

Abstract
Cardiac hypertrophy is one of the key structural changes that occurs in diabetic cardiomyopathy. Previous studies have indicated that the activation of NF-κB by calpain-1, a Ca2+-dependent cysteine protease, serves an important role in cardiac hypertrophy. The aim of the present study was to assess the effect of 30 and 60 mg/kg Lycium barbarum polysaccharide (LBP) treatment, the major active ingredient extracted from Lycium barbarum, on cardiac hypertrophy in streptozotocin (STZ) induced diabetic rats. In addition, the present study examined the possible underlying mechanisms of this effect by assessing calpain-1 expression and the NF-κB pathway. The mRNA expression of atrial natriuretic peptide (ANP) and brain natriuretic peptide (BNP) was determined by reverse transcription-quantitative PCR. Western blotting was used to detect the protein expressions of calpain-1, interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), intercellular adhesion molecule-1 (ICAM-1), vascular adhesion molecule-1 (VCAM-1) and toll-like receptor-4 (TLR-4) in the heart tissue. The results revealed that compared with non-diabetic rats, diabetic rats exhibited cardiac hypertrophy. Cardiac hypertrophy was defined by the following: Dysfunction of the cardiac hemodynamics, an increase in the ratios of left ventricular weight/body weight and heart weight/body weight and the increased expressions of ANP and BNP, which serve as hypertrophic markers in cardiac tissue. However, all of these changes were attenuated in diabetic rats treated with LBP. In addition, the protein expression of calpain-1 was increased in the heart tissue of diabetic rats compared with that of non-diabetic rats, where it was inhibited by LBP. LBP also decreased the protein expression of certain inflammatory mediators, IL-6, TNF-α, ICAM-1, VCAM-1 and TLR-4 in diabetic heart tissue. Furthermore, LBP treatment reduced the production of reactive oxygen species, upregulated the protein expression of endothelial nitric oxide synthase and downregulated the protein expression of inducible nitric-oxide synthase. Additionally, LBP increased the protein expression of p65, the subunit of NF-κB and inhibitory protein кB-α in the cytoplasm and reduced p65 expression in the nucleus. In conclusion, LBP improves cardiac hypertrophy, inhibits the expression of calpain-1 and inhibits the activation of NF-κB in diabetic rats.
Keywords: lycium barbarum polysaccharide, diabetic rat, cardiac hypertrophy, nuclear factor-κB, calpain-1
Introduction
Diabetic cardiomyopathy (DCM) is one of the most common complications of diabetes mellitus and is closely associated with the increasing incidence of heart failure (1). Although the duration and severity of hyperglycaemia are considered major risk factors for the development of DCM (2), the molecular mechanisms underlying DCM remain unknown. Cardiac hypertrophy is one of the key structural changes in DCM and is often preceded by the pathological phenotype of DCM (3). Calpain-1, a Ca2+-dependent cysteine protease, is implicated in a number of pathological conditions associated with cardiovascular diseases including cardiac hypertrophy (4–7). It has been previously reported that calpain-1 is upregulated in the heart tissue of rats treated with isoproterenol (8), highlighting the important role of calpain-1 in the development of sympathetic hypertrophic cardiomyopathy. The involvement of calpain-1 in hypertrophic cardiomyopathy largely depends on its mediation of oxidative stress and inflammation, which are the most important contributors to the onset and progression of diabetic cardiac hypertrophy (8,9). In addition, a previous study has revealed that the inhibition of the reactive oxygen species (ROS)-/NF-κB pathway protects cardiomyocytes from hypertrophy (10). There has been increasing evidence that inflammation and adhesion molecules are involved in diabetes and the progression of cardiac hypertrophy in diabetes (11,12). NF-κB serves an important role in the regulation of pro-inflammatory genes leading to the overproduction of inflammatory mediators, including tumor necrosis factor (TNF)-α, interleukin-6 (IL-6) and inducible nitric oxide synthase (iNOS) in the hearts of isoproterenol treated rats (13). It has been reported that the activity of calpain-1 increases in high glucose-treated cardiomyocytes and streptozotocin (STZ)-induced diabetes (2,8). In addition, calpain-1 has been reported to be involved in acute inflammatory processes via the activation of the NF-κB pathway (7,14). In conclusion, these results indicate that the improvement of cardiac hypertrophy may be achieved by inhibiting the expression of calpain-1 expression and the activation of NF-κB.
Lycium barbarum, a member of the Solanaceae family, produces red-colored fruits, commercially known as Goji berries and has been used as a traditional Chinese herbal medicine for thousands of years (15). Lycium barbarum polysaccharide (LBP) is the major active ingredient extracted from Lycium barbarum and has a number of important bioactivities, including anti-oxidation (16), immunomodulation (17) and anti-inflammation (18). In STZ-induced diabetic animals, LBP has been indicated to attenuate testicular dysfunction (19), protect peripheral neuropathy (20), improve male sexual dysfunction and fertility impairments in males (21), enhance spermatogenesis (22) and inhibit diabetic nephropathy (23). However, to the best of our knowledge, the protective effect of LBP on cardiac hypertrophy in diabetic rats has not yet been reported. Further investigation is required on whether this potential protective effect is targeted on calpain-1 expression and NF-κB pathway.
