

Abstract
5-HT activates the peristaltic reflex and is the neurotransmitter of a subset of myenteric interneurons. Hyperpolarizing afterpotential (AH)/type 2 neurons respond to 5-HT with a long-lived depolarization that is caused by the inhibition of a Ca2+-activated K+ conductance (gKCa). This effect is mediated by a G-protein-coupled receptor, 5-HT1P. 5-HT1P agonists specifically activate Gαo, the immunoreactivity of which was found to be highly abundant and membrane-associated in almost all enteric neurons. Responses of hyperpolarizing AH/type 2 neurons to 5-HT were inhibited by intracellular injection of GDPβS or anti-Gαo Fab fragments but were potentiated and prolonged by intracellular GTPγS. Responses to 5-HT were antagonized by pertussis toxin, downregulation of protein kinase C (PKC) and inhibitors of phosphatidylcholine phospholipase C (PC-PLC), PKC (including pseudosubstrate peptides, chelerythrine, and the α/β isoform-specific inhibitor Gö 6976), protein kinase A (PKA), and adenylate cyclase. Responses to 5-HT were mimicked by activators of PKC, and 5-HT induced a concentration-dependent increase in the membrane-associated PKC activity in isolated myenteric ganglia. Immunocytochemical studies suggested that the most abundant isoforms of PKC in enteric neurons are α and δ. These data suggest that signal transduction of the 5-HT1P-mediated slow response to 5-HT involves activation of PC-PLC by Gαo to liberate diacylglycerol, which stimulates PKC (most likely α). PKC probably activates adenylate cyclase, which through cAMP, activates PKA. Activation of both PKA and PKC lead to closure of gKCa.
Keywords: enteric nervous system, serotonin, 5-hydroxytryptamine, 5-HT1P receptor, protein kinase C, protein kinase A, phosphatidylcholine phospholipase C, Go, G-proteins, signal transduction
Responses of the gut to exogenous 5-HT may be nerve- or muscle-mediated, excitatory, or inhibitory (Mawe and Gershon, 1993; Gershon et al., 1994; Wade et al., 1994). The complexity of these responses is attributable to the multiplicity of 5-HT receptor subtypes in the enteric nervous system (ENS) (Gershon, 1995) and musculature (Engel et al., 1984). At least five subtypes are present on nerve, 5-HT1A (Galligan et al., 1988; Galligan and North, 1991), 5-HT1P (Branchek et al., 1984; Mawe et al., 1986; Mawe and Gershon, 1993), 5-HT2B (unpublished data), 5-HT3 (Mawe et al., 1986; Derkach et al., 1989), and 5-HT4 (Clarke et al., 1989; Craig and Clarke, 1990; Pan and Galligan, 1994) and two subtypes on muscle, 5-HT2A and 5-HT2B (Engel et al., 1984; Cohen et al., 1985; Foguet et al., 1992a,b; Kursar et al., 1992). Of these receptors, only 5-HT1P has been shown to play roles in specific physiological responses. These include the mediation of slow EPSPs in myenteric neurons (Takaki et al., 1985a,b; Mawe et al., 1989; Wade et al., 1991, 1994; Galligan, 1995) and the initiation of the peristaltic reflex (Kirchgessner et al., 1992, 1997; Wade et al., 1996).
The response of myenteric neurons that is 5-HT1P-mediated is known as the slow response (Wood, 1989; Gershon et al., 1994;Galligan, 1995). These responses are most common in hyperpolarizing afterpotential (AH)/type 2 neurons, a cell that is defined by a characteristic Ca2+-activated K+ conductance (gKca). This gKca gives rise to a pronounced hyperpolarizing afterpotential (AH) and also contributes to the resting membrane potential (Galligan, 1995). 5-HT inhibits gKCa, resulting in a slowly developing but long-lived depolarization, associated with an increase in input resistance and inhibition of the AH (Wood and Mayer, 1979; Grafe et al., 1980; Johnson et al., 1980; Morita et al., 1982; Hirst et al., 1985; North and Tokimasa, 1987; Galligan, 1995). Slow responses to 5-HT are identical to slow EPSPs evoked in the same neurons (Wood and Mayer, 1979), and both are blocked specifically byN-acetyl-5-hydroxytryptophyl-5-hydroxytryptophan amide (5-HTP-DP) (Takaki et al., 1985a; Mawe et al., 1986), renzapride (Mawe et al., 1989), and anti-idiotypic antibodies that recognize 5-HT receptors (Wade et al., 1994). The 5-HT1P receptor binds 5-HT with high affinity (KD ≈ 2–3 nm) (Branchek et al., 1984; Gershon et al., 1985; Mawe et al., 1986) and requires that potential ligands be hydroxylated indoles (Branchek et al., 1988; Mawe and Gershon, 1993).
5-HT receptor subtypes are defined by their primary structure, transductional properties, and operational characteristics (Humphrey et al., 1993). Although the transductional properties of the 5-HT1P receptor have not previously been fully characterized, following three observations have led to the suggestion that this receptor is a member of the heptahelical G-protein-coupled receptor family: (1) the binding of 3H-5-HT by the receptor is antagonized by GTPγS (Gershon et al., 1991; Fiorica-Howells et al., 1993); (2) intracellular injection of GTPγS potentiates and prolongs slow responses to 5-HT; and (3) similar injections of GDPβS inhibit slow responses (Wang et al., 1996). The G-protein to which the 5-HT1P receptor is coupled, moreover, is likely to be Go, because Go is activated specifically when enteric neuronal membranes are incubated with 5-HT or 5-hydroxyindalpine (Wang et al., 1996), a 5-HT1P agonist (Branchek et al., 1988). Activation of Go by either 5-HT or 5-hydroxyindalpine is antagonized by 5-HTP-DP (Wang et al., 1996). The current experiments were carried out to identify the signal transduction pathway responsible for 5-HT1P-mediated slow responses.
