

Highlights
-
•
Two-year-olds start to be sensitive to syntactic cues.
-
•
Interaction of syntax and semantics is high in very young children.
-
•
The reliance on syntactic cues shifts between the age of two and three.
Abstract
Sentence comprehension requires the assignment of thematic relations between the verb and its noun arguments in order to determine who is doing what to whom. In some languages, such as English, word order is the primary syntactic cue. In other languages, such as German, case-marking is additionally used to assign thematic roles. During development children have to acquire the thematic relevance of these syntactic cues and weigh them against semantic cues. Here we investigated the processing of syntactic cues and semantic cues in 2- and 3-year-old children by analyzing their behavioral and neurophysiological responses. Case-marked subject-first and object-first sentences (syntactic cue) including animate and inanimate nouns (semantic cue) were presented auditorily. The semantic animacy cue either conflicted with or supported the thematic roles assigned by syntactic case-marking. In contrast to adults, for whom semantics did not interfere with case-marking, children attended to both syntactic and to semantic cues with a stronger reliance on semantic cues in early development. Children’s event-related brain potentials indicated sensitivity to syntactic information but increased processing costs when case-marking and animacy assigned conflicting thematic roles. These results demonstrate an early developmental sensitivity and ongoing shift towards the use of syntactic cues during sentence comprehension.
1. Introduction
A fundamental step in language development is to learn the thematic relations indicated between individual words. Successful sentence comprehension depends on the understanding of thematic relations between the verb and its arguments in order to reveal who is doing what to whom. In linguistic terms, the who carries the thematic role of an agent and the whom that of a patient (Dowty, 1991). There are various means of encoding thematic roles across different languages. While in English the sentence-initial position of the noun phrase (NP) marks the role of an agent, other languages, such as German, additionally use unambiguous case-marking as a valid cue for the interpretation of thematic hierarchy (MacWhinney, Bates, & Kliegl, 1984). In German the nominative form der (.NOM) of the definite article preceding the noun in the NP indicates the subject and thereby the agent, while the accusative form den (.ACC) denotes the patient role, independent of word order. This is exemplified below in (a) the subject-first (Subject-Verb-Object, SVO) version and in (b) the object-first (Object-Verb-Subject, OVS) version of the following sentence.
-
(a)
Der.NOM Junge trinkt den.ACC Saft./The boy is drinking the juice.
-
(b)
Den.ACC Saft trinkt der.NOM Junge./The boy is drinking the juice.
In these example sentences the agent and patient roles are additionally indicated by the semantic cue of animacy with the animate noun taking the agent role and the inanimate noun taking the patient role. Case-markers for masculine nouns in German even allow a clear interpretation when no semantic cue is available, i.e., when both NPs are animate, as in (c) the SVO version and (d) the OVS version of the following sentence.
-
(c)
Der.NOM Junge grüßt den.ACC Lehrer./The boy is greeting the teacher.
-
(d)
Den.ACC Lehrer grüßt der.NOM Junge./The boy is greeting the teacher.
Conflicts in interpretation, however, may occur when an argument which is marked .NOM, signaling the agent role is inanimate, thereby also suggesting a patient role. Such semantically implausible sentences are syntactically correct and syntactically processable in principle. Thus, pitting syntactic against semantic cues would allow investigation of the weighting of syntactic and semantic information during sentence processing. This is of particular interest as weight of these cues may differ across early development as the developmental trajectories for the prioritization of semantic or syntactic cues may differ with syntactic processes being mastered somewhat later than semantic processes (for a review see Skeide and Friederici, 2016). Behavioral and neurophysiological studies suggest that syntactic knowledge appears to emerge between the age to 2 and 3 years (Oberecker et al., 2005, Oberecker and Friederici, 2006, Höhle et al., 2014). Semantic knowledge of single words starts to develop within the first year of life (Bergelson and Swingley, 2012) as does word learning and generalization, as indicated by event-related potential (ERP) measures (Friedrich et al., 2015, Friedrich and Friederici, 2017). Adult-like ERP effects reflecting semantic processes for single words are observed around the age of 14–17 months (Friedrich and Friederici, 2005a, Friedrich and Friederici, 2005b). At the sentence level, adult-like ERPs are found for semantic incongruity between words by the end of the second year of life (Holcomb et al., 1992, Hahne et al., 2004, Friedrich and Friederici, 2005c).
The current study investigated the developmental trajectory of how 2- and 3-year-old children use the syntactic cue of case-marking in the presence of semantically plausible and implausible cues (animacy) for sentence interpretation using ERP measures in addition to behavioral measures. In developmental science ERPs are the ultimate neurophysiological method as they allow for the monitoring of the brain’s activation on-line as a sentence is processed. Other neurophysiological methods, such as the time-sensitive magnetencephalography, are more difficult to apply in children, while functional resonance imaging (fMRI) has insufficient temporal resolution. Here we are manipulating the factors syntactic structure (SVO/OVS indicated by case markers) and semantic plausibility (animate/inanimate agent) as this allows us to uncover the possible interaction of syntactic and semantic aspects in sentence processing during early language development. We will now briefly review the relevant literature concerning case-marking in adults before moving to developmental studies.
2. Processing of case markers in adults
In behavioral studies adult German listeners show evidence of a subject-first processing preference (Gorrell, 2000) and a somewhat higher error rate in object-first sentences (e.g., Rösler et al., 1998, Bornkessel et al., 2002b, Grewe et al., 2007). This difficulty is also reflected in neurophysiological measurements, be it ERPs (e.g., Bornkessel et al., 2002b, Bornkessel and Schlesewsky, 2006, Schipke et al., 2012) or in functional magnetic resonance imaging (fMRI) data (Knoll et al., 2012, Skeide et al., 2014). In these studies the processing of case-marked OVS sentences was compared to SVO sentences.
Adult ERP studies of sentence processing that investigated case-marking in German sentences, without additional semantic cues, have reported a frontal negativity (100–450 ms) for the sentence-initial NP in response to the processing of case-marked, object-first sentences (Bornkessel et al., 2002b, Schipke et al., 2012). This effect has been called ‘topicalization’ or ‘scrambling’ negativity (Bornkessel & Schlesewsky, 2006). FMRI studies in adults, investigating the processing of case-marked, object-first sentences compared to subject-first sentences, reported increased activation in the left inferior frontal gyrus (IFG), in particular in the posterior portion of Broca’s area, BA 44 (Sakai, 2005, Friederici, 2011, Skeide et al., 2014). Thus, there are clear indications of increased processing cost for case-marked, object-first compared to case-marked, subject-first sentences in adults, as indicated by neuroscientific data (Bornkessel et al., 2002b, Grewe et al., 2007).
3. Processing case markers in children
There are a number of behavioral and neurophysiological studies on the processing of case markers during development. Most of these studies used so-called semantically reversible sentences, with two animate noun arguments in either an SVO or OVS structure in order to investigate the processing of syntactic information in the absence of semantic information.
Behavioral experiments have indicated that in semantically ambiguous sentences children do not consistently use the available syntactic cue of case-marking until the age of 7 years (Schaner-Wolles, 1989, Schipke et al., 2012), although they show above chance performance by the age of 3 years for subject-first sentences. In case-marked, subject-initial sentences both the word order and the case-marking provide information about the subject NP. In OVS sentences, however, the subject NP is sentence final. The latter creates a conflict between a canonical subject-first word order and the case-marking. In a sentence picture-matching task 2;7-year-old German children performed well on SVO sentences, that is, on sentences with canonical word order, but not on OVS sentences which have a non-canonical word order (Dittmar, Abbot-Smith, Lieven, & Tomasello, 2008). In an acting out task, Dittmar et al. (2008) tested German children at the ages of 2;0, 5;0 and 7;0 and found that for 2-year-olds, both word-order and case-marking cues were necessary to correctly mimic the action encoded in a test sentence. Five-year-olds were able to use the word-order cue alone, but not case-marking alone. Seven-year-olds were eventually able to assign correct thematic roles based on the case-marking cue alone. Similar behavioral findings were obtained for children aged 3;0, 4;6, and 6;0 years by Schipke et al. (2012). In this study, which used sentences with two animate nouns, the 3- and 6-year-old children performed at chance level for the object-first sentences whereas adults performed close to 100% correct. For subject-first sentences behavioral performance steadily increased with age from 64.72% to 96.39% (Schipke et al., 2012). In a more recent study, using similar stimulus material, the performance of 3-year-olds was considered at the individual level. It was found that 11 out of 21 children performed above chance for object-first sentences whereas 10 performed at chance (Vissiennon, Friederici, Brauer, & Wu, 2017). Although indicating high behavioral variability around this age, these data provide the first evidence that children, even from the age of 3 years, start to use case-marking as a possible cue to help sentence interpretation. In a study bringing word order and animacy into conflict, Chan, Lieven, and Tomasello (2009) showed an increase in reliance on word order from 2;6 to 3;6 to 4;6. The latter age group displayed the highest use of word order whereas the youngest age group understood only prototypical transitive sentences with a coalition of the animacy and word-order cue.