The current study hypothesized that LBP may protect diabetic rats from cardiac hypertrophy with the following taken into consideration: Calpain-1 mediates activation of the NF-κB pathway, which leads to oxidative stress and inflammation, serving an essential role in the development of cardiac hypertrophy, and LBP possesses antioxidative and anti-inflammatory effects (7,9). The present study also assessed the underlying mechanism of this protected effect by targeting calpain-1 expression and the NF-κB pathway.
Materials and methods
Chemicals and reagents
LBP with >98% purity was obtained from Ningxia Qiyuan Pharmaceutical Co., Ltd. The immunohistochemical kit was purchased from OriGene Technologies, Inc. Antibodies against NF-κB subunit (p65), inhibitory protein кB (IкB)-α, laminB and GAPDH were obtained from Abcam. Antibodies against iNOS, TNF-α, intercellular adhesion molecule (ICAM)-1, vascular adhesion molecule (VCAM)-1, Toll-Like Receptor 4 (TLR-4) and horseradish peroxidase goat anti-rabbit IgG (H+L) were obtained from ABclonal Biotechnology Co., Ltd. Endogenous nitric oxide synthase (eNOS), IL-6 and calpain-1 were obtained from Cell Signaling Technology Inc. STZ was obtained from Sigma-Aldrich (Merck KGaA).
STZ-induced diabetic model in rats
A total of 60 adult male Sprague-Dawley rats (180–200 g) were obtained from the Laboratory Animal Center of the Jinzhou Medical University, (Liaoning, China). The present study was approved by the Ethics Committee of Animal Experiments of the Jinzhou Medical University (approval number: LMU-2016-138; Liaoning, China). Animal procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (24). Rats were housed at a temperature of 20–22°C, a relative humidity of 50–60%, a 12 h light/dark cycle and with a free access to food and water. Rats were considered diabetic if they exhibited hyperglycemia (≥15 mmol/l) 72 h following the one-time intra peritoneal injection of STZ (50 mg/kg). Diabetic rats were divided into three groups: An STZ group (n=10), an STZ+LBP (60 mg/kg/d) group (n=10) and an STZ+LBP (30 mg/kg/d) group (n=10). The groups were administered saline solution [intragastric (i.g.)] and/or 60 and 30 mg/kg/d LBP (i.g.) for 12 weeks. An additional 10 healthy non-diabetic rats were used as the control group (n=10) and were administered the vehicle. Cardiac hypertrophy was defined by the following: Dysfunction of the cardiac hemodynamics, an increase in the ratios of left ventricular weight/body weight and heart weight/body weight and the increased expressions of atrial natriuretic peptide (ANP) and brain natriuretic peptide (BNP), which serve as hypertrophic markers in cardiac tissue.
Hemodynamics and heart weight index measurement
Hemodynamics was conducted after the rats were anesthetized with intraperitoneal injection of sodium pentobarbital (0.04 g/kg). Overdose of 20% urethane (1 g/kg) followed by exsanguination were used to sacrifice the rats following the hemodynamics. Hemodynamics and heart weight index was calculated according to previous reported methods (25). The BL-420S polygraph (Chengdu TME Technology Co. Ltd.) was used to record the left ventricular end-diastolic pressure (LVEDP), left ventricular systolic pressure (LVSP) and the maximal rate of left ventricular systolic and diastolic pressure (±dp/dtmax). The indexes of HW/BW and LVW/BW were defined as heart weight/body weight, and left ventricular weight/body weight, respectively.
Histological analysis
Heart tissues (thickness 5 µm) were fixed in 10% neutral formaldehyde buffer at 25°C for 24 h and embedded in paraffin. Tissues were subsequently stained with hematoxylin-eosin (HE; H staining for 5 min at 25°C, 1% E staining for 3 min at 25°C) to evaluate morphological changes.
mRNA expression with reverse transcription-quantitative PCR (RT-qPCR)
Heart tissue was homogenized and total mRNA was isolated using a TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). According to the manufacturer's protocol, total RNA (500 ng) was reverse-transcribed to cDNA using the HiScript® II One Step RT-PCR kit (Vazyme). The mRNA expression of atrial natriuretic peptide (ANP) and brain natriuretic peptide (BNP) was examined using a ChamQ SYBR qPCR Master Mix kit (Vazyme) with a BioRad iQ5 Real Time PCR system (Bio-Rad Laboratories, Inc.) according to the manufacturer's protocol. The sequences of the primers were as follows: ANP forward, 5′-CAGCACAATAGAGCCGCTGA-3′ and reverse, 5′-GGGCAGGAGCTTGAACACG-3′; BNP forward, 5′-GCAGAAGCTGCTGGAGCTGA-3′ and reverse, 5′-ATCCGGAAGGCGCTGTCTTG-3′; GAPDH forward, 5′-GAGACAGCCGCATCTTCTTG-3′ and reverse, 5′-ATACGGCCAAATCCGTTCAC-3′. The thermocycling conditions were as follows: Initial denaturation at 95°C for 30 sec; 40 cycles of annealing at 95°C for 10 sec and extension at 60°C for 30 sec. mRNA levels were normalized to the internal reference gene GAPDH according to the 2−ΔΔCq method (26).