MATERIALS AND METHODS
Tissue preparation. Male guinea pigs weighing 250–350 gm were stunned and exsanguinated. A segment of ileum was excised 10–20 cm proximal to the ileocecal junction and placed in oxygenated (95% O2/5% CO2) Krebs’ solution of the following composition (in mm): NaCl 121.3, KCl 5.95, CaCl2 2.5, NaHCO3 14.3, NaH2PO4 1.34, MgCl2 1.2, and glucose 11.5. The Krebs’ solution contained nifedipine and scopolamine (1 μm each) to block longitudinal muscle contractions while intracellular recordings were obtained. A 1.5 cm segment of ileum was cut open along the mesenteric border and pinned out flat (mucosal surface up) in a Petri dish lined with a silicone elastomer. Preparations of longitudinal muscle with adherent myenteric plexus (LMMP) were dissected by removing the mucosa, submucosa, and circular muscle with a fine forceps and scissors under microscopic control. A 5 mm2 LMMP segment was transferred to a small recording chamber (volume, 0.5 ml) that was coated with a silicone elastomer. The LMMP was stretched lightly and pinned with small stainless steel pins to the silicone coating of the bottom of the chamber. Preparations were superfused (3.5 ml/min; 36°C) with Krebs’ solution oxygenated with a mixture of 95% O2/5% CO2.
Intracellular recording. Individual myenteric ganglia were visualized at a magnification of 6.3×. Intracellular recordings were obtained from neurons using glass microelectrodes filled with 2m KCl (tip resistance, 90–160 MΩ). An amplifier with an active bridge circuit (Axoclamp 2A, Axon Instruments, Foster City, CA) was used to record the transmembrane potential difference and to inject current via the recording electrode.
Drugs, chemicals, and antibodies. 5-HT (Sigma Chemical, St. Louis, MO), forskolin (Research Biochemicals International, Natick, MA), histamine (Research Biochemical International), and the PKC activators phorbol 12,13-dibutyrate (PDBu, Research Biochemical International), (−)-7-octylindolactam V (LC Laboratories, Woburn, MA), and 1-oleoyl-2-acetyl-rac-glycerol (OAG, Sigma) were applied to neurons by ejection with pressure from a micropipette (filled with a 1.0 mm solution) or by addition to the fluid superfusing the preparations. In six cells used to evaluate the reproducibility of responses, the ratio of the amplitude of a second response to 5-HT to that of its predecessor was 0.97 ± 0.05 (p= 0.5669). The interval between trials of 5-HT was at least 5 min to avoid desensitization of 5-HT receptors. In all experiments, the effect of drugs on responses to 5-HT, histamine, or forskolin was not investigated until after reproducible responses to the agonist were obtained. Membrane-permeable potential antagonists of steps in signal transduction were added to the superfusing solutions. These included the phosphatidylcholine-phospholipase C (PC-PLC) inhibitor D609 (Kamiya Biomedical, Thousand Oaks, CA); the adenylate cyclase inhibitor 2′,5′-dideoxyadenosine (DDA, Biomol Research Laboratory, Plymouth Meeting, PA); and the PKC inhibitors chelerythrine (LC Laboratories), staurosporine (Research Biochemical International), K252a (Kamiya), Gö 6976, an indolcabazole that selectively inhibits the PKC isozymes, α, β1, β2, and γ, but not δ, ε, or ζ (Martiny-Baron et al., 1993) (Calbiochem, La Jolla, CA); and a myristolated pseudosubstrate sequence from PKC α and β, myr-FARKGALRQ (Biomol). Compounds that do not readily cross cell membranes were microinjected into neurons via the recording microelectrode. These included GTPγS (Sigma), GDPβS (Sigma), Fab fragments of antibodies to Gαo (preparation described below), the PKC α, β pseudosubstrate, RFARKGALRQKNV (PKC(19–31) (Kemp et al., 1994) (LC Labs, 1.0 mm), and the protein kinase A (PKA) inhibitor Rp-adenosine 3′5′-cyclic monophosphothiolate triethylamine cAMPS (Rp-cAMPS, 70 mm, Research Biochemical International). Because pertussis toxin (PTx, Research Biochemical International) penetrates membranes slowly, preparations were incubated for 6 hr ± PTx before determining the response to 5-HT.
Antibodies to Gαo were obtained from Upstate Biotechnology, Lake Placid, NY. Fab fragments (Mage and Lamoyi, 1987) were prepared by digestion with papain (in an enzyme-to-protein ratio of 1:100) in 500 μl of 20 mm sodium phosphate buffer, pH 7.4, that contained 20 μg of antibody to Gαo, dithiothreitol (1.0 mm), and EDTA (2.0 mm). Digestion was allowed to proceed at 37°C for 4–5 hr. After incubation, the reaction mixture was cooled to room temperature and transferred to dialysis tubing and dialyzed against PBS overnight. Protein A-Sepharose 4B beads were then added to the dialyzed mixture to bind Fc fragments of the antibody. The beads were then removed by centrifugation to provide a final clear solution containing Fab fragments that could be loaded into a micropipette.
