

ABSTRACT
Background
Protein ingestion during recovery from resistance-type exercise increases postexercise muscle protein synthesis rates. Whey protein has been reported to have greater anabolic properties than soy protein, an effect which may be attributed to the higher leucine content of whey.
Objective
The objective of this study was to compare postprandial myofibrillar (MyoPS) and mitochondrial (MitoPS) protein synthesis rates after ingestion of carbohydrate with whey, soy, or soy protein enriched with free leucine (to match the leucine content of whey) during recovery from a single bout of concurrent resistance- and endurance-type exercise in young healthy men.
Methods
In a randomized, double-blind, parallel-group design, 36 healthy young recreationally active men (mean ± SEM age: 23 ± 0.4 y) received a primed continuous infusion of l-[ring-13C6]-phenylalanine and l-[ring-3,5-2H2]-tyrosine and ingested 45 g carbohydrate with 20 g protein from whey (WHEY), soy (SOY), or leucine-enriched soy (SOY + LEU) after concurrent resistance- and endurance-type exercise. Blood and muscle biopsies were collected over a 360 min postexercise recovery period to assess MyoPS and MitoPS rates, and associated signaling through the mammalian target of rapamycin complex 1 (mTORC1).
Results
Postprandial peak plasma leucine concentrations were significantly higher in WHEY (mean ± SEM: 322 ± 10 μmol/L) and SOY + LEU (328 ± 14 μmol/L) compared with SOY (216 ± 6 μmol/L) (P < 0.05). Despite the apparent differences in plasma leucinemia, MyoPS (WHEY: 0.054 ± 0.002; SOY: 0.053 ± 0.004; SOY + LEU: 0.056 ± 0.004%·h−1; P = 0.83), and MitoPS (WHEY: 0.061 ± 0.004; SOY: 0.061 ± 0.006; SOY + LEU: 0.063 ± 0.004%·h−1; P = 0.96) rates over the entire 360 min recovery period did not differ between treatments. Similarly, signaling through mTORC1Ser2448, p70S6kThr389, 4E-BP1Thr37/46, and rpS6Ser235/236 was similar between treatments.
Conclusion
Postexercise MyoPS and MitoPS rates do not differ after co-ingestion of carbohydrate with 20 g protein from whey, soy, or leucine-enriched soy protein during 360 min of recovery from concurrent resistance- and endurance-type exercise in young, recreationally active men. This trial was registered at Nederlands Trial Register as NTR5098.
Keywords: muscle protein synthesis, young men, carbohydrate, dietary protein, whey, soy, leucine, concurrent exercise
Introduction
Resistance- (1) and endurance-type (2) exercise increase skeletal muscle protein synthesis (MPS) rates. However, these modes of exercise appear to differentially regulate myofibrillar (MyoPS) and/or mitochondrial (MitoPS) protein subfractional synthetic responses (3, 4). Protein/amino acid intake has been shown to further augment postexercise increases in mixed muscle protein synthesis and MyoPS rates after resistance- (5, 6) and endurance- (7, 8) type exercise, and as such, is commonly recommended to optimize postexercise muscle conditioning (9). However, whether or not protein ingestion can further augment postexercise MitoPS rates has not been established. In addition to ingesting protein to support postexercise muscle conditioning, carbohydrate ingestion is important to facilitate restoration of depleted muscle glycogen stores. Therefore, nutritional strategies to promote exercise recovery generally incorporate the combined intake of carbohydrate and protein (9). Dose-response studies suggest that ingestion of 20 g of a high-quality protein source is sufficient to maximally stimulate MPS rates during recovery from resistance-type exercise (10, 11). In addition, some studies have reported differences in the capacity of various protein sources to stimulate MPS after resistance-type exercise (12–16). Differences in the anabolic properties of various protein sources have been attributed to differences in their digestion and absorption kinetics and/or amino acid content (17). In terms of amino acids, the leucine content of a protein is believed to be of particular importance as leucine has been shown to activate translation initiation (18, 19) and stimulate MPS rates in vivo in humans (20).
Whey and soy protein are nutritionally complete, high-quality proteins (21). Whey protein has relatively high indispensable amino acid (∼50%) and leucine (∼10–12%) contents compared with other proteins (22), and undergoes rapid digestion and absorption after ingestion (23). Soy protein is also regarded as a rapidly digested dietary protein (24), but has lower indispensable amino acid (∼36%) and leucine (∼6%) contents than does whey protein (22). As a consequence, ingestion of soy protein is typically followed by a rapid, but more moderate postprandial rise in plasma amino acid levels (12, 16). Although soy is generally considered a high-quality source of dietary protein, it has been reported to be less effective than whey protein (12, 16) and bovine milk (13) in its capacity to increase MPS rates during recovery from a single bout of resistance-type exercise. This may relate to reported differences in how amino acids derived from soy and milk protein are used. For example, protein-derived amino acid splanchnic retention (24, 25), amino acid catabolism (24, 26), and/or amino acid oxidation (16) all seem greater after ingestion of soy compared to milk-derived proteins. Alternatively, the reduced capacity of soy protein to increase postprandial MPS rates compared to whey protein during recovery from resistance-type exercise may simply be attributed to a lower postprandial rise in leucine concentration (12, 16). In support of this notion, ingestion of a protein blend containing whey, soy, and sodium caseinate has been shown to be as effective as whey protein at stimulating postexercise MPS rates when matched for leucine contents (27). Furthermore, supplementation of soy protein enriched with branched-chain amino acids (leucine, isoleucine, valine) has been reported to enhance whole-body protein synthesis rates (28). Therefore, the capacity of soy protein ingestion to stimulate postexercise MPS rates may be enhanced by addition of free leucine.
The present study examined the effects of co-ingesting 20 g whey protein (WHEY), 20 g soy protein (SOY), or 20 g soy protein enriched with free leucine [to match the total leucine content of whey protein (SOY + LEU)], with 45 g carbohydrate on postprandial MyoPS and MitoPS rates during recovery from a single bout of concurrent resistance- and endurance-type exercise in young, healthy, recreationally active men. We hypothesized that WHEY and SOY + LEU would result in higher postexercise MyoPS and MitoPS rates compared to SOY.
Methods
Participants
Thirty-six healthy recreationally active men (age 23 ± 0.4 y; height 1.80 ± 0.01 m; weight 75.0 ± 1.3 kg; values are mean ± SEM) volunteered to participate in this parallel group, double-blind, randomized controlled trial. “Recreationally active” was defined as engaging in sports or structured exercise 1–3 d/wk. Participants’ characteristics are presented in Table 1. The characteristics and corresponding data from the whey protein treatment group (WHEY) are also presented in the accompanying manuscript (29). This study was part of a larger trial registered at the Nederlands Trial Register (NTR5098), and was conducted between March 2015 and May 2016 at Maastricht University in Maastricht, Netherlands. All participants were informed of the purpose of the study, the experimental procedures, and possible risks before providing informed written consent to participate. The procedures followed were in accordance with the ethical standards of the medical ethics committee of Maastricht University Medical Centre+ on human experimentation and in accordance with the Helsinki Declaration of 1975 as revised in October 2013. The study was independently monitored by the Clinical Trial Centre Maastricht (CTCM).
TABLE 1.