Neurophysiological studies in children, both neuroimaging and ERP, reveal interesting brain-related changes which can be related to the processing of semantically ambiguous, case-marked SVO and OVS sentences across development. An fMRI study with German children by Knoll et al. (2012) revealed an adult-like activation in the left IFG, housing Broca’a area, for OVS compared to SVO sentences in 4;8 to 6;8-year-olds, but only for children with high grammatical competence. The authors suggested that children at this age are already sensitive to the syntactic cues, but may use other strategies for sentence interpretation, for instance, relying on semantic cues instead of using case-marking. Another fMRI study varied word order and case-marking cues, again in semantically ambiguous sentences, (Vissiennon et al., 2017) and investigated the functional connectivity of the language network. They found a developmental difference in the connectivity between the posterior superior temporal cortex and different parts of Broca’s area in 3-year-olds and 6-year-olds. While at the age of 3 functional connectivity was stronger between the temporal cortex and the anterior portion of Broca’s area (BA 45), by 6 years of age the functional connectivity between the temporal cortex was stronger with the posterior portion of Broca’s area (BA 44). This distinction is interesting for two reasons. First, in the adult brain BA 45 is thought to support semantic and thematic aspects of processing, whereas BA 44 specifically processes syntax (Friederici, Chomsky, Berwick, Moro, & Bolhuis, 2017). Second, for the developing brain a functional MRI study has shown that syntactic processes segregate from semantic processes gradually (Skeide et al., 2014b). In that study, which varied sentence case-marking and semantic plausibility, the localization of case-marking only became gradually independent from semantics between the ages of 3 and 7 years. A more recent fMRI study, again investigating the processing of syntactic (case-marking) and semantic (animacy) cues during sentence comprehension (Wu, Vissiennon, Friederici, & Brauer, 2016), found developmental brain activation differences in relation to these cues. In adults, a functional dissociation of Broca's area in the left IFG was observed, with the main effect of syntactic case-marking in left BA 44 and the main effect of semantic animacy in left BA 45. In 5-year-old children, however, a main effect of animacy was evident in left BA 45 and posterior STG, whereas no main effect of case-marking was found in the left, fronto-temporal, language network. These data suggest that children at 5 years still rely mainly on semantic cues, such as animacy, during sentence comprehension.
A number of ERP studies have monitored brain activation in children and adults as a sentence is processed on-line, analyzing the first NP and the second NP separately (Schipke et al., 2012). In adults, ERPs for the processing of case-marked, object-first, semantically ambiguous sentences showed a frontal negativity (100–450 ms) for the sentence-initial NP (Bornkessel et al., 2002b, Schipke et al., 2012). A similar effect at the accusative-marked, object-first NP was found for 6-year-olds, but not for children between the ages of 3 and 6 years. Children at the age of 3 instead showed a late posterior positivity at the object-initial NP, which was interpreted as reflecting general processing difficulties with accusative-marked, sentence-initial NPs in the absence of semantic cues (Schipke et al., 2012).
The goal of the present study was to investigate the development of the functional independence of syntactic processes from semantic cues by systematically crossing these two aspects in the experiment. The ERP method was applied to permit a fine-grained analysis of processes occurring on-line as sentences unfold and to allow syntactic and semantic processes, within each NP, to be analyzed separately. This is possible because in German the NP determiner which carries the syntactic case marking feature is perceived prior to the following noun which carries the semantic feature of animacy.
4. The present study
Children and adults were presented with sentences in which we manipulated both the syntactic cue of case-marking (hereafter CM) and the semantic cue of animacy that congruently and incongruently assigned thematic roles to verb arguments in SVO and OVS sentences. The relative use of morphological cues, semantic cues, and word order for thematic role assignment has been systematically discussed for different languages in the Competition Model by Bates and MacWhinney (1989). In their model, it is proposed that during sentence comprehension different cue types are used depending on the validity and reliability of a given cue in a particular language. Moreover, the model predicts that coalitions of cues facilitate comprehension, whereas conflicting cues lead to processing costs. We used the model of Bates and MacWhinney to formulate the predictions for the current study.
At the behavioral level we expected the non-conflicting semantic cue to improve children’s comprehension of complex syntactic structures, leading to good performance even in OVS sentences. We hypothesized that children would have difficulties interpreting sentences when semantic cues were in conflict with CM cues. This effect was expected to be stronger for 2-year-olds than for 3-year-olds. Adults were expected to perform high even when cues were in conflict, as nominative case-marking in German is a reliable cue.
At the neurophysiological level predictions can only partly be based on prior work as the present study is the first to separately analyze the determiner and the noun of the relevant NPs. In the prior study by Schipke et al. (2012) the ERP analyses were conducted using the entire NP consisting of a determiner, carrying case-marking information, followed by a noun, carrying animacy information. In the present study we conducted a temporally more fine-grained analysis by separately analyzing the ERPs associated with the determiner and the subsequent animacy-carrying noun, both for the sentence-initial and sentence-final NP.
The relative timing of the availability of syntactic and semantic information is relevant because in principle the syntactic information (case) is sufficient to make a correct assignment of the respective NP’s thematic role. The subsequent semantic cue, i.e., the noun of the NP, is not necessary for thematic role assignment of case-marked NPs. A conflict between the case-marked determiner and the animacy-marked noun can only happen if the primary available syntactic case information and the secondary available semantic animacy information are processed to an equal degree, independent of the temporal availability.
For adults we predicted a clear CM effect at the determiner of the sentence-initial NP and only small effects on the same NP’s noun (Noun 1). This effect may be reflected in a negative deflection of the ERPs, as a negativity has been found previously at the first NP in OVS sentences, indicating topicalization costs (Bornkessel and Schlesewsky, 2006, Schipke et al., 2012). Moreover, if adults rely primarily on syntactic information, no interaction with the semantic factor of animacy should be found. For the sentence-final NP we predicted no CM effect on the determiner as roles are assigned already. For the noun no prediction was made, although the sentence-final, inanimate, subject-case marked noun is unexpected.
For children the expectations were different. Young children may have a hard time to process the sentence-initial determiner quickly and accurately. If so, no CM effect should be observed. Interaction effects should be found if case information is processed once the animacy information becomes available. The sentence-final NP could reflect these final interpretation processes.
The current study consisted of two experiments: a behavioral experiment (Experiment 1) and an ERP experiment (Experiment 2). These will be presented in turn.
5. Experiment 1
A behavioral picture-matching experiment investigated children’s ability to use CM by varying the syntactic structure of transitive sentences. We analyzed children’s responses to four types of sentences with unambiguously case-marked arguments, namely, SVO sentences with an animate and inanimate agent (Table 1, left), as well as OVS sentences with an animate and an inanimate agent (Table 1, right). In sentences with an animate agent, animacy was congruently mapped to thematic roles.
Table 1.
Conditions of the experiment.
| Semantics | Syntax |
|
|---|---|---|
| SVO | OVS | |
| Animate Agent |
Der Hund schiebt den Topf. the.NOM dog pushes the.ACC pot “The dog is pushing the pot.” |
Den Topf schiebt der Vogel the.ACC pot pushes the.NOM bird “The bird is pushing the pot.” |
| Inanimate Agent |
Der Keks schiebt den Esel. the.NOM biscuit pushes the.ACC donkey “The biscuit is pushing the donkey.” |
Den Igel schiebt der Schrank. the.ACC hedgehog pushes the.NOM wardrobe “The wardrobe is pushing the hedgehog.” |
In sentences with an inanimate agent, semantic features conflicted with typological hierarchy of thematic roles. Children were also presented with SVO and OVS sentences that contained two animate arguments (one third of the presented items) that were not analyzed in the present study.
5.1. Methods
5.1.1. Participants
The behavioral task was conducted with eighty-nine 2-year-old and fifty-six 3-year-old children. All children were tested with the “Test zum Satzverstehen von Kindern” (TSVK; Siegmüller, Kauschke, van Minnen, & Bittner, 2010) that assesses sentence comprehension using a picture-pointing procedure. Children were tested with age-appropriate versions of the TSVK. While 2-year-olds received 12 test items involving the comprehension of simple sentences with active verbs, 3-year-olds were tested using 36 items that included passive and reflexive verbs, past tense, word order variations and object-relative clauses. Several datasets from 2-year-olds had to be excluded from analyses due to: failure to respond to the training trials (n = 2), self-distraction or lack of motivation to complete the task (n = 30), history of hearing impairment or neurological disorder (n = 5), bilingual family environment (n = 1), one-side bias in pointing behavior (n = 9, calculated as pointing to one side in more than 80% of the trials), or missing data from the language development test (n = 1). Datasets from 3-year-olds were excluded from the analysis due to a history of hearing impairment (n = 4), bilingual family environment (n = 1), missing data (n = 9), below-norm performance in language development test (n = 1), failure to complete the test (n = 2), or one-side bias in pointing behavior (n = 4). After this exclusion procedure, a total of forty-one datasets from 2-year-old children (age range 24–35 months, M = 29.71 months, 23 girls) and thirty-five datasets from 3-year-old children (age range 37–47 months, M = 41.89 months, 20 girls) entered the analysis.
5.1.2. Materials
Sixty transitive sentences with the following structure [determiner – noun – verb – determiner – noun] were constructed using six verbs (schieben “to push”, ziehen “to pull”, werfen “to throw”, beißen “to bite”, treten “to kick”, hauen “to hit”), 11 nouns (Hund “dog”, Esel “donkey”, Vogel “bird”, Fuchs “fox”, Igel “hedgehog”, Tiger “tiger”, Keks “cookie”, Kuchen “cake”, Schrank “wardrobe”, Topf “pot”, Turm “tower”) and two determiners (der, den). All nouns were of masculine syntactic gender, because in German only these allow unambiguous CM. Lexical material was chosen by consulting a standard language test SETK-2 (Grimm, Aktas, & Frevert, 2000) and the data of 2-year-olds from the CHILDES corpus (MacWhinney, 2000).