Immunocytochemistry. Laminar preparations of LMMP or the dissected submucosa (containing the submucosal plexus) were fixed for 3 hr with 4% formaldehyde (freshly prepared from paraformaldehyde) in 0.1 m sodium phosphate buffer, pH 7.4, at room temperature, and washed three times with PBS. To locate proteins in the tissue by immunocytochemistry, free-floating LMMP or submucosal preparations were exposed to PBS containing 1.0% Triton X-100 and 10% horse serum for 30 min to permeabilize the tissue and reduce background staining. Gαo immunoreactivity was demonstrated with rabbit polyclonal antibodies (Upstate Biotechnology) at a concentration of 1.0 μg/ml. Preparations were incubated overnight in a humidified chamber at room temperature. Bound antibody was visualized by incubating tissues for 2 hr with biotinylated affinity-purified goat anti-rabbit IgG secondary antibodies (diluted 1:400; Kirkegaard & Perry, Gaithersburg, MD) at room temperature and then for 2 hr with avidin-FITC (diluted 1:200; Vector Laboratories, Burlingame, CA). Calbindin immunoreactivity, which marks 70–80% of neurons classified anatomically as Dogiel type II and physiologically as AH/type 2 (Pompolo and Furness, 1988; Furness et al., 1990), was located simultaneously in the same sections used to visualize Gαö immunoreactivity by double-label immunocytochemistry. To demonstrate calbindin immunoreactivity, preparations were incubated overnight at room temperature with mouse monoclonal antibodies to calbindin (diluted 1:100; Sigma), and bound primary antibodies were located with affinity-purified goat anti-mouse secondary antibodies labeled with tetramethylrhodamine isothiocyanate (TRITC, diluted 1:100; Kirkegaard and Perry). Double-label immunocytochemistry was made possible, because primary antibodies were raised in different species and immunoreactivity was visualized with species-specific secondary antibodies. Specific antibodies to the following isozymes of PKC, α, β1, β2, γ, δ, ε, ζ, ι/λ, and η (Jiang et al., 1994), were donated by Dr. Todd Sacktor (State University of New York, Downstate Medical Center). These were all polyclonal rabbit antibodies and thus were visualized, as described above, with biotinylated goat anti-rabbit secondary antibodies, and avidin-FITC. Immunostained tissues were examined with a Leica DMRB Microscope equipped for vertical fluorescence microscopy. FITC fluorescence was detected using a Leica “L-4” filter cube (exciting filter band pass, 470–490 nm; dichroic mirror reflection short pass, 510 nm; suppression filter bandwidth, 520 nm). TRITC fluorescence was detected using a Leitz “N-2.1” filter cube (exciting filter band pass, 546/14 nm; dichroic mirror reflection short pass, 580 nm; edge wavelength, 580 nm). There was no cross-detection between the FITC- and TRITC-selective dichroic mirror-filter cubes.
Isolation of myenteric ganglia. Ganglia were isolated from dissected LMMP preparations by a modification of the method described previously (Fiorica-Howells et al., 1993). This procedure takes advantage of the absence of collagen from the interior of the myenteric plexus and the role of glia in providing support for neurons (Gershon et al., 1994). When the LMMP is exposed to collagenase, therefore, the ganglia of the myenteric plexus remain largely intact, whereas the non-neuronal components of the preparation dissociate and form a suspension of single cells (Yau et al., 1989). When the resulting suspension of dissociated cells and intact ganglia is filtered through a wide-pore (8.0 μm) filter (Nucleopore), the individual cells pass though, whereas the ganglia are trapped on the filter (Fiorica-Howells et al., 1993). Studies using desmin immunoreactivity as a muscle marker and neuron-specific enolase as a neuronal marker have verified that the filtered preparations are highly enriched with ganglia and not contaminated by muscle.
For immunoblotting, the ganglia were scraped from the filters and homogenized in iced buffer [50 mm Tris, pH 7.5, 0.15m NaCl, 1% Triton X-100, 25 μg/ml each of the peptidase inhibitors leupeptin and aprotinin (Sigma), 2 mm EDTA, and 1.0 mm EGTA]. The homogenate was spun for 10 min in a microcentrifuge, and the proteins in the resulting supernatant were separated by electrophoresis through 10% PAGE. The proteins were then blotted onto nitrocellulose membranes and probed with G-protein-purified rabbit polyclonal antibodies selective for the α, β, γ, δ, ε, and ζ isozymes of PKC. Immunoreactive bands were demonstrated with goat anti-rabbit secondary antibodies conjugated to horseradish peroxidase. Horseradish peroxidase activity was visualized with TMB peroxidase substrate (Kirkegaard & Perry).
Protein kinase C-mediated protein phosphorylation and translocation. Isolated myenteric ganglia were incubated for 10 min at 37°C in Krebs’ solution with 1.0 μm 5-HT, 10.0 μm 5-HT, or 1.0 μm PDBu. Reactions were terminated by addition of 0.5 mm EGTA in Ca2+- free Krebs’ solution. Preparation of tissue fractions and assessment of PKC activity was performed as described previously (Friedman and Wang, 1989). All subsequent procedures were carried out at 4°C, unless otherwise indicated. Tissues were harvested and homogenized in 10 vol of buffer A (20.0 mm Tris-HCl, pH 7.5, 0.32m sucrose, 2.0 mm EDTA, 0.5 mmEGTA, 50.0 μg/ml leupeptin, 0.1% 2-mercaptoethanol, and 0.2 mm phenylmethylsulfonyl fluoride), and the homogenate was centrifuged at 800 × g for 10 min. The supernatant obtained was sonicated (Kontes Micro Cell Disrupter) and centrifuged at 25,000 × g for 15 min. The resultant supernatant was removed, diluted to 1.0 ml with buffer A, chromatographed on diethylaminoethyl cellulose (DE52, Whatman, Maidstone, UK) anion exchange columns and used as the cytosolic fraction. The pellet was resuspended in 1.0 ml buffer A with 0.2% Nonidet P-40 and solubilized on ice for 1 hr. The sample was centrifuged at 25,000 ×g for 15 min. The resultant supernatant was chromatographed on DE52 columns, and the eluate was used as the membrane extract. Samples were applied to 1.0 ml DE52 columns equilibrated in buffer A, and the columns were washed with 5.0 ml of buffer A followed by 0.5 ml of 20 mm NaCl in buffer A. Enzyme was eluted with 0.75 ml of 200 mm NaCl in buffer A, and the eluate was used immediately for assessing PKC activity.
