

Abstract
The physiological mechanisms underlying leaf growth inhibition under salt stress are not fully understood. Apoplastic pH is considered to play an important role in cell wall loosening and tissue growth and was demonstrated to be altered by several growth-limiting environmental conditions. In this study we have evaluated the possibility that inhibition of maize (Zea mays) leaf elongation by salinity is mediated by changes in growing cell wall acidification capacity. The kinetics of extended apoplast pH changes by leaf tissue of known expansion rates and extent of growth reduction under stress was investigated (in vivo) and was found similar for non-stressed and salt-stressed tissues at all examined apoplast salinity levels (0.1, 5, 10, or 25 mm NaCl). A similar rate of spontaneous acidification for the salt and control treatments was demonstrated also in in situ experiments. Unlike growing cells that acidified the external medium, mature nongrowing cells caused medium alkalinization. The kinetics of pH changes by mature tissue was also unchanged by salt stress. Fusicoccin, an enhancer of plasmalemma H+-ATPase activity level, greatly stimulated elongation growth and acidification rate to a similar extent in the control and salt treatments. That the ability of the growing tissue to acidify the apoplast did not change under same salt stress conditions that induced inhibition of tissue elongation rate suggests that salinity does not inhibit cell growth by impairing the acidification process or reducing the inherent capacity for cell wall acidification.
Inhibition of leaf elongation is one of the primary effects of salt stress. The physiological mechanisms underlying leaf growth inhibition are not fully understood (Munns, 1993; Lazof and Bernstein, 1998). In grasses during an important period of leaf development, leaf elongation is confined to a basal elongation zone located near the point of leaf attachment to the node. Leaf longitudinal expansion is dependent on irreversible expansion of cells located at this basal region. Salt stress reduces leaf growth rate by shortening the length of the leaf elongating zone and decreasing the growth intensity in its central and distal portions (Bernstein et al., 1993a, 1993b). Leaf growth inhibition by salinity must be expected to occur via an effect on this region (Lazof and Bernstein, 1998).
Cell and tissue expansion are generated by turgor-driven wall extension (Tomos and Pritchard, 1994). A modified capacitance of cell walls to irreversibly yield was suggested to be the major growth-limiting factor in long term growth inhibition under salt stress (Cramer and Bowman, 1991; Neumann et al., 1994). Apoplast pH was suggested to play an important role in cell wall loosening and growth (McQueen-Mason et al., 1993) and in numerous plant tissues including leaves of maize (Zea mays), increased rates of growth are associated with increased acidification of the cell wall space (for example, Van Volkenburgh and Boyer, 1985; Jahn et al., 1996; Peters et al., 1998; Stahlberg and Van Volkenburgh, 1999). Decreased cell wall pH is generally thought to promote wall-loosening events necessary for growing cell enlargement (Rayle and Cleland, 1992; Cosgrove, 1997), and inhibition of cell wall acidification may therefore reduce cell growth rate.
Several environmental conditions that affect growth were shown to alter apoplast acidification. For example, growth inhibition by water stress is accompanied by an increase in apoplastic pH and a decrease in acidification rate (Van Volkenburgh and Boyer, 1985; Hartung et al., 1988), and cold temperature shock increases extracellular pH of the motor cells of Mimosa pudica (Kumon and Suda, 1985), and gravitrophic stimulation enhances acid efflux and cell elongation along the upper surface of horizontally placed roots (Mulkey and Evans, 1981). The salt stress-induced growth inhibition might also be associated with modifications of apoplast acidification.
Some indications suggest possible inhibitory effects of Na+ and Cl− on H+ efflux and apoplast acidification (Newman et al., 1987; Ben-Hayyim and Ran, 1990; Wilson and Shannon, 1995). This might play a role in growth sensitivity under salt stress, since salinized cell walls contain higher levels of Na+ and Cl− than non-stressed walls, and cultivar salt tolerance sometimes correlates with a higher level of exclusion from cell wall and cytoplasm (Hajibagheri et al., 1987; Flowers et al., 1991; Lazof and Bernstein, 1998).
In addition to possible effects on cell wall loosening, extracellular pH plays a regulatory role in plasma membrane transport processes. The trans-membrane proton electrochemical gradient drives plasma membrane transport of inorganic nutrients (Niu et al., 1995). If salt stress induces changes in cell wall acidification rates or cell wall pH value, it might alter rates of membrane transport processes into the growing cell and therefore the availability of nutritional solutes for growth processes.
The possibility that inhibition of leaf cell growth by salt stress is associated with alteration in acidification of the apoplast of growing cells is the subject of this report. We investigated the effect of salt stress on tissue capacity for apoplast acidification in relation to inhibition of relative elemental growth rate (REGR; e.g. growth intensity) along the growing zone. The results provide insights regarding the possible role of proton extrusion response in the process of cell growth inhibition under salt stress.
RESULTS AND DISCUSSION
Effect of Salinity on Leaf Development
Figure 1A shows the results of nondestructive daily leaf length measurements. Salinity (80 mm NaCl) reduced leaf growth and shoot development. The effect of salt stress on leaf growth was apparent from leaf 3 (Fig. 1A). Maximal leaf length attained was reduced 20% and 30% in the third and fourth leaves, respectively, and maximal elongation rate of the fourth leaf was decreased 47% by the stress (Fig. 1B). We opted to use leaf 4 from 14-d-old plants as our experimental system because salt stress effects on the plastochron (the period between emergence of successive leaves) were only apparent from leaf 5 on (data not shown), so that on d 14, control and stress plants could be considered to still be in the same developmental stage. On the other hand, salt stress effects on growth had already become obvious on d 14; the length of leaf 4 was reduced 38% (Fig. 1A) and the elongation rate was reduced 44% (Fig. 1B).
