

Abstract
Recent studies indicate that regulation of the actin cytoskeleton is important for protein trafficking, but its precise role is unclear. We have characterized the ARF1-dependent assembly of actin on the Golgi apparatus. Actin recruitment involves Cdc42/Rac and requires the activation of the Arp2/3 complex. Although the actin-binding proteins mAbp1 (SH3p7) and drebrin share sequence homology, they are differentially segregated into two distinct ARF-dependent actin complexes. The binding of Cdc42 and mAbp1, which localize to the Golgi apparatus, but not drebrin, is blocked by occupation of the p23 cargo-protein-binding site on coatomer. Exogenously expressed mAbp1 is mislocalized and inhibits Golgi transport in whole cells. The ability of ARF, vesicle-coat proteins, and cargo to direct the assembly of cytoskeletal structures helps explain how only a handful of vesicle types can mediate the numerous trafficking steps in the cell.
INTRODUCTION
A rapidly growing body of evidence shows that the cytoskeleton is important for the regulation and organization of both the exocytic and endocytic functions of the secretory pathway. For example, recent evidence indicates roles for the actin cytoskeleton during clathrin-mediated endocytosis (Qualmann et al., 2000) and for transport at the Golgi apparatus. An actin/spectrin/ankyrin cytoskeleton has been implicated in protein transport to and from the Golgi (De Matteis and Morrow, 2000). Most importantly, when actin is disrupted with the toxin cytochalasin B, protein transport through the Golgi apparatus is inhibited (Hirschberg et al., 1998). Similarly, when spectrin binding to the Golgi is blocked (Godi et al., 1998), or when the function of the Rho-family GTP-binding protein, Cdc42—a regulator of actin rearrangement—is disrupted (Wu et al. 2000), protein transport from the ER to the Golgi is inhibited. In addition to the defects in the early secretory pathway, Cdc42 function is important for late Golgi protein transport and sorting in polarized epithelial cells (Kroschewski et al. 1999; Müsch et al., 2001).
Consistent with the defects in protein transport described above are biochemical and immunoelectron microscopy studies showing that actin, actin-binding proteins, and myosin are bound to Golgi membranes and Golgi-derived vesicles (Heimann et al., 1999; De Matteis and Morrow, 2000; Valderrama et al., 2000). A study showing that disrupting actin has effects on Golgi morphology also provides good evidence that the actin cytoskeleton functions at the Golgi (Valderrama et al., 1998). Together, the above studies suggest an important role for the actin cytoskeleton and signaling pathways that affect actin in protein trafficking to and from the Golgi apparatus. Despite this progress, the precise role that actin plays in protein trafficking at the Golgi remains to be determined.
One clue to the role of actin at the Golgi apparatus is the finding that ADP-ribosylation factor (ARF), the GTP-binding protein that plays an essential role in regulating the assembly of transport vesicles (Mellman and Warren, 2000), also regulates the actin cytoskeleton. At the plasma membrane, ARF6 regulates clathrin/AP2-independent endocytosis and the assembly of filopodia and cortical actin (Radhakrishna et al., 1999; Boshans et al., 2000; Mostov et al., 2000). At the Golgi apparatus, ARF1 has been shown to regulate the binding of actin and spectrin to the membrane (Godi et al., 1998; Fucini et al., 2000). We have shown previously that at least two pools of actin with distinct properties are assembled on Golgi membranes upon ARF activation (Fucini et al., 2000). Phosphatidylinositol (PI)-4-kinase, PI-5-kinase, and Rho family GTP-binding proteins have been implicated as effector proteins in ARF-mediated regulation of the actin cytoskeleton (D'Souza-Schorey et al., 1997; Godi et al., 1999; Honda et al., 1999; Boshans et al., 2000).
Besides the clear role for ARF1 in Golgi function, numerous studies indicate that the Golgi apparatus is a site for Cdc42 function. Cdc42, and components of Cdc42 signaling pathways are localized to the Golgi in several cell types (Erickson et al., 1996; McCallum et al., 1998; Kroschewski et al. 1999). Studies using whole cells indicate that Cdc42 levels on the Golgi are sensitive both to brefeldin A (BFA), an inhibitor of ARF activation, and to the expression of mutant ARF isoforms (Erickson et al., 1996). Importantly, a recent study shows that Cdc42 interacts with the γ-COP subunit of coatomer, the major coat component of Golgi-derived COPI transport vesicles (Wu et al., 2000). This interaction is implicated in regulating protein trafficking and in the ability of a mutant form of Cdc42 to transform cells.
To help elucidate the role of actin in Golgi function and vesicular transport, we have further characterized the ARF-dependent actin assembly at the Golgi. We show that interactions between proteins involved in transport vesicle assembly and proteins involved in Cdc42-dependent actin signaling function to assemble specific actin structures on the Golgi. Our data indicate a role for actin early in vesicle assembly.
MATERIALS AND METHODS
Reagents
Rat-liver Golgi membranes and bovine brain cytosol were isolated as described previously (Malhotra et al.,1989). Recombinant myristoylated ARF1 was expressed in Escherichia coli and purified using DEAE-sepharose as described previously (Helms et al., 1993). Coatomer was purified as described by Waters et al. (1992). Clostridium difficile toxin B (Tech Lab Inc., Blacksburg, VA), C. botulinum exoenzyme C3 (Cytoskeleton, Denver, CO), and BFA (Calbiochem, La Jolla, CA) were obtained commercially. The following antibodies were used in these studies: antiactin (Sigma, St. Louis, MO), anti–β-COP (Sigma), anti-Cdc42 (Zymed Laboratories Inc., South San Francisco, CA), anti-Rac (Cytoskeleton), anti-Rho (Cytoskeleton), anti-drebrin (MBL), anti–α-mannosidase II (Covance Research Products, Denver, PA).
Expression of WASP-CA and N-WASP-CA GST Fusion Proteins
The glutathione S-transferase (GST)-fusion proteins containing the C-terminal CA regions of human WASP or rat N-WASP were obtained by expression in E. coli as described previously (Miki et al., 1996; Rohatgi et al., 1999). Briefly, cDNA fragments encoding the amino acids 449–505 of human WASP and amino acids 450–501 of rat N-WASP were amplified by PCR using synthetic oligonucleotide primers and then inserted into the BamHI/EcoRI sites of pGEX 4T-2 (Amersham-Pharmacia Biotechnology, Piscataway, NJ). The recombinant plasmids were expressed in E. coli and purified from lysates by elution from glutathione-sepharose beads (Amersham-Pharmacia Biotechnology) according to the manufacturer's instructions. For control incubations, GST alone was expressed from the pGEX 4T-2 plasmid and purified exactly like the fusion proteins.
Generating Anti-mAbp1 Antibodies
A rat EST clone representing the C-terminal 30 amino acids (aa 406–436) of Abp1 was kindly provided by Dr. M. B. Soares (GenBank accession no. AA859856). This amino acid sequence is located in the putative SH3 domain and is identical to the mAbp1 C terminus (Kessels et al., 2000). PCR-amplified products of this sequence were ligated into the pGex4T-2 vector (Amersham-Pharmacia), and the GST-fusion protein was expressed in E. coli and purified as mentioned above.
To raise polyclonal antibodies against mAbp1 (aa 406–436), the GST-fusion protein (500 μg) was mixed 1:1 with complete Freund's adjuvant (Difco Labs, Detroit, MI) and injected into rabbits. After an initial 3-week interval, rabbits were boosted using incomplete adjuvant every 2 weeks. Rabbits were bled, and positive sera were identified by immunoblot analysis of a dilution series of purified GST-mAbp1 (aa 406–436), bovine brain cytosol, and a phalloidin precipitate of bovine brain cytosol. Anti-mAbp1 antibodies were affinity purified by binding to recombinant mAbp1 immobilized on nitrocellulose and eluting with 100 mM glycine buffer, pH 2.5. Anti-GST antibodies were subtracted from the serum by passing it through GST-coupled beads.