Pre-recorded stimuli were used to maintain a standard experimental procedure. A trained female native speaker recorded the auditory experimental material in a child-directed manner. After recording, sentences were digitized (44.1 kHz/16bit sampling rate, mono). These sentences were also used in the ERP experiment (Experiment 2) and required additional editing. Since no processing efforts should take place at the baseline, the editing included inserting a 50-ms silence period at the beginning and the end of each sentence to eliminate the presence of irrelevant acoustic material. Sentences were normalized using the root mean square of the speech amplitude. Cartoon pictures were created for each individual sentence (for an example of the visual stimuli, see Supplementary Material 1). All the characters were of the same size and had similar physical features such as a body and extremities. The action was depicted by a typical motion pose of the character, e.g., keeping the paws up for a throwing action of a dog. While natural extremities were used in the case of animate actors, inanimate actors received the minimum physical features that were required for a specific action to be conveyed, such as a paw in a throwing or a kicking action. No other animal-like features, such as faces or other body parts, were added to the inanimate actors. By doing so, we ensured that it would be possible to depict the action and at the same time the inanimate character would not receive an overall animate appearance.
Each cartoon item contained two scenes that were horizontally aligned on a 210 × 297 mm sheet of paper. One scene corresponded to the stimulus sentence, whereas in the other scene the thematic roles were reversed. Right-left allocation of corresponding and not corresponding scenes was pseudo-randomized. Each child was presented with a selection of 30 (in the case of 2-year-olds) or 36 (in the case of 3-year-olds) sentences that were pseudo-randomized in several lists prior to the experiment.
5.1.3. Procedure
The behavioral experiment consisted of a short introduction, training, and testing phases. During the introduction phase, children were familiarized with animals and objects in a game-like manner. Participants were presented with the single images of animals/objects one at a time and asked to name them. If the response was correct, the experimenter just repeated the name of the animal/object. If the child recognized the object/animal but used another word (e.g., an onomatopoeia such as wauwau ‘bow-wow’ for a dog), the experimenter agreed but corrected the word, repeating the target der Hund ‘the dog’ 3 to 4 times. In case the child did not respond, the experimenter just repeated the word 3 to 4 times while pointing to the picture.
During the training phase, participants were seated on their parent’s lap. The experimenter sat either to the left or right of the child, assigned randomly. The experimenter placed a picture book in front of the child and explained that she and the child were going to play a game. The experimenter was going to tell a short story and the child would be asked to point to the corresponding picture. Two training items were used to ensure the child understood the task. Feedback was provided for the training items. These trials were excluded from evaluation.
In the testing phase that followed, the experimenter showed the picture and said: “Now, the aunty from the boxes [pointing to loudspeakers] will tell a new story. Please point to the picture that fits the story.” Stimuli were presented using Presentation (Neurobehavioral Systems, Inc, Albany, CA, USA) with the average intensity of 50 dB via loudspeakers that were located at approximately 140 cm in front of the participants.
The child was encouraged to listen to the sentence, “Now listen to the aunty…” and then point to the picture, “Show me!”. In case the child was distracted during stimulus presentation, the sentence was presented again. Children were allowed to point after the presentation was finished. The pointing task had no time limitations. Experimenters were required to avoid positive/negative evaluation of the responses. Parents were only solicited for motivational assistance if necessary. They were asked to refrain from repeating sentences and giving verbal or nonverbal feedback throughout the experiment. Trials were not analyzed if these requirements were not fulfilled.
5.1.4. Data analysis
When the child pointed to the scene that corresponded to the sentence they heard it was counted as a correct response. Only unambiguous responses were scored. In case the child pointed at both pictures, did not listen to the story attentively, or did not look at the pictures, the answer was considered a miss. Accordingly, 51 of 820 trials were excluded from evaluation in 2-year-olds and 16 of 840 in 3-year-olds (for details, see Supplementary Material 2). Differences in the accuracy of responses were evaluated using repeated measures analysis of variance (ANOVA) with factors for Syntax [SVO, OVS], Agent Animacy [Animate, Inanimate], and Age [2-year-olds, 3-year-olds]. Following previous research, the accuracy data were also analyzed in relation to the chance level (50%) using a one-sample t-test.
5.2. Results
Figure 1 presents a summary of performance accuracy in the picture-matching experiment. The repeated measures ANOVA revealed a main effect of Age, F(1,74) = 17.70, p < .001, a main effect of Syntax, F(1,74) = 27.19, p < .001, and a main effect of Agent Animacy, F(1,74) = 4.28, p = .042. The interaction of Syntax and Age was significant, F(1, 74) = 4.32, p = .041. No other interaction effect reached significance.
Fig. 1.
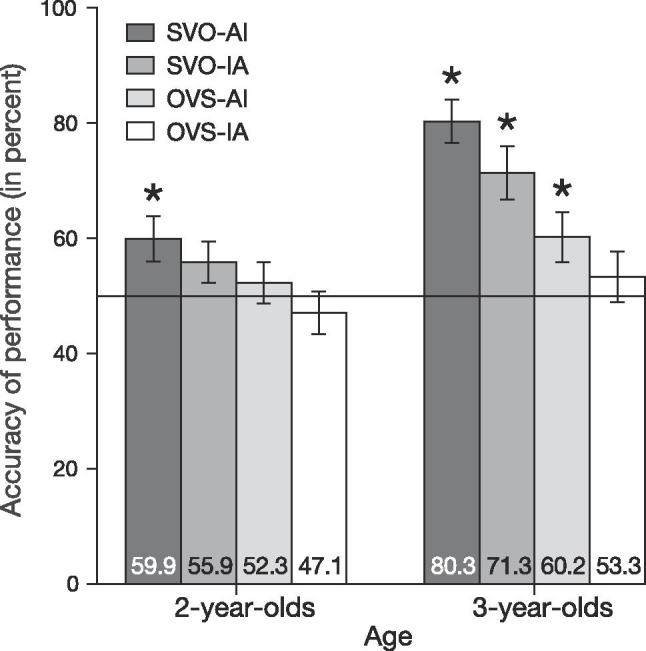
Performance accuracy of 2- and 3-year-old children in the behavioral task. Asterisk indicates significant deviation from chance level, as assessed by a one-sample t-test, p < .05. Error bars indicate one standard error (SE).
Individual planned comparisons revealed that all children performed above chance level in SVO sentences with congruent animacy (2-year-olds: t(40) = 2.52, p = .016; 3-year-olds: t(34) = 8.09, p < .001). Three-year-old children also performed above chance level for SVO sentences with inanimate agents, t(34) = 4.62, p < .001, and OVS sentences with animate agents, t(34) = 2.38, p = .023.
5.3. Discussion
In the current picture-matching experiment, we investigated young children’s use of case information in sentences with congruent/incongruent semantic cues of animacy. Results suggest that children acquire the functional meaning of case-marking between two and three years of age. The youngest participants performed above chance on SVO sentences only when word order, CM and animacy were in coalition. This is in line with previous studies that pointed to early understanding of fully prototypical sentences in which all the cues (CM, word order, and animacy) are assigning the same thematic roles (e.g., Dittmar et al., 2008). Three-year-old children performed above chance for all sentence types except for those in which the animate object was in the sentence-initial position. Three-year-olds, however, were not distracted by the inanimate agent in SVO sentences, that is, in sentences where word order and CM indicate the same thematic role. Importantly, the 3-year-old children in our experiment performed above chance level on OVS sentences, where word order was non-canonical, but only when animacy and CM were not conflicting, i.e., assigning identical thematic roles. Three-year-olds had difficulty processing OVS sentences when the semantic cue conflicted with the CM. In other words, 3-year-olds clearly profited from the supporting semantic cue and demonstrated their awareness of the functional meaning of CM.
One might argue that 3-year-olds’ high performance in SVO sentences was exclusively driven by the word order cue, with CM having no value for sentence interpretation. However, if this were the case, 3-year-olds should have performed below chance level when word order was reversed (i.e. in OVS sentences), especially in sentences with an inanimate agent. Comparable effects were observed by Schipke et al. (2012). In their study, 3-year-old participants performed not just at chance level but significantly below, for OVS sentences with two animate nouns. This indicates that the absence of a semantic cue led young children to use word order, which resulted in systemic errors for these sentences.
The acquisition of CM is certainly far from being complete in 3-year-olds, as this age group did not exclusively rely on CM when conflicting semantic information was present. Our results suggest it is a combination of CM and semantic cue that contribute to above chance performance in complex sentences for 3-year-olds.
6. Experiment 2
In Experiment 2, we conducted an ERP study in order to identify the neural correlates of children’s syntactic comprehension abilities, as compared to adults. Experiments 1 and 2 used the same sentence material but differed in the granularity of their analyses. In Experiment 1, the congruency between thematic roles assigned by syntactic and semantic features was analyzed with respect to the entire sentence. In Experiment 2, we explored the use of syntactic and semantic features within each noun phrase. The noun-animacy and syntactic complexity were crossed yielding four types of sentences. In SVO and OVS sentences, the sentence-initial noun was animate, and the sentence-final noun was inanimate (Table 2, a/b). In SVO and OVS sentences, the first noun was inanimate and the second was animate (Table 2, c/d). The participants were also presented with SVO and OVS sentences that contained two animate arguments. These sentences were not analyzed in the present study.
Table 2.
Conditions of the ERP experiment.
| Syntax | Semantics |
|
|---|---|---|
| Animate N1 | Inanimate N1 | |
| SVO | (a) SVO Der Vogel fängt den Topf. the.NOM bird catches the.ACC pot “The bird is getting the pot.” |
(c) SVO Der Schrank trägt den Hund. the.NOM wardrobe carries the.ACC dog “The wardrobe is carrying the dog.” |
| OVS | (b) OVS Den Hund hebt der Ball. the.ACC dog lifts the.NOM ball “The ball is lifting the dog.” |
(d) OVS Den Topf tritt der Igel. the.ACC pot kicks the.NOM hedgehog “The hedgehog is kicking the pot.” |
The ERP analyses were time-locked to the onset of the two NP constituents. Each NP contained a determiner (D1/D2) and a noun (N1/N2). We will now provide our hypotheses only for the first NP. The hypotheses regarding the second NP will be presented after the discussion of the ERP results for NP1.