Characteristics of young recreationally active men who ingested nutritional treatments consisting of carbohydrate co-ingested with whey, soy, or free leucine-enriched soy protein after a single bout of concurrent exercise1
| Nutritional treatment group | ||||
|---|---|---|---|---|
| WHEY | SOY | SOY + LEU | P value | |
| Age, y | 23 ± 1 | 23 ± 1 | 23 ± 1 | 0.98 |
| Height, m | 1.80 ± 0.01 | 1.80 ± 0.02 | 1.80 ± 0.02 | 0.97 |
| Weight, kg | 76.0 ± 2.0 | 75.2 ± 2.5 | 73.8 ± 2.4 | 0.79 |
| BMI, kg/m2 | 23.4 ± 0.6 | 23.1 ± 0.6 | 22.9 ± 0.7 | 0.81 |
| Fat + bone-free mass, kg | 58.6 ± 1.5 | 59.6 ± 1.8 | 56.9 ± 1.5 | 0.50 |
| Fat mass, kg | 15.3 ± 0.9 | 13.6 ± 0.9 | 14.4 ± 1.0 | 0.45 |
| % Fat | 19.8 ± 0.9 | 17.7 ± 0.9 | 19.2 ± 1.0 | 0.25 |
| Systolic blood pressure, mm Hg | 123 ± 3 | 131 ± 3 | 123 ± 3 | 0.10 |
| Diastolic blood pressure, mm Hg | 64 ± 4 | 68 ± 2 | 66 ± 3 | 0.62 |
| Leg press 1-RM, kg | 271 ± 13 | 265 ± 16 | 281 ± 11 | 0.70 |
| Leg extension 1-RM, kg | 121 ± 5 | 124 ± 7 | 124 ± 6 | 0.95 |
| Maximal workload, W | 266 ± 13 | 267 ± 12 | 267 ± 10 | 1.00 |
Values are mean ± SEM. n = 12 for WHEY, 12 for SOY, and 12 for SOY + LEU. Data were analyzed with use of a 1-factor ANOVA. WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY; 1-RM, 1 repetition maximum.
Preliminary testing
Participants aged 20–30 y inclusive, with a BMI >19.0 and <25.0 (kg/m2) underwent an initial screening session to assess height, weight, blood pressure, and body composition (by dual-energy X-ray absorptiometry; Discovery A, Hologic). Participants were deemed healthy based on their responses to a medical questionnaire and screening results. After assessment of baseline anthropometrics, participants were familiarized with the exercise testing protocol and the exercise equipment. All exercise testing during the preliminary testing visit was supervised by >1 of the study investigators. Participants underwent estimates of 1 repetition maximum (1-RM) strength on the supine leg press (Technogym BV) and seated leg extension (Technogym BV) exercise through use of the multiple repetition testing procedure (30). Before testing each exercise, participants performed 10 submaximal repetitions to become familiar with the equipment and to have exercise technique assessed and adjusted by 1 of the study investigators. Working sets were then performed with progressively increased loads until failure to perform a valid estimation within 3–6 repetitions of the set. A repetition was considered valid if the subject was able to complete it in a controlled manner as determined by a study investigator. A 2-min inter-set rest period was allowed between successive sets. After estimates of 1-RM on the leg press and leg extension exercise, peak power output was determined during an incremental test to volitional fatigue on a cycle ergometer (Lode BV). Participants began cycling at a workload equivalent to 2 W/kg bodyweight for 150 s, after which the workload was increased by 25 W every 150 s until volitional fatigue was reached, defined as the inability to maintain a cadence >60 revolutions/min. All equipment settings were noted and replicated during the experimental test day. The pretesting and experimental trials were separated by >5 d.
Study design
Participants were randomly assigned to ingest a beverage (590 mL) containing 45 g of carbohydrate with 20 g whey protein (WHEY), 20 g soy protein (SOY), or 20 g soy protein supplemented with free leucine to match the leucine content of whey protein (SOY + LEU). The carbohydrate powder was supplied by PepsiCo Inc, and was composed of dextrose and maltodextrin. Whey protein concentrate (Nutri Whey 800F) and soy protein isolate (Unisol DP IP Non GMO) were obtained from FrieslandCampina DMV B.V. and Vitablend Nederland B.V., respectively. The leucine was obtained from Frutarom. Details of the amino acid, protein, and carbohydrate contents of the nutritional treatment are shown in Table 2. Random assignment was performed with a computerized list randomizer (https://www.random.org/lists/), and participants were sequentially allocated to a treatment according to the random assignment list.
TABLE 2.
Amino acid (l-form), protein, and carbohydrate contents of nutritional treatments consisting of carbohydrate co-ingested with whey, soy, or free leucine-enriched soy protein after a single bout of concurrent exercise in young recreationally active men1
| Nutritional treatment group | |||
|---|---|---|---|
| WHEY | SOY | SOY + LEU | |
| Amino acid content, g | |||
| Alanine | 1.02 | 0.76 | 0.76 |
| Arginine | 0.62 | 1.34 | 1.34 |
| Asparagine | 2.36 | 2.04 | 2.04 |
| Cysteine | 0.56 | 0.22 | 0.22 |
| Glutamine | 3.66 | 3.36 | 3.36 |
| Glycine | 0.38 | 0.74 | 0.74 |
| Histidine | 0.44 | 0.46 | 0.46 |
| Isoleucine | 1.14 | 0.86 | 0.86 |
| Leucine | 2.58 | 1.44 | 1.44 |
| Lysine | 2.14 | 1.12 | 1.12 |
| Methionine | 0.48 | 0.22 | 0.22 |
| Phenylalanine | 0.78 | 0.92 | 0.92 |
| Proline | 1.02 | 0.90 | 0.90 |
| Serine | 0.94 | 0.92 | 0.92 |
| Threonine | 1.08 | 0.68 | 0.68 |
| Tryptophan | 0.42 | 0.24 | 0.24 |
| Tyrosine | 0.74 | 0.66 | 0.66 |
| Valine | 1.06 | 0.88 | 0.88 |
| Added free amino acids, g | |||
| Leucine | 0.00 | 0.00 | 1.142 |
| Totals, g | |||
| Leucine | 2.58 | 1.44 | 2.58 |
| ƩNEAA | 11.30 | 10.94 | 10.94 |
| ƩEAA | 10.12 | 6.82 | 7.96 |
| ƩAA | 21.42 | 17.76 | 18.90 |
| Protein3 | 20.00 | 20.00 | 20.00 |
| Carbohydrate | 45.00 | 45.00 | 45.00 |
AA, amino acids; EAA, essential amino acids; NEAA, nonessential amino acids;WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
The added leucine in SOY + LEU was in addition to 20 g total protein.
Total protein was calculated as nitrogen × 6.38 for whey and nitrogen × 6.25 for soy protein.
Diet and physical activity
All participants were instructed to refrain from strenuous physical activity and alcohol consumption for 3 d before the experimental trial. In addition, all participants were instructed to fill out food intake and physical activity questionnaires for 2 d before the experimental trial. On the evening before the experimental trial, all participants consumed a prepackaged standardized meal containing 55% energy as carbohydrate, 30% energy as fat, and 15% energy as protein before 2000, after which they remained fasted.