Golgi-binding Reactions
One- or two-stage Golgi-binding assays were carried out and membranes were reisolated as described previously (Fucini et al., 2000), except that cytosol was omitted from the first stage of the two-stage reactions and replaced by recombinant ARF1 and/or BFA as indicated in the figures. The final reaction volume was 0.5 ml. The final GTPγS concentration was 20 μM when included. For the experiments using BFA, both the membranes and the cytosol for the second-stage reaction were incubated with 400 μM BFA or with the methanol solvent (2% final) as a control. Reactions containing toxin B also included 100 μM UDP-glucose as a substrate for glucosylation and 100 μM NAD was included in the incubations with C3 exoenzyme as a substrate for ADP-ribosylation. For the extraction of actin with high salt, the Golgi membranes were washed with 250 mM potassium chloride and reisolated by centrifugation as described previously for the extraction of COPI-coated vesicles (Fucini et al., 2000). Coatomer-depleted cytosol was prepared by fractionating the bovine brain cytosol using a Sephacryl S-200 column. The column fractions were analyzed by Western blotting with anti-Cdc42 and anti–β-COP antibodies. The fractions devoid of coatomer were pooled and concentrated using a Centricon filtration device (Millipore, Bedford, MA).
Western Blotting
Proteins were fractionated using SDS-PAGE and blotted onto PVDF membranes using standard protocol for the Bio-Rad minigel and blotting apparatuses. After the transfer, the membranes were incubated with appropriate dilutions of the indicated primary antibodies. The signal was visualized using HRP-conjugated secondary antibodies (Bio-Rad, Hercules, CA) and ECL (Amersham-Pharmacia). Where indicated, the signals were quantitated using densitometry.
Immunofluorescence
NRK cells were plated onto coverslips, and after 24 h the cells were washed with phosphate-buffered saline (PBS), fixed with 4% formaldehyde, and permeabilized using 0.1% Triton X-100 for 4 min at room temperature. The formaldehyde was quenched with 50 mM ammonium chloride for 10 min at room temperature. The cells were washed three times with PBS and blocked with 2.5% donkey serum in PBS for 2 h at 4°C. Appropriate dilutions of the anti-mAbp1, anti–mannosidase II, and anti–β-COP antibodies in the blocking buffer were added to the cells for 2 h at 4°C. The cells were washed three times with PBS and FITC-conjugated anti-mouse and Texas-Red–conjugated anti-rabbit secondary antibodies were added in blocking buffer for 1 h. The cells were washed three times, mounted on slides, and analyzed on a confocal microscope (Bio-Rad).
VSV G Protein Transport Assay
The cell line Gts-NRK (a gift from Dr. V. Malhotra) that stably expresses the temperature-sensitive ts045-VSVG was maintained by growth at the permissive temperature 32°C in α-MEM plus 5% fetal calf serum. The plasmid for the expression of myc-tagged, full-length mAbp1 was constructed using pRK5 as described in Kessels et al. (2001). The plasmid pEGFP-C3 (CLONTECH, Palo Alto, CA) was used for the expression of GFP in controls. For the assays, cells were grown on coverslips and transfected with the mAbp1 expression plasmids using Lipofectin reagent (Life Technologies, Rockville, MD). Transfected cells were incubated for 14–16 h at the restrictive temperature (39.5°C) to accumulate VSV G protein in the ER. VSV G protein was released from the temperature block by switching the cells to 32°C media containing 10 μg/ml cycloheximide. The cells were incubated for 15 min at 32°C and processed for immunofluorescence. VSV G protein was visualized using the mAb P5D4 and the myc-tagged mAbp1 proteins were detected using a chicken polyclonal anti-myc antibody (Aves Labs, Tygard, OR).
RESULTS
ARF Activation Is Necessary but not Sufficient for Actin Binding to Golgi Membranes
Using a cell-free binding assay, we showed previously that actin assembles in a GTPγS-dependent and BFA-sensitive manner on Golgi membranes (Fucini et al., 2000). To prove that ARF is involved in actin recruitment to the Golgi and to provide a system for the further dissection of signaling events involved in GTPγS-dependent actin assembly on the Golgi, we investigated whether this process could be reconstituted with recombinant myristoylated ARF1.
If the BFA effects on Golgi actin assembly are mediated through inhibiting the nucleotide exchange activity for ARF, then binding activated ARF to the membrane before the addition of BFA is predicted to prevent the inhibitory effects of this toxin. Therefore, two-stage Golgi-binding assays were carried out to examine the effects of purified ARF1 and BFA on Golgi actin assembly (Figure 1). In the first stage, Golgi membranes were pretreated with GTPγS alone or GTPγS plus ARF1 and then incubated with BFA or a solvent control. The Golgi membranes were reisolated and incubated in a second-stage reaction that contained cytosol and GTPγS. BFA inhibited both actin assembly and the recruitment of coatomer during the second-stage incubation when recombinant ARF1 was omitted from the first-stage incubation (Figure 1A, lanes 1 and 2). By contrast, when activated ARF1 was preloaded onto the membrane, BFA had no effect on actin levels or on coatomer binding (lanes 3 and 4). This result shows that the effects of BFA on both coat binding and actin assembly are mediated by inhibiting ARF activation. Thus, activated ARF is clearly necessary for actin assembly on the Golgi membranes.
Figure 1.
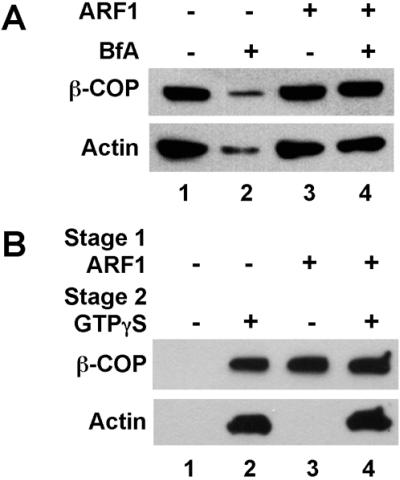
ARF1 activation is necessary but not sufficient for actin binding to the Golgi. (A) Western blot analysis of two-stage Golgi-binding assays reconstituted with GTPγS alone (lanes 1 and 2) or GTPγS plus recombinant myristoylated ARF1 (0.03 mg/ml; lanes 3 and 4). After a 10-min preincubation to allow the ARF1 to bind, brefeldin A (BfA; lanes 2 and 4) or methanol carrier (lanes 1 and 3) was added and the first-stage incubation was continued for an additional 10 min. The membranes were reisolated and added to a second-stage incubation with cytosol, GTPγS, and either BFA (lanes 2 and 4) or the methanol carrier (lanes 1 and 3). After the second-stage incubation, the membranes were isolated by flotation and the levels of bound actin and β-COP were determined. (B) Western blot probed with anti-actin and anti–β-COP for two-stage Golgi-binding assays. GTPγS alone (lanes 1 and 2) or GTPγS plus myristoylated recombinant ARF1 (lanes 3 and 4) was included in the first-stage incubation. The second stage incubation contained cytosol alone (lanes 1 and 3) or cytosol plus GTPγS (lanes 2 and 4). After the second stage, the membranes were isolated by flotation and the levels of bound actin and coatomer were analyzed as above.
The reconstitution of actin assembly with purified recombinant ARF1 provided a means to further characterize the role of ARF in this process. In this regard, two-stage incubations were carried out to determine whether ARF is not only necessary but also sufficient to mediate the GTPγS-dependent actin assembly on the Golgi (Figure 1B). In the first stage, Golgi membranes were incubated with or without ARF1 and GTPγS and reisolated by centrifugation as before. The membranes were then incubated in a second stage with either cytosol alone or cytosol plus GTPγS. As expected, whenever GTPγS was included in the second stage together with cytosol, both actin assembly and coatomer binding were observed (Figure 1B, lanes 2 and 4). When recombinant ARF1 was prebound to the membrane in stage 1, no additional GTPγS was required in the second stage for coatomer binding (lane 3). Unexpectedly, when ARF1 was prebound to the Golgi membranes (lanes 3 and 4), actin assembly was observed in the second stage only when GTPγS was included in the incubation. Thus, ARF activation is necessary for both actin assembly and coatomer binding (Figure 1A), yet it is sufficient only for coatomer binding (Figure 1B). The GTPγS requirement for actin assembly in the second-stage incubation implicates a second GTP-binding protein, in addition to ARF, in regulating actin levels on the Golgi membranes.