Based on previous studies, adults were expected to show a topicalization negativity at D1 (Matzke et al., 2002, Schipke et al., 2012) because the topicalization negativity was observed between 100 and 400 ms after NP onset, which starts with the determiner. The results of Experiment 1 demonstrated that 3-year-olds did not completely acquire the functional meaning of CM but relied on a combination of syntactic and semantic information. This suggests that neither 2-year-olds nor 3-year-olds would show an adult-like ERP response at D1.
On the basis of Schipke et al. (2012), who found a late positivity between 700 and 800 ms after the onset of the first NP in their 3-year-olds, we argued that our participants within the same age group would be sensitive to syntactic information at the sentence-initial position. Therefore, we assumed that OVS sentences would elicit a similar positivity at N1, at least in the group of 3-year-olds. Integration of syntactic information from CM with semantic information from the noun appears to take place at N1. Since preschool children were shown to use semantic information in Experiment 1, we expected to observe an increase in processing cost if D1 and N1 assigned different thematic roles through N1 animacy cue.
6.1. Methods
6.1.1. Participants
Ninety-six 2-year-olds and fifty-seven 3-year-olds participated in the ERP study. Informed parental consent was obtained for all children before the experiment. In addition to the TSVK (Siegmüller et al., 2010) for both age groups, parents of 2-year-olds also completed the vocabulary and grammar subcomponents of the FRAKIS questionnaire (Szagun, Stumper, & Schramm, 2009).
EEG recording was not possible for seven 2-year-old and two 3-year-old children. We excluded the recordings of several 2-year-olds due to: poor quality of the EEG data, i.e., at least 15 artefact-free trials for each condition (n = 20), history of neurological or hearing disease (n = 5), bilingual family environment (n = 1), below-norm language development in one of the FRAKIS subtests (n = 11), missing FRAKIS data (n = 3), or missing TSVK data (n = 1). Several datasets of the 3-year-olds were excluded due to: poor quality EEG data (n = 8), neurological or hearing disease history (n = 4), bilingual environment (n = 1), missing data (n = 8), or below-norm performance on TSVK (n = 1). The final datasets consisted of EEG recordings from forty-eight 2-year-old (age range 24–35 months, M = 29 months, 24 girls) and thirty-three 3-year-old children (age range 37–47 months, M = 41.76 months, 18 girls).
Thirty-four adults took part in the present experiment (age range 21–35 years, M = 27.32 years, 15 female). They were all healthy, native German speakers. Three participants were excluded from the analysis due to a lack of reliable EEG data (n = 2) and technical problems during the recording (n = 1). The final dataset consisted of 31 EEG recordings (age range 21–35 years, M = 27.13 years, 14 female).
6.1.2. Materials
Stimulus material for the ERP experiment consisted of 120 critical trials. These included the 40 sentences that were used in the behavioral experiment and 80 additional sentences that were created using the same nouns and six new verbs (kneifen “to pinch”, heben “to lift”, fangen “to catch”, kratzen “to scratch”, tragen “to carry”, schlagen “to beat”). The stimuli were balanced such that each referent took the agent/patient role and appeared together with each verb equally often. Participants were also presented with 30 SVO and 30 OVS sentences that contained two animate arguments, which were not analyzed in the present study.
6.1.3. Procedure
Prior to the ERP experiment, the animals and objects featured in the stimuli were introduced to the children. For the EEG recordings participants were seated on their parent’s lap in an electrically shielded cabin. During the presentation of the stimuli, a silent cartoon film was shown which did not involve any animal or action present in the experimental stimuli. Stimuli were presented acoustically using Presentation (Neurobehavioral Systems, Inc, Albany, CA, USA) with the average intensity of 50 dB and an interstimulus interval (stimulus offset to stimulus onset) of 2500 ms. Children were instructed to listen to the sentences and to carefully pay attention to “who is doing what to whom”, and they were reminded that there would be a post-experiment test about the sentences to which they were about to listen. Parents wore earplugs and were instructed to avoid communicating with the child. The experiment was divided into two blocks of 10 min. Additional breaks were taken if necessary.
EEG data was recorded at 129 electrode sites using Geodesic Sensor Nets (Electrical Geodesics, Inc., Eugene, OR, USA) with an operating impedance of 50 kΩ. The data were digitized online at a rate of 500 Hz and referenced to the Cz electrode.
6.1.4. Data analysis
Electrophysiological data were processed using the software tools EEGLAB (Delorme and Makeig, 2004) and ERPLAB (Lopez-Calderon & Luck, 2014). The EEG data were down-sampled offline to 250 Hz, band-pass filtered between 0.3 and 20 Hz, and re-referenced to the linked mastoids (electrode sites 57 and 100). Algorithms exploiting Independent Component Analysis were used to correct stereotyped artifacts such as eye movements. Epochs, time-locked to the onset of the two determiners (500 ms) and the two nouns (1000 ms), were extracted (see Fig. 2). Due to the absence of a condition-independent time period, suitable for each constituent during auditory sentence presentation, no pre-stimulus baseline correction was applied (for discussion and similar approach, see Friederici et al., 2000, Wolff et al., 2008, Maess et al., 2016). Only completely artifact-free sentences were included in the individual ERP generation.
Fig. 2.
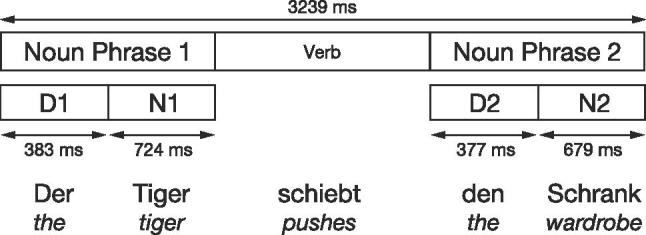
The average duration (in ms) of the sentence and its constituents. ERP measures were time locked, separately, to the determiner (D1, D2) and the subsequent noun (N1, N2).
Statistical analyses were performed on the mean amplitudes of consecutive time windows (TWs) of 50 ms as in previous studies (Gunter et al., 1997, Hahne and Friederici, 2001, Hoeks et al., 2004, Diaz et al., 2011, Schipke et al., 2012). The determiners were analyzed using 10 TWs between 0 and 500 ms (0–50 ms, 50–100 ms, etc). The nouns were analyzed in 20 TWs between 0 and 1000 ms. Nine regions of interest were defined in anterior-posterior (AP) and lateral (LP) planes: anterior-left, anterior-middle, anterior-right, central-left, central-middle, central-right, posterior-left, posterior-middle, and posterior-right.
The impact of syntactic and semantic cues on sentence processing was tested using a repeated measures ANOVA with factors for Syntax [SVO, OVS], N1 Animacy [animate, inanimate], and distributional factors AP [anterior, central, posterior] and LP [left, middle, right]. Corrections using the Greenhouse-Geisser method were applied. To correct for type I errors, effects were considered only if they were confirmed in at least two consecutive TWs. Voltage topographies were computed using mean amplitudes between the first and the last TW with reliable effects.
6.2. Results
6.2.1. Noun phrase 1
Since we were primarily interested in the use of syntactic cues, and their interaction with animacy, but not in the main effects of animacy per se, the main effects of animacy are reported in Supplementary Material 3.
6.2.2. Determiner 1
When comparing SVO and OVS sentences we see that the first determiner of the OVS structures “den” elicited an anterior-central negative response in adults (Fig. 3) that was confirmed by ANOVA between 250 and 400 ms (Table 3). The ANOVA revealed no main effects of Syntax in either 2- or in 3-year-old children.
Fig. 3.

ERP response to SVO and OVS sentences relative to (A) the onset of D1 in adults (upper panel), 3-year-olds (middle panel), and 2-year-olds (bottom panel), (B) the onset of N1 in adults (upper panel), 3-year-olds (middle panel), and 2-year-olds (bottom panel).
Table 3.
Syntax effects as calculated at D1 for adults.
| TW | Main effect |
Area-specific effect |
|||||
|---|---|---|---|---|---|---|---|
| df | F | p | Area | df | F | p | |
| 250–300 | 1, 30 | 10.38 | 0.003 | anterior, central | 1, 30 | ≥8.78 | ≤0.006 |
| 300–350 | 1, 30 | 20.40 | <0.001 | ||||
| 350–400 | 1, 30 | 13.10 | 0.001 | AM, AR, CL, CM, CR, PL | 1, 30 | ≥4.55 | ≤0.041 |
Note. TW, time window; AM, anterior-middle; AR, anterior-right; CL, central-left; CM, central-middle; CR, central-right; PL, posterior-left.
6.2.3. Noun 1
The processing of the first noun of the OVS relative to SVO sentences was associated with an early positivity in all three age groups (Fig. 3). Whereas the ANOVA substantiated the anterior-central effect of Syntax in adults between 150 and 400 ms, the early positivity in 3-year-olds started around the word onset and lasted until 200 ms (Table 4). A similar effect was observed in 2-year-olds. This was statistically confirmed in the left scalp area between 50 and 100 ms, and over the whole scalp between 100 and 200 ms.
Table 4.