The standard assay mixture (250 μl) containing 24.0 mm Tris-HCl, pH 7.5, 20.0 mm NaCl, 0.1 mm EGTA, 0.4 mm EDTA, 0.03% 2-mercaptoethanol, 60 μg/ml leupeptin, 0.04 mm phenylmethylsulfonyl fluoride, 0.25 mg/ml, histone type III-s (Sigma), 1.2 mmCaCl2, 20 μg/ml phosphatidyl-l-serine, 8.0 nm phorbol-12-myristate,13-acetate, 10.0 mmMg(CH3CO2)2, and 0.03 mm [32P]ATP (500,000 cpm, DuPont, Boston, MA) was preincubated at 30°C for 5 min, and the reaction was initiated by the addition of eluted protein. After 1 min, the reaction was terminated by transferring 125 μl onto a 2 × 4 cm phosphocellulose (Whatman P81) strip that was subsequently immersed in 75.0 mm phosphoric acid (10.0 ml per strip). The strips were washed three times (2 min per wash) in fresh phosphoric acid and air dried. The strips were then placed in scintillation fluid, and radioactivity was determined by liquid scintillation spectrometry (LKB RACKBETA). PKC activity was defined as the phosphorylation that occurred in the presence of phosphatidyl-l-serine and phorbol-12-myristate,13-acetate and expressed as pmol32Pi-incorporated per unit protein of column eluate. Protein was determined by the method of Lowry et al. (Lowry et al., 1951).
RESULTS
5-HT evokes a uniphasic slow depolarization in AH/type 2 neurons when receptors for 5-HT receptor subtypes other than 5-HT1Pare antagonized
Recordings were made from cells that were classified as AH/type 2 neurons. Criteria used in classification included (1) the presence of an AH, and (2) a Ca2+ shoulder on the falling phase of the action potential (Gershon et al., 1994; Wood, 1994). A total of 235 AH/type 2 neurons were studied with a mean resting membrane potential of 73 ± 1 mV and an input resistance of 108 ± 5 MΩ. Except where otherwise stated, the 5-HT1A antagonist NAN-190 (0.3 μm) and the 5-HT3/4 dual antagonist tropisetron (1.0 μm) were added to the superfusing medium so that the other subtypes of 5-HT receptor to which these cells are known to respond, would not interfere with 5-HT1P-mediated responses. Tetrodotoxin (1.0 μm) was also present to confine recordings to postsynaptic events in the impaled neurons. In a series of 81 AH/type 2 cells studied under these conditions, 71 responded to the microejection of 5-HT (Fig. 1). The response was uniphasic under these conditions and consisted of a prolonged (104 ± 6 sec) membrane depolarization (16 ± 1 mV) associated with an increase in input resistance (mean increase = 102 ± 8%). No response to 5-HT was observed in 10 of the 81 neurons; such cells were not studied further. Superfused 5-HT (1.0 μm) reduced the amplitude of the AH from 14 ± 1 mV (n = 14) in control preparations to 7 ± 3 mV (n = 5; p < 0.02). The duration of the AH was also reduced by 5-HT from 13 ± 2 sec in control preparations to 6 ± 2 sec (p < 0.05).
Fig. 1.

5-HT induces a slow depolarization in AH/type 2 cells during which membrane conductance decreases and the AH is inhibited. A, In the presence of the 5-HT1Aantagonist NAN-190 and the 5-HT3/4 antagonist tropisetron, the response to 5-HT is a uniphasic depolarization associated with an increase in input resistance. The top trace depicts the membrane potential, and the bottom trace indicates current injected through the recording pipette. The downward deflections in the top trace represent the electrotonic responses to the injections of hyperpolarizing current pulses.B, At a faster sweep speed, the pronounced AH (top trace) can be seen after the action potential in the control record. The addition of 5-HT (1.0 μm) to the superfusing medium blocks the AH. The bottom tracedepicts depolarizing current injected through the recording pipette to evoke action potentials.
The slow response to 5-HT is Go protein-dependent
The hypothesis that the slow response of AH/type 2 neurons to 5-HT is mediated by a Go protein was tested. Initial studies were carried out to determine whether the previous report (Wang et al., 1996) that slow responses to 5-HT are potentiated by the intracellular injection of GTPγS and inhibited by GDPβS could be confirmed in preparations in which responses to subtypes of 5-HT receptor other than 5-HT1P were blocked. Subsequent experiments evaluated the sensitivity of the slow response to inhibition by PTx, which would be expected to antagonize a Go-mediated response (Casey and Gilman, 1988). Finally, the effects of the intracellular injection of Fab fragments of antibodies to Gαo into AH/type 2 neurons were ascertained. To study the effects of the guanine nucleotides on responses to 5-HT, recordings were made with beveled electrodes loaded with GTPγS or GDPβS. Control recordings were obtained first and, after stable responses to 5-HT were observed, the cells were injected with the guanine nucleotide by passing a negative DC current and simultaneously applying pressure to the recording electrode. The amplitude of control responses to 5-HT and that of responses to 5-HT recorded 10–15 min after the injection of GTPγS or GDPβS were measured and their ratios determined (Fig. 2). Intracellular injection of GTPγS more than doubled the amplitude of the response to 5-HT (p < 0.05;n = 6); furthermore, after the intracellular injection of GTPγS, the depolarization induced by the microejection of 5-HT was greatly prolonged and the membrane potential did not fully return to the resting level. In contrast, intracellular injection of GDPβS reduced the amplitude of the response to 5-HT to ∼35% of that of the control (Fig. 2) (p < 0.001; n= 6).
Fig. 2.

The slow response to 5-HT is inhibited by the intracellular injection of GDPβS and potentiated by the intracellular injection of GTPγS.