Serum IL-6 and TNF-α determination
Serum contents of IL-6 and TNF-α were determined using IL-6 ELISA kit (cat. no. R6000B) and TNF-α ELISA kit (cat. no. RTA00) obtained from R&D Systems, Inc. according to the manufacturer's protocols.
Detection of ROS in heart tissues
The oxidation-sensitive fluorescent dye dihydroethidium (DHE) was used to assess ROS content in situ. Heart tissues were embedded in tissue freezing medium (O.C.T. Compound 4583; Sakura Finetek USA, Inc.) and cut in a cryostat (LeicaCM1850; Leica Microsystems GmbH). Transverse heart tissue sections (5 µm) were disposed on glass slides, in order to reach the equilibrium for 30–60 min at 37°C in PBS. The sections were then incubated with DHE at 37°C for 30 min in the dark and control sections received an identical volume of PBS. Fluorescent images were subsequently obtained with an optical microscope (magnification, ×200; LeicaDFC 300FX: Leica Microsystems GmbH). Total and background pixel intensity were measured and used to correct the DHE intensity of each image. Quantification of DHE intensity was performed using ImageJ software (version 1.4.3.67, National Institutes of Health).
Immunohistochemical analysis
Tissue samples were treated with endogenous peroxidase blocker (cat. no. PV-6001; OriGene Technologies, Inc.) at 25°C for 10 min, and incubated with anti-p65 antibodies (1:100, cat. no. ab16502; Abcam) overnight at 4°C. This was followed by an incubation with horseradish peroxidase-labeled goat anti-rabbit IgG polymer (cat. no. PV-6001; OriGene Technologies, Inc.) at 25°C for 20 min. Tan or brown pigmented cell nuclei were considered to indicate a positive expression. Images were captured using a fluorescence microscope (Leica DMI300B). The results were analyzed using the Image-Pro Plus 6.0 (Media Cybernetics, Inc.) image analysis system. In each section, five positive colored regions were randomly selected and the average intensity of nuclear staining was determined.
Western blot analysis
Heart tissues were homogenized in phenylmethane sulfonyl fluoride and RIPA lysis buffer (cat. no. P0013B, Beyotime Institute of Biotechnology) and centrifuged at 12,000 × g for 20 min at 4°C. Nuclear proteins were separated using Active Motif's Nuclear and Cytoplasm Extraction kit (Dakewe Biotech Co., Ltd.) according to the manufacturer's protocol. A total of 40 µg of protein/lane from all samples was loaded onto 10% SDS-PAGE, transferred to polyvinylidene fluoride membranes. The membranes were then blocked for 1.5 h at 25°C with 10% bovine serum albumin (cat. no. 10735108001; Roche Applied Science) and incubated with anti-calpain-1 (1:1,000, cat. no. 2556S), anti-p65 (1:500, cat. no. ab86299), IкB-α (1:2,000, cat. no. ab7217), GAPDH (1:500, cat. no. ab9483), IL-6 (1:1,000, cat. no. 3732S), TNF-α (1:1,500, cat. no. A0277), ICAM-1 (1:1,000, cat. no. A559), VCAM-1 (1:1,000, cat. no. A0279), TLR-4 (1:1,500, cat. no. A5258), eNOS (1:1,000, cat. no. 9572S), iNOS (1:1,000, cat. no. A0312) and Lamin B (1:500, cat. no. ab8982) overnight at 4°C. Following extensive washing (TBST), membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (1:1,000, cat. no. AS014; ABclonal Biotechnology Co., Ltd) for 2 h at 25°C and the immunoreactivity in protein bands was visualized via enhanced chemiluminescence (Gaomin ECL Chemiluminescence kit; Wanleibio Co., Ltd.). Western blotting results were quantified with Wright Cell Imaging Facility ImageJ Launcher software (version 1.4.3.67, National Institute of Health). Each experiment was performed at least in triplicate.
Statistical analysis
Results are presented as the mean ± standard error of the mean. Statistical analysis of data was performed using a one-way analysis of variance with Bonferroni's correction for multiple group comparisons. A Student's t-test was used for comparisons between two groups with SPSS 17.0 software (SPSS, Inc.). P<0.05 was considered to indicate a statistically significant difference.