ARF-dependent Actin Assembly Requires Cdc42
Members of the Rho family of GTP-binding proteins are principally involved in regulating dynamic rearrangements of the actin cytoskeleton (Hall and Nobes, 2000) and play important regulatory roles in protein trafficking (Garrett et al., 2000). Thus, these proteins seemed to be good candidates for the second GTP-binding protein involved in the ARF1-dependent actin assembly on the Golgi membranes. We tested this possibility by measuring the Golgi-associated actin levels in the presence of specific inhibitors of the Rho-related GTP-binding proteins. When C. difficile toxin B, which inhibits Rho-family members through glucosylation (Boquet, 1999), was added to the Golgi-binding assay, actin assembly was blocked in a dose-dependent manner (Figure 2A). Although toxin B, completely blocked the ARF/GTPγS-dependent assembly of actin on the Golgi membranes, it had little effect on coatomer binding (Figure 2A). This result indicates that ARF-dependent actin assembly, but not ARF-mediated vesicle-coat assembly requires the activity of a Rho-like GTP-binding protein. To further define which Rho family member acts downstream of ARF in this process, we used exoenzyme C3 from C. botulinum, which specifically ADP-ribosylates Rho, but not Rac and Cdc42 (Boquet, 1999). The addition of exoenzyme C3 had little or no effect on Golgi actin assembly (Figure 2B). C3 exoenzyme was also without effect on coatomer recruitment. These results suggest that a Rho family member, but not Rho itself, is required for actin binding.
Figure 2.

Actin binding requires the ARF-dependent regulation of Cdc42/Rac levels on the Golgi membranes. “Float-up” Golgi binding assays were carried out in the presence of the indicated concentrations of toxin B (A) or exoenzyme C3 (B). The levels of bound actin and β-COP were determined by Western blotting. Plotted is the average value from three independent experiments. The bars represent the SE of the mean. In B one half of each error bar is omitted for clarity. (C) Western blots probed with antibodies against Rho, Rac, and Cdc42 from a “float-up” binding assay (lanes 1 and 2). The incubations were carried out without (lane 1) or with (lane 2) GTPγS. Lane 3 contains 20 μg of the bovine brain cytosol preparation used for these studies. Lane 4 contains 12 μg of the rat-liver Golgi-membrane preparation used for the binding assays. (D) Western blot probed with the anti-Cdc42 and anti-Rac antibodies for two-stage Golgi binding assays carried out with or without ARF1 and brefeldin A (BfA) in the first stage as indicated. The conditions were identical to those used in Figure 1A. (E) Golgi-binding assays were carried out with whole cytosol (lanes 1 and 2) or coatomer-depleted cytosol (lanes 3–6). Purified coatomer (0.4 mg/ml) was added where indicated. Shown are Western blots probed with anti-Cdc42 antibodies.
Based on these results, one way that ARF could regulate actin assembly is by affecting the levels of the Rho-family GTP-binding proteins on the Golgi membranes. We tested this possibility by examining the levels of Cdc42, Rho, and Rac on Golgi membranes using the Golgi-binding assays (Figure 2, C and D). All three Rho family members are present in the brain cytosol and in the membrane preparations used for these experiments (Figure 2C, lanes 3 and 4). Interestingly, the binding of Cdc42 and Rac to the Golgi membranes, but not Rho, was found to be GTPγS dependent (Figure 2C, lanes 1 and 2). The levels of Rac and Cdc42 on the membrane were markedly increased after the incubation (cf. lanes 2 and 4), indicating that the bulk of the bound protein was derived from the cytosol. To determine if ARF was involved in the GTPγS-dependent localization of Cdc42 and Rac to the membranes, we examined the effects of both BFA and recombinant myristoylated ARF1 protein on Cdc42 and Rac binding. The levels of Cdc42 and Rac bound to the Golgi membranes appear to be strictly dependent on ARF activity. Figure 2D shows that in the presence of BFA, there is a significant decrease in the levels of these two GTP-binding proteins bound to the Golgi membranes. This decrease is prevented by the presence of prebound ARF1 (Figure 2D, cf. lanes 2 and 4). Therefore, ARF1-regulated changes in Cdc42 and Rac levels on the Golgi correlate precisely with ARF-mediated changes in actin levels. The fact that Rho levels are not sensitive to ARF activation (Figure 2C) is consistent with the results using C3 exoenzyme (Figure 2B). Taken together, the results indicate that Rho itself does not act downstream of ARF1 in actin binding to the Golgi membranes.
The results from the toxin and binding experiments suggest that Rac or Cdc42 are likely candidates to mediate actin assembly on the Golgi. Rac and Cdc42 are often observed to behave similarly in vitro, and both interact with CRIB (Cdc42/Rac interactive-binding) domains (Hoffman and Cerione, 2000). Several lines of evidence suggest that Cdc42 may be important for Golgi function. Cdc42 and the Cdc42 effector protein, IQGAP, are found to be localized to the Golgi apparatus, and the localization of Cdc42 is disrupted by the expression of dominant-negative but not wild-type ARF (Erickson et al., 1996; McCallum et al., 1998). We have also examined the subcellular localization of the Rho-family members in NRK cells and found that Cdc42 localizes to the Golgi apparatus.
The recently described interaction between coatomer and Cdc42 (Wu et al., 2000) could be responsible for mediating the ARF-sensitive localization of Cdc42 to the Golgi membrane. We tested this possibility by examining whether coatomer was necessary for Cdc42 binding to the membranes. Figure 2E shows that Cdc42 does not bind Golgi from coatomer-depleted cytosol. Addition of purified coatomer complex restored the GTPγS-dependent binding (lanes 5 and 6). These results show that ARF mediates Cdc42 recruitment to the Golgi through the binding interaction between coatomer and Cdc42.
Cdc42 and Rac can act through the WASP/Scar proteins to trigger actin polymerization via the protein complex Arp2/3 (Mullins, 2000). The C-terminal (Arp2/3-binding) domain of WASP, containing a region homologous to cofilin (C) and an acidic region (A), is a dominant inhibitor of Arp2/3 activation (Machesky and Insall, 1998; Rohatgi et al., 1999). We used GST fusion proteins containing the C-terminal (CA) domains for both the WASP protein and the more ubiquitously expressed isoform N-WASP to test whether Arp2/3 activation was required for ARF-dependent actin assembly on the Golgi membrane. Figure 3 shows that addition of either the WASP-CA domain (lanes 2–5) or the N-WASP-CA domain (lanes 6–9) inhibited the ARF/GTPγS-dependent actin binding in a dose-dependent manner. By contrast, the WASP-CA domains had no effect on coatomer binding to the membrane. A control incubation with GST affected neither actin assembly nor coatomer binding (Figure 3, lanes 1 and 11). These results indicate that Cdc42 acts through the Arp2/3 complex during ARF-dependent actin assembly on the Golgi membranes.
Figure 3.
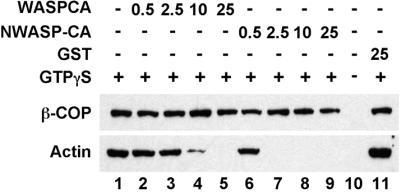
Actin assembly on the Golgi membranes requires Arp2/3 activation. Shown is a Western blot probed for β-COP and actin for Golgi-binding assays. GST-WASP-CA fusion protein (lanes 2–5) or the GST-NWASP-CA fusion protein (lanes 6–9) containing the dominant inhibitory Arp2/3-binding domains were added at the indicated concentrations (in μg/ml). After the incubation, the Golgi membranes were reisolated by flotation. In lane 11, purified GST protein was added as a control. GTPγS was included in all of the incubations except for lane 10.
mAbp1 and Drebrin Define Two Distinct ARF-dependent Actin Pools on Golgi Membranes
We have previously reported that activation of ARF leads to the binding of at least two distinct actin/actin-binding protein complexes on Golgi membranes (Fucini et al., 2000). The two actin-based complexes were differentiated based on their sensitivity to cytochalasin D, whether they could be extracted from the membrane with salt and whether they contained the actin-binding protein, drebrin (Fucini et al., 2000). Drebrin was specifically bound to actin upon ARF activation, whereas other actin-binding proteins present in the cytosol were excluded (Fucini et al., 2000).