Figure 1.

Effect of salinity on maize leaf growth and shoot development. Plants were grown at 1 mm NaCl (control) or 80 mm NaCl (salt). Data represent results of nondestructive daily leaf length measurements of leaves 1 through 4 (A), where each trace represents the growth of one leaf, and elongation rate (B) of leaf 4 throughout its development, calculated from the leaf length data. Data are means ± se (n = 20). If not shown, error bars are smaller than the symbol size.
The effect of salt stress on maize leaf growth and shoot development is similar to its effect on sorghum (Bernstein et al., 1993a) and lettuce (Lazof et al., 1991). In the three species salinity decreases rates of leaf elongation, maximal leaf length attained, and plastochron duration.
Distribution of Elongation Growth Intensity
We evaluated the effect of NaCl on the spatial distribution of longitudinal expansion along the leaf elongation zone. Salt stress shortened the length of the elongation zone by 25%, from 67 to 50 mm, and reduced the maximal REGR value from 0.061 to 0.040 h−1 (Fig. 2). The extent of growth inhibition varied with spatial location. REGR of the basal 10 mm of the elongation zone was similar in salt and control leaves and in both treatments the zone of maximum REGR was located between 20 and 30 mm from leaf base. In the zone between 20 to 40 mm above the leaf base REGR was greatly reduced by stress.
Figure 2.

Spatial distribution of REGR along the elongation zone of maize leaves growing at 1 mm NaCl (control) or 80 mm NaCl (salt). Data evaluated from prick hole-marking experiments of leaf 4 of maize. Data represent means ± se (n = 10); regression curves are fourth order polynomials, with 95% confidence interval fitted to the original unaveraged segmental R data. The numerical methods are detailed in “Materials and Methods.”
A methodological problem stems from the fact that during prick-hole experiments, leaf elongation is decreased in pricked plants compared with undisturbed controls. In our experiments pricking-induced relative growth reduction was practically identical in unstressed and stressed plants, 48.7% and 47.8% (n = 10), in the control and salt treatments, respectively.
In conclusion, our present data (Fig. 2) support the idea that reduction of REGR in the distal elongation zone is a general characteristic of salinity-induced growth inhibition in grass leaves (Bernstein et al., 1993a, 1993b). Causal investigations into the growth response to salt stress will obviously have to focus on this region. Growing tissue located 20 to 40 mm from the leaf base was chosen as the experimental system for the study of salt stress effect on apoplast acidification. Because this region includes the zone of maximal growth intensity in the control and salt plants and the zone where growth is most affected by stress, changes in cell wall acidification capacity, if they are induced by salt stress and affect cell growth, should be prominent in this location.
Effect of Salt Stress on Excised Leaf Segment Growth
Studies involving estimation of apoplast acidification in this study involved measurements of excised growing segments. Since excision might alter the growth response to stress, proper correlation of growth inhibition with apoplast acidification required analysis of the effect of stress on excised segment growth. The extent of salt stress-induced growth inhibition was not affected by the sectioning (Fig. 3). Similar to the intact tissue on the leaf, growth of the excised segments from salt-stressed plants was inhibited 46.7% in comparison with non-stressed segments (during the 1st h following sectioning). The extent of growth inhibition by salt stress declines with time, but at a slow rate, such that 5 h after sectioning, growth inhibition of the stressed segments was still prominent (42%; 0.52 and 0.3 mm h−1 for control and salt treatments, respectively).
Figure 3.

Elongation rates of excised leaf segments of tissue located 25 to 40 mm from the base of leaf 4. Plants were grown at 1 mm NaCl (control) or 80 mm NaCl (salt). Elongation rate data were calculated from digital images of the segments, taken at 1-h intervals. Data represent means ± se (n = 20).
Effect of Salt Stress on the Kinetics of Apoplast Acidification by Growing Leaf Tissue Segments
Direct measurements of cell wall pH are difficult and not free of artifacts or estimations. Cell wall pH can also be estimated indirectly from measurements of tissue external incubation medium (Cleland, 1973). Such measurements do not provide accurate numerical values of apoplast pH, but they purvey information on the kinetics of apoplast pH changes (Grignon and Sentenac, 1991; Peters et al., 1998). We hereby compared the kinetics of external medium pH changes by control and salinized segments of growing tissue of known local expansion rates and extent of growth reduction under stress.
Excised segments of growing leaf tissue from both salt-stressed and non-stressed plants spontaneously acidified the external medium (Fig. 4). The kinetics of acidification was similar for salt-stressed and non-stressed growing tissues. The pH of the external solution acidified at a nearly constant rate of 0.2 pH units per hour until pH 4.7 was reached after 10 to 11 h. The rate of acidification, as well as the lowest value of pH achieved was similar for both control and salt-stressed growing tissues.
Figure 4.

Effect of salt stress on spontaneous acidification kinetics of external solution by growing tissue segments of maize leaves. Plants were grown at 1 mm NaCl (control) or 80 mm NaCl (salt). pH values, recorded with a data logger, were measured with a pH electrode inserted into 2 mL of aerated solution containing 488 mg of growing leaf tissue (14–20 1-cm segments). The solution contained 0.1, 5, 10, or 25 mm of NaCl (in addition to 1 mm KCl and 0.1 mm CaCl2). The inset figure presents kinetics of pH changes in an aerated solution without the presence of plant tissue (. . . . .), with the presence of control growing tissue (—), and “corrected” pH kinetics, for the control tissue, which takes into account acidification due to aeration (+++). Experiments were conducted at least six times for each treatment; representative traces ± se are shown.
