---|---|---|---|
| BCSLCs | 35.06±7.33* | 3354±83.35* | 94.72±7.08* | 6287.67±162.19* |
| UMUC3 | 12.62±3.53 | 178.63±19.45 | 25.17±1.97 | 194.67±9. 65 |
Abbreviations: BCSLCs, bladder cancer stem-like cells; NCTD, norcantharidin.
BCSLCs are relatively quiescent regarding DNA replication, but show high levels of initiation protein Cdc6 binding onto chromatin
To determine the proliferative status of BCSLCs, we conducted Edu incorporation assays. The proportion of Edu-positive cells in BCSLCs population was much lower than that in UMUC3 population (Figure 2A and B). These results suggest that BCSLCs are in a stationary phase and that DNA replication is significantly inhibited. However, as an initiation protein of DNA replication, Cdc6 shows considerable expression and binding to chromatin in BCSLCs. Moreover, the amount of chromatin-binding Cdc6 (Cdc6-C) in BCSLCs was higher than that in UMUC3 cells (Figure 2C and D).
Figure 2.

Edu assays and protein expression of Cdc6 in BCSLCs. (A) Edu assays were conducted to evaluate DNA synthesis (proliferation ability). (B) Proportion of Edu+ cells (%). (C) Cdc6 protein levels were analyzed by Western blotting. Cdc6-S: non-chromatin binding Cdc6; Cdc6-C: chromatin-binding Cdc6. (D) Relative OD of Western blot bands is presented. (*P<0.05).
Abbreviations: BCSLCs, bladder cancer stem-like cells; Cdc6-S, non-chromatin binding Cdc6; Cdc6-C, chromatin-binding Cdc6.
The effect of NCTD on BCSLCs in vitro: NCTD inhibited the expression of Cdc6-c, inhibited activation of the ATR pathway, inhibited DDR, and enhanced the killing effect of cisplatin on BCSLCs in vitro
Chromatin-binding proteins were collected after BCSLCs were treated with different concentrations of NCTD. The result showed that NCTD inhibited the expression of chromatin-binding Cdc6 (Cdc6-C) in BCSLCs, and the inhibition was enhanced with increasing NCTD concentrations (Figure 3A and B). Next, chromatin protein and total protein were collected after BCSLCs and UMUC3 cells were treated with or without NCTD (50 μM). The expression of ATR-C, phosphorylated Chk1 (p-Chk1) and phosphorylated Cdc25C (p-Cdc25C) was detected by Western blotting. The results showed the following: 1) The expression of ATR-C, p-Chk1, and p-Cdc25C in BCSLCs was significantly higher than that in UMUC3 cells. 2) The ATR pathway was significantly activated in BCSLCs compared with UMUC3 cells. 3)The expression of ATR-C, p-Chk1 and p-Cdc25C in UMUC3 cells and BCSLCs treated with NCTD was significantly increased, and NCTD inhibited the activation of the ATR pathway in UMUC3 cells and BCSLCs (Figure 3C–F). The cck-8 assay showed that the relative cell viability (mean) of the cisplatin group was 0.62, while that of the NCTD group was 0.59, and that of the combo group was 0.32. NCTD enhanced the killing effect of cisplatin on BCSLCs (Figure 3G).
Figure 3.
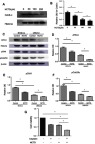
The effect of NCTD on BCSLCs in vitro. (A) The protein expression of Cdc6-C in BCSLCs after NCTD treatment for 48 hr (0 μM, 50 μM, 100 μM and 200 μM) was detected by Western blotting. (B) The relative OD for Cdc6-C was determined from the Western blot band. (C) The expression of ATR-C, pChk1, pCdc25C in BCSLCs and UMUC3 cells after treatment with or without NCTD (50 μM) was detected by Western blotting. (D–F) The relative OD data for ATR-C, pChk1, pCdc25C was determined from the Western blot band. (G) Relative cell viability of different groups is shown. (*P<0.05).