Experimental protocol
At ∼0745, participants arrived at the laboratory in the overnight postabsorptive state. A catheter was inserted into an antecubital vein for stable isotope amino acid infusion, while a second catheter was subsequently inserted into a dorsal hand vein on the contralateral arm for arterialized blood sampling. To obtain arterialized blood samples, the hand was placed in a hot box (60°C) for 10 min before sample collection (31). After taking a baseline blood sample (t = −150 min), the plasma phenylalanine pool was primed with a single dose of l-[ring-13C6]-phenylalanine (2.25 μmol/kg) and l-[ring-3,5-2H2]-tyrosine (0.867 μmol/kg), and a continuous intravenous infusion of l-[ring-13C6]-phenylalanine (0.05 μmol kg−1 min−1) and l-[ring-3,5-2H2]-tyrosine (0.019 μmol kg−1 min−1) was initiated (t = −150 min) with use of a calibrated IVAC 598 pump. After resting in a supine position for 60 min, a second arterialized blood sample was drawn (t = −90 min). After resting for another 30 min, participants initiated (t = −60 min) the concurrent exercise intervention (described subsequently). A third blood sample was drawn (t = −30 min) during the transition from resistance- to endurance-type exercise. Immediately after the exercise intervention (t = 0 min), an arterialized blood sample was drawn and a muscle biopsy sample was collected from the vastus lateralis of a randomly selected leg. Subsequently, participants received a 590 mL beverage corresponding to their randomly assigned treatment allocation [i.e., WHEY (n = 12), SOY (n = 12), SOY + LEU (n = 12)]. The beverages were enriched to 4% l-[ring-13C6]-phenylalanine to minimize dilution of the steady-state plasma l-[ring-13C6]-phenylalanine precursor pool implemented by the constant infusion. Arterialized blood samples were then collected at t = 15, 30, 60, 90, 120, 150, 180, 240, 300, and 360 min in the postprandial period. Second and third muscle biopsy samples were collected at t = 120 and t = 360 min to determine postprandial MyoPS and MitoPS rates from t = 0–120, 120–360, and 0–360 min. Blood samples were collected into EDTA-containing tubes and centrifuged at 1000 × g for 15 min at 4°C. Aliquots of plasma were frozen in liquid nitrogen and stored at −80°C. Biopsy samples were collected with use of a 5-mm Bergström needle custom-adapted for manual suction. Samples were obtained from separate incisions from the middle region of the vastus lateralis, ∼15 cm above the patella and ∼3 cm below entry through the fascia, under 1% xylocaine local anesthesia with adrenaline (1:100,000). Muscle samples were freed from any visible non-muscle material, immediately frozen in liquid nitrogen, and stored at −80°C until further processing. When the experimental protocol was complete, cannulae were removed and subjects were fed and assessed for ∼30 min before leaving the laboratory. For a schematic representation of the infusion protocol, see Figure 1.
FIGURE 1.

Schematic representation of the experimental design.
Concurrent exercise protocol
Resistance-type exercise
Participants began with a standardized warm-up on the supine leg press (1 × 10 repetitions at ∼50% estimated 1-RM), followed by 4 sets of 8 repetitions at ∼80% of their previously estimated 1-RM. Participants then carried out the same exercise protocol (i.e., same number of sets and repetitions at % estimated 1-RM) on the seated leg extension machine. Each set was separated by 2 min of passive recovery during which time the subject remained seated. Range of motion was set from ∼70–155° for the leg press and from ∼75–165° for the leg extension. Strong verbal encouragement was provided by 1 of the study investigators during each set.
Endurance-type exercise
After the resistance-type exercise, participants performed 30 min of continuous cycling at ∼60% of their previously determined maximal workload (Wmax). Participants were allowed ad libitum access to water during cycling. Visual feedback for pedal frequency and elapsed time were provided to participants and strong verbal encouragement was provided by 1 of the study investigators.
Plasma and muscle tissue analyses
Plasma analyses
Details of analysis relating to the determination of plasma glucose, insulin, and amino acid concentrations as well as plasma l-[ring-13C6]-phenylalanine, l-[ring-13C6]-tyrosine, and l-[ring-3,5-2H2]-tyrosine enrichments are presented in Supplemental Methods.
Muscle tissue analyses
A piece of wet muscle (∼100 mg) was homogenized on ice with use of a Teflon pestle in ice-cold homogenization buffer (10 μL/mg; 1 M sucrose, 1 M Tris/HCl, 1 M KCl, 1 M EDTA) containing protease/phosphatase inhibitor cocktail tablets (Complete Protease Inhibitor Mini-Tabs; and PhosSTOP, Roche Applied Science). After ∼5–10 min of hand homogenization, the homogenate was centrifuged at 700 ×g for 15 min at 4°C to pellet a myofibrillar protein-enriched fraction. The supernatant was transferred to another tube and centrifuged at 12,000 × g for 20 min at 4°C to pellet a mitochondrial protein-enriched fraction. The resulting supernatant was used for Western Blot analysis. Additional details regarding the preparation and analysis of skeletal muscle samples for measurement of myofibrillar and mitochondrial protein-bound phenylalanine enrichment, and intramuscular signaling via Western Blot are presented in Supplementary Methods.
Calculations
The FSR of myofibrillar and mitochondrial protein enriched fractions was calculated through use of the standard precursor-product equation
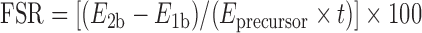 |
(1) |
where Eb is the increment in myofibrillar or mitochondrial protein-bound l-[ring-13C6]-phenylalanine enrichment mole % excess (MPE) between 2 muscle biopsy samples, Eprecursor is the weighted mean plasma l-[ring-13C6]-phenylalanine enrichment (MPE) during the tracer incorporation period, and t is the tracer incorporation time in h. Weighted mean plasma enrichments were calculated by taking the measured enrichment between consecutive time points and correcting for the time between these sampling time points. For calculation of postprandial FSR, biopsy samples at t = 0, 120, and 360 min were used.
Statistical analysis
Subjects’ characteristics, 1-RM strength, and Wmax data were analyzed with use of a 1-factor (treatment) ANOVA. Blood glucose and plasma insulin were analyzed with use of a 2-factor (treatment × time) repeated-measures ANOVA. Plasma leucine, phenylalanine, and tyrosine concentrations were analyzed with use of a 2-factor (treatment × time) repeated-measures ANOVA. Leucine, phenylalanine, and tyrosine AUC was analyzed with use of a 1-factor (treatment) ANOVA. Plasma enrichments were analyzed with use of a 2-factor (treatment × time) repeated-measures ANOVA. Myofibrillar and mitochondrial FSR during early and late recovery (i.e., 0–120 and 120–360 min) and protein phosphorylation status (i.e., 0, 120, and 360 min) were analyzed with use of a 2-factor (treatment × time) repeated-measures ANOVA. The aggregate myofibrillar and mitochondrial FSR (i.e., 0–360 min) was analyzed with use of a 1-factor (treatment) ANOVA. A power calculation was performed with differences in postprandial myofibrillar protein FSR as the primary outcome measure with the use of a standard deviation of 0.0065%·h−1 in all treatments, and a difference in FSR of 0.008%·h−1 between treatments (or ∼20% when expressed as relative difference between treatments). With a power of 80% and a significance level of 0.05, the final number of participants to be included was calculated as n = 12 per group. Tukey's post hoc analysis was performed whenever a significant F ratio was found to isolate specific differences. Statistical analyses were performed with a software package (IBM SPSS Statistics for Windows, version 21.0, IBM Corp.). Means were considered to be significantly different for P values <0.05.
Results
Plasma analyses
Plasma glucose concentrations (Figure 2A) were transiently increased from t = 15–60 min after ingestion of the protein-carbohydrate containing treatment beverages (P < 0.001). Similarly, plasma insulin concentrations (Figure 2B) were increased during the postprandial period (P < 0.001) from t = 15–90 min after beverage intake. Plasma leucine concentrations (Figure 3A) were increased in each treatment group during the postprandial period, with WHEY (from t = 15–150 min) and SOY + LEU (from t = 15–180 min) resulting in higher leucine concentrations when compared with SOY (P-interaction < 0.001). Plasma leucine AUC (Figure 3B) over the 360 min postprandial period was greater in WHEY and SOY + LEU compared with SOY (P < 0.001). Plasma phenylalanine concentrations (Figure 3C) were transiently increased after protein-carbohydrate co-ingestion, with higher concentrations in response to SOY at t = 60 and t = 300 min, and both SOY and SOY + LEU from t = 90–180 min compared with WHEY during the postprandial period (P-interaction < 0.01). Plasma phenylalanine AUC (data not shown) was greater in SOY and SOY + LEU compared with WHEY (P = 0.003). Plasma tyrosine concentrations (Figure 3D) were increased from t = 15–120 min during the postprandial period (P < 0.001). Plasma tyrosine AUC (data not shown) did not differ between treatments (P = 0.68).