These findings are in accord with in vivo studies demonstrating that under optimal conditions, plasma membrane ATPase activity is not modulated by high salt concentration (Mills and Hodges, 1988; Brüggemann and Janiesch, 1989; Ben-Hayyim and Ran, 1990), but not with studies of the halophyte Atriplex nummularia, or with tomato roots, which demonstrated enhanced H+ transport activity (Niu et al., 1993), or reduced Mg2+-dependent ATPase activity under salinity (Gronwald et al., 1990). Similar to our results for maize leaves, acidification rate in maize roots was not affected by salt stress (Zidan et al., 1990).
Because apoplast acidification requires the presence of CO2 and O2 in the medium (Cerana et al., 1989; Marrè et al., 1989), we chose to aerate the medium with air rather than O2. To exclude the possibility that the kinetics of medium acidification was a result of the continuous bubbling of air through the solution (and consequently high CO2 pressure) rather than of H+ efflux from the tissue, pH changes in an aerated medium were also recorded without the presence of leaf tissue. The contribution of aeration to the acidification kinetics was small and declined with time (Fig. 4, inset). The “corrected” acidification kinetics curve (Fig. 4, inset) from which acidification due to aeration was subtracted was only slightly lower than the “uncorrected” acidification kinetics curve. The corrected pH value after 2 and 8 h of aeration was lower than the uncorrected value by 0.13 and 0.01 pH units, respectively.
In the intact plant, growing cells of the salt-stressed plant are exposed to apoplastic solution of different ionic composition than growing cells of the non-stressed plant. To exclude the possibility that the kinetics of acidification differ when cells are exposed to apoplastic solution of higher Na+ or Cl− concentrations, as is typical of growing tissues under stress, we measured the kinetics of acidification by salt-stressed growing tissue segments bathed in incubation solution of different NaCl concentrations. Twenty-five millimolars Na+ was selected as the highest level of salt to be examined since this concentration was found to correlate with levels of apoplastic Na+ in the stressed growing zone tissue (B.G. Neves-Piestun and N. Bernstein, unpublished data). Bulk tissue Na+ and Cl− concentrations of the leaf growing segment were 40 and 70 mm, respectively (B.G. Neves-Piestun and N. Bernstein, unpublished data). Elevation of NaCl concentration in the external medium from 0.1 to 5, 10, or 25 mm NaCl did not change the kinetics of acidification by the salt-stressed leaf growing tissue (Fig. 4). Similar to acidification by non-stressed growing cells, the acidification rate remained 0.2 pH unit per h and the lowest pH value reached was 4.7 ± 0.3.
In all treatments the spontaneous acidification of the external medium began 2 h after the growing tissue was dissected and infiltrated with the incubation medium. Similar transitional phase for spontaneous acidification of external medium by growing tissue segments was also characteristic for maize roots (Moloney et al., 1981) and maize coleoptiles (Jahn et al., 1996; Peters et al., 1998). This latent period may in part be due to a wounding response following dissection from the plant, or the artificial changes in apoplast characteristics. In all treatments the acidification continued until a steady phase at pH value 4.7 ± 0.3 was reached. If the pH value of this steady phase is endogenously regulated (Peters and Felle, 1991), then the acidification of the external medium can be considered a transient change geared toward restoring the endogenously regulated pH value. This “spontaneous acidification,” although probably different from intact growing tissue behavior, could provide an indication of the tissue tendency to acidify the apoplast and restore its equilibrium pH level under given conditions. In this sense, although a transient phenomenon and a result of breakdown of cellular energetic, it allows comparative estimation of differences in “spontaneous” acidification rates between the control and salt treatments. The similarities in spontaneous acidification rates by control and stressed tissue might hence reflect similarities in endogenous pH regulation. Furthermore, if one accepts the notion that the steady pH level is representative of the tissue endogenously regulated pH level, then our experimental conditions allow comparative estimation of differences in apoplastic pH levels at different treatments. The present results might therefore suggest that the regulated level is unchanged by salt stress. The absolute apoplast pH value however cannot be determined by this method.
Comparison of temporal profiles of spontaneous acidification kinetics (Fig. 4) with temporal profiles of tissue growth rates (Fig. 3) reveals that in both the control and salt treatments, leaf tissue elongation does not correlate with rates of spontaneous apoplast acidification. During the 1st and 2nd h after excision, while the segment elongation growth was maximal, the apoplastic pH first rapidly increased with time and was then maintained briefly at a constant level. After this initial phase, spontaneous acidification proceeded at a steady rate, whereas the segment elongation rate decreased with time. At the end of the steady acidification phase when the pH maintained steady level, tissue growth had ceased all together. Unlike maize coleoptiles in which a temporal correlation between apoplastic acidification kinetics and segment growth was demonstrated (Peters et al., 1998), in maize leaf excised segments, spontaneous acidification responses do not correlate with growth intensity.
Because the kinetics of “spontaneous acidification” by salt-stressed and non-stressed plants were identical (during incubation in 0.1 mm NaCl) and did not temporally correlate with growth intensity, and since a 250-fold increase in NaCl concentration (from 0.1–25 mm) did not alter the kinetics of the “spontaneous acidification,” we conclude that stressed and non-stressed tissue respond similarly to the artificial conditions imposed by our experimental setup (e.g. excision, chemical stimuli resulting from the medium composition being slightly acidic, chemical effect of artificial “apoplast” composition, CO2 and O2 pressures, and illumination). The results suggest that increases in Na+ or Cl− concentration in the leaf apoplast in the range typical of salinization do not affect tissue spontaneous acidification.