Abbreviations: BCSLCs, bladder cancer stem-like cells; Cdc6-C, chromatin-binding Cdc6; NCTD, norcantharidin; ATR-C, Chromatin-binding ATR; pChk1, phosphorylated check point kinase 1; pCdc25C, phosphorylated Cdc25C.
NCTD enhanced killing effect of cisplatin on BCSLCs in vivo
The nude mice with subcutaneous tumors were randomly divided into four groups, treated as planned with intraperitoneal injection, and the volume of the tumor was measured every 3 days. The observation lasted 21 days. The results showed that 1) NCTD or cisplatin alone slowed tumor growth, but did not effectively reduce tumor volume (the tumor volume of the NCTD group decreased significantly compared with that of the control group after day 9, and tumor growth among mice in the cisplatin group was slowed to a certain extent after day 18, but the tumor volume of both group still increased gradually). 2) The combo group showed the strongest inhibitory effect on tumor growth. The tumor volume of the combo group was significantly smaller than that of the other three groups (P<0.05), and the tumor volume decreased on the 18th day, compared with the 15th day, under continuous treatment, indicating that NCTD reversed the resistance of the tumor to cisplatin, enhanced the anti-tumor effect of cisplatin and reduced the tumor volume (Figure 4A and B, Table 4). Then, the tumor tissues were obtained, embedded and sectioned for immunohistochemical detection. Immunohistochemistry assays showed that 1) Cdc6 expression in the NCTD group was significantly weaker than it was in the control group (P<0.05). Cdc6 expression in the combo group was rarely detected. Cdc6 expression in the cisplatin group was weaker than it was in the control group (Figure 4C and D). 2) Expression of ATR in the NCTD group and the control group was not obvious, but it was significantly increased in the cell nucleus of the Cisplatin group cells. Cisplatin activated the ATR pathway, while the expression of ATR in the combo group was decreased, indicating that NCTD inhibited the activation of the ATR pathway (Figure 4C and E). 3) γ-H2AX was expressed to the greatest degree in the combo group, indicating that NCTD enhanced the DNA damaging effect of cisplatin (Figure 4C and F). This finding confirmed that NCTD inhibited the protein expression of Cdc6 and the ATR pathway in tumor tissues, and it reduced the DDR activity and enhanced the DNA damage caused by cisplatin.
Figure 4.

The effect of NCTD in BCSLCs in vivo. (A) Tumor size of each group on day 21 is shown. (B) Growth curve of subcutaneous tumors is presented for each group. (C) Representative graph of Immunohistochemistry shows Cdc6, ATR and γ-H2AX (brown represents positive expression). (D) Histogram of ATR immunohistochemical score is displayed. (E) Histogram of Cdc6 immunohistochemical score is presented. (F) Histogram of γ-H2AX immunohistochemical score is shown. (*P<0.05).
Abbreviations: Cdc6-C, chromatin-binding Cdc6; NCTD, norcantharidin; Cdc6, cell division cycle 6; ATR, ataxia telangiectasia mutated and RAD3-related.
Table 4.
Volume of subcutaneous tumors in each group of mice (mean ± SD) (n=5)
| Groups | Control (mm3) | Cisplatin (mm3) | NCTD (mm3) | Combo (mm3) |
|---|---|---|---|---|
| Day 0 | 87.06±8.23 | 86.36±7.46 | 86.04±6.58 | 86.61±6.84 |
| Day 3 | 138.19±20.98 | 121.47±12.29 | 126.59±11.43 | 120.52±12.49 |
| Day 6 | 237.62±40.56 | 224.57±29.44 | 195.65±25.18 | 156.91±25.07 |
| Day 9 | 587.07±69.82 | 553.97±54.38 | 370.65±46.75 | 266.84±31.91 |
| Day 12 | 918.33±89.55 | 879.11±72.39 | 679.06±70.41 | 408.67±43.75 |
| Day 15 | 1389.18±128.8 | 1290.78±109.26 | 889.91±89.76 | 530.89±58.27 |
| Day 18 | 1808.87±133.67 | 1491.26±105.61 | 1131.43±70.78 | 443.42±39.47 |
| Day 21 | 2572.22±157.47 | 1895.34±79.43 | 1370.67±56.52 | 373.62±25.65 |
Abbreviation: NCTD, norcantharidin.