A protein, mAbp1 (SH3p7), has been described that shares homology with the actin-binding domain of drebrin, but unlike drebrin it has a C-terminal SH3 domain (Sparks et al., 1996; Larbolette et al., 1999; Kessels et al., 2000). Both the mammalian and the yeast homologues of Abp1 as well as other SH3-domain containing actin-binding proteins have been implicated in endocytosis (Wesp et al. 1997; Kessels et al., 2001; reviewed by Qualmann et al., 2000). Given a potential role in protein trafficking for both homologues, we decided to further characterize drebrin and mAbp1 levels during ARF-dependent actin assembly on Golgi membranes. We generated an antibody against an mAbp1-GST fusion protein (see MATERIALS AND METHODS). This antibody recognizes a protein of ∼55 kDa that binds to phalloidin polymerized F-actin (Figure 4, lanes 1 and 2) and labels NRK cells expressing mAbp1 from a plasmid. We used this antibody together with an antibody against drebrin to characterize the binding of both of these proteins to Golgi membranes upon ARF activation (Figure 4). Surprisingly, although drebrin and mAbp1 share sequence homology through their N-terminal actin-binding domains, we find that they are largely segregated into the two different ARF1-dependent actin complexes on the Golgi membranes. Consistent with our previous findings (Fucini et al., 2000), drebrin is present in a complex that can be extracted from the Golgi membranes with 250 mM KCl (Figure 4, lane 8). This complex is sensitive to the actin-depolymerizing toxin cytochalasin D (lane 9). We now find that this drebrin-enriched pool of actin is largely devoid of mAbp1. A second actin pool is resistant to salt extraction from the membrane and is resistant to cytochalasin D treatment (Fucini et al., 2000). The mAbp1 protein is present exclusively in this cytochalasin D-resistant actin pool (cf. lanes 4 and 5). The assembly of both actin structures requires the activity of ARF and thus can be blocked by the addition of BFA (lanes 6 and 10). Hence, ARF activation leads to the assembly of at least two distinct actin pools, each with its own unique composition of actin-binding proteins.
Figure 4.
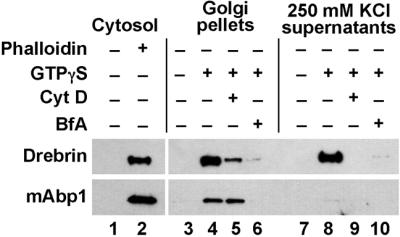
The F-actin binding proteins drebrin and mAbp1 are differentially segregated into two ARF-dependent Golgi actin pools. Golgi membranes were incubated with cytosol as done previously for binding assays. GTPγS, 20 μM cytochalasin D (CytD) or brefeldin A (BfA), were added to the incubations where indicated. After the incubation the membranes were washed with 250 mM potassium chloride and pelleted by centrifugation. Shown is a Western blot of the pellets (lanes 3–6) and the supernatants (lanes 7–10) probed with antidrebrin and anti-mAbp1 antibodies. Lanes 1 and 2: the pellet fraction after the sedimentation of cytosol that had been incubated alone (lane 1) or in the presence of 200 μM phalloidin to assemble F-actin.
Given the ARF-dependent binding of drebrin and mAbp1 to isolated Golgi membranes, we reasoned that these actin-binding proteins may be involved in Golgi function and thus should be found on Golgi membranes in whole cells. In this regard, we have analyzed the subcellular localization of drebrin and mAbp1. Drebrin was found localized throughout the cell and thus could potentially participate in numerous cellular processes including Golgi transport. mAbp1, on the other hand, has previously been shown to localize to membrane ruffles and to the periplasmic region of the cell (Kessels et al., 2000). This localization was shown to be sensitive to Rac activation. We find that the periplasmic mAbp1 is further enriched at a juxtanuclear structure that colocalizes with the Golgi marker β-COP (Figure 5). We also observe the staining of cell surface structures resembling membrane ruffles or lamellipodia (Figure 5A). The subcellular localization of mAbp1 is consistent with a role in Golgi function.
Figure 5.

mAbp1 is localized to the Golgi apparatus and distinct cell-surface structures. Confocal micrographs of NRK cells that have been colabeled with antibodies against mAbp1 (A) and the Golgi marker β-COP (B). (C) A merged image of colabeled cells in A and B. mAbp1 was visualized with Texas Red–conjugated anti-rabbit secondary antibodies. β-COP was visualized with a FITC-conjugated anti-mouse secondary antibody. For the merged image (C), overlapping signals appear yellow. Bar: B, 5 μm.
Coatomer-p23 Interactions Affect the Binding of mAbp1 to Golgi Membranes
Because both the mAbp1-enriched and the drebrin-enriched actin pools are assembled upon ARF activation, we were interested in mechanisms downstream of ARF that might specify their selective recruitment to the Golgi membrane. We found that inhibiting Rac/Cdc42 activity with toxin B (Figure 6A) or blocking the activation of the Arp2/3 complex with WASP-CA (Figure 6B) prevents the binding of both mAbp1 and drebrin to the Golgi membranes. The GTPase dependence of the mAbp1 recruitment to the Golgi membranes is consistent with the observation that the lamellipodial recruitment of mAbp1 is controlled by signaling pathways leading to the activation of Rho-family GTPases (Kessels et al., 2000). Exoenzyme C3 has no effect on the binding of either mAbp1 or drebrin indicating again that Rho is not involved in regulating the Golgi actin. Although mAbp1 recruitment is cytochalasin D resistant, it does require actin assembly because it is sensitive to latrunculin A. Thus, our data indicate that both drebrin and mAbp1 are recruited to the Golgi through the Cdc42/Arp2/3-dependent assembly of actin.
Figure 6.
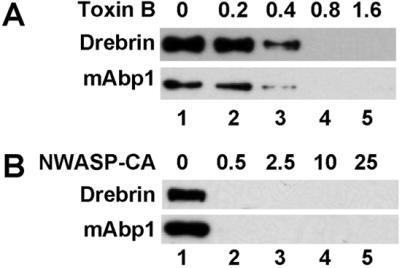
Assembly of both Golgi actin pools requires Rac/Cdc42 and Arp2/3 activation. Shown is a Western blot of Golgi-binding assays probed with antibodies against drebrin and mAbp1. GTPγS plus either Toxin B (A) or the N-WASP-CA (B) were added to the incubation at the indicated concentrations in μg/ml. After the incubations, the Golgi membranes were isolated by flotation.
Given that ARF regulates both coatomer binding and actin assembly, we decided to explore whether additional connections exist between coatomer function and the recruitment of the actin binding proteins and regulatory proteins to the Golgi membranes. For these experiments, we used a γ-COP–binding peptide corresponding to the C terminus of the putative cargo receptor protein, p23 (Harter and Wieland, 1998; Bremser et al., 1999). This peptide was shown previously to disrupt the interaction between coatomer and Cdc42 (Wu et al., 2000). The p23/p24 family of proteins is abundant integral membrane components of both COPI- and COPII-coated transport vesicles (Schimmöller et al., 1995; Stamnes et al., 1995; Sohn et al., 1996) and interact with coat proteins via their C-terminal cytosolic domain (Fiedler et al., 1996; Sohn et al., 1996; Dominguez et al., 1998). They may act as receptor proteins for the packaging or quality control of other types of vesicle cargo (Muniz et al., 2000; Springer et al., 2000). When the p23 C-terminal peptide was added to the incubation, it blocked the ARF-dependent binding of Cdc42 and Rac to the Golgi membranes (Figure 7A). The effects of the p23 C-terminal peptide were specific to Rac and Cdc42 because the levels of Rho were unaffected by the presence of the peptide (Figure 7B). These results are consistent with our finding (Figure 2E) that ARF recruits Cdc42 to the Golgi membrane through its interaction with coatomer.
Figure 7.