Syntax effects as calculated at N1 for adults, 3-year-olds, and 2-year-olds.
| TW | Main effect |
Area-specific effect |
|||||
|---|---|---|---|---|---|---|---|
| df | F | p | Area | df | F | p | |
| Adults | |||||||
| 150–200 | 1, 30 | 9.03 | 0.005 | ||||
| 200–250 | 1, 30 | 6.95 | 0.013 | anterior, central | 1, 30 | ≥7.51 | ≤0.010 |
| 250–300 | 1, 30 | 7.17 | 0.012 | ||||
| 300–350 | 1, 30 | 6.45 | 0.017 | ||||
| 350–400 | 1, 30 | 7.49 | 0.010 | ||||
| 3-year-olds | |||||||
| 0–50 | 1, 32 | 4.62 | 0.039 | ||||
| 50–100 | 1, 32 | 5.10 | 0.031 | ||||
| 100–150 | 1, 32 | 14.01 | 0.001 | ||||
| 150–200 | 1, 32 | 10.91 | 0.002 | ||||
| 2-year-olds | |||||||
| 50–100 | left | 1, 47 | 6.38 | 0.015 | |||
| 100–150 | 1, 47 | 8.25 | 0.006 | anterior, central | 1, 47 | ≥7.21 | ≤0.010 |
| 150–200 | 1, 47 | 4.99 | 0.030 | anterior | 1, 47 | 7.96 | 0.007 |
| 150–200 | left, middle | 1, 47 | ≥5.13 | ≤0.028 | |||
Note. TW, time window.
Crucially, interactions involving Animacy, Syntax, and distributional factors were observed at N1 in both developmental groups but not in adults. In 3-year-olds, such interactions between 100 and 300 ms were primarily driven by animacy differences in OVS sentences with animate object nouns evoking a more negative ERP response. Animate nouns – often considered to be agents – are unexpected in sentence-initial OVS sentences. ERPs to inanimate and animate nouns differed significantly in OVS conditions in all TWs between 100 and 300 ms (Table 5). The analysis of animacy effects in SVO sentences revealed no significant differences.
Table 5.
Interactions between Syntax and Animacy as calculated at N1 for 3- and 2-year-olds.
| TW | Interaction effects |
Effects of animacy in SVO and OVS |
||||||
|---|---|---|---|---|---|---|---|---|
| Effect | df | F | p | df | F | p | ||
| 3-year-olds | ||||||||
| 100–150 | S × A × AP | 1.7, 52.7 | 6.03 | 0.007 | OVS: cent | 1, 32 | 4.76 | 0.037 |
| 150–200 | S × A | 1, 32 | 6.17 | 0.018 | OVS | 1, 32 | 5.52 | 0.025 |
| 150–200 | S × A × AP | 1.6, 52.5 | 7.83 | 0.002 | OVS | 1, 32 | 5.47 | 0.026 |
| 200–250 | S × A | 1, 32 | 4.91 | 0.034 | OVS | 1, 32 | 11.88 | 0.002 |
| 200–250 | S × A × AP × LP | 4, 128 | 2.44 | 0.050 | OVS | 1, 32 | 11.90 | 0.002 |
| OVS: ant, cent | 1, 32 | ≥11.10 | ≤0.002 | |||||
| 250–300 | S × A × AP × LP | 3.1, 100.1 | 2.66 | 0.050 | OVS: AL, CL, CM | 1, 32 | ≥4.19 | ≤0.049 |
| 2-year-olds | ||||||||
| 0–50 | S × A × AP | 1.3, 60.7 | 4.60 | 0.027 | SVO | 1, 47 | 4.50 | 0.039 |
| 50–100 | S × A × AP | 1.4, 65.3 | 6.85 | 0.006 | SVO | 1, 47 | 6.46 | 0.014 |
| SVO: ant, cent | 1, 47 | ≥6.19 | ≤0.017 | |||||
| 300–350 | S × A × AP | 1.5, 71.0 | 4.09 | 0.031 | OVS: ant | 1, 47 | 8.68 | 0.005 |
| 350–400 | S × A × AP | 1.6, 74.1 | 6.61 | 0.005 | OVS: ant | 1, 47 | 5.40 | 0.024 |
| 400–450 | S × A | 1, 47 | 8.69 | 0.005 | SVO | 1, 47 | 6.02 | 0.018 |
| 400–450 | S × A × AP × LP | 3.1, 145.1 | 3.13 | 0.026 | SVO | 1, 47 | 6.04 | 0.018 |
| OVS: post | 1, 47 | 8.12 | 0.006 | |||||
| 450–500 | S × A | 1, 47 | 21.01 | <0.001 | SVO | 1, 47 | 15.51 | <0.001 |
| 500–550 | S × A | 1, 47 | 7.71 | 0.003 | SVO | 1, 47 | 14.12 | <0.001 |
| 550–600 | S × A | 1, 47 | 3.41 | 0.018 | SVO | 1, 47 | 7.62 | 0.008 |
Note. TW, time window; S, Syntax; A, Animacy; AP, anterior-posterior plane; LP, lateral plane; AL, anterior-left; CL, central-left; CM, central-middle; ant, anterior; cent, central.
In 2-year-olds, interactions between Syntax, Animacy, and distributional factors were found between 0 and 100 ms and 300 and 600 ms (Table 5). Separate analyses of syntactic conditions showed reliable effects of animacy between 0 and 100 ms and 400 and 600 ms in SVO but not in OVS sentences.
6.3. Discussion: Noun phrase 1
The current ERP experiment compared 2- and 3-year-olds’ online processing of syntactic cues in the presence of semantic cues, to adults. We will first discuss the ERP findings at D1 and N1 before proceeding to NP2 data.
The analysis of D1 for adult participants, as expected, showed an early negative effect with anterior-central distribution (250–400 ms) for processing OVS structures. This result is in agreement with experiments on adults’ processing of syntactic complexity (Rösler et al., 1998, Matzke et al., 2002, Schlesewsky et al., 2003, Schipke et al., 2012). Our data show that adults recognize the structure of an OVS sentence as early as at the first case-marked determiner. This was not the case in either group of children.
The analysis of N1 revealed two important findings. First, OVS sentences elicited an early positivity in all age groups. Comparable effects, in the absence of semantic cues, were argued to represent sensitivity to CM in 3-year-old children (Schipke et al., 2012). In the current data, the positivity peaked within the P1-P2 range, following a typical developmental pattern (Ponton et al., 2000, Čeponienė et al., 2005). Due to the very early onset of the effect and the absence of a baseline prior to the noun, we assume that the positivity was triggered by the differences between the two D1 forms, pointing to a slow processing of the case information carried by the determiner. The P2 finding is in agreement with previous interpretations of P2 as an independent, attention-related component that is associated with stimulus awareness (Crowley and Colrain, 2004, Čeponienė et al., 2005). Thus, the early positivity in our data may be associated with an attention-related process due to the unexpected, accusative-marked determiner in the sentence-initial position.
Second, analyses of N1 demonstrated that children do not only detect an accusative determiner, but actually use syntactic information for sentence interpretation. In 3-year-olds, the negative semantic effect between 100 and 300 ms at N1 was larger in OVS sentences as compared to SVO sentences. The observed ERP effect appears to reflect a mismatch between the thematic role assignments that are triggered by syntactic and semantic cues. The processing of the accusative-marked argument, that assigns a non-agentive role to the noun, conflicts with the agent role assigned by semantic features of the noun, as in Den Tiger…/The.ACC tiger…. Similar effects of animacy have been reported in the N400 range in previous experiments with English-speaking (Weckerly and Kutas, 1999) and German-speaking adults (Frisch and Schlesewsky, 2001, Schlesewsky and Bornkessel, 2004, Schlesewsky and Bornkessel-Schlesewsky, 2009). In the latter studies, the ERP epoch was time-locked to the NP onset, in contrast to the noun onset, as was done in the present analysis. Considering the mean duration of the first determiner (ca. 380 ms), we suggest that the negativity observed in 3-year-olds reflects the increasing cost for processing conflicting cues. When interpreting NP1 data, it is plausible to conclude that 3-year-olds consider CM information of the determiner but also attend to the semantic information provided by the noun. The observed effects cannot be attributed to a reliance on other cues, such as animacy or word order, since both strategies assign NP1 the role of the agent.
In 2-year-olds, sentence-initial, inanimate nouns elicited a greater positivity than animate nouns, between 400 and 600 ms in SVO sentences but not in the OVS sentences. Late positivities in the P600 range have been reported in previous studies on adult processing of animacy-based thematic violations (e.g., Hoeks et al., 2004, Kim and Osterhout, 2005, Kuperberg et al., 2007). Most of these studies focused on the anomalies within verb-argument dependencies and interpreted the late positivity as being elicited by a conflict between the processing streams assessing syntactic, semantic, and/or thematic information. Similar to these conflict-based models of interpretation, we assume that the observed positive ERP effect reflects a mismatch between the thematic role assignments that are triggered by CM and animacy. The conflict arises due to the agentive role assigned by the nominative CM and the non-agentive role assigned by semantic features of the noun, as in Der Schrank…/The.NOM wardrobe…. We take this interaction in role assignment to indicate an awareness, in 2-year-olds, of the functional meaning of the nominative case marker. One could argue that the increasing processing cost stems from the conflict between animacy and word order, without any impact of CM. However, if 2-year-olds actually ignored CM, similar animacy effects should have been observed at N1 in OVS sentences. Since that was not the case in our data, we suggest that 2-year-old participants were sensitive to CM at the sentence-initial constituent.
Given the differences between adults and children in their processing of NP1, it is reasonable to assume that there are also developmental differences in the processing of NP2. The analysis of the first NP revealed that semantics does not intervene with syntax in adults. The adult sentence comprehension system recognizes the syntactic structure immediately at D1. Based on the present D1 data and the results of Schipke et al. (2012), we hypothesized that adults would not require the additional retrieval of syntactic information to process the remaining noun phrase, i.e., no effects were expected at the determiner in NP2.