FIGURE 2.
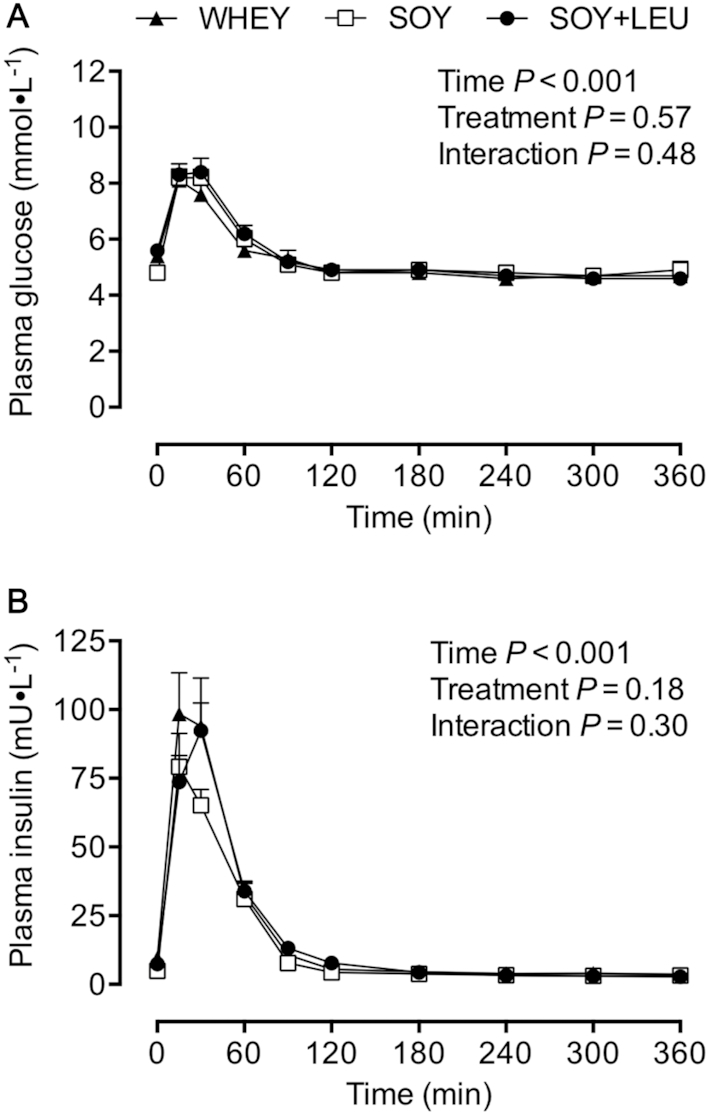
Plasma glucose (A) and insulin (B) concentrations during postabsorptive conditions (t = 0 min), and during postprandial conditions (t = 15–360 min) after beverage intake during recovery from a single bout of concurrent exercise in young men. Data for glucose and insulin were analyzed with use of a 2-factor repeated measures ANOVA. Values are mean ± SEM. n = 12. WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
FIGURE 3.

Plasma leucine (A), leucine AUC (B), phenylalanine (C), and tyrosine (D) concentrations during postabsorptive conditions (t = 0 min), and during postprandial conditions (t = 15–360 min) after beverage intake during recovery from a single bout of concurrent exercise in young men. Data for leucine, phenylalanine, and tyrosine were analyzed with use of a 2-factor repeated measures ANOVA. Data for leucine AUC were analyzed with use of a 1-factor ANOVA. Values are mean ± SEM. n = 12. Labeled means within a time without a common letter differ, P < 0.05. WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
Stable isotope tracer analyses
Plasma l-[ring-13C6]-phenylalanine enrichments (Figure 4) were different between SOY and SOY + LEU at t = 0, 15, and 60 min, different between WHEY and SOY + LEU at t = 30 and 120 min, and different between WHEY and SOY at t = 60, 90, 120, and 180 min (P-interaction < 0.01).
FIGURE 4.
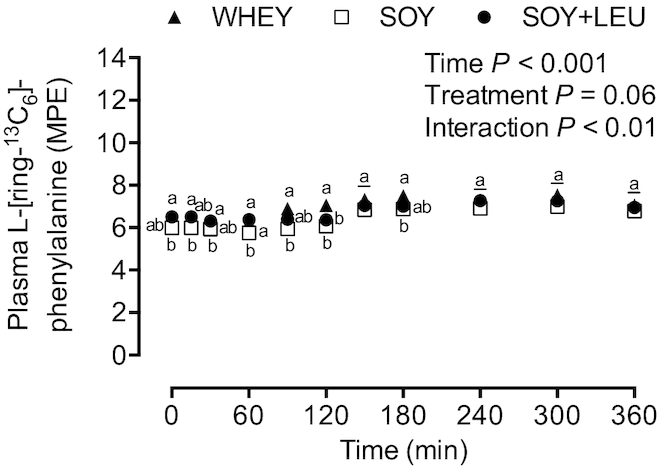
Plasma l-[ring-13C6]-phenylalanine enrichments during postabsorptive conditions (t = 0 min), and during postprandial conditions (t = 15–360 min) after beverage intake during recovery from a single bout of concurrent exercise in young men. Data were analyzed with use of a 2-factor repeated measures ANOVA. Values are mean ± SEM. n = 12. Labeled means within a time without a common letter differ, P < 0.05. MPE, mole % excess; WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
Postprandial MyoPS rates (i.e., myofibrillar FSR), assessed during early (0–120 min) and late (120–360 min) postexercise recovery (Figure 5A), did not differ between treatments (P = 0.69). MyoPS rates were higher during early (0–120 min) compared with late (120–360 min) postexercise recovery (P = 0.01). Aggregate (i.e., 0–360 min) MyoPS rates (Figure 5B) did not differ between treatments (P = 0.83). Postprandial MitoPS rates (i.e., mitochondrial FSR), assessed during early (0–120 min) and late (120–360 min) postexercise recovery (Figure 6A), were greater in SOY + LEU compared to WHEY and SOY during early, but not late, postexercise recovery (P-interaction = 0.03). Aggregate (i.e., 0–360 min) MitoPS rates (Figure 6B) did not differ between treatments (P = 0.96).
FIGURE 5.

Myofibrillar protein FSR over 0–120 and 120–360 min (A), and over 0–360 min (B) after beverage intake during recovery from a single bout of concurrent exercise in young men. Time-course (A) data were analyzed with use of a 2-factor repeated measures ANOVA. Aggregate (B) data were analyzed with use of a 1-factor ANOVA. Boxes represent 25th to 75th percentiles. Horizontal lines and crosses within boxes represent medians and means, respectively. Whiskers represent minimums and maximums. n = 12. FSR, fractional synthetic rate; WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
FIGURE 6.