mAbp1 binding is selectively blocked by the presence of the coatomer-binding domain of p23 protein. (A) Golgi-binding assays were carried out with GTPγS and the p23 C-terminal peptide (250 μM) as indicated. After the reaction, the membranes were reisolated by sedimentation. Shown is a Western blot of the reisolated Golgi membranes probed for coatomer (β-COP), drebrin, mAbp1, Cdc42, and Rac. (B) A Western blot of Golgi-binding assays carried out as in A and probed with anti-mAbp1 and Rho. (C) A Western blot of Golgi-binding assays into which the p23 peptide was added at the indicated concentrations in μM. The membranes were reisolated by flotation after the incubation.
In addition to analyzing the effects of the peptide on the Rho-family GTP-binding proteins, we have investigated whether the coatomer/Cdc42 interaction influences the association of actin and actin-binding proteins with the Golgi membranes. Figure 7 shows that the addition of the p23 C-terminal peptide blocked the assembly of the mAbp1-containing pool of actin. Addition of the peptide did not block the binding of drebrin to the Golgi membranes (Figure 7, A and C). Actin levels were reduced, but not completely blocked, upon addition of the peptide, indicating the existence of both a peptide-sensitive and a peptide-resistant actin pool (Figure 7C). Coatomer binding was unaffected by the peptide. The inhibition of mAbp1 binding to the Golgi membranes occurs at a peptide concentration between 100 and 500 μM (Figure 7C), the same concentration range previously shown to disrupt the binding interaction between coatomer and Cdc42 (Wu et al., 2000). Together, these results indicate that the mAbp1-enriched actin, but not the drebrin-enriched actin pool, is regulated by the p23-sensitive interaction between coatomer and Cdc42/Rac.
mAbp1 Is Involved in Protein Transport
Disrupting Cdc42 function has profound effects on Golgi trafficking (Kroschewski et al. 1999; Wu et al., 2000; Müsch et al., 2001). We surmised that these effects could be manifested through the ARF-Cdc42/Rac-Arp2/3–dependent assembly of the mAbp1 actin pool. We tested this directly by examining the effects of exogenously expressed mAbp1 on protein trafficking at the Golgi apparatus. For these experiments, we measured anterograde transport using NRK cells expressing the temperature-sensitive mutant form of the vesicular stomatitis virus glycoprotein (ts045-VSVG). The temperature-sensitive VSV G protein accumulates in the ER at the restrictive temperature, 39°C and is transported from the ER to the Golgi apparatus at the permissive temperature, 32°C (for examples, see Hirschberg et al., 1998; Kroschewski et al., 1999; Wu et al., 2000).
Figure 8B shows that after an incubation at 39°C, the VSV G protein is accumulated in a dispersed ER compartment. If the cells are then switched to 32°C, the VSV G is transported to the Golgi apparatus, which can be observed as a compact perinuclear structure (Figure 8D). The Golgi localization of VSV G protein after the shift to the permissive temperature was confirmed by demonstrating colocalization with α-mannosidase II. In transfected cells overexpressing exogenous myc-mAbp1, a significant inhibition of VSV G transport to the Golgi was observed (Figure 8, C and D). A count of three independent experiments showed that although the majority (513/602 or 85%) of untransfected cells displayed Golgi localized VSV G, only 38% (106/262) of the myc-mAbp1–expressing cells displayed Golgi localization. A control plasmid encoding GFP (see MATERIALS AND METHODS) had only a minimal effect on VSV-G transport, with 72% (215/300) of the cells displaying Golgi localization. Even more pronounced effects on VSVG transport were observed if only cells expressing mAbp1 at high levels were examined. This result is consistent with a role for mAbp1 in coatomer-mediated transport because a similar inhibition of ER to Golgi transport has been observed in both mammalian cells and yeast when COPI vesicle transport is compromised (Peter et al., 1993; Gaynor and Emr, 1997). Expression of mAbp1 does not lead to general disruption of Golgi morphology because mannosidase II localization is largely unaffected (Figure 8, E and F). Unlike the endogenous protein (Figure 5), the overexpressed myc-mAbp1 protein is localized throughout the cell (Figure 8, A, C, and E). It is possible that this aberrant localization of mAbp1 leads to the improper targeting of other proteins that normally interact with mAbp1 at the Golgi membrane, thus explaining the defects in protein transport. The similarity in defects observed upon disrupting Cdc42 function (Wu et al., 2000) and disrupting mAbp1 function (Figure 8) is consistent with these proteins functioning together in a pathway.
Figure 8.
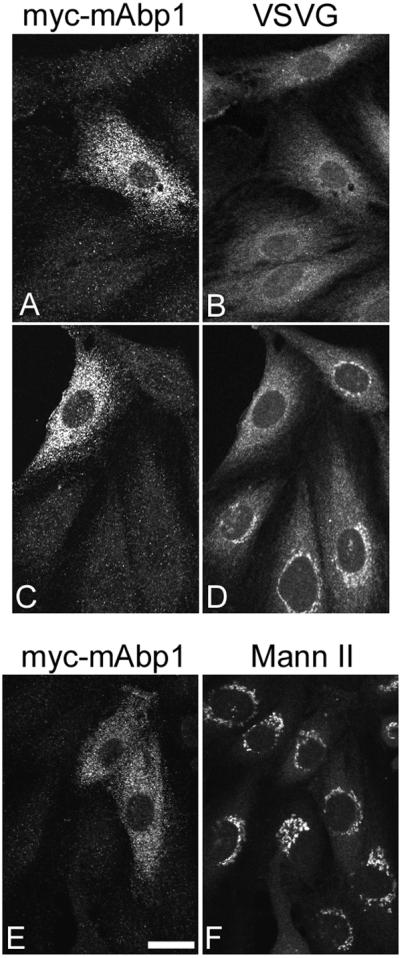
mAbp1 is required for anterograde trafficking to the Golgi apparatus. NRK cells expressing ts045-VSVG were transfected with a plasmid encoding myc-tagged mAbp1 and grown at the restrictive temperature (39°C) for 14 h (A and B). (C–F) The 39°C incubation was followed by an incubation at the permissive temperature (32°C) for 15 min. The transfected cells were identified by indirect immunofluorescence using an anti-myc antibody (A, C, and E). The VSV G protein was visualized with an anti–VSV-G antibody (B and D). In F, the cells were decorated with anti–α-mannosidase II (Mann II) to observe the Golgi morphology. The left and right panels for each set represent the same field of cells. Bar: E, 5 μm.
In summary, we have characterized a Golgi-localized actin-binding protein, mAbp1, that is specifically regulated by the coat-binding domain of the vesicle cargo protein, p23, and functions in anterograde protein trafficking to the Golgi. These findings have important implications for the role of actin in trafficking and the mechanisms of vesicle targeting.
DISCUSSION
What Role Does Actin Play in Vesicle Formation and Targeting?
We have uncovered a signaling pathway, initiated by ARF activation that leads to the recruitment of specific actin-binding proteins, mAbp1 and drebrin, to the Golgi membrane. Components of this pathway, mAbp1 (Figure 8) and Cdc42 (Kroschewski et al. 1999; Wu et al., 2000; Müsch et al., 2001), are required for normal protein transport to or from the Golgi apparatus. Our findings suggest specific roles for actin in vesicle trafficking at the Golgi apparatus.
An anticipated role for the cytoskeleton in transport is in the targeting and physical translocation of vesicles or other transport intermediates. In general, two types of mechanisms have been postulated for actin-based translocation of organelles and vesicles. The first involves moving organelles or vesicles along actin microfilaments via myosin motors (Pruyne et al., 1998; Stow and Heimann, 1998). A second mechanism involves actin comet-tails in which the polymerization of actin itself generates the force to translocate organelles or vesicles (Cameron et al., 2000). Cdc42, WASP, and Arp2/3 are involved in the comet-tail–based movement of endosomal or Golgi-derived membranes (Rozelle et al., 2000; Taunton et al., 2000).