Results
LBP attenuates cardiac hypertrophy in STZ-induced diabetic rats
In the control group, HE staining of the heart tissue revealed that the morphology of cells was normal and that myocardial fibers were arranged in neat rows (Fig. 1A). However, STZ-induced diabetic rats demonstrated wide myocardial fibers and disarranged myocytes (Fig. 1A). The subsequent administration of LBP (30 and 60 mg/kg/d) improved the observed pathological changes, indicating that LBP prevents effects of STZ in heart tissue. The results of the current study also indicated that STZ increased the ratios of HW/BW (Fig. 1B) and LVW/BW (Fig. 1C), and upregulated mRNA and protein expression of various hypertrophic markers, including ANP and BNP (Fig. 1D-G) when compared with the control. However, LBP treatment reduced the ratios of HW/BW and LVW/BW, and downregulated the mRNA and protein expression of ANP and BNP compared with STZ alone. The aforementioned results indicate that LBP treatment attenuates cardiac hypertrophy induced by STZ.
Figure 1.

LBP attenuates cardiac hypertrophy in STZ-induced diabetic rats. (A) Histological changes of heart tissue (scale bar, 40 µm). (B) HW/BW ratios. (C) LVW/BW ratio. (D) mRNA expression of ANP. (E) mRNA expression of BNP. (F) Protein expression of ANP. (G) Protein expression of BNP. Data are expressed as the mean ± SEM (A, n=4, B and C, n=10, D-G, n=4). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin; HW/BW, heart weight/body weight; LVW/BW, left ventricle weight/body weight; ANP, atrial natriuretic peptide; BNP, brain natriuretic peptide.
LBP lowers blood glucose levels of STZ-induced diabetic rats
Compared with the control group, it was revealed that there was nearly a two-fold increase in fasting blood glucose levels following STZ injection (Fig. 2A). However, blood glucose levels were significantly decreased in the STZ+LBP (60) group, but not in the STZ+LBP (30) group when compared with the STZ group.
Figure 2.

LBP lowers blood glucose and improves the cardiac hemodynamics of STZ-induced diabetic rats. (A) Fasting blood glucose levels. (B) LVSP and (C) LVEDP levels following different treatments. (D) The +dp/dtmax and (E) -dp/dtmax of rats. Data are expressed as the mean ± SEM (n=10). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin; LVSP, left ventricular systolic pressure; LVEDP, left ventricular end-diastolic pressure; dp/dtmax, maximal rate of left ventricular systolic pressure.
LBP improves cardiac hemodynamics in STZ-induced diabetic rats
STZ treatment was revealed to increase LVSP (Fig. 2B) and LVEDP (Fig. 2C), and decrease +dp/dtmax (Fig. 2D) and-dp/dtmax (Fig. 2E) compared with the control group. However, LBP treatment was demonstrated to decrease LVSP and LVEDP in a dose-dependent manner. Furthermore, LBP treatment and increased ±dp/dtmax compared with STZ group.
LBP reduces superoxide anion generation and regulates the protein expression of eNOS and iNOS in STZ-induced diabetic rats
DHE staining indicated a significant increase in superoxide anion generation in heart tissue following STZ treatment when compared with the control. However, LBP treatment was revealed to reduce superoxide anion generation in a dose-dependent manner compared with the STZ group (Fig. 3A and B). STZ treatment was also revealed to downregulate the protein expression of eNOS and upregulate the expression of iNOS compared with the control group. Furthermore, the effect of STZ was reversed when introducing LBP treatment (Fig. 3C and D).
Figure 3.

LBP attenuated superoxide anion generation and the protein expression of eNOS and iNOS in STZ-induced diabetic rats. (A) DHE staining (scale bar, 40 µm) and (B) their respective fold changes in STZ-induced rats receiving different treatments. (C) Protein expression of eNOS and (D) iNOS. Data are expressed as the mean ± SEM (n=4). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin; DHE, dihydroethidium; eNOS, endogenous nitric oxide synthase; iNOS, inducible nitric oxide synthase.
LBP decreases inflammatory cytokine production in STZ-induced diabetic rats
Compared with the control group, STZ increased IL-6 (Fig. 4A) and TNF-α (Fig. 4B) content in rat serum, and upregulated the protein expression of IL-6 (Fig. 4C), TNF-α (Fig. 4D), ICAM-1 (Fig. 4E), VCAM-1 (Fig. 4F) and TLR-4 (Fig. 4G) in rat heart tissue. However, subsequent LBP treatment decreased serum IL-6 and TNF-α in a dose-dependent manner and downregulated the protein expression of IL-6, TNF-α, ICAM-1, VCAM-1 and TLR-4 compared with the STZ group. These results indicate that the inhibition of cardiac hypertrophy by LPB may be potentially attributed to its anti-inflammatory properties.
Figure 4.

LBP downregulates inflammatory cytokine production in STZ-induced diabetic rats. Serum (A) IL-6 and (B) TNF-α content in STZ-induced rats. The expression of (C) IL-6, (D) TNF-α, (E) ICAM-1, (F) VCAM-1 and (G) TLR-4 as determined via western blotting. Data are expressed as the mean ± SEM (A and B, n=10; C-G, n=4). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin; IL-6, interleukin-6; TNF-α, tumor necrosis factor-α; ICAM-1, intercellular adhesion molecule-1; VCAM-1, vascular adhesion molecule-1; TLR-4, Toll like receptor-4.