Mitochondrial protein FSR over 0–120 and 120–360 min (A), and over 0–360 min (B) after beverage intake during recovery from a single bout of concurrent exercise in young men. Time-course (A) data were analyzed with use of a 2-factor repeated measures ANOVA. Aggregate (B) data were analyzed with use of a 1-factor ANOVA. Boxes represent 25th to 75th percentiles. Horizontal lines and crosses within boxes represent medians and means, respectively. Whiskers represent minimums and maximums. n = 12 for WHEY, 11 for SOY, and 12 for SOY + LEU. Labeled means within a time without a common letter differ, P < 0.05. FSR, fractional synthetic rate; WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
Muscle tissue signaling
The phosphorylation status of mTORSer2448 (Figure 7A) was not different between treatments (P = 0.65), but was increased during the postprandial period after concurrent exercise at both t = 120 min and t = 360 min (P < 0.01). The phosphorylation status of p70S6k (ribosomal protein S6 kinase)Thr389 (Figure 7B) was not different between treatments (P = 0.17), and was not increased during the postprandial period after concurrent exercise (P = 0.17). 4E-BP1 (eukaryotic initiation factor 4E binding protein 1)Thr37/46 phosphorylation (Figure 7C) was increased during the postprandial period after concurrent exercise at t = 120 min, and more so at t = 360 min when compared to t = 0 min (P < 0.01), with no differences between treatments (P = 0.18). The phosphorylation of rpS6 (ribosomal protein S6)Ser235/236 (Figure 7D) was increased at t = 120 min, and more so at t = 360 min during the postprandial period after concurrent exercise when compared to t = 0 min (P < 0.01), with no differences between treatments (P = 0.66). Representative Western Blot images are shown in Figure 8.
FIGURE 7.

Phosphorylation of mTORSer2448 (A), p70S6kThr389 (B), 4E-BP1Thr37/46 (C), and rpS6Ser235/236 (D) relative to the total abundance of their corresponding protein during postabsorptive conditions (t = 0 min), and during postprandial conditions (t = 120 and 360 min) after beverage intake during recovery from a single bout of concurrent exercise in young men. Data at t = 120 min and t = 360 min are expressed as fold-change from t = 0 min. Data were analyzed with use of a 2-factor repeated measures ANOVA. Values are mean ± SEM. n = 12. mTOR, mammalian target of rapamycin; p70S6k, ribosomal protein S6 kinase; rpS6, ribosomal protein S6; WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY; 4E-BP1, eukaryotic initiation factor 4E binding protein 1.
FIGURE 8.

Representative Western Blot images for phosphorylated (p) and total mammalian target of rapamycin (mTORSer2448), ribosomal protein S6 kinase (p70S6kThr389), eukaryotic initiation factor 4E binding protein 1 (4E-BP1Thr37/46), and ribosomal protein S6 (rps6Ser235/236) during postabsorptive conditions (t = 0 min), and during postprandial conditions (t = 120 and 360 min) after beverage intake during recovery from a single bout of concurrent exercise in young men. WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY.
Discussion
In the present study, we did not observe higher MyoPS rates during recovery from concurrent resistance- and endurance-type exercise after co-ingestion of 45 g of carbohydrate with 20 g whey (WHEY) or 20 g soy protein enriched with free leucine (SOY + LEU) when compared with 20 g soy protein (SOY). This lack of difference was observed despite substantially greater postprandial plasma leucine concentrations after WHEY and SOY + LEU when compared with SOY. Similarly, although we observed greater MitoPS rates during early (t = 0–120 min) recovery from concurrent exercise after SOY + LEU when compared to WHEY and SOY, late (t = 120–360 min) and overall aggregate (t = 0–360 min) MitoPS rates did not differ among treatments.
Ingestion of dietary protein and exercise (i.e., skeletal muscle contraction) represent 2 of the most potent regulators of skeletal muscle protein metabolism (32). The increase in postprandial amino acid availability after protein ingestion results in a marked stimulation of MPS rates (for review see (33)). The postprandial increase in MPS rates is attributed to an increase in circulating indispensable amino acids (IAAs) (34), with leucine (20) being of particular relevance. The postprandial rise in dispensable amino acid concentrations appears to be of less relevance to the stimulation of MPS rates (35). The increase in MPS rates in response to amino acids is further augmented by prior resistance-type exercise (6). Studies examining the dose-dependent relationship between protein ingestion and MPS rates after resistance-type exercise in young adults have demonstrated that ingestion of ∼20 g of a high-quality protein source is sufficient to maximize postexercise mixed muscle protein synthesis (11) and MyoPS (10) rates. Ingestion of more protein does not further stimulate protein synthesis, but is instead oxidized (11) and directed towards ureagenesis (10). As such, in the present study we provided subjects with 20 g whey or soy protein to compare these protein sources in their capacity to support MyoPS and MitoPS rates during recovery from a single bout of combined resistance- and endurance-type exercise (i.e., concurrent exercise).
Whey protein is among the highest quality sources of dietary protein because of its amino acid profile (high IAA, branched-chain amino acid, and leucine contents), rapid digestibility, and robust capacity to stimulate postprandial MPS rates (22). Soy protein is also a rapidly digested high-quality protein source with a favorable IAA content, but ingestion of soy protein has been reported to be less effective in stimulating MPS rates when compared with the ingestion of whey (12, 16) and bovine milk (13) protein during recovery from resistance-type exercise. The lower capacity of soy protein to stimulate postexercise MPS rates when compared to whey may be attributed to its low(er) leucine content (6–8% compared with 10–12% leucine). As such, we also examined the impact of ingesting soy protein enriched with free leucine (to match that of whey protein) on postexercise MyoPS and MitoPS rates. Peak plasma leucine concentrations were considerably higher after WHEY (322 ± 10 μmol/L; ∼152 ± 10% increase) when compared to SOY (216 ± 6 μmol/L; ∼75 ± 7% increase) protein. Fortification of soy protein with free leucine (SOY + LEU) increased postprandial peak leucine concentrations substantially, with levels similar to WHEY (328 ± 14 μmol/L; ∼165 ± 15% increase). In agreement, total plasma leucine exposure (AUC) during the entire 6 h (360 min) recovery period was greater after WHEY and SOY + LEU when compared with SOY (Figure 3B). Despite the differences in postprandial plasma leucinemia between treatments (Figure 3A and B), we observed no differences in MyoPS rates after WHEY, SOY, or SOY + LEU when assessed during the early (from 0–120 min), late (120–360 min), or aggregate 6 h (0–360 min) postexercise recovery periods (Figure 5A and B). Similarly, aside from greater early MitoPS rates after SOY + LEU, we observed no differences in MitoPS rates among treatments.