Although we cannot rule out a direct role for ARF-regulated actin assembly in vesicle translocation, we believe our results are more consistent with a role earlier in vesicle assembly. Our results suggest that mAbp1 actin assembly is activated upon recruitment of the coatomer-Cdc42 complex by ARF. Importantly, the presence of cargo proteins is predicted to inactivate this signaling pathway through two mechanisms. First, some types of cargo proteins such as the KDEL receptor (ERD2) may recruit ARF-GAP to the site of vesicle assembly (Aoe et al., 1997). This would result in the inactivation of ARF and inhibit further coat assembly and actin polymerization. Second, we show in this study that p23 proteins could disrupt the interaction between coatomer and Cdc42 that also would result in an inhibition of mAbp1 binding. The p23/p24 family of proteins can be considered cargo proteins in that they are very abundant in transport vesicles yet they may not be necessary for vesicle formation (Schimmöller et al., 1995; Stamnes et al., 1995; Springer et al., 2000). Therefore, the pathway leading to mAbp1 actin assembly is very likely most active after coat binding but before cargo recruitment.
Given the timing of mAbp1 actin assembly, we propose that instead of being directly involved in translocation, the mAbp1 actin may be involved in limiting the interactions of a nascent vesicle with the targeting or scission machinery. Coupling these interactions to the presence of cargo proteins could prevent the premature release or translocation of the vesicles. A prediction of this model is that the transport of some proteins may become inefficient, because of the release of incompletely filled vesicles, upon disruption of mAbp1 actin assembly. Such inefficiency could explain some of the trafficking defects observed upon disrupting Cdc42 or mAbp1 function.
With regard to the above model, recent results indicate that the GTP-binding protein dynamin, which is known to play a central role in the release of clathrin-coated vesicles, also binds to the SH3-domains of mAbp1, cortactin, and syndapin (McNiven et al., 2000; Qualmann and Kelly; 2000; Kessels et al., 2001). The syndapin and mAbp1 interactions are critical for endocytosis (Qualmann and Kelly; 2000; Kessels et al., 2001). This raises the possibility that the actin cytoskeleton, together with dynamin, plays a role in regulating vesicle scission (Qualmann et al., 2000). Further studies will be required to test whether actin-binding proteins can regulate dynamin function, vesicle release, and/or vesicle translocation at the Golgi.
The Mechanism for Actin Assembly on the Golgi Apparatus
We show that in addition to ARF, actin assembly involves recruitment of activated Cdc42 and/or Rac to the Golgi membrane and requires activation of the Arp2/3 complex. The ARF-mediated recruitment of Cdc42 to the Golgi membranes occurs through the direct interaction between the γ-COP subunit of coatomer and Cdc42 (Wu et al., 2000). The ARF dependence, coatomer dependence, and the sensitivity to γ-COP–binding peptides that we observe for Cdc42 recruitment to Golgi membranes are all consistent with this model. The interaction between coatomer and p23/p24 proteins has also been shown to affect the function of ARFGAP (Goldberg, 2000), suggesting that occupation of this site on γ-COP may play multiple important regulatory roles during vesicle assembly or targeting (Donaldson and Lippincott-Schwartz, 2000). Our data indicate that Cdc42 causes actin assembly, and subsequently mAbp1 recruitment, by causing Arp2/3 to become activated. The yeast Abp1 protein was recently shown to activate Arp2/3 directly (Goode et al., 2001). It is unlikely, however, that the mammalian protein shares this property because it is missing acidic domains required for Arp2/3 activation by the yeast homolog.
Unlike mAbp1 binding to the Golgi membrane, the ARF-dependent assembly of the drebrin-enriched pool of actin was insensitive to the addition of the p23 peptide, even although it was sensitive to the Rho-family inhibitor toxin B. One possibility is that the drebrin-enriched actin pool is assembled by the recruitment of Cdc42/Rac via interactions with a second type of ARF-dependent coat protein that does not interact with p23, such as the AP1/clathrin coats. Another possibility is that a different Rho-family GTP-binding protein mediates the assembly of the drebrin-enriched actin pool. In this regard, several novel GTP-binding proteins related to Cdc42 have been described recently (Neudauer et al., 1998; Vignal et al., 2000).
Although our data show a requirement for Rho-family members and Arp2/3 in ARF-dependent actin assembly on the Golgi membranes, other studies indicate that ARF acts through PI-4-kinase or PI-5-kinase to mediate changes in the actin cytoskeleton and membrane ruffling (Godi et al., 1999; Honda et al., 1999; Radhakrishna et al., 1999; reviewed by Mostov et al., 2000). These two mechanisms for ARF action are not mutually exclusive and it is likely that both PI metabolism and Cdc42-Arp2/3 signaling play a role in ARF-dependent regulation of the actin cytoskeleton on the Golgi membranes (Godi et al., 1998 and 1999; Fucini et al., 2000).
Do Vesicle Contents and Coats Direct Cytoskeleton-mediated Targeting Mechanisms?
Given the large number of transport steps in a cell and the relatively small number of vesicle types that have been characterized, it is likely that each type of vesicle will have the capacity to mediate several different trafficking steps. For instance, COPI vesicles have been implicated in both anterograde- and retrograde-directed trafficking at the Golgi apparatus as well as in endosomal trafficking (Daro et al., 1997; Gu et al., 1997; Orci et al., 1997; Pelham and Rothman, 2000). Our results suggest that the types of cargo proteins entering a vesicle during budding could influence the actin-dependent regulation of the targeting or fission machinery. It is possible that the distinct ARF-dependent actin pools that we observed may mediate different vesicle-type-specific functions.
As the molecular mechanisms involved in vesicle assembly and vesicle fusion become better understood, a remaining challenge is to elucidate the catalytic and regulatory mechanisms that allow precise and efficient sorting and targeting of proteins and lipids so that the correct compartmentalization of cellular components can be maintained over the lifetime of a cell. Clarifying the role of the cytoskeleton and its regulation in protein trafficking will be an important step toward understanding these processes. The recent identification of specific proteins and mechanisms that are involved at the interface between protein trafficking and cytoskeletal regulation will allow for many exciting advances in this area in the near future.
ACKNOWLEDGMENTS
The authors thank Drs. Lois Weisman, Jatinder Ahluwalia, and Jeffrey Pessin for helpful discussions. This work was supported by grants to M.S. from the American Cancer Society, the Roy J. Carver Charitable Trust, and the American Heart Association Heartland Affiliate.
Footnotes
Article published online ahead of print. Mol. Biol. Cell 10.1091/mbc.01–11-0547. Article and publication date are at www.molbiolcell.org/cgi/10.1091/mbc.01–11-0547.