Discussion
CSCs comprise a subpopulation of cancer cells capable of self-renewal and differentiation into mature tumor cells.28 they play an important role in tumor growth and a decisive role in maintaining malignant proliferation, invasion, metastasis, recurrence and drug resistance of tumors.29 The recurrence of solid tumors may be due to the inability of traditional chemotherapy and radiotherapy to eliminate CSCs. Eliminating CSCs is a key issue in eradicating tumors.30 Researchers have successfully isolated and identified CSCs from various tumors.30–34 Our previous study also reported the successful separation and identification of CSCs from MB49 bladder cancer cell line.35 The tumorigenic ability was obviously greater than it was in the MB49 cell line. and chemotherapy resistance of CSCs is greater than that of the MB49 cells. In this study, we found that bladder cancer stem-like cells (BCSLCs) can also be enriched in cancer cell populations by cisplatin screening and show multiple chemotherapeutic drug resistance.
The enhanced DDR response is an important mechanism for the drug resistance of cancer stem cells.6 ATR plays two main roles in the DDR process: First, ATR binds to DNA damage sites to make time for DNA repair by activating the checkpoint kinase 1 (Chk1) checkpoint pathway and continuing to phosphorylate the Cdc25 protein, leading to multistage cell cycle arrest. At the same time, ATR interacts with other related proteins to bind to the damaged area of DNA and participate in DNA repair.36,37 DNA-damaging drugs such as cisplatin can activate the ATR pathway and stimulate DNA repair in cells. Abnormal regulation of DNA repair is associated with chemotherapy resistance.38–40
Therefore, ATR plays an important regulatory role in the activation of the detection point pathway during DNA damage. In our study, we found that the expression of chromatin-binding ATR (ATR-C), pChk1 and pCdc25C in BCSLCs after cisplatin treatment was significantly higher than that in UMUC3 cells. It is highly possible that the chemotherapy resistance of BCSLCs may be related to excessive activation of the ATR-Chk1 pathway.
Cdc6 overexpression has been detected in a variety of cancer types, and high levels of Cdc6 are not merely the result of cancer proliferation. Cdc6 itself has the ability to promote malignant cell transformation and is associated with poor prognosis in patients with cancer.41,42 In this study, we found that DNA replication was reduced in BCSLCs, however, the protein expression of chromatin-binding Cdc6 was higher. A significant proportion of Cdc6 remains in the nucleus and binds to chromatin, which is positively associated with the ATR pathway activated in BCSLCs. This observation suggests that these chromatin-binding Cdc6 proteins may be involved in the chemotherapy resistance of BCSLCs and that the ATR-related detection point pathway is regulated by the interaction between Cdc6 and ATR.
When we treated BCSLCs with norcantharidin, the protein level of chromatin- bound Cdc6 decreased, activation of the ATR-Chk1 pathway was reduced and the sensitivity of cells to cisplatin was enhanced both in vivo and in vitro, DDR was reduced and DNA damage was increased. Norcantharidin enhanced the killing effect of cisplatin on BCSLCs.
Conclusion
Bladder cancer stem-like cells (BCSLCs) were obtained by enrichment of UMUC3 cells with the long-term cisplatin treatment and evaluated in the current study. Compared with UMUC3 cells, BCSLCs have more malignant characteristics (stronger proliferation, migration and invasion, MDR, and tumorigenic ability). The expression level of chromatin-binding Cdc6 in BCSLCs was higher than that in UMUC3 cells after cisplatin treatment.
NCTD enhanced the killing effect of cisplatin on BCSLCs in vitro and vivo. The mechanism is based on the ability of NCTD to reduce chromatin-binding Cdc6 expression, inhibit the activation of the ATR-Chk1 pathway, and inhibit DDR activity. The study provided an experimental basis for the clinical application of NCTD.