In contrast, children process D1 and integrate the obtained grammatical information with semantic features of N1. As shown in Experiment 1, 3-year-olds comprehend OVS sentences in which syntactic and semantic cues are congruent. Taken together, the behavioral and the first noun phrase ERP data suggest that 3-year-olds may require additional syntactic information to successfully process the remainder of the sentence. We predicted that, contrary to adults, 3-year-olds systematically consider CM at D2. Since 2-year-olds were only able to comprehend simple SVO sentences in Experiment 1, we did not expect them to show such an effect.
6.4. Results: Noun phrase 2
6.4.1. Determiner 2
For adults, as expected, no significant difference between OVS and SVO sentences was observed at the second determiner. For children, the nominative case marker at D2 in the OVS sentences elicited a positive, anteriorly-distributed ERP response between 200 and 300 ms, in comparison to the canonical accusative case marker, in both 2- and 3-year-olds (Fig. 4, Table 6).
Fig. 4.

ERP response to SVO and OVS sentences relative to (A) the onset of D2 in adults (upper panel), 3-year-olds (middle panel), and 2-year-olds (bottom panel), (B) the onset of N2 in adults (upper panel), 3-year-olds (middle panel), and 2-year-olds (bottom panel).
Table 6.
Syntax effects as calculated at D2 for 3- and 2-year-olds.
| TW | Main effect |
Area-specific effects |
|||||
|---|---|---|---|---|---|---|---|
| df | F | p | Area | df | F | p | |
| 3-year-olds | |||||||
| 200–250 | anterior, central | 1, 32 | ≥4.83 | ≤0.035 | |||
| 250–300 | 1, 32 | 8.32 | 0.007 | ||||
| 2-year-olds | |||||||
| 200–250 | anterior | 1, 47 | 5.17 | 0.028 | |||
| 200–250 | AL, AM, AR, CM | 1, 47 | ≥4.04 | ≤0.05 | |||
| 250–300 | anterior | 1, 47 | 5.44 | 0.024 | |||
| 400–450 | 1, 47 | 15.97 | <0.001 | anterior, central | 1, 47 | ≥15.13 | <0.001 |
| 450–500 | 1, 47 | 4.79 | 0.034 | anterior, central | 1, 47 | ≥5.14 | ≤0.028 |
Note. TW, time window; AL, anterior-left; AM, anterior-middle; AR, anterior-right; CM, central-middle.
6.4.2. Noun 2
At the second noun, processing of OVS sentences was associated with an early negativity in all age groups (Fig. 4). This observation was substantiated by a significant main effect of Syntax between 150 and 300 ms for adults, between 50 and 150 ms for 3-year-olds, and between 50 and 300 ms for 2-year-olds (Table 7). The effect showed an anterior-central distribution. In 2-year-olds, the early negativity was followed by a small late positivity that only reached significance between 500 and 650 ms.
Table 7.
Syntax effects as calculated at N2 for adults, 3-year-olds, and 2-year-olds.
| TW | Main effect |
Area-specific effects |
|||||
|---|---|---|---|---|---|---|---|
| df | F | p | Area | df | F | p | |
| Adults | |||||||
| 150–200 | 1, 30 | 8.71 | 0.006 | ||||
| 200–250 | 1, 30 | 7.65 | 0.010 | ||||
| 250–300 | 1, 30 | 7.06 | 0.012 | ||||
| 3-year-olds | |||||||
| 50–100 | anterior-right | 1, 32 | 9.41 | 0.004 | |||
| 100–150 | 1, 32 | 4.39 | 0.044 | ||||
| 2-year-olds | |||||||
| 50–100 | 1, 47 | 4.48 | 0.040 | ||||
| 100–150 | 1, 47 | 7.72 | 0.008 | ||||
| 150–200 | 1, 47 | 4.24 | 0.045 | ||||
| 200–250 | 1, 47 | 10.12 | 0.003 | anterior, central | 1, 47 | ≥8.15 | ≤0.006 |
| 250–300 | right | 1, 47 | 5.01 | 0.030 | |||
| 500–550 | 1, 47 | 7.85 | 0.007 | ||||
| 550–600 | 1, 47 | 9.13 | 0.004 | ||||
| 600–650 | 1, 47 | 4.28 | 0.044 | ||||
6.5. Discussion: Noun phrase 2
We had hypothesized that adults and children would show distinct patterns of processing of the sentence-final NP. While no effects were expected in adults at D2, processing cost at this sentence constituent were predicted for 3-year-olds.
As predicted, no effects were found at D2 in adults. Sentence processing did not require additional efforts at the second determiner, which is in line with previous studies of adults (Friederici et al., 1998, Frisch et al., 2002, Schipke et al., 2012). Three-year-olds showed an anteriorly-distributed positivity for OVS compared to SVO sentences at D2. This effect might reflect the processing of the unexpected nominative form at NP2, since prototypically a patient role is expected at the end of the sentence (but cf. Schipke, Friederici, & Oberecker, 2011). Alternatively, the positivity may relate to an effort of syntactic evaluation. A similar effect, the so-called P345, was observed in adult studies with sentences that required disambiguation or thematic reanalysis (Mecklinger et al., 1995, Friederici and Mecklinger, 1996, Rösler et al., 1998, Bornkessel et al., 2002a, Bornkessel et al., 2003; cf. Silva-Pereyra, Rivera-Gaxiola, & Kuhl, 2005). In line with these studies, the early effect may be associated with the diagnosis of the need for reanalysis when the maintained subject-first interpretation has to be revised (Friederici, 1998). All in all, the observed positive response at D2 implies that 3-year-olds are sensitive to the type of determiner presented and recognize its function for sentence syntactic-thematic structure.
Interestingly, it seems that 2-year-olds were also able to consider syntactic information at D2. Moreover, our youngest participants also showed a late positivity at N2. A similar effect could be observed in 3-year-olds, although it did not reach significance. A comparable positivity has been elicited in adults with sentences that require reanalysis and reordering (Kaan et al., 2000, Fiebach et al., 2001, Friederici et al., 2001, Meyer et al., 2012), and in toddlers as a response to syntactic violations (Oberecker et al., 2005, Oberecker and Friederici, 2006). One may speculate that 2-year-olds retrieve and attempt to functionally interpret syntactic information during the processing of complex sentences. However, the results of Experiment 1 indicate that these efforts do not result in correct behavioral outcomes. This may be due to the fact that 2-year-olds have not yet acquired the full functional meaning of the accusative case. Note that the analysis of syntax-semantic interactions at N1 indicated 2-year-olds’ ability to interpret a nominative case-marker, while the grammatical meaning of the accusative case marker had been apparently ignored. Whereas 3-year-olds revise a sentence structure using their knowledge of the accusative, younger children do not appear to complete the interpretation.
Finally, the processing of the accusative D2 was associated with a positive deflection at the subsequent N2, in the same way as at NP1. Again, the positivity peaked in the P1-P2 range and showed that all age groups were sensitive to differences between case forms at both determiner positions.
From the current results, it can be concluded that young children start to become aware of the functional role of CM in thematic role assignment and attempt to use it to assign thematic roles in complex OVS sentences. However, their sentence comprehension is still strongly dependent on semantic information, in particular when information congruency can improve sentence comprehension. When semantic information is incongruent it can also heavily distract their syntactic analysis. It is only later in development that the syntactic processing system gains independence from the semantic processing system (Skeide et al., 2014b).
7. General discussion
The present study examined the early developmental trajectory of the processing of syntactic case-marking information and its possible interaction with semantic factors for the assignment of thematic roles during sentence comprehension. We hypothesized that pre-kindergarten children start to become aware of the functional meaning of syntactic cues, such as case-marking, for the assignment of thematic roles in transitive sentences between the ages of 2 and 3 years. To investigate these processes, in the presence of supporting and conflicting semantic information, the semantic factor of animacy was manipulated along with case-marking in behavioral and ERP experiments. The behavioral experiment showed that children’s sentence comprehension takes advantage of supportive semantic information. If thematic roles are congruently assigned by case-marking and animacy information, 3-year-olds but not 2-year-olds can correctly interpret accusative, case-marked, object-first structures at an above-chance level.
The ERP data in Experiment 2 allowed us to unveil the processing of case-marking and animacy information, for sentence-initial and -final NPs, in a fine-grained manner by analyzing the case-marked determiner and the animacy carrying noun in succession. For adults, the first case marker in the sentence-initial determiner was sufficient to recognize the upcoming sentence structure. Semantic information carried by the noun did not intervene with the syntactic analysis of any sentence. The ERP data of the children, on the other hand, point towards a slowly emerging awareness of case-marking information encoded in the determiner of the NPs. Three-year-olds showed awareness of the functional meaning of the sentence-initial, accusative, case-marked determiner, as they tried to integrate its syntactic meaning with the incongruent semantic information of the sentence-initial noun. They also attended to the second determiner and seemed to launch behaviorally effective reanalysis mechanisms in processing OVS sentences. Two-year-olds were not able to use the sentence-initial accusative case-marking for sentence interpretation, although they were sensitive to the differences between the nominative and accusative forms in general. Thus, our youngest participants, focused on the sentence-initial nominative determiner and appeared to attempt integrating the conflicting role assignment information coming from inanimate nouns. Moreover, they attended to the second determiner, which seemed to trigger a revision of thematic roles that, however, did not result in correct sentence interpretation.