Based on previous research comparing postexercise MPS rates after ingestion of soy compared to whey (12, 16) or milk (13) protein, we hypothesized that WHEY and SOY + LEU would result in higher MyoPS and MitoPS rates when compared with SOY. Previous research comparing the postprandial metabolic use of dietary nitrogen from soy and total milk protein demonstrated that milk protein better supported “peripheral” protein synthesis (i.e., skeletal muscle) whereas soy protein better stimulated splanchnic protein synthesis rates when assessed under resting conditions (24, 25). Subsequent work from Wilkinson and colleagues (13) demonstrated that ingestion of milk (∼18 g protein) resulted in a more positive amino acid balance across the leg and greater MPS rates when compared to the ingestion of an isonitrogenous, isoenergetic, and macronutrient composition-matched amount of soy protein during recovery from resistance-type exercise. It was proposed that the attenuated postprandial rise in protein-derived amino acid availability after milk as opposed to soy protein ingestion may have been responsible for the greater leg amino acid uptake and higher MPS rates during recovery from resistance-type exercise (13). However, later work from Tang and colleagues (12) demonstrated that more rapidly digested whey protein resulted in higher postprandial MPS rates when compared with the ingestion of soy and micellar casein protein. Whereas micellar casein has been classified as a more slowly digested dietary protein (23), the protein digestion and amino acid absorption kinetics of soy protein seem to be more similar to whey (25). Therefore, potential differences in the anabolic properties of soy compared with whey protein ingestion during postexercise recovery are unlikely to be related to differences in protein digestion and amino acid absorption kinetics and may be more related to differences in amino acid composition of the proteins. The IAA and leucine contents of whey protein are considerably greater than those of soy protein (36). Leucine is a major nutrient regulator of translation initiation (18, 19) that can stimulate MPS rates in humans (20). Supplementing soy protein with BCAAs (leucine, isoleucine, and valine) has also been reported to increase its postprandial anabolic properties in elderly and clinical populations (28). Consequently, we hypothesized that fortifying 20 g soy protein with free leucine (SOY + LEU) to match the leucine content of 20 g whey protein would increase its anabolic properties and augment postprandial MyoPS and MitoPS rates when compared to the ingestion of soy only (SOY). In contrast to our hypotheses, we did not detect any differences in postprandial MyoPS rates after WHEY, SOY, or SOY + LEU during the early (0–120 min), late (120–360 min), as well as aggregate (0–360 min) periods of recovery after concurrent exercise. Similarly, aside from greater postprandial MitoPS rates after SOY + LEU during the early (0–120 min) period of recovery after concurrent exercise, MitoPS rates did not differ between treatments over the late (120–360 min) or aggregate (0–360 min) postexercise recovery periods.
The absence of differences in MyoPS rates during recovery from concurrent exercise after ingestion of 20 g whey, soy, or free leucine-enriched soy, implies that exercise and/or nutrient signals that regulate postexercise MyoPS rates were equivalent between treatment groups. Because the current study did not incorporate a nonprotein control treatment, we cannot determine the independent contribution of whey or soy protein ingestion to MyoPS rates during recovery from concurrent exercise. In an accompanying study (29), we show that co-ingestion of 20 g protein with 45 g carbohydrate resulted in only marginal increases in MyoPS rates (∼16%) when compared to the ingestion of 45 g carbohydrate only during recovery from concurrent exercise. Ingested carbohydrate and the associated increase in insulin availability may have stimulated increases in blood flow (37), microvascular perfusion (38), amino acid transport (39), and/or a suppression of protein degradation in skeletal muscle tissue (39). In addition, carbohydrate co-ingestion with protein may have delayed the digestion and absorption kinetics of the ingested proteins (40). As such, potential differences in MyoPS rates after ingestion of various sources of isolated dietary protein may be less apparent when co-ingested with carbohydrate. Consequently, greater doses of protein (e.g., 30 g) may be required to robustly increase MyoPS rates when protein is co-ingested with carbohydrate during recovery from concurrent exercise and, as such, may be necessary to reveal any differences in the anabolic properties of ingested whey, soy, or free leucine-enriched soy after exercise.
Currently, there is limited information available on the nutritional regulation of MitoPS in human muscle. Early studies demonstrated that amino acid provision via intravenous infusion may stimulate MitoPS rates at rest (41, 42). More recent studies have demonstrated that orally ingested protein can increase MitoPS rates at rest (43, 44), and that the source of ingested protein may modulate the response of MitoPS (44). However, studies to date evaluating the effect of protein ingestion on MitoPS rates in humans have failed to find support for the notion that protein ingestion positively augments MitoPS rates after exercise (7, 45–47). The results of the present study, and those reported in our accompanying study (29), are the first to report the effect of different sources of ingested protein on postprandial MitoPS after exercise. The reason for the greater increase in early (0–120 min) postprandial MitoPS rates after SOY + LEU in the current study is unclear, but may be a result of differences in the metabolism of free versus protein-bound leucine. Leucine has been reported to increase mitochondrial content and mitochondrial biogenesis-related gene expression in C2C12 myotubes (48, 49). Nonetheless, the effect was short-lived as MitoPS rates did not differ between treatments over the late (120–360 min) or aggregate (0–360 min) postexercise recovery periods.
In line with the absence of any major differences in MyoPS or MitoPS rates after WHEY, SOY, or SOY + LEU during recovery from concurrent exercise, we did not detect any differences in the phosphorylation status of the intracellular signaling proteins between treatments (Figure 7A–D). To date, few studies have compared signaling responses of protein targets of the mTORC1 pathway after ingestion of different sources of dietary protein during postexercise recovery (50–53). Anthony and colleagues (50) reported that rats fed carbohydrate plus whey protein demonstrated greater increases in mTORSer2448 and p70S6kThr389 phosphorylation compared to rats fed carbohydrate and soy protein after treadmill exercise (50). In accordance, Mitchell and colleagues (51) reported that ingestion of 30 g soy protein was less effective than whey protein at sustaining postprandial increases in p70S6kThr389 phosphorylation during recovery from resistance-type exercise in older adults. The reason for the lack of differences in signaling responses in mTORSer2448, p70S6kThr389, 4E-BP1Thr37/46, and rpS6Ser235/236 after ingestion of whey, soy, or free leucine-enriched soy protein despite substantial differences in plasma leucine availability remains unclear but may relate to the timing of biopsy sampling (i.e., at 120 and 360 min during the postprandial period) as well as the postexercise conditions in which these measurements were performed. A bout of concurrent exercise may have already increased amino acid availability to the muscle by increasing endogenous amino acid release, and/or stimulating skeletal muscle blood flow. These factors may have made the contribution of the postprandial release of exogenous amino acids less relevant to the changes in mTORC1 and its downstream targets. Alternatively, as peak signaling responses via mTORC1 and/or its downstream targets may occur early after protein intake (54), we may have missed divergent signaling responses among protein treatments that occurred before the biopsy taken at 120 minutes into the postprandial period after concurrent exercise. Either way, the lack of differences in myocellular signaling responses among treatments are in line with the absence of major differences in postexercise MyoPS and MitoPS rates.
In conclusion, co-ingestion of carbohydrate with 20 g whey, soy, or free leucine-enriched soy protein do not lead to differences in overall postexercise MyoPS or MitoPS rates during recovery from a single bout of concurrent resistance- and endurance-type exercise in recreationally active young men. Increasing the leucine content of soy protein to match that of whey protein does not further increase postexercise MyoPS or MitoPS rates during 360 min of recovery from concurrent resistance- and endurance-type exercise.
Supplementary Material
Acknowledgments
We thank Joan Senden, Joy Goessens, and Annemie Gijsen for their analytical assistance. The authors’ responsibilities were as follows: TAC-V, IR, and LJCvL: conceived and designed the research; TAC-V, PJMP, JSJS, WMP, AHZ, and HS: conducted the research; TAC-V, AHZ, and HS: analyzed the data; TAC-V and LJCvL: interpreted results of experiments; TAC-V: prepared figures; TAC-V: drafted the manuscript; TAC-V and LJCvL: edited and revised the manuscript; and all authors: read and approved the final paper.
Notes
Supported by PepsiCo/Gatorade Sports Science Institute.
Author disclosures: TAC-V, PJMP, JSJS, WMP, AHZ, HS, and LBV, no conflicts of interest. IR is an employee of the Gatorade Sports Science Institute, a division of PepsiCo Inc. The views expressed in this manuscript are those of the authors and do not necessarily reflect the position or policy of PepsiCo Inc. LJCvL has received research grants, consulting fees, speaking honoraria, or a combination of these, from Friesland Campina and PepsiCo.
Supplemental methods are available from the “Supplementary data” link in the online posting of the article and from the same link in the online table of contents at https://academic.oup.com/jn/.