To study the effects of PTx, we initially tried to obtain control responses to 5-HT and then to maintain impalements long enough, after the addition of PTx (1.0 μg/ml) to the medium, to determine whether the slow response to 5-HT was affected by the toxin. Unfortunately, PTx is very slow to take effect when it is applied extracellularly. As a result, very long impalements (4–6 hr) were necessary to investigate the action of PTx. The membrane properties of AH/type 2 cells, even in control experiments, were found to be unstable during such long impalements; consequently, although responses to 5-HT were found eventually be inhibited in cells exposed to PTx (Wang et al., 1996), it was difficult to be certain that the inhibition was a specific effect of PTx. To circumvent this problem and to maximize the effectiveness of exposure to PTx, preparations were incubated with PTx for 6 hr before recordings were obtained. Controls were incubated similarly, but PTx was not present in the medium. As an additional control, to evaluate possible nonspecific effects of incubation with PTx, the responses of AH/type 2 neurons to forskolin were studied in the same cells used to investigate responses to 5-HT. Forskolin, which activates adenylate cyclase, evokes a slow depolarization similar to that elicited by 5-HT, but the response to forskolin (Nemeth et al., 1986; Bertrand and Galligan, 1995) would not be expected to be PTx-sensitive. The ratios of the amplitude of the 5-HT response to that of forskolin were computed for control cells and cells exposed to PTx. After incubation ± PTx, therefore, AH/type 2 cells were identified and their responses to 5-HT and forskolin determined (Fig.3). The amplitude of the response to 5-HT was significantly lower (p < 0.0001) in cells incubated with PTx (2 ± 1 mV; n = 8) than in controls (14 ± 1 mV; n = 35). The amplitude of the response to forskolin was also somewhat lower in cells incubated with PTx (7 ± 1 mV; n = 8) than in controls (15 ± 1 mV; n = 35). The ratio of the amplitudes of responses to 5-HT to those of forskolin (Fig. 3), however, was 1.05 ± 0.09 in control cells and 0.34 ± 0.07 after exposure to PTx (p < 0.0001). The response of AH/type 2 neurons to 5-HT thus was much more strongly affected by exposure to PTx than were the responses of the same cells to forskolin. These observations indicate that the slow response to 5-HT is PTx-sensitive.
Fig. 3.

Slow responses to 5-HT are inhibited by pertussis toxin (PTx). Preparations were superfused with PTx (1.0 μg/ml) for 6 hr. Control preparations were superfused for an equivalent period of time but in the absence of PTx. To control for nonspecific effects of prolonged superfusion, responses of AH/type 2 cells to 5-HT were compared with the similar responses of the same cells to forskolin in the presence or absence of PTx. The graph shows the ratio of the 5-HT to the forskolin response for control and PTx-treated preparations.
Preliminary studies revealed that AH/type 2 neurons could not consistently be injected through the recording micropipette with whole antibodies to Gαo. Preparations in which injections were attempted were fixed and later immunostained with biotinylated secondary antibodies and avidin-FITC to determine whether antibodies to Gαo had actually entered cells. In practice, the pipettes filled with whole antibodies tended to clog, leading to increases in electrode tip resistance, and antibodies only rarely entered impaled cells in quantities adequate for immunocytochemical detection. Fab fragments were therefore prepared and used for intracellular injection. In contrast to whole antibodies, the Fab fragments consistently entered injected cells. An initial control was to inject cells with Fab fragments of anti-Gαs. Although these injections did not inhibit slow responses to 5-HT, the effects of anti-Gαo and anti-Gαs could not be compared in the same cells. To determine whether injected anti-Gαo exerts nonspecific effects, therefore, responses to 5-HT in anti-Gαo-injected neurons were studied in the absence of tropisetron, so that the 5-HT3-mediated fast response to 5-HT (Mawe et al., 1986; Derkach et al., 1989) could be measured simultaneously with the 5-HT1P-mediated slow response and used as a control. Before the injection of anti-Gαo Fab fragments, therefore, the response to 5-HT was biphasic instead of monophasic. A rapidly developing transient depolarization (the fast response, which is known to be 5-HT3-mediated) (Mawe et al., 1986; Wade et al., 1991) preceded the much more slowly developing and longer-lived 5-HT1P-mediated slow response. A second control was to determine the effect of injection of anti-Gαo Fab fragments on responses of AH/type 2 neurons to histamine. Histamine evokes a slow depolarization similar to that elicited by 5-HT; however, the response to histamine is thought to be attributable to the activation of adenylate cyclase (Wood, 1994) and, thus, is probably coupled to a Gs protein. Intracellular injection of anti-Gαo Fab fragments was found to substantially inhibit the slow response to 5-HT, but these injections had little or no effect on fast responses to 5-HT of the same neurons (Fig.4A,B) and did not significantly affect slow responses of these cells to histamine (Fig.4A, ii). The amplitude of the slow response to 5-HT was reduced by injection of anti-Gαo Fab fragments from 13 ± 1 mV to 7 ± 1 mV (n = 23; p < 0.001). In contrast, the mean amplitude of the fast response in the same cells was 14.0 ± 1.8 mV before injection and 14.5 ± 2.1 mV (n = 16; pis not significant). These data support the hypothesis that the 5-HT1P receptor is coupled to a Go protein.
Fig. 4.