LBP downregulates the protein expression of calpain-1 in STZ-induced diabetic rats
STZ increased the protein expression of calpain-1 in rat heart tissue compared with the control group (Fig. 5). However, subsequent LBP treatment dose-dependently downregulated the protein expression of calpain-1 when compared with the STZ group.
Figure 5.
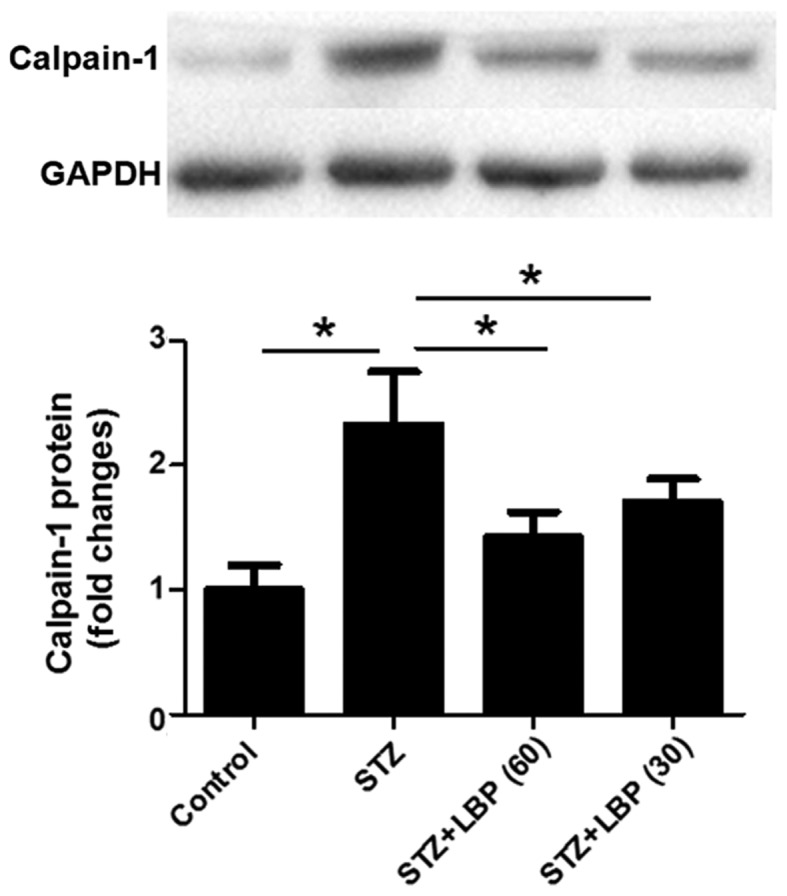
LBP reduces the protein expression of calpain-1 in STZ-induced diabetic rats. Data are expressed as the mean ± SEM (n=4). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin.
LBP inhibits the nuclear translocation of NF-κB in STZ-induced diabetic rats
Immunohistological staining of the p65 antibody indicated that STZ increased the number of p65 nuclear inputs compared with the control, which was decreased following LBP treatment (Fig. 6A and B). Western blot analysis revealed that compared with the control, STZ increased the protein expression of p65 in the nucleus (Fig. 6C), and decreased the expression of p65 (Fig. 6D) and IкB-α (Fig. 6E) in the cytoplasm of heart tissues. However, LBP treatment reversed the effect of STZ in a dose-dependent manner.
Figure 6.

LBP inhibits the nuclear translocation of NF-κB in the heart tissue of STZ-induced diabetic rats. (A) Immunohistological staining images (scale bar, 40 µm) and (B) their respective fold changes. (C) The expression of nuclear p65, (D) cytoplasmic p65 and (E) cytoplasmic IкB-α as determined via western blotting. Data are expressed as the mean ± SEM (n=4). *P<0.05. LBP, lycium barbarum polysaccharide; STZ, streptozotocin; IкB-α, inhibitory protein кB.
Discussion
In the present study, it was revealed that STZ-induced diabetic rats exhibited cardiac hypertrophy represented by upregulation of mRNA and protein expression levels of ANP and BNP, two hypertrophic markers (27), accompanied by an increased protein expression of calpain-1 and NF-κB activation in hypertrophic heart tissue. The results indicated that an increased protein expression of calpain-1 and the activation of NF-κB may serve key roles in the development of cardiac hypertrophy in diabetic rats. It was also demonstrated that the intragastrical administration of LBP attenuated cardiac hypertrophy, as it decreased the expression of calpain-1 and inhibited the activation of the NF-κB pathway. These results indicate that the improvement of cardiac hypertrophy by LBP treatment may be associated with the inhibition of calpain-1 expression and NF-κB pathway activation.