Kinetics of Apoplast Acidification by Mature (Nongrowing) Tissue
Unlike growing tissues, mature tissues do not necessarily acidify their external medium. Changes in mature cells apoplast acidification rates or pH values might be tissue specific or location specific and influenced by localized effects or physiological processes (Aloni et al., 1988; Zidan et al., 1990; Kim et al., 1999; Peters and Felle, 1999). Mature, nongrowing cells of our leaf system alkalinized the external medium (Fig. 5A). Similar alkalinization kinetics were typical of both non-stressed and stressed growing cells. The increase in pH does not necessarily reflect a lack of endogenous pH regulation in the mature tissue. The apoplastic pH value of mature tissue might be endogenously regulated to a higher level than was reached after 10 h of measurements. Longer measurement durations or analyses with higher initial pH values are required to resolve this issue. The similarities in alkalinization kinetics by control and stressed tissue suggest that the hypothetical pH regulation in the mature tissue as well is not changed by salt stress.
Figure 5.

Comparative kinetics of pH changes induced by excised segments of mature tissue (A; in vivo experiments), and growing tissue of intact leaves (B; in situ experiments). Plants were grown at 1 mm NaCl (control) or 80 mm NaCl (salt). The mature tissue analyzed was located 150 to 170 mm from the base of leaf 4, the same leaf as used for the growing tissue analysis study. These experiments were replicated five times and representative traces ± se are shown. For the in situ experiments, growing tissue located 25 mm from the leaf base was analyzed. A 0.02-mL drop of a solution was placed on the growing tissue of the leaf; pH changes in the drop were measured with a micro-pH electrode. The inset figure in B presents kinetics of pH changes in a 0.02-mL drop, without the presence of plant tissue. The in situ experiments were conducted five times and representative traces ± se are shown.
Effect of Salt Stress on Apoplast Acidification by Growing Tissues of an Intact Plant
In studies involving excised segments a distinction between effects pertinent to the excision and physiological responses relevant to the intact plant is essential for proper interpretation of experimental results. To ensure that the similarity in acidification kinetics by stressed and non-stressed excised growing segments (Fig. 4) is characteristic of growing tissue of intact plants, we have also followed acidification in situ in a small solution drop placed on the epidermis of an intact leaf while still attached to the plant. The pattern of spontaneous acidification kinetics was similar in the in vivo (excised segments; Fig. 4) and the in situ (intact leaf; Fig. 5B) experiments. In the in situ experiments as well the rate of spontaneous acidification was similar for salt and control treatments (Fig. 5B). This again suggests that the artificial conditions imposed by our experimental setup have similar effects on tissues of both treatments. It also excludes the possibility that altered supply of an organic or inorganic compound from other plant parts to the tissue in question induce differential acidification responses in the two treatments.
Similar to the excised segments experiments in both treatments of the situ experiments, a short phase of alkalinization preceded the spontaneous acidification phase. This suggests that this initial transient phase is not induced by direct damage to the excised tissue, but possibly by the sudden change in the apoplast physical and/or chemical condition. It should be noted that the in situ experiments are also not completely free of wounding effects (removal of leaves 1, 2, and 3). We cannot exclude the possibility that the incisions at the older leaf bases imposed some effect, possibly even hormonal, on the growing tissue of leaf 4.
The length of time required for establishment of steady pH level (4 min) was much shorter, and the resulting acidification rate (2.8 pH units per hour) was 14 times higher in the intact plant experiments, probably as a result of the smaller ratio of external solution to growing tissue. In accordance with this, the acidification rate of the excised segments was about 7% of the intact plant acidification rate, and the ratio of tissue surface area to “extended apoplast” volume at the excised segment experiments was similarly about 10% of the intact plants value (0.00023 and 0.002 mL mm−2 at the excised and intact tissue experiments, respectively).
The medium pH reached a steady level in the intact plants as well, supporting the notion that the “steady level” obtained in the excised segments experiments is not an artifact resulting from segment excision, but rather a representation of endogenous regulation of apoplast pH.
The kinetics of pH change by mature tissue of intact plants was also similar in stressed and non-stressed plants (data not shown). Similar to the excised segments experiments, a continuous alkalinization of the “extended apoplast” (constant rate of 2.7 pH units per hour) was observed.
Effect of Fusicoccin (FC)
Acidification of the medium suggests proton extrusion. To exclude the possibility that external medium acidification was due to nonspecific leakage of acidic compounds from compartments such as the vacuole rather than proton extrusion, FC, a powerful enhancer of the plasmalemma H+-ATPase activity, was added to the external solution. It was added to the incubation medium 5 h after segment excision when the acidification rate was already established at 0.2 pH units per h (Fig. 4) and stress-induced inhibition of segment growth was 42% (Fig. 3). At this time, excised segment elongation rate was still rapid (0.52 and 0.3 mm h−1 for non-stressed and stressed segments, respectively; Fig. 3) and acidification due to aeration was minimal (0.001 pH units h−1; Fig. 4, inset). FC stimulated the elongation rate of the excised tissue segments (Table I). The elongation rate of the non-stressed and stressed segments increased 314% and 244%, respectively. Similar growth-stimulating effects of FC on dicotyledonous and monocotyledonous tissues have been reported by numerous authors (including Schopfer, 1993; Jahn et al., 1996).