Taken together, these results provide evidence for an early sensitivity to case-marking information that children, at the age of 2 and 3 years, are not yet able to use for unambiguous thematic role assignment. It appears that children’s ability to fully use case-marking is only present by the age of 7 years (Dittmar et al., 2008). The present behavioral findings are consistent with those of Vissiennon et al., 2017, Knoll et al., 2012, who showed that young children start to become aware of syntactic information encoded in determiners. However, both our behavioral and ERP data demonstrate that 2-year-old and 3-year-old children also attend to the lexico-semantic features of verb arguments, although their interpretation is not completely driven by this factor. Our temporally fine-grained ERP analyses indicate that children realize the unexpected, accusative-marked determiner of NP1 in the sentence-initial position, but unlike adults, do not use this information to construct the underlying OVS sentence structure. This is indicated in the ERPs by the positivity associated with the nominative, case-marked determiner of NP2 in the sentence-final position. These findings suggest that syntax-oriented strategies start to develop as early as 2–3 years of age, with the realization of the functional significance of morphosyntactic cues increasing gradually and approaching adult-like levels after the age of 6.
8. Conclusion
The present study shows that awareness of the functional meaning of syntactic cues, such as case-marking, starts to emerge between the ages of 2 and 3 years in German children. It offers the first neurophysiological insights into the processing of structurally complex sentences during the initial stages of syntax development. Both 2- and 3-year-olds are aware of the functional relevance of case-marking and begin to understand its functional meaning for sentence interpretation, although at the same time, they still attend to semantic cues for thematic interpretation. These conclusions are supported in particular by the electrophysiological findings that demonstrate the awareness of syntactic information, as well as the semantic-syntactic integration efforts, during sentence comprehension in early development.
Acknowledgments
Acknowledgements
This work was supported by a grant from the European Research Council (ERC-2010-360 AdG 269505 awarded to A.D.F.). We would like to thank Kodjo Vissiennon and Riccardo Cafiero for their help with the preparation of stimulus material and Katja Kirsche, Ulrike Barth, Anne Wienholz, and Wiebke Bruchmüller for their assistance with data collection.
Author contributions
ASF, CS, ADF, and JB conceived and planned the experiments. ASF carried out the experiments. ASF, CS, and TG analyzed the data. ASF, TG, and ADF contributed to the interpretation of results. ASF and ADF wrote the manuscript.
Footnotes
Supplementary data to this article can be found online at https://doi.org/10.1016/j.bandc.2018.09.003.
Appendix A. Supplementary material
The following are the Supplementary data to this article:
References
- Bates E., MacWhinney B. Cambridge University Press; New York: 1989. Functionalism and the competition model; pp. 3–73. [Google Scholar]
- Bergelson E., Swingley D. At 6–9 months, human infants know the meanings of many common nouns. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(9):3253–3258. doi: 10.1073/pnas.1113380109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bornkessel I., Schlesewsky M. The extended argument dependency model: A neurocognitive approach to sentence comprehension across languages. Psychological Review. 2006;113(4):787–821. doi: 10.1037/0033-295X.113.4.787. [DOI] [PubMed] [Google Scholar]
- Bornkessel I., Schlesewsky M., Friederici A.D. Beyond syntax: Language-related positivities reflect the revision of hierarchies. NeuroReport. 2002;13(3):361–364. doi: 10.1097/00001756-200203040-00022. [DOI] [PubMed] [Google Scholar]
- Bornkessel I., Schlesewsky M., Friederici A.D. Grammar overrides frequency: Evidence from the online processing of flexible word order. Cognition. 2002;85(2):B21–B30. doi: 10.1016/s0010-0277(02)00076-8. [DOI] [PubMed] [Google Scholar]
- Bornkessel I., Schlesewsky M., Friederici A.D. Contextual information modulates initial processes of syntactic integration: The role of inter-versus intrasentential predictions. Journal of Experimental Psychology-Learning Memory and Cognition. 2003;29(5):871–882. doi: 10.1037/0278-7393.29.5.871. [DOI] [PubMed] [Google Scholar]
- Čeponienė R., Alku P., Westerfield M., Torki M., Townsend J. ERPs differentiate syllable and nonphonetic sound processing in children and adults. Psychophysiology. 2005;42(4):391–406. doi: 10.1111/j.1469-8986.2005.00305.x. [DOI] [PubMed] [Google Scholar]
- Chan A., Lieven E., Tomasello M. Children's understanding of the agent-patient relations in the transitive construction: Cross-linguistic comparisons between Cantonese, German, and English. Cognitive Linguistics. 2009;20(2):267–300. [Google Scholar]
- Crowley K.E., Colrain I.M. A review of the evidence for P2 being an independent component process: Age, sleep and modality. Clinical Neurophysiology. 2004;115:732–744. doi: 10.1016/j.clinph.2003.11.021. [DOI] [PubMed] [Google Scholar]
- Delorme A., Makeig S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 2004;134(1):9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Diaz B., Sebastian-Galles N., Erdocia K., Mueller J.L., Laka I. On the cross-linguistic validity of electrophysiological correlates of morphosyntactic processing: A study of case and agreement violations in Basque. Journal of Neurolinguistics. 2011;24(3):357–373. [Google Scholar]
- Dittmar M., Abbot-Smith K., Lieven E., Tomasello M. German children's comprehension of word order and case marking in causative sentences. Child Development. 2008;79(4):1152–1167. doi: 10.1111/j.1467-8624.2008.01181.x. [DOI] [PubMed] [Google Scholar]
- Dowty D. Thematic proto-roles and argument selection. Language. 1991;67(3):547–619. [Google Scholar]
- Fiebach C.J., Schlesewsky M., Friederici A.D. Syntactic working memory and the establishment of filler-gap dependencies: Insights from ERPs and fMRI. Journal of Psycholinguistic Research. 2001;30(3):321–338. doi: 10.1023/a:1010447102554. [DOI] [PubMed] [Google Scholar]
- Friederici A.D. Kluwer Academic Publishers; Dordrecht: 1998. Diagnosis and reanalysis: Two processing aspects the brain may differentiate; pp. 177–200. [Google Scholar]
- Friederici A.D. The brain basis of language processing: From structure to function. Physiological Reviews. 2011;91(4):1357–1392. doi: 10.1152/physrev.00006.2011. [DOI] [PubMed] [Google Scholar]
- Friederici A.D., Chomsky N., Berwick R.C., Moro A., Bolhuis J.J. Language, mind and brain. Nature Human Behaviour. 2017;1:713–722. doi: 10.1038/s41562-017-0184-4. [DOI] [PubMed] [Google Scholar]
- Friederici A.D., Mecklinger A. Syntactic parsing as revealed by brain responses: First-pass and second-pass parsing processes. Journal of Psycholinguistic Research. 1996;25(1):157–176. doi: 10.1007/BF01708424. [DOI] [PubMed] [Google Scholar]
- Friederici A.D., Mecklinger A., Spencer K.M., Steinhauer K., Donchin E. Syntactic parsing preferences and their on-line revisions: A spatio-temporal analysis of event-related brain potentials. Cognitive Brain Research. 2001;11(2):305–323. doi: 10.1016/s0926-6410(00)00065-3. [DOI] [PubMed] [Google Scholar]
- Friederici A.D., Steinhauer K., Mecklinger A., Meyer M. Working memory constraints on syntactic ambiguity resolution as revealed by electrical brain responses. Biological Psychology. 1998;47(3):193–221. doi: 10.1016/s0301-0511(97)00033-1. [DOI] [PubMed] [Google Scholar]
- Friederici A.D., Wang Y., Herrmann C.S., Maess B., Oertel U. Localization of early syntactic processes in frontal and temporal cortical areas: A magnetoencephalographic study. Human Brain Mapping. 2000;11(1):1–11. doi: 10.1002/1097-0193(200009)11:1<1::AID-HBM10>3.0.CO;2-B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedrich M., Friederici A.D. Lexical priming and semantic integration reflected in the event-related potential of 14-month-olds. NeuroReport. 2005;16(6):653–656. doi: 10.1097/00001756-200504250-00028. [DOI] [PubMed] [Google Scholar]
- Friedrich M., Friederici A.D. Phonotactic knowledge and lexical-semantic processing in one-year-olds: Brain responses to words and nonsense words in picture contexts. Journal of Cognitive Neuroscience. 2005;17(11):1785–1802. doi: 10.1162/089892905774589172. [DOI] [PubMed] [Google Scholar]
- Friedrich M., Friederici A.D. Semantic sentence processing reflected in the event-related potentials of one- and two-year-old children. NeuroReport. 2005;16(16):1801–1804. doi: 10.1097/01.wnr.0000185013.98821.62. [DOI] [PubMed] [Google Scholar]
- Friedrich M., Friederici A.D. The origins of word learning: Brain responses of 3-month-olds indicate their rapid association of objects and words. Developmental Science. 2017;20(2) doi: 10.1111/desc.12357. [DOI] [PubMed] [Google Scholar]
- Friedrich M., Wilhelm I., Born J., Friederici A.D. Generalization of word meanings during infant sleep. Nature Communications. 2015;6:6004. doi: 10.1038/ncomms7004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frisch S., Schlesewsky M. The N400 reflects problems of thematic hierarchizing. NeuroReport. 2001;12(15):3391–3394. doi: 10.1097/00001756-200110290-00048. [DOI] [PubMed] [Google Scholar]