Lycium barbarum has been used as a traditional Chinese herbal medicine for thousands of years (15). LBP is the major active ingredient extracted from Lycium barbarum. In STZ-induced diabetic animals, LBP has been indicated to attenuate testicular dysfunction (19), protect peripheral neuropathy (20), improve male sexual dysfunction and fertility impairments (21), enhance spermatogenesis (22) and inhibit diabetic nephropathy (23). The results of the present study indicated that LBP improves cardiac hypertrophy in STZ-induced diabetic rats. The results of the aforementioned studies and present study indicate that LBP may effectively attenuate diabetic complications (19,22,23). These results also provide experimental evidence for the effective use of LBP in patients with diabetes. In terms of practical applications, Cai et al (28) reported that LBP may serve as a potential treatment aided-agent for patients with diabetes.
Increasing evidence has indicated that inflammation and oxidative stress serve key roles in the pathogenesis of cardiac hypertrophy (2,25,29). The present study also demonstrated that LBP inhibited the expression of various inflammatory molecules including IL-6, TNF-α, ICAM-1, VCAM-1 and TLR-4 in the serum and/or heart tissue of STZ-induced rats. In addition, LBP reduced the production of ROS and regulated the expression of iNOS and eNOS in diabetic heart tissue. These results indicate that the protective effect of LBP on cardiac hypertrophy in diabetes may be partly associated with the inhibition of inflammation and oxidative stress. Similarly, Du et al (18) reported that LBP reduces oxidative stress and inflammation, subsequently leading to anti-diabetic and anti-nephritis effects in a high-fat diet (12% protein, 5% fat, 67% carbohydrate, 5% cholesterol, and 5% other additives) and STZ-induced diabetic rats. In conclusion, these results indicated that anti-oxidation and anti-inflammation may serve as key underlying mechanisms for the improvement of diabetic complications by LBP.
Guo et al (29) reported that NF-κB activation is an important underlying mechanism of diabetic cardiac hypertrophy in STZ-induced type 1 diabetic mice and high glucose-treated H9c2 cardiomyocytes. The aforementioned study supports those of the present study as the protein expression of calpain-1 was increased and NF-κB was activated in the hypertrophic heart tissue of diabetic rats. In addition, it has been reported that calpain-1 accumulates in the mitochondria and promotes diabetic cardiomyopathy (30). It has been previously demonstrated that calpain-1 is upregulated in the hypertrophic heart tissue of rats treated with isoproterenol (8). In conclusion, the results of the current study revealed that calpain-1 and NF-κB serve important roles in the pathogenesis of cardiac hypertrophy. In accordance with the present results indicating that LBP inhibits the activation of NF-κB, Du et al (18) also reported that LBP mediates anti-diabetic and anti-nephritic effects in a high-fat diet and STZ-induced diabetic rats by regulating the NF-κB pathway.
The main limitation of the present study is that the cause-effect relations among parameters including calpain-1 expression, NF-κB activation, inflammatory molecules and oxidation indicators cannot be precisely determined. It is possible in the future to overcome this limitation by conducting animal experiments and/or by in vitro studies, using inhibitors of calpain-1 and NF-κB.
In conclusion, the present study demonstrated that LBP treatment inhibited cardiac hypertrophy in STZ-induced diabetic rats, an effect that may be associated with the inhibition of calpain-1 expression and NF-κB activation. These results provide an insight into the underlying mechanism of LBP and an experimental basis for the potential therapeutic use of LBP in diabetic cardiac hypertrophy.
Acknowledgements
Not applicable.
Funding
The current study was supported by the National Natural Science Foundation of China (grant no. 81870329), the Talent Fund of Liaoning Medical University (grant no. 2014-18) and the Natural Science Foundation of Liaoning Province (grant no. 2018020325).
Availability of data and materials
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' contributions
QL, QH and ML performed the experiments, analyzed the data and prepared the manuscript. FT and HW wrote and revised the manuscript, and designed the experiments.
Ethical approval and consent to participate
The experimental protocols were approved by the Committee on the Ethics of Animal Experiments of Jinzhou Medical University, China (Approval number: LMU-2016-138).
Patient consent for publication
Not applicable.
Conflict of interest
The authors declare that there is no conflict of interest associated with this work.