Abbreviations used: 4E-BP1, eukaryotic initiation factor 4E binding protein 1; FSR, fractional synthetic rate; IAA, indispensable amino acids; MitoPS, mitochondrial protein synthesis; MPE, mole % excess; MPS, muscle protein synthesis; mTORC1, mammalian target of rapamycin complex 1; MyoPS, myofibrillar protein synthesis; p70S6k, ribosomal protein S6 kinase; rpS6, ribosomal protein S6; SOY, 45 g carbohydrate co-ingested with 20 g soy protein; SOY + LEU, 45 g carbohydrate co-ingested with 20 g soy protein enriched with leucine to match the leucine content of WHEY; WHEY, 45 g carbohydrate co-ingested with 20 g whey protein; 1-RM, 1 repetition maximum.
References
- 1. Chesley A, MacDougall JD, Tarnopolsky MA, Atkinson SA, Smith K. Changes in human muscle protein synthesis after resistance exercise. J Appl Physiol (1985). 1992;73:1383–8. [DOI] [PubMed] [Google Scholar]
- 2. Carraro F, Stuart CA, Hartl WH, Rosenblatt J, Wolfe RR. Effect of exercise and recovery on muscle protein synthesis in human subjects. Am J Physiol. 1990;259:E470–6. [DOI] [PubMed] [Google Scholar]
- 3. Wilkinson SB, Phillips SM, Atherton PJ, Patel R, Yarasheski KE, Tarnopolsky MA, Rennie MJ. Differential effects of resistance and endurance exercise in the fed state on signalling molecule phosphorylation and protein synthesis in human muscle. J Physiol. 2008;586:3701–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Donges CE, Burd NA, Duffield R, Smith GC, West DW, Short MJ, Mackenzie R, Plank LD, Shepherd PR, Phillips SM, et al.. Concurrent resistance and aerobic exercise stimulates both myofibrillar and mitochondrial protein synthesis in sedentary middle-aged men. J Appl Physiol (1985). 2012;112:1992–2001. [DOI] [PubMed] [Google Scholar]
- 5. Tipton KD, Ferrando AA, Phillips SM, Doyle D Jr., Wolfe RR. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am J Physiol. 1999;276:E628–34. [DOI] [PubMed] [Google Scholar]
- 6. Biolo G, Tipton KD, Klein S, Wolfe RR. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol. 1997;273:E122–9. [DOI] [PubMed] [Google Scholar]
- 7. Breen L, Philp A, Witard OC, Jackman SR, Selby A, Smith K, Baar K, Tipton KD. The influence of carbohydrate-protein co-ingestion following endurance exercise on myofibrillar and mitochondrial protein synthesis. J Physiol. 2011;589:4011–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Howarth KR, Moreau NA, Phillips SM, Gibala MJ. Coingestion of protein with carbohydrate during recovery from endurance exercise stimulates skeletal muscle protein synthesis in humans. J Appl Physiol (1985). 2009;106:1394–402. [DOI] [PubMed] [Google Scholar]
- 9. Thomas DT, Erdman KA, Burke LM. American college of sports medicine joint position statement. Nutrition and athletic performance. Med Sci Sports Exerc. 2016;48:543–68. [DOI] [PubMed] [Google Scholar]
- 10. Witard OC, Jackman SR, Breen L, Smith K, Selby A, Tipton KD. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am J Clin Nutr. 2014;99:86–95. [DOI] [PubMed] [Google Scholar]
- 11. Moore DR, Robinson MJ, Fry JL, Tang JE, Glover EI, Wilkinson SB, Prior T, Tarnopolsky MA, Phillips SM. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am J Clin Nutr. 2009;89:161–8. [DOI] [PubMed] [Google Scholar]
- 12. Tang JE, Moore DR, Kujbida GW, Tarnopolsky MA, Phillips SM. Ingestion of whey hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J Appl Physiol. 2009;107:987–92. [DOI] [PubMed] [Google Scholar]
- 13. Wilkinson SB, Tarnopolsky MA, Macdonald MJ, Macdonald JR, Armstrong D, Phillips SM. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am J Clin Nutr. 2007;85:1031–40. [DOI] [PubMed] [Google Scholar]
- 14. Burd NA, Gorissen SH, van Vliet S, Snijders T, van Loon LJ. Differences in postprandial protein handling after beef compared with milk ingestion during postexercise recovery: a randomized controlled trial. Am J Clin Nutr. 2015;102:828–36. [DOI] [PubMed] [Google Scholar]
- 15. Burd NA, Yang Y, Moore DR, Tang JE, Tarnopolsky MA,Phillips SM . Greater stimulation of myofibrillar protein synthesis with ingestion of whey protein isolate v. micellar casein at rest and after resistance exercise in elderly men. Br J Nutr. 2012;108:958–62. [DOI] [PubMed] [Google Scholar]
- 16. Yang Y, Churchward-Venne TA, Burd NA, Breen L, Tarnopolsky MA, Phillips SM. Myofibrillar protein synthesis following ingestion of soy protein isolate at rest and after resistance exercise in elderly men. Nutr Metab (Lond). 2012;9:57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Tang JE, Phillips SM. Maximizing muscle protein anabolism: the role of protein quality. Curr Opin Clin Nutr Metab Care. 2009;12:66–71. [DOI] [PubMed] [Google Scholar]
- 18. Crozier SJ, Kimball SR, Emmert SW, Anthony JC, Jefferson LS. Oral leucine administration stimulates protein synthesis in rat skeletal muscle. J Nutr. 2005;135:376–82. [DOI] [PubMed] [Google Scholar]
- 19. Anthony JC, Yoshizawa F, Anthony TG, Vary TC, Jefferson LS, Kimball SR. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J Nutr. 2000;130:2413–9. [DOI] [PubMed] [Google Scholar]
- 20. Wilkinson DJ, Hossain T, Hill DS, Phillips BE, Crossland H, Williams J, Loughna P, Churchward-Venne TA, Breen L, Phillips SM et al.. Effects of leucine and its metabolite β-hydroxy-β-methylbutyrate on human skeletal muscle protein metabolism. J Physiol. 2013;591:2911–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Rutherfurd SM, Fanning AC, Miller BJ, Moughan PJ. Protein digestibility-corrected amino acid scores and digestible indispensable amino acid scores differentially describe protein quality in growing male rats. J Nutr. 2015;145:372–9. [DOI] [PubMed] [Google Scholar]
- 22. Devries MC, Phillips SM. Supplemental protein in support of muscle mass and health: advantage whey. J Food Sci. 2015;80(Suppl 1):A8–A15. [DOI] [PubMed] [Google Scholar]
- 23. Boirie Y, Dangin M, Gachon P, Vasson MP, Maubois JL, Beaufrere B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc Natl Acad Sci U S A. 1997;94:14930–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Bos C, Metges CC, Gaudichon C, Petzke KJ, Pueyo ME, Morens C, Everwand J, Benamouzig R, Tome D. Postprandial kinetics of dietary amino acids are the main determinant of their metabolism after soy or milk protein ingestion in humans. J Nutr. 2003;133:1308–15. [DOI] [PubMed] [Google Scholar]
- 25. Fouillet H, Mariotti F, Gaudichon C, Bos C, Tome D. Peripheral and splanchnic metabolism of dietary nitrogen are differently affected by the protein source in humans as assessed by compartmental modeling. J Nutr. 2002;132:125–33. [DOI] [PubMed] [Google Scholar]
- 26. Luiking YC, Deutz NE, Jakel M, Soeters PB. Casein and soy protein meals differentially affect whole-body and splanchnic protein metabolism in healthy humans. J Nutr. 2005;135:1080–7. [DOI] [PubMed] [Google Scholar]