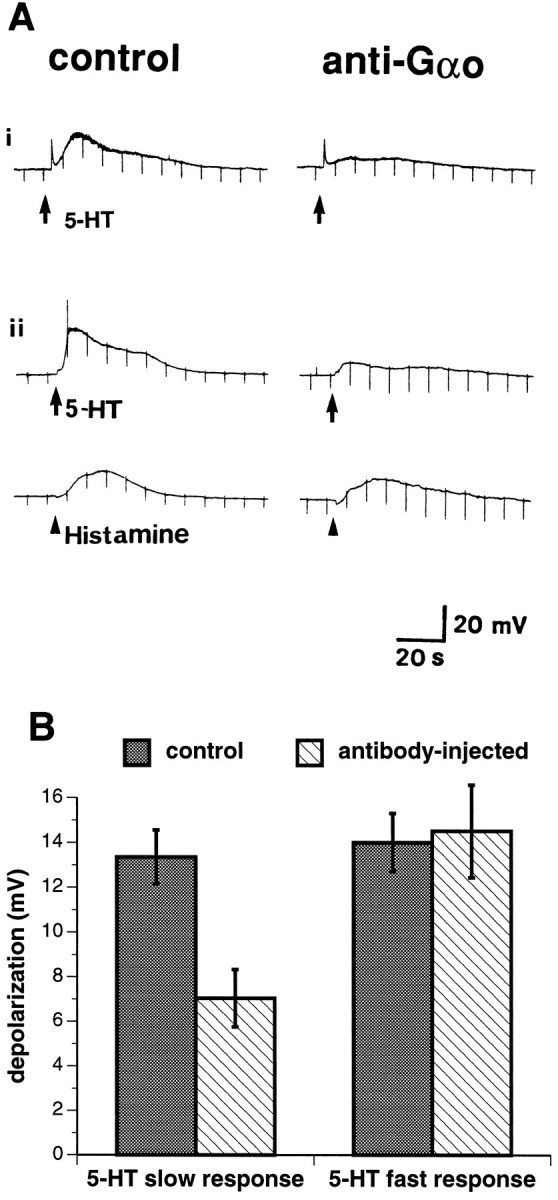
Intracellular injection of anti-GαoFab fragments inhibits slow responses to 5-HT. A,i, In the absence of tropisetron, 5-HT evokes a biphasic response (control), consisting of a fast 5-HT3-mediated depolarization, followed by the 5-HT1P-mediated slow depolarization. The injection of anti-Gαo Fab fragments antagonizes the slow but not the fast response to 5-HT. ii, Control responses of the same neuron to 5-HT and histamine are shown. The control response to 5-HT is again biphasic; the fast response is very small. An action potential is seen during the depolarization. Injection of anti-Gαo Fab fragments inhibits the slow response to 5-HT far more than that to histamine. The mean ratio of the amplitude of slow responses to histamine after injection of anti-Gαo Fab fragments to control responses obtained in the same cells (1.3 ± 0.1) was not significantly different from 1 (n = 6;p = 0.0804). B, The effects of intracellular injection of anti-Gαo Fab fragments on fast and slow responses of AH/type 2 neurons to 5-HT are compared.
Enteric neurons express Gαo immunoreactivity
The presence of Gαo in enteric neurons and their processes was investigated by immunocytochemistry. Gαoimmunoreactivity was observed to be widely distributed in neurons of both the submucosal (Fig.5A,B) and myenteric (Fig. 5C) plexuses. The immunoreactivity was strikingly marginal in these cells and appeared to be concentrated in their plasma membranes (Fig. 5B,C). The cell bodies of neurons thus appeared as negative images outlined by Gαo-immunoreactive membranes. Gαoimmunoreactivity was also abundant in the processes of these neurons and thus was seen in interganglionic connectives, as well as in ganglia. Calbindin immunoreactivity, which has been used as a marker for AH/type 2 neurons and is found in the majority of these cells (Pompolo and Furness, 1988; Furness et al., 1990) was demonstrated in the same preparations to determine whether there was a coincident cellular distribution of the two markers (Fig.5C,D). All calbindin-immunoreactive neurons (Fig. 5D) were ringed by Gαo-immunoreactive membranes (Fig. 5C); however, Gαo immunoreactivity was more widely distributed than that of calbindin and also ringed cells that did not contain calbindin. Gαo immunoreactivity was not observed in the intestinal musculature, but it was abundant in the ganglia and interganglionic connectives of the submucosal plexus (Fig.5A). In the submucosa, Gαo immunoreactivity was not confined to neural tissue but was also found in the smooth muscle of arteries (Fig. 5A). Within submucosal ganglia, the localization of Gαo immunoreactivity was similar to that seen in the myenteric plexus. Again, Gαo immunoreactivity was concentrated around neurons, which appeared as negative images (Fig. 5B). Because submucosal neurons were as Gαo immunoreactive as myenteric neurons, we evaluated the response of submucosal AH neurons to 5-HT. In the presence of tetrodotoxin (1.0 μm), tropisetron (1.0 μm), and NAN 190 (0.3 μm), 5-HT evoked a slow response that was very similar to that observed in AH/type 2 neurons of the myenteric plexus (Fig. 5E).
Fig. 5.

Gαo immunoreactivity is present in most submucosal and myenteric neurons. A, Both ganglia and interganglionic connectives of the submucosal plexus contain Gαo immunoreactivity. Connective tissue cells of the submucosa do not exhibit Gαo immunoreactivity; however, the smooth muscle of submucosal arteries (arrow) and associated paravascular nerve fibers are also Gαoimmunoreactive. B, At higher magnification, the Gαo immunoreactivity within a submucosal ganglion can be seen to outline the membranes of component neurons. C, The Gαo immunoreactivity of the neurons of a myenteric ganglion (visualized with FITC) is also marginal and outlines neuronal plasma membranes. D, The same section as shown inC, but the filters are set to reveal calbindin immunoreactivity, which is visualized with TRITC. Calbindin-immunoreactive neurons are ringed by Gαoimmunoreactivity; however, so too are most of the neurons of the ganglion. Scale bars, 30 μm. E, An intracellular record obtained from a submucosal AH neuron showing a typical 5-HT-evoked slow response associated with an increase in input resistance.
Activators of PKC mimic the slow response to 5-HT
Because the slow response to 5-HT was found to be associated with the activation of a Go protein, it seems unlikely that the response is directly linked to adenylate cyclase, which is normally activated by Gs. The hypothesis that the slow response to 5-HT is PKC-dependent was therefore tested. Each of three different activators of PKC mimicked the slow response to 5-HT. When applied by microejection onto the surface of AH/type 2 neurons, PDBu induced a slow depolarization (Fig. 6A,record 1) with a mean amplitude of 16 ± 2 mV (n = 8) associated with an increase in input resistance of 87 ± 15%. The duration of the slow depolarization was extremely long (>10 min). A similar, but even longer-lasting, response was evoked by adding PDBu (1.0 μm) to the superfusing medium (Fig. 6A, record 2). Almost identical responses were elicited by OAG or (−)-7-octylindolactam V (data not shown). In addition to the slow depolarization evoked by microejection of PDBu, the AH, which is inhibited by superfused 5-HT (Figs. 1B, 6B) was also inhibited by superfusion of PDBu (1.0 μm) or OAG (100 μm) (Fig. 6A, record 3,B).