References
- 1.Zhang J, Qiu H, Huang J, Ding S, Huang B, Wu Q, Jiang Q. Naringenin exhibits the protective effect on cardiac hypertrophy via EETs-PPARs activation in streptozocin-induced diabetic mice. Biochem Biophys Res Commun. 2018;502:55–61. doi: 10.1016/j.bbrc.2018.05.119. [DOI] [PubMed] [Google Scholar]
- 2.Li Y, Ma J, Zhu H, Singh M, Hill D, Greer PA, Arnold JM, Abel ED, Peng T. Targeted inhibition of calpain reduces myocardial hypertrophy and fibrosis in mouse models of type 1 diabetes. Diabetes. 2011;60:2985–2994. doi: 10.2337/db10-1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Negishi K. Echocardiographic feature of diabetic cardiomyopathy: Where are we now? Cardiovasc Diagn Ther. 2018;8:47–56. doi: 10.21037/cdt.2018.01.03. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yan M, Chen K, He L, Li S, Huang D, Li J. Uric acid induces cardiomyocyte apoptosis via activation of calpain-1 and endoplasmic reticulum stress. Cell Physiol Biochem. 2018;45:2122–2135. doi: 10.1159/000488048. [DOI] [PubMed] [Google Scholar]
- 5.Hua Y, Nair S. Proteases in cardiometabolic diseases: Pathophysiology, molecular mechanisms and clinical applications. Biochim Biophys Acta. 2015;1852:195–208. doi: 10.1016/j.bbadis.2014.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen B, Zhao Q, Ni R, Tang F, Shan L, Cepinskas I, Cepinskas G, Wang W, Schiller PW, Peng T. Inhibition of calpain reduces oxidative stress and attenuates endothelial dysfunction in diabetes. Cardiovasc Diabetol. 2014;13:88. doi: 10.1186/1475-2840-13-88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yu L, Yin M, Yang X, Lu M, Tang F, Wang H. Calpain inhibitor I attenuates atherosclerosis and inflammation in atherosclerotic rats through eNOS/NO/NF-κB pathway. Can J Physiol Pharmacol. 2018;96:60–67. doi: 10.1139/cjpp-2016-0652. [DOI] [PubMed] [Google Scholar]
- 8.Mei M, Tang F, Lu M, He X, Wang H, Hou X, Hu J, Xu C, Han R. Astragaloside IV attenuates apoptosis of hypertrophic cardiomyocyte through inhibiting oxidative stress and calpain-1 activation. Environ Toxicol Pharmacol. 2015;40:764–773. doi: 10.1016/j.etap.2015.09.007. [DOI] [PubMed] [Google Scholar]
- 9.Xu T, Zhang B, Yang F, Cai C, Wang G, Han Q, Zou L. HSF1 and NF-κB p65 participate in the process of exercise preconditioning attenuating pressure overload-induced pathological cardiac hypertrophy. Biochem Biophys Res Commun. 2015;460:622–627. doi: 10.1016/j.bbrc.2015.03.079. [DOI] [PubMed] [Google Scholar]
- 10.Xu C, Tang F, Lu M, Yang J, Han R, Mei M, Hu J, Zhou M, Wang H. Astragaloside IV improves the isoproterenol-induced vascular dysfunction via attenuating eNOS uncoupling-mediated oxidative stress and inhibiting ROS-NF-κB pathways. Int Immunopharmacol. 2016;33:119–127. doi: 10.1016/j.intimp.2016.02.009. [DOI] [PubMed] [Google Scholar]
- 11.Zuo G, Ren X, Qian X, Ye P, Luo J, Gao X, Zhang J, Chen S. Inhibition of JNK and p38 MAPK-mediated inflammation and apoptosis by ivabradine improves cardiac function in streptozotocin-induced diabetic cardiomyopathy. J Cell Physiol. 2019;234:1925–1936. doi: 10.1002/jcp.27070. [DOI] [PubMed] [Google Scholar]
- 12.Wang Y, Huang X, Ma Z, Wang Y, Chen X, Gao Y. Ophiopogonin D alleviates cardiac hypertrophy in rat by upregulating CYP2J3 in vitro and suppressing inflammation in vivo. Biochem Biophys Res Commun. 2018;503:1011–1019. doi: 10.1016/j.bbrc.2018.06.110. [DOI] [PubMed] [Google Scholar]
- 13.Yun JM, Jialal I, Devaraj S. Epigenetic regulation of high glucose-induced proinflammatory cytokine production in monocytes by curcumin. J Nutr Biochem. 2011;22:450–458. doi: 10.1016/j.jnutbio.2010.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zou J, Li H, Chen X, Zeng S, Ye J, Zhou C, Liu M, Zhang L, Yu N, Gan X, et al. C/EBPβ knockdown protects cardiomyocytes from hypertrophy via inhibition of p65-NFκssssB. Mol Cell Endocrinol. 2014;390:18–25. doi: 10.1016/j.mce.2014.03.007. [DOI] [PubMed] [Google Scholar]
- 15.Tang HL, Chen C, Wang SK, Sun GJ. Biochemical analysis and hypoglycemic activity of a polysaccharide isolated from the fruit of Lycium barbarum L. Int J Biol Macromol. 2015;77:235–242. doi: 10.1016/j.ijbiomac.2015.03.026. [DOI] [PubMed] [Google Scholar]