- 27. Reidy PT, Walker DK, Dickinson JM, Gundermann DM, Drummond MJ, Timmerman KL, Fry CS, Borack MS, Cope MB, Mukherjea R et al.. Protein blend ingestion following resistance exercise promotes human muscle protein synthesis. J Nutr. 2013;143:410–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Engelen MP, Rutten EP, De Castro CL, Wouters EF, Schols AM, Deutz NE. Supplementation of soy protein with branched-chain amino acids alters protein metabolism in healthy elderly and even more in patients with chronic obstructive pulmonary disease. Am J Clin Nutr. 2007;85:431–9. [DOI] [PubMed] [Google Scholar]
- 29. Churchward-Venne TA, Pinckaers PJ, Smeets JS, Peeters WM, Zorenc AH, Schierbeek H, Rollo I, Verdijk LB, van Loon LJ. Myofibrillar and mitochondrial protein synthesis rates do not differ in young men following ingestion of carbohydrate with milk protein, whey, or micellar casein during recovery after concurrent resistance- and endurance-type exercise. J Nutr. 2019;149(2):198–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Mayhew JL, Prinster JL, Ware JS, Zimmer DL, Arabas JR, Bemben MG. Muscular endurance repetitions to predict bench press strength in men of different training levels. J Sports Med Phys Fitness. 1995;35:108–13. [PubMed] [Google Scholar]
- 31. Abumrad NN, Rabin D, Diamond MP, Lacy WW. Use of a heated superficial hand vein as an alternative site for the measurement of amino acid concentrations and for the study of glucose and alanine kinetics in man. Metabolism. 1981;30:936–40. [DOI] [PubMed] [Google Scholar]
- 32. Rennie MJ, Wackerhage H, Spangenburg EE, Booth FW. Control of the size of the human muscle mass. Annu Rev Physiol. 2004;66:799–828. [DOI] [PubMed] [Google Scholar]
- 33. Reidy PT, Rasmussen BB. Role of ingested amino acids and protein in the promotion of resistance exercise-induced muscle protein anabolism. J Nutr. 2016;146:155–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Smith K, Reynolds N, Downie S, Patel A, Rennie MJ. Effects of flooding amino acids on incorporation of labeled amino acids into human muscle protein. Am J Physiol. 1998;275:E73–8. [DOI] [PubMed] [Google Scholar]
- 35. Volpi E, Kobayashi H, Sheffield-Moore M, Mittendorfer B, Wolfe RR. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78:250–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. van Vliet S, Burd NA, van Loon LJ. The skeletal muscle anabolic response to plant- versus animal-based protein consumption. J Nutr. 2015;145:1981–91. [DOI] [PubMed] [Google Scholar]
- 37. Baron AD. Hemodynamic actions of insulin. Am J Physiol. 1994;267:E187–202. [DOI] [PubMed] [Google Scholar]
- 38. Coggins M, Lindner J, Rattigan S, Jahn L, Fasy E, Kaul S, Barrett E. Physiologic hyperinsulinemia enhances human skeletal muscle perfusion by capillary recruitment. Diabetes. 2001;50:2682–90. [DOI] [PubMed] [Google Scholar]
- 39. Biolo G, Williams BD, Fleming RY, Wolfe RR. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes. 1999;48:949–57. [DOI] [PubMed] [Google Scholar]
- 40. Gorissen SH, Burd NA, Hamer HM, Gijsen AP, Groen BB, van Loon LJ. Carbohydrate coingestion delays dietary protein digestion and absorption but does not modulate postprandial muscle protein accretion. J Clin Endocrinol Metab. 2014;99:2250–8. [DOI] [PubMed] [Google Scholar]
- 41. Bohe J, Low A, Wolfe RR, Rennie MJ. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: a dose-response study. J Physiol. 2003;552:315–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Bohe J, Low JF, Wolfe RR, Rennie MJ. Latency and duration of stimulation of human muscle protein synthesis during continuous infusion of amino acids. J Physiol. 2001;532:575–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Beals JW, Mackenzie RWA, van Vliet S, Skinner SK, Pagni BA, Niemiro GM, Ulanov AV, Li Z, Dilger AC, Paluska SA et al.. Protein-rich food ingestion stimulates mitochondrial protein synthesis in sedentary young adults of different BMIs. J Clin Endocrinol Metab. 2017;102:3415–24. [DOI] [PubMed] [Google Scholar]
- 44. Walrand S, Gryson C, Salles J, Giraudet C, Migne C, Bonhomme C, Le Ruyet P, Boirie Y. Fast-digestive protein supplement for ten days overcomes muscle anabolic resistance in healthy elderly men. Clin Nutr. 2016;35:660–8. [DOI] [PubMed] [Google Scholar]
- 45. Abou Sawan S, van Vliet S, Parel JT, Beals JW, Mazzulla M, West DWD, Philp A, Li Z, Paluska SA, Burd NA et al.. Translocation and protein complex co-localization of mTOR is associated with postprandial myofibrillar protein synthesis at rest and after endurance exercise. Physiol Rep. 2018;6:1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Coffey VG, Moore DR, Burd NA, Rerecich T, Stellingwerff T, Garnham AP, Phillips SM, Hawley JA. Nutrient provision increases signalling and protein synthesis in human skeletal muscle after repeated sprints. Eur J Appl Physiol. 2011;111:1473–83. [DOI] [PubMed] [Google Scholar]
- 47. Camera DM, West DW, Phillips SM, Rerecich T, Stellingwerff T, Hawley JA, Coffey VG. Protein ingestion increases myofibrillar protein synthesis after concurrent exercise. Med Sci Sports Exerc. 2015;47:82–91. [DOI] [PubMed] [Google Scholar]
- 48. Liang C, Curry BJ, Brown PL, Zemel MB. Leucine modulates mitochondrial biogenesis and SIRT1-AMPK signaling in C2C12 myotubes. J Nutr Metab. 2014;2014:239750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Sun X, Zemel MB. Leucine modulation of mitochondrial mass and oxygen consumption in skeletal muscle cells and adipocytes. Nutr Metab (Lond). 2009;6:26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Anthony TG, McDaniel BJ, Knoll P, Bunpo P, Paul GL, McNurlan MA. Feeding meals containing soy or whey protein after exercise stimulates protein synthesis and translation initiation in the skeletal muscle of male rats. J Nutr. 2007;137:357–62. [DOI] [PubMed] [Google Scholar]
- 51. Mitchell CJ, Della Gatta PA, Petersen AC, Cameron-Smith D, Markworth JF. Soy protein ingestion results in less prolonged p70S6 kinase phosphorylation compared to whey protein after resistance exercise in older men. J Int Soc Sports Nutr. 2015;12:6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Reitelseder S, Agergaard J, Doessing S, Helmark IC, Lund P, Kristensen NB, Frystyk J, Flyvbjerg A, Schjerling P, van Hall G et al.. Whey and casein labeled with L-[1-13C]leucine and muscle protein synthesis: effect of resistance exercise and protein ingestion. Am J Physiol Endocrinol Metab. 2011;300:E231–42. [DOI] [PubMed] [Google Scholar]
- 53. Dideriksen KJ, Reitelseder S, Petersen SG, Hjort M, Helmark IC, Kjaer M, Holm L. Stimulation of muscle protein synthesis by whey and caseinate ingestion after resistance exercise in elderly individuals. Scand J Med Sci Sports. 2011;21:E372–83. [DOI] [PubMed] [Google Scholar]
- 54. Atherton PJ, Etheridge T, Watt PW, Wilkinson D, Selby A, Rankin D, Smith K, Rennie MJ. Muscle full effect after oral protein: time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am J Clin Nutr. 2010;92:1080–8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.