Acknowledgments
This project was funded by the Natural Science Foundation of Guangdong Province (Grant No: 2015A030313289) (Wanlong Tan) and the National Natural Science Foundation of China (Grant No: 81672915) (Jinlong Li).
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Antoni S, Ferlay J, Soerjomataram I et al. Bladder cancer incidence and mortality: a global overview and recent trends. Eur Urol. 2017;71(1):96–108. doi: 10.1016/j.eururo.2016.06.010 [DOI] [PubMed] [Google Scholar]
- 2.Hussain SA, James ND. The systemic treatment of advanced and metastatic bladder cancer. Lancet Oncol. 2003;4(8):489–497. doi: 10.1016/S1470-2045(03)01168-9 [DOI] [PubMed] [Google Scholar]
- 3.Vaishampayan U. Systemic therapy of advanced urothelial cancer. Curr Treat Options Oncol. 2009;10(3–4):256–266. doi: 10.1007/s11864-009-0101-9 [DOI] [PubMed] [Google Scholar]
- 4.Blanpain C, Mohrin M, Sotiropoulou PA, Passegue E. DNA-damage response in tissue-specific and cancer stem cells. Cell Stem Cell. 2011;8(1):16–29. doi: 10.1016/j.stem.2010.12.012 [DOI] [PubMed] [Google Scholar]
- 5.Colak S, Medema JP. Cancer stem cells–important players in tumor therapy resistance. Febs J. 2014;281(21):4779–4791. doi: 10.1111/febs.12758 [DOI] [PubMed] [Google Scholar]
- 6.Sugihara E, Saya H. Complexity of cancer stem cells. Int J Cancer. 2013;132(6):1249–1259. doi: 10.1002/ijc.27961 [DOI] [PubMed] [Google Scholar]
- 7.Maugeri-Sacca M, Bartucci M, De Maria R. DNA damage repair pathways in cancer stem cells. Mol Cancer Ther. 2012;11(8):1627–1636. doi: 10.1158/1535-7163.MCT-11-0824-T [DOI] [PubMed] [Google Scholar]
- 8.Yu WK, Wang Z, Fong CC, et al. Chemoresistant lung cancer stem cells display high DNA repair capability to remove cisplatin-induced DNA damage. Br J Pharmacol. 2017;174(4):302–313. doi: 10.1111/bph.13690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vantaku V, Donepudi SR, Ambati CR, et al. Expression of ganglioside GD2, reprogram the lipid metabolism and EMT phenotype in bladder cancer. Oncotarget. 2017;8(56):95620–95631. doi: 10.18632/oncotarget.21038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vantaku V, Dong J, Ambati CR, et al. Multi-omics integration analysis robustly predicts high-grade patient survival and identifies CPT1B effect on fatty acid metabolism in Bladder Cancer. Clin Cancer Res. 2019. clincanres.1515.2018. doi: 10.1158/1078-0432.CCR-18-1515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cimprich KA, Cortez D. ATR: an essential regulator of genome integrity. Nat Rev Mol Cell Bio. 2008;9(8):616–627. doi: 10.1038/nrm2450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Keith CT, Schreiber SL. PIK-related kinases: DNA repair, recombination, and cell cycle checkpoints. Science. 1995;270(5233):50–51. [DOI] [PubMed] [Google Scholar]
- 13.Engelman JA, Luo J, Cantley LC. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet. 2006;7(8):606–619. doi: 10.1038/nrg1749 [DOI] [PubMed] [Google Scholar]
- 14.Evrin C, Fernandez-Cid A, Riera A, et al. The ORC/Cdc6/MCM2-7 complex facilitates MCM2-7 dimerization during prereplicative complex formation. Nucleic Acids Res. 2014;42(4):2257–2269. doi: 10.1093/nar/gkt1148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hermand D, Nurse P. Cdc18 enforces long-term maintenance of the S phase checkpoint by anchoring the Rad3-Rad26 complex to chromatin. Mol Cell. 2007;26(4):553–563. doi: 10.1016/j.molcel.2007.04.010 [DOI] [PubMed] [Google Scholar]