Table I.
Effect of FC on growth of excised segments
| Elongation Rate of Excised Segments
| ||
|---|---|---|
| Treatment | Control | Salt |
| mm h−1 | ||
| − FC | 0.360 ± 0.050 | 0.250 ± 0.040 |
| + FC | 1.490 ± 0.019 | 0.860 ± 0.016 |
Elongation rates of segments excised from non-stressed (control) and salt-stressed (salt) plants were analyzed with or without the presence of FC in the incubation medium. Elongation rates were evaluated from digital images of the segments taken at the time of FC application and 1 h thereafter. Data are means ± se (n = 10 segments).
FC caused an immediate, dramatic decrease in medium pH (Fig. 6). Following its application the acidification rate increased from 0.2 to 1.5 pH units per hour in both control and salt-stressed segments. About 0.8 h after FC application, the acidification rate decreased and proceeded in progressively lower rates until pH 3.6 ± 0.3 was reached 2 h after FC application. Acidification kinetics for both treatments were identical.
Figure 6.

Effect of 0.01 μm FC on the acidification kinetics of external solution by growing tissue segments of maize leaf. Plants were grown at 1 mm NaCl (control) or 80 mm NaCl (salt). The arrow indicates the timing of FC application. The experiment was conducted five times, and representative traces ± se are shown.
Similar effects of FC on medium pH were reported for several growing plant tissues including maize coleoptiles (Peters and Felle, 1991) and maize roots (Felle, 1998). The FC-induced pH decrease in the medium in which growing segments are incubated is conventionally interpreted as an FC-stimulated, energy-linked H+ extrusion by plasmalemma ATPase (Marrè, 1979; Schopfer, 1993). Considering that in our system, enhancement of acidification was observed in the presence of FC, the pH changes of the external medium reported in Figure 4 suggest an association with proton pumping into the cell wall space, and not a nonspecific cellular leakage.
It is important to note that the enhanced rate of acidification induced by FC is indicative of the tissue's inherent potential for acidification. The strong biochemical stimulation of the plasmalemma H+-ATPase activity by FC allows a rough comparative estimate of active ATPase density in the tissue. Dissimilarities in growing tissue responses to FC might suggest differences in functional ATPase density. The FC-induced acidification of the external medium by salt-stressed and non-stressed growing tissue was found to follow similar kinetics, suggesting that active ATPase density in the tissue was not altered by salt stress (Fig. 6).
That the potential of the growing tissue to acidify the apoplast did not change under the same salt stress conditions that induced inhibition of tissue elongation rate suggests that salinity does not inhibit cell growth by impairing the acidification process or reducing the capacity for cell wall acidification.
In summary, our results suggest that salt-stressed and non-stressed growing cells regulate their apoplast pH endogenously and that salt stress does not alter the ability of the growing tissue to regulate its external medium pH.
Taking into account two observations: (a) that similar rates of spontaneous acidification were observed in stressed and non-stressed segments differing in their elongation rate (Figs. 3 and 4), and (b) that the capacity of the growing tissue to acidify the apoplast did not change under the same salt-stress conditions that induced inhibition of tissue elongation rate (FC experiments; Fig. 6; Table I), we conclude that salinity does not inhibit maize leaf cell growth by impairing the acidification process or reducing the inherent capacity for cell wall acidification.
MATERIALS AND METHODS
Plant Material
Seeds of maize (Zea mays cv G.S. 46, Galilee Seeds, Haifa, Israel) were soaked in aerated solution (2 mm KCl and 1 mm CaCl2) for 6 h and were later sown on moist vermiculite in plastic boxes. The vermiculite was pretreated with 15 mm Ca(NO3)2 for 2 h, rinsed twice, and was later soaked in 0.1 concentration-modified Hoagland solution (Epstein, 1972) for 2 h prior to sowing. The boxes were covered and kept in the dark at 25°C until d 4 when illumination started (400 μE s−1 m−2, 16-h photoperiod, relative humidity of 60% and 80% during the day and night, respectively). On d 7, plants with similar lengths of leaf 1 and 2 (80 ± 10 mm) were selected and transferred to aerated one-quarter-strength modified Hoagland solution (Bernstein et al., 1995). Micronutrients were supplied as in one-half Hoagland concentration, except that iron was added as 50 μm Fe-EDTA and 20 μm Fe(NH4SO4)2 and Na level was elevated to 1 mm. Solution pH was adjusted to 5.7 with addition of KOH. Growth chamber conditions remained as described above.
Salinization began with the transfer to hydroponics on d 7. At this time leaf 3 was not yet visible and leaf 4 was shorter than 1 cm. NaCl concentration in the medium was elevated in three daily steps (to 20, 50, and finally 80 mm). Control plants remained at a total concentration of sodium and chloride of 1 mm each.
Growth Measurements
Daily leaf length measurements were used for evaluation of shoot growth, calculations of leaf elongation rate, and selection of leaf insertion number for this study. Leaf length was measured daily with a ruler to the nearest 0.5 mm from the base of the plant to the tip of the leaf (Bernstein et al., 1993b). Results from 20 replicated leaves were averaged.