REFERENCES
- Aoe T, Cukierman E, Lee A, Cassel D, Peters PJ, Hsu VW. The KDEL receptor, ERD2, regulates intracellular traffic by recruiting a GTPase-activating protein for ARF1. EMBO J. 1997;16:7305–7316. doi: 10.1093/emboj/16.24.7305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boquet P. Bacterial toxins inhibiting or activating small GTP-binding proteins. Ann NY Acad Sci. 1999;886:83–90. doi: 10.1111/j.1749-6632.1999.tb09403.x. [DOI] [PubMed] [Google Scholar]
- Boshans RL, Szanto S, van Aelst L, D'Souza-Schorey C. ADP-ribosylation factor 6 regulates actin cytoskeleton remodeling in coordination with Rac1 and RhoA. Mol Cell Biol. 2000;20:3685–3694. doi: 10.1128/mcb.20.10.3685-3694.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bremser M, Nickel W, Schweikert M, Ravazzola M, Amherdt M, Hughes CA, Sollner TH, Rothman JE, Wieland FT. Coupling of coat assembly and vesicle budding to packaging of putative cargo receptors. Cell. 1999;96:495–506. doi: 10.1016/s0092-8674(00)80654-6. [DOI] [PubMed] [Google Scholar]
- Cameron LA, Giardini PA, Soo FS, Theriot JA. Secrets of actin-based motility revealed by a bacterial pathogen. Nat Rev Mol Cell Biol. 2000;1:110–119. doi: 10.1038/35040061. [DOI] [PubMed] [Google Scholar]
- Daro E, Sheff D, Gomez M, Kreis T, Mellman I. Inhibition of endosome function in CHO cells bearing a temperature-sensitive defect in the coatomer (COPI) component epsilon-COP. J Cell Biol. 1997;139:1747–1759. doi: 10.1083/jcb.139.7.1747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Matteis MA, Morrow JS. Spectrin tethers and mesh in the biosynthetic pathway. J Cell Sci. 2000;113:2331–2343. doi: 10.1242/jcs.113.13.2331. [DOI] [PubMed] [Google Scholar]
- Dominguez M, Dejgaard K, Fullekrug J, Dahan S, Fazel A, Paccaud JP, Thomas DY, Bergeron JJ, Nilsson T. gp25L/emp24/p24 protein family members of the cis-Golgi network bind both COP I and II coatomer. J Cell Biol. 1998;140:751–765. doi: 10.1083/jcb.140.4.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donaldson JG, Lippincott-Schwartz J. Sorting and signaling at the Golgi complex. Cell. 2000;101:693–696. doi: 10.1016/s0092-8674(00)80881-8. [DOI] [PubMed] [Google Scholar]
- D'Souza-Schorey C, Boshans RL, McDonough M, Stahl PD, Van Aelst L. A role for POR1, a Rac1-interacting protein, in ARF6-mediated cytoskeletal rearrangements. EMBO J. 1997;16:5445–5454. doi: 10.1093/emboj/16.17.5445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erickson JW, Zhang C, Kahn RA, Evans T, Cerione RA. Mammalian Cdc42 is a brefeldin A-sensitive component of the Golgi apparatus. J Biol Chem. 1996;271:26850–26854. doi: 10.1074/jbc.271.43.26850. [DOI] [PubMed] [Google Scholar]
- Fiedler K, Veit M, Stamnes MA, Rothman JE. Bimodal interaction of coatomer with the p24 family of putative cargo receptors. Science. 1996;273:1396–1399. doi: 10.1126/science.273.5280.1396. [DOI] [PubMed] [Google Scholar]
- Fucini RV, Navarrete A, Vadakkan C, Lacomis L, Erdjument-Bromage H, Tempst P, Stamnes M. Activated ADP-ribosylation factor assembles distinct pools of actin on Golgi membranes. J Biol Chem. 2000;275:18824–18829. doi: 10.1074/jbc.M000024200. [DOI] [PubMed] [Google Scholar]
- Garrett WS, Chen LM, Kroschewski R, Ebersold M, Turley S, Trombetta S, Galan JE, Mellman I. Developmental control of endocytosis in dendritic cells by Cdc42. Cell. 2000;102:325–334. doi: 10.1016/s0092-8674(00)00038-6. [DOI] [PubMed] [Google Scholar]
- Gaynor EC, Emr SD. COPI-independent anterograde transport: cargo-selective ER to Golgi protein transport in yeast COPI mutants. J Cell Biol. 1997;136:789–802. doi: 10.1083/jcb.136.4.789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godi A, Pertile P, Meyers R, Marra P, Di Tullio G, Iurisci C, Luini A, Corda D, De Matteis MA. ARF mediates recruitment of PtdIns-4-OH kinase-beta and stimulates synthesis of PtdIns(4,5)P2 on the Golgi complex. Nat Cell Biol. 1999;1:280–287. doi: 10.1038/12993. [DOI] [PubMed] [Google Scholar]
- Godi A, et al. ADP ribosylation factor regulates spectrin binding to the Golgi complex. Proc Natl Acad Sci USA. 1998;95:8607–8612. doi: 10.1073/pnas.95.15.8607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg J. Decoding of sorting signals by coatomer through a GTPase switch in the COPI coat complex. Cell. 2000;100:671–679. doi: 10.1016/s0092-8674(00)80703-5. [DOI] [PubMed] [Google Scholar]
- Goode BL, Rodal AA, Barnes G, Drubin DG. Activation of the Arp2/3 complex by the actin filament binding protein Abp1p. J Cell Biol. 2001;153:627–34. doi: 10.1083/jcb.153.3.627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu F, Aniento F, Parton RG, Gruenberg J. Functional dissection of COP-I subunits in the biogenesis of multivesicular endosomes. J Cell Biol. 1997;139:1183–1195. doi: 10.1083/jcb.139.5.1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall A, Nobes CD. Rho GTPases: molecular switches that control the organization and dynamics of the actin cytoskeleton. Philos Trans R Soc Lond B Biol Sci. 2000;355:965–970. doi: 10.1098/rstb.2000.0632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harter C, Wieland FT. A single binding site for dilysine retrieval motifs and p23 within the gamma subunit of coatomer. Proc Natl Acad Sci USA. 1998;95:11649–11654. doi: 10.1073/pnas.95.20.11649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heimann K, Percival JM, Weinberger R, Gunning P, Stow JL. Specific isoforms of actin-binding proteins on distinct populations of Golgi-derived vesicles. J Biol Chem. 1999;274:10743–10750. doi: 10.1074/jbc.274.16.10743. [DOI] [PubMed] [Google Scholar]
- Helms JB, Palmer DJ, Rothman JE. Two distinct populations of ARF bound to Golgi membranes. J Cell Biol. 1993;121:751–760. doi: 10.1083/jcb.121.4.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirschberg K, Miller CM, Ellenberg J, Presley JF, Siggia ED, Phair RD, Lippincott-Schwartz J. Kinetic analysis of secretory protein traffic and characterization of golgi to plasma membrane transport intermediates in living cells. J Cell Biol. 1998;143:1485–1503. doi: 10.1083/jcb.143.6.1485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman GR, Cerione RA. Flipping the switch: the structural basis for signaling through the CRIB motif. Cell. 2000;102:403–406. doi: 10.1016/s0092-8674(00)00045-3. [DOI] [PubMed] [Google Scholar]
- Honda A, et al. Phosphatidylinositol 4-phosphate 5-kinase alpha is a downstream effector of the small G protein ARF6 in membrane ruffle formation. Cell. 1999;99:521–532. doi: 10.1016/s0092-8674(00)81540-8. [DOI] [PubMed] [Google Scholar]
- Kessels MM, Engqvist-Goldstein AE, Drubin DG. Association of mouse actin-binding protein 1 (mAbp1/SH3P7), an src kinase target, with dynamic regions of the cortical actin cytoskeleton in response to Rac1 activation. Mol Biol Cell. 2000;11:393–412. doi: 10.1091/mbc.11.1.393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessels MM, Engqvist-Goldstein AE, Drubin DG, Qualmann B. Mammalian Abp1, a signal-responsive F-actin-binding protein, links the actin cytoskeleton to endocytosis via the GTPase dynamin. J Cell Biol. 2001;153:351–66. doi: 10.1083/jcb.153.2.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroschewski R, Hall A, Mellman I. Cdc42 controls secretory and endocytic transport to the basolateral plasma membrane of MDCK cells. Nat Cell Biol. 1999;1:8–13. doi: 10.1038/8977. [DOI] [PubMed] [Google Scholar]
- Larbolette O, Wollscheid B, Schweikert J, Nielsen PJ, Wienands J. SH3P7 is a cytoskeleton adapter protein and is coupled to signal transduction from lymphocyte antigen receptors. Mol Cell Biol. 1999;19:1539–1546. doi: 10.1128/mcb.19.2.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machesky LM, Insall RH. Scar1 and the related Wiskott-Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex. Curr Biol. 1998;8:1347–1356. doi: 10.1016/s0960-9822(98)00015-3. [DOI] [PubMed] [Google Scholar]