Fig. 6.
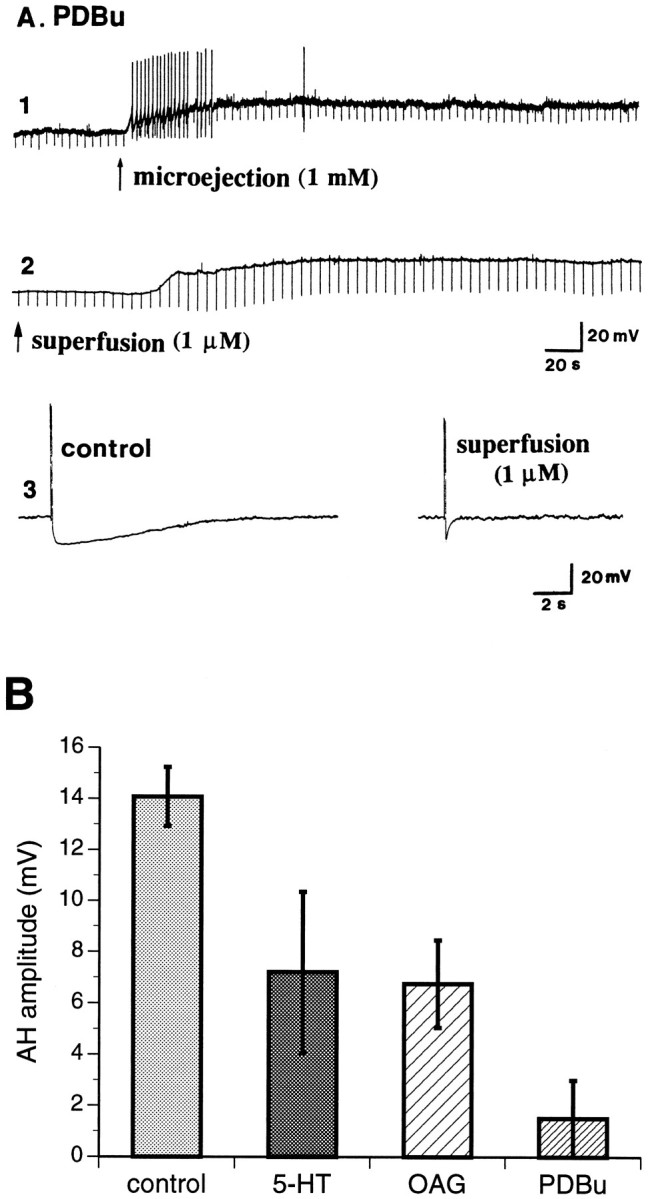
Activators of PKC mimic slow responses to 5-HT.A, The phorbol ester PDBu evokes a slow depolarization in AH/type 2 neurons associated with an increase in input resistance, no matter whether it is applied by microejection (1) from a pipette positioned close to the impaled neuron or by superfusion (2) in the ambient medium. Superfusion of PDBu also inhibits the AH (3) that follows the action potential in these cells. Note that although the effects of PDBu are qualitatively similar to those of 5-HT, the response to PDBu is much longer lasting.B, The effects of 5-HT (1.0 μm), OAG (100 μm), and PDBu (1.0 μm) on the amplitude of the AH in AH/type 2 neurons are compared. All three compounds inhibit the AH; PDBu is the most potent.
PKC antagonists inhibit slow responses of AH/type 2 neurons to 5-HT
The observation that activators of PKC mimic the slow response of AH/type 2 neurons to 5-HT suggests that PKC might be involved in the transduction of responses mediated by the 5-HT1P receptor. To determine whether PKC actually does play such a role, the ability of PKC inhibitors to antagonize slow responses to 5-HT was determined. Superfusion of stau- rosporine (1.0 μm) (data not illustrated) and K-252a (3.0–10 μm) (Fig.7B,C), both of which inhibit PKC, antagonized slow responses of AH/type 2 neurons to 5-HT. These compounds, however, do not discriminate sufficiently well between PKC and other kinases to enable their antagonism of slow responses to 5-HT to be attributed specifically to inhibition of PKC. Studies were thus carried out with more selective PKC antagonists. These included the pseudosubstrate peptide PKC[19–31] and the membrane permeable PKC inhibitors chelerythrine (3.0 μm), Gö 6976 (10.0 μm), and myr-FARKGALRQ (40 μm) (Martiny-Baron et al., 1993). Because PKC[19–31] does not cross cell membranes, it was injected into AH/type 2 neurons through the recording micropipette. Superfusion with chelerythrine (Fig. 7A–C), Gö 6976 (Fig. 7A), or myr-FARKGALRQ (Fig.7A) and injection of PKC[19–31] (Fig.7A–C) all inhibited slow responses to 5-HT. For these experiments, control responses to 5-HT were obtained and, when these were stable in amplitude, PKC[19–31] was injected into cells or the membrane permeable antagonists were added to the superfusing medium. The ratio of the amplitude of the response to 5-HT after the injection of PKC[19–31] or superfusion of chelerythrine to that of the immediate preceding control response was determined (Fig. 7C). Each cell thus served as its own control. A ratio of 1.0 would have been obtained if PKC[19–31] or chelerythrine had failed to antagonize slow responses to 5-HT. In fact, the ratios obtained for both of the specific PKC inhibitors were significantly less than 1.0 [p < 0.001 for PKC[19–31](n = 12); p < 0.01 for chelerythrine (n = 8)]. Similarly, a ratio of less than 1.0 was also obtained for the less PKC-specific K252a [p < 0.05 (n = 3)] (Fig. 7C). The membrane-permeable myristolated peptide myr-FARKGALRQ also reduced the amplitude of the slow response to 5-HT from 22.0 ± 2.0 mm to 7.0 ± 2.0 mV; in contrast to the amplitude of slow responses, the amplitude of fast responses to 5-HT before addition of myr-FARKGALRQ, 12.5 ± 6.5 mV, was not significantly different from that obtained after exposure to the peptide, 14.5 ± 7.5 mV (n = 3).