Spatial distribution of elongation was calculated from short-term displacements of prick-holes placed along the growing zone of leaf 4, the selected leaf for analysis in this study. Leaves were marked as described previously (Bernstein et al., 1993b), except that initial distance between marks was 5 mm (W.S. Peters and N. Bernstein, unpublished data). In short, 4 h after the beginning of the light period, on d 14, the base of the shoot was pricked with a series of 20 fine insect-mounting pins (size 00; Hamilton Bell Co., Montvale, NJ) fixed at roughly 5 mm distance on a Plexiglas holder. The actual distance between pins was measured to the nearest 12 μm under a dissection microscope before and after each experiment. The pins were removed and after growth had continued for another 5 h, leaf 4 was carefully freed from older enclosing leaf sheaths. The final positions of the pin marks were measured to the nearest 12 μm. Experiments were performed four times for salinized and unstressed plants each, with 12 pricked and 10 unpricked control plants (to allow assessment of pricking-induced growth inhibition) at every occasion. Pricked leaves were included in the analysis only if the first prick hole was located less than 1.5 mm above the leaf base.
Segmental relative growth rate (R) was calculated, as of Equation 1, where L0 and Lt denote initial and final distances between two marks, respectively, and t the duration of the experiment (Green, 1976).
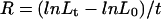 |
1 |
Profiles of REGR were then created by plotting segmental R versus average segment position (Peters and Bernstein, 1997) and fitting polynomials (fourth order) to the data points. In addition, segmental R were averaged for parallel segments on replicated leaves, and plotted (±se) versus average segment position. Results from both methods yielded similar growth profiles and are presented in Figure 2.
Elongation rates of 15 mm-long excised segments located 25 to 40 mm from the base of leaf 4 were analyzed. The segments were cut from each replicated leaf 4 h after the transition to the light period, 7 d after commencement of salinization (14 d after germination). The excised segments were placed in vials containing at least 1.125 mL of incubation medium (1 mm KCl, 0.1 mm CaCl2, and 0.1 mm NaCl at pH 5.5, according to Peters and Felle, 1991, with modifications) and lightly vacuum infiltrated. The solution was constantly bubbled with air. The vials were kept in a growth chamber under constant light conditions at 25°C. Digital images of the segments were captured every hour with a digital camera (MSV-800, Applitec, Israel). Segment length was measured from the digital images and segment elongation rate was numerically calculated. Results from 20 replicated segments were averaged for each treatment.
In some experiments FC, a phytotoxin known as a “super-auxin” because of its effect on growth (Cleland, 1976), was added to the segment incubation medium to a final concentration of 0.01 μm (in 0.0001% [w/v] ethanol). FC was therefore prepared as an aqueous 1 μm stock solution (pH 6.24) by adding 0.250 mg of FC previously dissolved in 0.1 mL of ethanol to the appropriate volume of deionized water at 75°C. The stock was stored for a maximum period of 6 h at 4°C prior to application.
Apoplastic pH Measurements
Excised leaf segments, as well as intact plants, were used for estimation of apoplastic acidification. Growing segments were sampled from leaves of same age, developmental stage, and insertion number as the ones sampled for segment elongation rate analyses. A 20-mm-long leaf segment (located 20–40 mm from the leaf base) was cut from the growing zone of leaf 4 of stressed and non-stressed plants. The tissue was cut along the mid-vein and again perpendicular to the vein into two 1-cm sections and placed base down in a 6-mL transparent vial. Sections from seven to 10 replicated leaves were placed in 3 mL of aerated solution and lightly vacuum infiltrated. As in the excised segments elongation rate experiments, aeration rate was 15 to 18 mL min−1, and the same ratio of tissue weight/solution volume (244 mg mL−1) was used in both the control and the salt treatments to ensure that changes in acidification kinetics would not result from differences in cell wall biomass per sample. Cell wall volume fraction per tissue is similar in the appropriate control and salt-stress cells (B.G. Neves-Piestun and N. Bernstein, unpublished data). The vial, covered with parafilm, was kept in a growth chamber. In some experiments higher Na+ concentrations were used, Na+ levels were then elevated from 0.1 to 5, 10, and 25 mm by the addition of NaCl. The solution pH was constantly recorded with a Russell CW711 combination pH electrode with a Mettler 340 pH meter connected to a data logger CR10X (Campbell Scientific Inc., Logan, UT). pH changes were also recorded in an aerated incubation solution without the presence of the tissue segments to estimate the extent of CO2-induced medium acidification. The initial pH of the incubation solution (which might affect spontaneous acidification; Aloni et al., 1988) was adjusted to 5.5, a stimulating pH level for ATPase activity (Ballesteros et al., 1998).
pH changes were also recorded in the presence of FC in the incubation solution. In addition to its effect on growth, FC is also known to strongly stimulate plasma membrane ATPase activity (Malek and Baker, 1977) and therefore, apoplastic acidification. FC was added to the incubation medium according to the procedure described above.
In other experiments the kinetics of medium acidification by mature, nongrowing leaf tissue was followed as well. Mature tissue, located 150 to 170 mm from the base of the same leaf used for growing tissue analysis, was excised and prepared as described for growing tissue. The ratio between the tissue and the solution volume was kept similar to the growing tissue experiments.
Acidification of external medium by growing and mature tissue of intact plants (in situ measurements) was investigated, as well, to evaluate possible wounding stress interaction in the kinetics of medium acidification. For growing tissue analysis, leaves 1 to 3 were removed from the shoot of the intact plant and the growing region of leaf 4 was thus exposed. The excision marks were covered with paraffin, and the exposed growing zone was protected with parafilm to create a humid chamber. A 0.02-mL drop of a solution containing 1 mm KCl, 0.1 mm CaCl2 and 0.1 mm NaCl at pH 6.1 was placed on the growing tissue of the leaf, 25 mm from its base, in the same zone used for excised segments study. For mature tissue analysis the drop was placed 160 mm from the base of leaf 4, the middle of the segment used for excised segments mature tissue experiments. A PHR-146 micro-combination pH electrode (Lazar Research Laboratories, Los Angeles) was lowered into the drop and the pH of the solution was continuously recorded as described above. pH changes were also recorded in a solution drop of same volume placed on the surface of a glass Petri dish, rather than on a growing leaf tissue.