- Malhotra V, Serafini T, Orci L, Shepherd JC, Rothman JE. Purification of a novel class of coated vesicles mediating biosynthetic protein transport through the Golgi stack. Cell. 1989;58:329–336. doi: 10.1016/0092-8674(89)90847-7. [DOI] [PubMed] [Google Scholar]
- McCallum SJ, Erickson JW, Cerione RA. Characterization of the association of the actin-binding protein, IQGAP, and activated Cdc42 with Golgi membranes. J Biol Chem. 1998;273:22537–22544. doi: 10.1074/jbc.273.35.22537. [DOI] [PubMed] [Google Scholar]
- McNiven MA, Kim L, Krueger EW, Orth JD, Cao H, Wong TW. Regulated interactions between dynamin and the actin-binding protein cortactin modulate cell shape. J Cell Biol. 2000;151:187–198. doi: 10.1083/jcb.151.1.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellman I, Warren G. The road taken: past and future foundations of membrane traffic. Cell. 2000;100:99–112. doi: 10.1016/s0092-8674(00)81687-6. [DOI] [PubMed] [Google Scholar]
- Miki H, Miura K, Takenawa T. N-WASP, a novel actin-depolymerizing protein, regulates the cortical cytoskeletal rearrangement in a PIP2-dependent manner downstream of tyrosine kinases. EMBO J. 1996;15:5326–5335. [PMC free article] [PubMed] [Google Scholar]
- Mostov KE, Verges M, Altschuler Y. Membrane traffic in polarized epithelial cells. Curr Opin Cell Biol. 2000;12:483–490. doi: 10.1016/s0955-0674(00)00120-4. [DOI] [PubMed] [Google Scholar]
- Mullins RD. How WASP-family proteins, and the Arp2/3 complex convert intracellular signals into cytoskeletal structures. Curr Opin Cell Biol. 2000;12:91–96. doi: 10.1016/s0955-0674(99)00061-7. [DOI] [PubMed] [Google Scholar]
- Muniz M, Nuoffer C, Hauri HP, Riezman H. The Emp24 complex recruits a specific cargo molecule into endoplasmic reticulum-derived vesicles. J Cell Biol. 2000;148:925–930. doi: 10.1083/jcb.148.5.925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müsch A, Cohen D, Kreitzer G, Rodriguez-Boulan E. cdc42 regulates the exit of apical, and basolateral proteins from the trans-Golgi network. EMBO J. 2001;20:2171–2179. doi: 10.1093/emboj/20.9.2171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neudauer CL, Joberty G, Tatsis N, Macara IG. Distinct cellular effects and interactions of the Rho-family GTPase TC10. Curr Biol. 1998;8:1151–1160. doi: 10.1016/s0960-9822(07)00486-1. [DOI] [PubMed] [Google Scholar]
- Orci L, Stamnes M, Ravazzola M, Amherdt M, Perrelet A, Söllner TH, Rothman JE. Bidirectional transport by distinct populations of COPI-coated vesicles. Cell. 1997;90:335–349. doi: 10.1016/s0092-8674(00)80341-4. [DOI] [PubMed] [Google Scholar]
- Pelham HR, Rothman JE. The debate about transport in the Golgi–two sides of the same coin? Cell. 2000;102:713–719. doi: 10.1016/s0092-8674(00)00060-x. [DOI] [PubMed] [Google Scholar]
- Pruyne DW, Schott DH, Bretscher A. Tropomyosin-containing actin cables direct the Myo2p-dependent polarized delivery of secretory vesicles in budding yeast. J Cell Biol. 1998;143:1931–1945. doi: 10.1083/jcb.143.7.1931. [DOI] [PubMed] [Google Scholar]
- Peter F, Plutner H, Zhu H, Kreis TE, Balch WE. Beta-COP is essential for transport of protein from the endoplasmic reticulum to the Golgi in vitro. J Cell Biol. 1993;122:1155–1167. doi: 10.1083/jcb.122.6.1155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qualmann B, Kelly RB. Syndapin isoforms participate in receptor-mediated endocytosis and actin organization. J Cell Biol. 2000;148:1047–1061. doi: 10.1083/jcb.148.5.1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qualmann B, Kessels MM, Kelly RB. Molecular links between endocytosis and the actin cytoskeleton. J Cell Biol. 2000;150:F111–116. doi: 10.1083/jcb.150.5.f111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radhakrishna H, Al-Awar O, Khachikian Z, Donaldson JG. ARF6 requirement for Rac ruffling suggests a role for membrane trafficking in cortical actin rearrangements. J Cell Sci. 1999;112:855–866. doi: 10.1242/jcs.112.6.855. [DOI] [PubMed] [Google Scholar]
- Rohatgi R, Ma L, Miki H, Lopez M, Kirchhausen T, Takenawa T, Kirschner MW. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell. 1999;97:221–231. doi: 10.1016/s0092-8674(00)80732-1. [DOI] [PubMed] [Google Scholar]
- Rozelle AL, Machesky LM, Yamamoto M, Driessens MH, Insall RH, Roth MG, Luby-Phelps K, Marriott G, Hall A, Yin HL. Phosphatidylinositol 4,5-bisphosphate induces actin-based movement of raft-enriched vesicles through WASP-Arp2/3. Curr Biol. 2000;10:311–320. doi: 10.1016/s0960-9822(00)00384-5. [DOI] [PubMed] [Google Scholar]
- Schimmöller F, Singer-Kruger B, Schroder S, Kruger U, Barlowe C, Riezman H. The absence of Emp24p, a component of ER-derived COPII-coated vesicles, causes a defect in transport of selected proteins to the Golgi. EMBO J. 1995;14:1329–1339. doi: 10.1002/j.1460-2075.1995.tb07119.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohn K, Orci L, Ravazzola M, Amherdt M, Bremser M, Lottspeich F, Fiedler K, Helms JB, Wieland FT. A major transmembrane protein of Golgi-derived COPI-coated vesicles involved in coatomer binding. J Cell Biol. 1996;135:1239–1248. doi: 10.1083/jcb.135.5.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparks AB, Hoffman NG, McConnell SJ, Fowlkes DM, Kay BK. Cloning of ligand targets: systematic isolation of SH3 domain-containing proteins. Nat Biotechnol. 1996;14:741–744. doi: 10.1038/nbt0696-741. [DOI] [PubMed] [Google Scholar]
- Springer S, Chen E, Duden R, Marzioch M, Rowley A, Hamamoto S, Merchant S, Schekman R. The p24 proteins are not essential for vesicular transport in Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 2000;97:4034–4039. doi: 10.1073/pnas.070044097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamnes MA, Craighead MW, Hoe MH, Lampen N, Geromanos S, Tempst P, Rothman JE. An integral membrane component of coatomer-coated transport vesicles defines a family of proteins involved in budding. Proc Natl Acad Sci USA. 1995;92:8011–8015. doi: 10.1073/pnas.92.17.8011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stow JL, Heimann K. Vesicle budding on Golgi membranes: regulation by G proteins and myosin motors. Biochim Biophys Acta. 1998;1404:161–171. doi: 10.1016/s0167-4889(98)00055-x. [DOI] [PubMed] [Google Scholar]
- Taunton J, Rowning BA, Coughlin ML, Wu M, Moon RT, Mitchison TJ, Larabell CA. Actin-dependent propulsion of endosomes and lysosomes by recruitment of N-WASP. J Cell Biol. 2000;148:519–530. doi: 10.1083/jcb.148.3.519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valderrama F, Babia T, Ayala I, Kok JW, Renau-Piqueras J, Egea G. Actin microfilaments are essential for the cytological positioning and morphology of the Golgi complex. Eur J Cell Biol. 1998;76:9–17. doi: 10.1016/S0171-9335(98)80012-5. [DOI] [PubMed] [Google Scholar]
- Valderrama F, Luna A, Babia T, Martinez-Menarguez JA, Ballesta J, Barth H, Chaponnier C, Renau-Piqueras J, Egea G. The golgi-associated COPI-coated buds and vesicles contain beta/gamma-actin. Proc Natl Acad Sci USA. 2000;97:1560–1565. doi: 10.1073/pnas.97.4.1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vignal E, De Toledo M, Comunale F, Landopoulou A, Gauthier-Rouviere C, Blangy A, Fort P. Characterization of TCL, a new GTPase of the Rho family related to TC10 and Cdc42. J Biol Chem. 2000;275:36457–36464. doi: 10.1074/jbc.M003487200. [DOI] [PubMed] [Google Scholar]
- Waters MG, Beckers CJM, Rothman JE. Purification of coat protomers. Methods Enzymol. 1992;219:331–337. doi: 10.1016/0076-6879(92)19033-3. [DOI] [PubMed] [Google Scholar]
- Wesp A, Hicke L, Palecek J, Lombardi R, Aust T, Munn AL, Riezman H. End4p/Sla2p interacts with actin-associated proteins for endocytosis in Saccharomyces cerevisiae. Mol Biol Cell. 1997;8:2291–2306. doi: 10.1091/mbc.8.11.2291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu WJ, Erickson JW, Lin R, Cerione RA. The gamma-subunit of the coatomer complex binds Cdc42 to mediate transformation. Nature. 2000;405:800–804. doi: 10.1038/35015585. [DOI] [PubMed] [Google Scholar]