- 16.Yoshida K, Sugimoto N, Iwahori S, et al. CDC6 interaction with ATR regulates activation of a replication checkpoint in higher eukaryotic cells. J Cell Sci. 2010;123(Pt 2):225–235. doi: 10.1242/jcs.058693 [DOI] [PubMed] [Google Scholar]
- 17.Wang WX, Zhang WJ, Peng ZL, Yang KX. [Expression and clinical significance of CDC6 and hMSH2 in cervical carcinoma]. Sichuan Da Xue Xue Bao Yi Xue Ban. 2009;40(5):857–860. [PubMed] [Google Scholar]
- 18.Shu GS, Yang LP, Yang ZL, Jiang S. Expression of CDC6 and GDF-9 and their clinicopathological significances in benign and malignant lesions of the gallbladder. Cancer Biomark. 2012;11(2–3):107–114. doi: 10.3233/CBM-2012-0267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Puerto GC, Vargas ML, Kouznetsov VV. Cantharidin-based small molecules as potential therapeutic agents. Chem Biol Drug Des. 2013;82(5):477–499. doi: 10.1111/cbdd.12180 [DOI] [PubMed] [Google Scholar]
- 20.Deng L, Tang S. Norcantharidin analogues: a patent review (2006-2010). Expert Opin Ther Pat. 2011;21(11):1743–1753. doi: 10.1517/13543776.2011.539205 [DOI] [PubMed] [Google Scholar]
- 21.Deng LP, Dong J, Cai H, Wang W. Cantharidin as an antitumor agent: a retrospective review. Curr Med Chem. 2013;20(2):159–166. doi: 10.2174/092986713804806711 [DOI] [PubMed] [Google Scholar]
- 22.Zhang S, Li G, Ma X, et al. Norcantharidin enhances ABT-737-induced apoptosis in hepatocellular carcinoma cells by transcriptional repression of Mcl-1. Cell Signal. 2012;24(9):1803–1809. doi: 10.1016/j.cellsig.2012.05.009 [DOI] [PubMed] [Google Scholar]
- 23.Yu T, Hou F, Liu M, et al. Norcantharidin anti-angiogenesis activity possibly through an endothelial cell pathway in human colorectal cancer. Asian Pac J Cancer Prev. 2012;13(2):499–503. doi: 10.7314/APJCP.2012.13.2.499 [DOI] [PubMed] [Google Scholar]
- 24.Yu CC, Ko FY, Yu CS, et al. Norcantharidin triggers cell death and DNA damage through S-phase arrest and ROS-modulated apoptotic pathways in TSGH 8301 human urinary bladder carcinoma cells. Int J Oncol. 2012;41(3):1050–1060. doi: 10.3892/ijo.2012.1511 [DOI] [PubMed] [Google Scholar]
- 25.Xiao W, Dai B, Zhu Y, Ye D. Norcantharidin induces autophagy-related prostate cancer cell death through Beclin-1 upregulation by miR-129-5p suppression. Tumour Biol. 2016;37: 15643––15648.. doi: 10.1007/s13277-015-4488-6 [DOI] [PubMed] [Google Scholar]
- 26.Chen S, Qu X, Wan P, et al. Norcantharidin inhibits pre-replicative complexes assembly of HepG2 cells. Am J Chin Med. 2013;41(3):665–682. doi: 10.1142/S0192415X13500468 [DOI] [PubMed] [Google Scholar]
- 27.Chen S, Wan P, Ding W, et al. Norcantharidin inhibits DNA replication and induces mitotic catastrophe by degrading initiation protein Cdc6. Int J Mol Med. 2013;32(1):43–50. doi: 10.3892/ijmm.2013.1359 [DOI] [PubMed] [Google Scholar]
- 28.Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414(6859):105–111. doi: 10.1038/35102167 [DOI] [PubMed] [Google Scholar]