ACKNOWLEDGMENTS
We thank Dr. W.S. Peters for comments on early versions of the manuscript. The seeds used in this project were a gift from Galilee-seeds, Israel.
Footnotes
This work was supported by the U.S.-Israel Binational Agriculture Research and Development Fund (grant no. 2360–93). This paper is contribution no. 610/00 from the Volcani Center, Bet-Dagan, Israel.
LITERATURE CITED
- Aloni B, Daie J, Wyse RE. Regulation of apoplastic pH in source leaves of Vicia faba by gibberellic acid. Plant Physiol. 1988;88:367–369. doi: 10.1104/pp.88.2.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballesteros E, Kerkeb B, Donaire JP, Belver A. Effects of salt stress on H+-ATPase activity of plasma membrane-enriched vesicles isolated from sunflower roots. Plant Sci. 1998;134:181–190. [Google Scholar]
- Ben-Hayyim G, Ran U. Salt-induced cooperativity in ATPase activity of plasma membrane-enriched fractions from cultured Citrus cells: kinetic evidence. Physiol Plant. 1990;80:210–216. [Google Scholar]
- Bernstein N, Laüchli A, Silk WK. Kinematics and dynamics of sorghum (Sorghum bicolor L.) leaf development at various Na/Ca salinities: I. Elongation growth. Plant Physiol. 1993a;103:1107–1114. doi: 10.1104/pp.103.4.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernstein N, Silk WK, Laüchli A. Growth and development of sorghum leaves under conditions of NaCl stress: spatial and temporal aspects of leaf growth inhibition. Planta. 1993b;191:433–439. [Google Scholar]
- Bernstein N, Silk WK, Läuchli A. Growth and development of sorghum leaves under conditions of NaCl stress: possible role of some mineral elements in growth inhibition. Planta. 1995;196:699–705. [Google Scholar]
- Brüggemann W, Janiesch P. Comparison of plasma membrane ATPase from salt-treated and salt-free grown Plantago maritima L. J Plant Physiol. 1989;140:20–25. [Google Scholar]
- Cerana R, Patti V, Lado P. Changes in ethanol, lactate and malate content in Acer pseudoplatanus cells: effects of fusicoccin and O2 availability. Physiol Plant. 1989;75:183–187. [Google Scholar]
- Cleland RE. Auxin-induced hydrogen ion excretion from Avena coleoptiles. Proc Natl Acad Sci USA. 1973;70:3092–3093. doi: 10.1073/pnas.70.11.3092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleland RE. Kinetics of hormone induced H+ secretion. Plant Physiol. 1976;58:210–213. doi: 10.1104/pp.58.2.210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosgrove DJ. Relaxation in a high-stress environment: the molecular bases of extensible walls and cell enlargement. Plant Cell. 1997;9:1031–1041. doi: 10.1105/tpc.9.7.1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer GR, Bowman DC. Kinetics of maize leaf elongation: I. Increased yield threshold limits short-term steady-state elongation rates after exposure to salinity. J Exp Bot. 1991;42:1417–1426. [Google Scholar]
- Epstein E. Mineral Nutrition of Plants: Principles and Perspectives. New York: John Wiley & Sons; 1972. [Google Scholar]
- Felle HH. The apoplastic pH of the Zea mays roots cortex as measured with pH-sensitive microelectrodes: aspects of regulation. J Exp Bot. 1998;49:987–995. [Google Scholar]
- Flowers TJ, Hajibagheri MA, Yeo AR. Ion accumulation in the cell walls of rice plants growing under saline conditions: evidence for the Oertli hypothesis. Plant Cell Environ. 1991;14:319–325. [Google Scholar]
- Green PB. Growth and cell pattern formation on an axis: critique of concepts, terminology, and modes of study. Bot Gaz. 1976;137:187–202. [Google Scholar]
- Grignon C, Sentenac H. pH and ionic conditions in the apoplast. Annu Rev Plant Physiol Plant Mol Biol. 1991;42:103–28. [Google Scholar]
- Gronwald JW, Suhayda CG, Tal M, Shanon MC. Reduction in plasma membrane ATPase activity of tomato roots by salt stress. Plant Sci. 1990;66:145–153. [Google Scholar]
- Hajibagheri MA, Harvey DMR, Flowers T. Quantitative ion distribution within root cells of salt-sensitive and salt-tolerant maize varieties. New Phytol. 1987;105:367–379. doi: 10.1111/j.1469-8137.1987.tb00874.x. [DOI] [PubMed] [Google Scholar]
- Hartung W, Radin JW, Hendrix DL. Abscisic acid movement into the apoplastic solution of water-stressed cotton leaves. Plant Physiol. 1988;86:908–913. doi: 10.1104/pp.86.3.908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jahn T, Johansson F, Lüthen H, Volkmann D, Larsson C. Reinvestigation of auxin and fusicoccin stimulation of the plasma-membrane H+-ATPase activity. Planta. 1996;199:359–365. [Google Scholar]
- Kim TK, Silk WK, Cheer AY. A mathematical model for pH patterns in the rhizospheres of growth zones. Plant Cell Environ. 1999;22:1527–1538. [Google Scholar]
- Kumon K, Suda S. Changes in the extracellular pH of the motor cells of Mimosa pudica L. during movement. Plant Cell Physiol. 1985;26:375–377. [Google Scholar]