- Frisch S., Schlesewsky M., Saddy D., Alpermann A. The P600 as an indicator of syntactic ambiguity. Cognition. 2002;85(3):B83–B92. doi: 10.1016/s0010-0277(02)00126-9. [DOI] [PubMed] [Google Scholar]
- Gorrell P. The subject-before-object preference in german clauses. In: Hemforth K.L.B., editor. German sentence processing. Springer; Dordrecht: 2000. [Google Scholar]
- Grewe T., Bornkessel-Schlesewsky I., Zysset S., Wiese R., von Cramon D.Y., Schlesewsky M. The role of the posterior superior temporal sulcus in the processing of unmarked transitivity. Neuroimage. 2007;35(1):343–352. doi: 10.1016/j.neuroimage.2006.11.045. [DOI] [PubMed] [Google Scholar]
- Grimm H., Aktas M., Frevert S. Hogrefe; Göttingen: 2000. SETK-2: Sprachentwicklungstest für zweijährige Kinder. [Google Scholar]
- Gunter T.C., Stowe L.A., Mulder G. When syntax meets semantics. Psychophysiology. 1997;34(6):660–676. doi: 10.1111/j.1469-8986.1997.tb02142.x. [DOI] [PubMed] [Google Scholar]
- Hahne A., Eckstein K., Friederici A.D. Brain signatures of syntactic and semantic processes during children's language development. Journal of Cognitive Neuroscience. 2004;16(7):1302–1318. doi: 10.1162/0898929041920504. [DOI] [PubMed] [Google Scholar]
- Hahne A., Friederici A.D. Processing a second language: Late learners' comprehension mechanisms as revealed by event-related brain potentials. Bilingualism: Language and Cognition. 2001;4(2):123–141. [Google Scholar]
- Hoeks J.C.J., Stowe L.A., Doedens G. Seeing words in context: The interaction of lexical and sentence level information during reading. Cognitive Brain Research. 2004;19(1):59–73. doi: 10.1016/j.cogbrainres.2003.10.022. [DOI] [PubMed] [Google Scholar]
- Höhle B., Hörnig R., Weskott T., Knauf S., Krüger A. Effects of focus and definiteness on children's word order: Evidence from German five-year-olds' reproductions of double object constructions. Journal of Child Language. 2014;41(4):780–810. doi: 10.1017/S0305000913000196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb P.J., Coffey S.A., Neville H.J. Visual and auditory sentence processing – A developmental analysis using event-related brain potentials. Developmental Neuropsychology. 1992;8(2–3):203–241. [Google Scholar]
- Kaan E., Harris A., Gibson E., Holcomb P. The P600 as an index of syntactic integration difficulty. Language and Cognitive Processes. 2000;15(2):159–201. [Google Scholar]
- Kim A., Osterhout L. The independence of combinatory semantic processing: Evidence from event-related potentials. Journal of Memory and Language. 2005;52(2):205–225. [Google Scholar]
- Knoll L.J., Obleser J., Schipke C.S., Friederici A.D., Brauer J. Left prefrontal cortex activation during sentence comprehension covaries with grammatical knowledge in children. Neuroimage. 2012;62(1):207–216. doi: 10.1016/j.neuroimage.2012.05.014. [DOI] [PubMed] [Google Scholar]
- Kuperberg G.R., Kreher D.A., Sitnikova T., Caplan D.N., Holcomb P.J. The role of animacy and thematic relationships in processing active English sentences: Evidence from event-related potentials. Brain and Language. 2007;100(3):223–237. doi: 10.1016/j.bandl.2005.12.006. [DOI] [PubMed] [Google Scholar]
- Lopez-Calderon J., Luck S.J. ERPLAB: an open-source toolbox for the analysis of event related potentials. Frontiers in Human Neuroscience. 2014;8:213. doi: 10.3389/fnhum.2014.00213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacWhinney B. Erlbaum; Hillsdale, N.J: 2000. The CHILDES project: Tools for analyzing talk. [Google Scholar]
- MacWhinney B., Bates E., Kliegl R. Cue validity and sentence interpretation in English, German, and Italian. Journal of Verbal Learning and Verbal Behavior. 1984;23(2):127–150. [Google Scholar]
- Maess B., Schröger E., Widmann A. High-pass filters and baseline correction in M/EEG analysis. Commentary on: “How inappropriate high-pass filters can produce artefacts and incorrect conclusions in ERP studies of language and cognition”. Journal of Neuroscience Methods. 2016;266:164–165. doi: 10.1016/j.jneumeth.2015.12.003. [DOI] [PubMed] [Google Scholar]
- Matzke M., Mai H., Nager W., Russeler J., Munte T. The costs of freedom: An ERP – Study of non-canonical sentences. Clinical Neurophysiology. 2002;113(6):844–852. doi: 10.1016/s1388-2457(02)00059-7. [DOI] [PubMed] [Google Scholar]
- Mecklinger A., Schriefers H., Steinhauer K., Friederici A.D. Processing relative clauses varying on syntactic and semantic dimensions: An analysis with event-related potentials. Memory & Cognition. 1995;23(4):477–494. doi: 10.3758/bf03197249. [DOI] [PubMed] [Google Scholar]
- Meyer L., Obleser J., Kiebel S.J., Friederici A.D. Spatiotemporal dynamics of argument retrieval and reordering: An FMRI and EEG study on sentence processing. Frontiers in Psychology. 2012;3 doi: 10.3389/fpsyg.2012.00523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberecker R., Friederici A.D. Syntactic event-related potential components in 24-month-olds' sentence comprehension. NeuroReport. 2006;17(10):1017–1021. doi: 10.1097/01.wnr.0000223397.12694.9a. [DOI] [PubMed] [Google Scholar]
- Oberecker R., Friedrich M., Friederici A.D. Neural correlates of syntactic processing in two-year-olds. Journal of Cognitive Neuroscience. 2005;17(10):1667–1678. doi: 10.1162/089892905774597236. [DOI] [PubMed] [Google Scholar]
- Ponton C.W., Eggermont J.J., Don M., Waring M.D., Kwong B., Cunningham J., Trautwein P. Maturation of the mismatch negativity: effects of profound deafness and cochlear implant use. Audiology and Neurootology. 2000;5(3–4):167–185. doi: 10.1159/000013878. http://doi.org/13878 [DOI] [PubMed] [Google Scholar]
- Rösler F., Pechmann T., Streb J., Röder B., Hennighausen E. Parsing of sentences in a language with varying word order: Word-by-word variations of processing demands are revealed by event-related brain potentials. Journal of Memory and Language. 1998;38(2):150–176. [Google Scholar]
- Sakai K.L. Language acquisition and brain development. Science. 2005;310(5749):815–819. doi: 10.1126/science.1113530. [DOI] [PubMed] [Google Scholar]
- Schaner-Wolles C. Strategies in acquiring grammatical relations in German: Word order or case marking. Folia Linguistica. 1989;23(1–2):131–156. [Google Scholar]
- Schipke C.S., Friederici A.D., Oberecker R. Brain responses to case-marking violations in German preschool children. NeuroReport. 2011;22(16):850–854. doi: 10.1097/WNR.0b013e32834c1578. [DOI] [PubMed] [Google Scholar]
- Schipke C.S., Knoll L.J., Friederici A.D., Oberecker R. Preschool children's interpretation of object-initial sentences: Neural correlates of their behavioral performance. Developmental Science. 2012;15(6):762–774. doi: 10.1111/j.1467-7687.2012.01167.x. [DOI] [PubMed] [Google Scholar]
- Schlesewsky M., Bornkessel-Schlesewsky I. Birgit Rausing language program conference seres in linguistics; Lund, Sweden: 2009. When semantic P600s turn into N400s: On cross-linguistic differences in online verb-argument linking; pp. 75–97. [Google Scholar]
- Schlesewsky M., Bornkessel I. On incremental interpretation: Degrees of meaning accessed during sentence comprehension. Lingua. 2004;114(9–10):1213–1234. [Google Scholar]
- Schlesewsky M., Bornkessel I., Frisch S. The neurophysiological basis of word order variations in German. Brain and Language. 2003;86(1):116–128. doi: 10.1016/s0093-934x(02)00540-0. [DOI] [PubMed] [Google Scholar]
- Siegmüller J., Kauschke C., van Minnen S., Bittner D. Urban & Fischer Verlag; München: 2010. Test zum Satzverstehen von Kindern (TSVK) [Google Scholar]
- Silva-Pereyra J., Rivera-Gaxiola M., Kuhl P.K. An event-related brain potential study of sentence comprehension in preschoolers: Semantic and morphosyntactic processing. Brain Research Cognitive Brain Research. 2005;23(2–3):247–258. doi: 10.1016/j.cogbrainres.2004.10.015. [DOI] [PubMed] [Google Scholar]
- Skeide M.A., Brauer J., Friederici A.D. Syntax gradually segregates from semantics in the developing brain. Neuroimage. 2014;100:106–111. doi: 10.1016/j.neuroimage.2014.05.080. [DOI] [PubMed] [Google Scholar]
- Skeide M.A., Friederici A.D. The ontogeny of the cortical language network. Nature Reviews Neuroscience. 2016;17(5):323–332. doi: 10.1038/nrn.2016.23. [DOI] [PubMed] [Google Scholar]
- Szagun G., Stumper B., Schramm S.A. Pearson; Frankfurt (a.M.): 2009. FRAKIS: Fragebogen zur frühkindlichen Sprachentwicklung (FRAKIS) und FRAKIS-K (Kurzform) [Google Scholar]
- Vissiennon K., Friederici A.D., Brauer J., Wu C.Y. Functional organization of the language network in three- and six-year-old children. Neuropsychologia. 2017;98:24–33. doi: 10.1016/j.neuropsychologia.2016.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weckerly J., Kutas M. An electrophysiological analysis of animacy effects in the processing of object relative sentences. Psychophysiology. 1999;36(5):559–570. [PubMed] [Google Scholar]
- Wolff S., Schlesewsky M., Hirotani M., Bornkessel-Schlesewsky I. The neural mechanisms of word order processing revisited: Electrophysiological evidence from Japanese. Brain and Language. 2008;107(2):133–157. doi: 10.1016/j.bandl.2008.06.003. [DOI] [PubMed] [Google Scholar]
- Wu C.Y., Vissiennon K., Friederici A.D., Brauer J. Preschoolers' brains rely on semantic cues prior to the mastery of syntax during sentence comprehension. Neuroimage. 2016;126:256–266. doi: 10.1016/j.neuroimage.2015.10.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.