- 29.Ferreira-Teixeira M, Parada B, Rodrigues-Santos P, et al. Functional and molecular characterization of cancer stem-like cells in bladder cancer: a potential signature for muscle-invasive tumors. Oncotarget. 2015;6(34):36185–36201. doi: 10.18632/oncotarget.5517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ojha R, Jha V, Singh SK, Bhattacharyya S. Autophagy inhibition suppresses the tumorigenic potential of cancer stem cell enriched side population in bladder cancer. Biochim Biophys Acta. 2014;1842(11):2073–2086. doi: 10.1016/j.bbadis.2014.07.007 [DOI] [PubMed] [Google Scholar]
- 31.Garcia-Gomez I, Elvira G, Zapata AG, et al. Mesenchymal stem cells: biological properties and clinical applications. Expert Opin Biol Ther. 2010;10(10):1453–1468. doi: 10.1517/14712598.2010.519333 [DOI] [PubMed] [Google Scholar]
- 32.Hirschmann-Jax C, Foster AE, Wulf GG, et al. A distinct “side population” of cells with high drug efflux capacity in human tumor cells. Proc Natl Acad Sci U S A. 2004;101(39):14228–14233. doi: 10.1073/pnas.0400067101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ginestier C, Hur MH, Charafe-Jauffret E, et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007;1(5):555–567. doi: 10.1016/j.stem.2007.08.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Richardson GD, Robson CN, Lang SH, et al. CD133, a novel marker for human prostatic epithelial stem cells. J Cell Sci. 2004;117(Pt 16):3539–3545. doi: 10.1242/jcs.01222 [DOI] [PubMed] [Google Scholar]
- 35.Zhu Y, Pang S, Luo Y, et al. A modified method by differential adhesion for enrichment of bladder cancer stem cells. Int Braz J Urol. 2016;42(4):817–824. doi: 10.1590/S1677-5538.IBJU.2015.0409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Smith J, Tho LM, Xu N, Gillespie DA. The ATM-Chk2 and ATR-Chk1 pathways in DNA damage signaling and cancer. Adv Cancer Res. 2010;108:73–112. [DOI] [PubMed] [Google Scholar]
- 37.Fokas E, Prevo R, Hammond EM, Brunner TB, McKenna WG, Muschel RJ. Targeting ATR in DNA damage response and cancer therapeutics. Cancer Treat Rev. 2014;40(1):109–117. doi: 10.1016/j.ctrv.2013.03.002 [DOI] [PubMed] [Google Scholar]
- 38.Bouwman P, Jonkers J. The effects of deregulated DNA damage signalling on cancer chemotherapy response and resistance. Nat Rev Cancer. 2012;12(9):587–598. doi: 10.1038/nrc3342 [DOI] [PubMed] [Google Scholar]
- 39.Galluzzi L, Senovilla L, Vitale I, et al. Molecular mechanisms of cisplatin resistance. Oncogene. 2012;31(15):1869–1883. doi: 10.1038/onc.2011.627 [DOI] [PubMed] [Google Scholar]
- 40.Galluzzi L, Vitale I, Michels J, et al. Systems biology of cisplatin resistance: past, present and future. Cell Death Dis. 2014;5:e1257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liu Y, Hock JM, Van Beneden RJ, Li X. Aberrant overexpression of FOXM1 transcription factor plays a critical role in lung carcinogenesis induced by low doses of arsenic. Mol Carcinog. 2014;53(5):380–391. doi: 10.1002/mc.21989 [DOI] [PubMed] [Google Scholar]
- 42.Champeris TS, Kanellakis N, Symeonidou IE, et al. Licensing of DNA replication, cancer, pluripotency and differentiation: an interlinked world? Semin Cell Dev Biol. 2014;30::174–180. doi: 10.1016/j.semcdb.2014.03.013 [DOI] [PubMed] [Google Scholar]