- Lazof D, Bernstein N. The NaCl-induced inhibition of shoot growth: the case for disturbed nutrition with special consideration of calcium nutrition. Adv Bot Res. 1998;29:113–189. [Google Scholar]
- Lazof D, Bernstein N, Laüchli A. Growth and development of the Lactuca sativa shoot as affected by NaCl stress: consideration of leaf developmental stages. Bot Gaz. 1991;152:72–76. [Google Scholar]
- Malek T, Baker DA. Effect of fusicoccin on proton cotransport of sugars and the phloem loading of Ricinus communis L. Plant Sci Lett. 1977;11:233–239. [Google Scholar]
- Marrè E. Fusicoccin: a tool on plant physiology. Annu Rev Plant Physiol. 1979;30:273–288. [Google Scholar]
- Marrè MT, Albergoni FG, Moroni A, Marrè E. Light-induced activation of electrogenic H+ extrusion and K+ uptake in Elodea densa depends on photosynthesis and is mediated by the plasma membrane H+ ATPase. J Exp Bot. 1989;40:343–352. [Google Scholar]
- McQueen-Mason SJ, Fry SC, Durachko DM, Cosgrove DJ. The relationship between xyloglucan endotransglycosylase and in vitro cell wall extension in cucumber hypocotyls. Planta. 1993;190:327–331. doi: 10.1007/BF00196961. [DOI] [PubMed] [Google Scholar]
- Mills D, Hodges TK. Characterization of plasma membrane ATPase from roots of Atriplex nummularia. J Plant Physiol. 1988;132:513–519. [Google Scholar]
- Moloney MM, Elliott MC, Cleland RE. Acid growth effects in maize roots: evidence for a link between auxin-economy and proton extrusion in the control of root growth. Planta. 1981;152:285–291. doi: 10.1007/BF00388251. [DOI] [PubMed] [Google Scholar]
- Mulkey TJ, Evans ML. Geotropism in corn roots: evidence for its mediation by differential acid efflux. Science. 1981;212:70–71. doi: 10.1126/science.212.4490.70. [DOI] [PubMed] [Google Scholar]
- Munns R. Physiological processes limiting plant growth in saline soils: some dogmas and hypotheses. Plant Cell Environ. 1993;16:15–24. [Google Scholar]
- Neumann PM, Azaizeh H, Leon D. Hardening of roots cell walls: a growth inhibitory response to salinity stress. Plant Cell Environ. 1994;17:303–309. [Google Scholar]
- Newman IA, Kochian LV, Grusak MA, Lucas W. Fluxes of H+ and K+ in corn roots. Plant Physiol. 1987;84:1177–1184. doi: 10.1104/pp.84.4.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu X, Bressan RA, Hasegawa PM, Pardo JM. Ion homeostasis in NaCl stress environments. Plant Physiol. 1995;109:735–742. doi: 10.1104/pp.109.3.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu X, Narasimhan ML, Salzman RA, Bressan RA, Hasegawa PM. NaCl regulation of plasma membrane H+-ATPase gene expression in a glycophyte and a halophyte. Plant Physiol. 1993;103:713–718. doi: 10.1104/pp.103.3.713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters WS, Bernstein N. The determination of relative elemental growth rate profiles from segmental growth rates: a methodological evaluation. Plant Physiol. 1997;113:1395–1404. doi: 10.1104/pp.113.4.1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters WS, Felle H. Control of Apoplast pH in corn coleoptile segments: I. The endogenous regulation of cell wall pH. J Plant Physiol. 1991;137:655–661. [Google Scholar]
- Peters WS, Felle H. The correlation of profiles of surface pH and elongation growth in maize roots. Plant Physiol. 1999;121:905–912. doi: 10.1104/pp.121.3.905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters WS, Luthen H, Böttger M, Felle H. The temporal correlation of changes in apoplast pH and growth rate in maize coleoptile segments. Aust J Plant Physiol. 1998;25:21–25. [Google Scholar]
- Rayle DL, Cleland RE. The acid growth theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992;99:1271–1274. doi: 10.1104/pp.99.4.1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schopfer P. Determination of auxin-dependent pH changes in coleoptile cell walls by a Null-Point Method. Plant Physiol. 1993;103:351–357. doi: 10.1104/pp.103.2.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stahlberg R, Van Volkenburgh E. The effect of light on membrane potential, apoplastic pH and cell expansion in leaves of Pisum sativum L. var. Argenteum Planta. 1999;208:188–195. [Google Scholar]
- Tomos D, Pritchard J. Biophysical and biochemical control of cell expansion in roots and leaves. J Exp Bot. 1994;45:1721–1731. [Google Scholar]
- Van Volkenburgh E, Boyer JS. Inhibitory effects of water deficit on maize leaf elongation. Plant Physiol. 1985;77:190–194. doi: 10.1104/pp.77.1.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson C, Shannon MC. Salt-induced Na+/H+ antiport in root plasma membrane of a glycophytic and halophytic species of tomato. Plant Sci. 1995;107:147–157. [Google Scholar]
- Zidan I, Azaizeh H, Neumann PM. Does salinity reduce growth in maize root epidermal cells by inhibiting their capacity for cell wall acidification? Plant Physiol. 1990;93:7–11. doi: 10.1104/pp.93.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]