- 16.Yang DM, Zhang JQ, Fei YF. Lycium barbarum polysaccharide attenuates chemotherapy-induced ovarian injury by reducing oxidative stress. J Obstet Gynaecol Res. 2017;43:1621–1628. doi: 10.1111/jog.13416. [DOI] [PubMed] [Google Scholar]
- 17.Chen S, Liang L, Wang Y, Diao J, Zhao C, Chen G, He Y, Luo C, Wu X, Zhang Y. Synergistic immunotherapeutic effects of Lycium barbarum polysaccharide and interferon-α2b on the murine Renca renal cell carcinoma cell line in vitro and in vivo. Mol Med Rep. 2015;12:6727–6737. doi: 10.3892/mmr.2015.4230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Du M, Hu X, Kou L, Zhang B, Zhang C. Lycium barbarum polysaccharide mediated the antidiabetic and antinephritic effects in diet-streptozotocin-induced diabetic sprague dawley rats via regulation of NF-κB. Biomed Res Int. 2016;2016:3140290. doi: 10.1155/2016/3140290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Shi GJ, Zheng J, Han XX, Jiang YP, Li ZM, Wu J, Chang Q, Niu Y, Sun T, Li YX, et al. Lycium barbarum polysaccharide attenuates diabetic testicular dysfunction via inhibition of the PI3K/Akt pathway-mediated abnormal autophagy in male mice. Cell Tissue Res. 2018;374:653–666. doi: 10.1007/s00441-018-2891-1. [DOI] [PubMed] [Google Scholar]
- 20.Liu SY, Chen L, Li XC, Hu QK, He LJ. Lycium barbarum polysaccharide protects diabetic peripheral neuropathy by enhancing autophagy via mTOR/p70S6K inhibition in Streptozotocin-induced diabetic rats. J Chem Neuroanat. 2018;89:37–42. doi: 10.1016/j.jchemneu.2017.12.011. [DOI] [PubMed] [Google Scholar]
- 21.Shi GJ, Zheng J, Wu J, Qiao HQ, Chang Q, Niu Y, Sun T, Li YX, Yu JQ. Protective effects of Lycium barbarum polysaccharide on male sexual dysfunction and fertility impairments by activating hypothalamic pituitary gonadal axis in streptozotocin-induced type-1 diabetic male mice. Endocr J. 2017;64:907–922. doi: 10.1507/endocrj.EJ16-0430. [DOI] [PubMed] [Google Scholar]
- 22.Shi GJ, Zheng J, Wu J, Qiao HQ, Chang Q, Niu Y, Sun T, Li YX, Yu JQ. Beneficial effects of Lycium barbarum polysaccharide on spermatogenesis by improving antioxidant activity and inhibiting apoptosis in streptozotocin-induced diabetic male mice. Food Funct. 2017;8:1215–1226. doi: 10.1039/C6FO01575A. [DOI] [PubMed] [Google Scholar]
- 23.Zhao R, Li QW, Li J, Zhang T. Protective effect of Lycium barbarum polysaccharide 4 on kidneys in streptozotocin-induced diabetic rats. Can J Physiol Pharmacol. 2009;87:711–719. doi: 10.1139/Y09-068. [DOI] [PubMed] [Google Scholar]
- 24.National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals, corp-author. National Academies Press (US); Washington, DC: 2011. Guide for the Care and Use of Laboratory Animals, 8th edition. [Google Scholar]
- 25.Tang F, Lu M, Yu L, Wang Q, Mei M, Xu C, Han R, Hu J, Wang H, Zhang Y. Inhibition of TNF-α-mediated NF-κB Activation by Ginsenoside Rg1 Contributes the Attenuation of Cardiac Hypertrophy Induced by Abdominal Aorta Coarctation. J Cardiovasc Pharmacol. 2016;68:257–264. doi: 10.1097/FJC.0000000000000410. [DOI] [PubMed] [Google Scholar]
- 26.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 27.Alvarez BV, Quon AL, Mullen J, Casey JR. Quantification of carbonic anhydrase gene expression in ventricle of hypertrophic and failing human heart. BMC Cardiovasc Disord. 2013;13:2. doi: 10.1186/1471-2261-13-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cai H, Liu F, Zuo P, Huang G, Song Z, Wang T, Lu H, Guo F, Han C, Sun G. Practical application of antidiabetic efficacy of Lycium barbarum polysaccharide in patients with type 2 diabetes. Med Chem. 2015;11:383–390. doi: 10.2174/1573406410666141110153858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Guo Y, Zhuang X, Huang Z, Zou J, Yang D, Hu X, Du Z, Wang L, Liao X. Klotho protects the heart from hyperglycemia-induced injury by inactivating ROS and NF-κB-mediated inflammation both in vitro and in vivo. Biochim Biophys Acta Mol Basis Dis. 2018;1864:238–251. doi: 10.1016/j.bbadis.2017.09.029. [DOI] [PubMed] [Google Scholar]
- 30.Ni R, Zheng D, Xiong S, Hill DJ, Sun T, Gardiner RB, Fan GC, Lu Y, Abel ED, Greer PA, Peng T. mitochondrial calpain-1 disrupts atp synthase and induces superoxide generation in type 1 diabetic hearts: A novel mechanism contributing to diabetic cardiomyopathy. Diabetes. 2016;65:255–268. doi: 10.2337/db15-0963. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.