Fig. 7.

Inhibitors of PKC antagonize slow responses to 5-HT. A, Slow responses to 5-HT of an AH/type 2 neuron are antagonized by the intracellular injection of the PKC pseudosubstrate PKC[19–31] (top trace,left) and by the superfusion of the membrane permeable PKC pseudosubstrate myr-PKC[19–27] (myr-FARKGALRQ;bottom trace, left). Slow responses are also inhibited by the superfusion of chelerythrine (3.0 μm; top trace, right) and by the selective PKC α and β antagonist Gö 6976 (10 μm; bottom trace,right). B,C, The abilities of PKC[19–31] chelerythrine and K252a to reduce the amplitude of the 5-HT-induced slow depolarization of AH/type 2 cells are compared.
Downregulation of PKC antagonizes slow responses to 5-HT
The observations that six different inhibitors of PKC each antagonized the slow response of AH/type 2 neurons to 5-HT supports the idea that PKC plays a critical role in the transduction of responses mediated by the 5-HT1P receptor. If so, then the downregulation of PKC would be expected to interfere with slow responses to 5-HT. To downregulate PKC, dissected preparations of LMMP were cultured overnight in the presence of PDBu (10 μm). Control preparations were cultured similarly, but in the absence of PDBu. The amplitude of slow responses of AH/type 2 neurons to 5-HT in the preparations exposed overnight to PDBu was compared with that of slow responses to 5-HT in the controls. To test the possibility that a decrease in the amplitude of slow responses to 5-HT in cells chronically exposed to PDBu might have been attributable to nonspecific effects, responses to forskolin were also obtained from each of the cells used for testing responses to 5-HT. Responses to forskolin, which stimulates adenylate cyclase, would not be expected to be dependent on PKC; moreover, the slow depolarizing response of AH/type 2 neurons to forskolin (amplitude, 16 ± 1.0 mV; duration, 165 ± 9.0 sec, increase in input resistance of 107 ± 11.8%; n = 46) is very similar to the slow response of these cells to 5-HT. The ratio of the amplitudes of the slow responses of each neuron to 5-HT to that of the same cells to forskolin was determined. In preparations that were not exposed to PDBu, the amplitudes of slow responses to 5-HT and forskolin were about equal (Fig.8A) (n = 32); however, in preparations that were treated with PDBu, the amplitudes of the responses of AH/type 2 neurons to 5-HT were quite small and considerably less than those to forskolin (Fig. 8B) (n = 11). As a result, the 5-HT to forskolin ratio was significantly reduced by the chronic exposure of preparations to PDBu (Fig. 8C) (p < 0.001). Chronic exposure to PDBu also reduced the duration of slow responses to 5-HT and the associated increase in input resistance. The ratio of the duration of the 5-HT response to that of forskolin fell from 0.7 ± 0.1 in untreated preparations to 0.4 ± 0.1 in those exposed to PDBu (p < 0.05). The ratio of the maximal change in input resistance during the 5-HT response to that during the response to forskolin decreased from 1.3 ± 0.2 in untreated preparations to 0.3 ± 0.2 in preparations treated chronically with PDBu (p < 0.001). These observations indicate that the slow response of AH/type 2 neurons is inhibited specifically by downregulation of PKC.
Fig. 8.

Downregulation of PKC inhibits the slow response of AH/type 2 neurons to 5-HT. A, In control preparations, the amplitudes of the slow depolarizations evoked by 5-HT and forskolin are approximately equal. B, After overnight exposure of preparations to PDBu to downregulate PKC, the 5-HT-induced slow depolarization is reduced in amplitude far more than is the response to forskolin. C, In control preparations, the ratio of the amplitude of the 5-HT to the forskolin response is about 1; however, this ratio is greatly reduced as a result of the inhibition of the response to 5-HT after long-term exposure of preparations to PDBu.
5-HT activates PKC in isolated myenteric ganglia
To test directly whether 5-HT activates PKC in situ, we measured the membrane association of PKC activity ± the exposure of isolated myenteric ganglia to 5-HT. As a positive control, the membrane association of PKC activity was also followed in preparations exposed to PDBu. The action of 5-HT was investigated in isolated ganglia to avoid interference from effects 5-HT might exert on smooth muscle. The isolated ganglia, trapped on filters, were equilibrated for 5 min at 37°C in oxygenated Krebs’ solution, to which was added the 5-HT1A antagonist NAN-190 (0.3 μm), the 5-HT2 antagonist ketanserin (10 μm), and the 5-HT3/4 dual antagonist tropisetron (1.0 μm) to block responses to subtypes of 5-HT receptor other than 5-HT1P. 5-HT (1.0 or 10.0 μm) or PDBu (1.0 μm) were then applied for 10 min in the continued presence of the cocktail of antagonists. Control preparations were incubated similarly in the absence of 5-HT. After incubation, all preparations were washed for 5 min with iced Krebs’ solution and transferred to iced buffer for homogenization. PKC activity was measured in both the membrane and supernatant fractions. Exposure to 5-HT induced a concentration-dependent increase in the membrane associated PKC activity (Table 1). At 10 μm, the response to 5-HT was ∼75% that to PDBu.
Table 1.
PKC translocation in myenteric ganglia
| PKC activity | |||
|---|---|---|---|
| Membrane bound | Cytosol | M/C ratio | |
| (pmol/min/μg protein) | |||
| Control | 2.09 ± 0.08 | 2.90 ± 0.11 | 0.72 ± 0.03 |
| 5-HT (1.0 μm) | 2.72 ± 0.16* | 2.23 ± 0.06* | 1.22 ± 0.07* |
| 5-HT (10.0 μm) | 3.17 ± 0.19* | 1.96 ± 0.03* | 1.61 ± 0.10* |
| PDBu (1.0 μm) | 3.86 ± 0.04* | 1.78 ± 0.04* | 2.17 ± 0.05* |
The ability of the PKC α/β pseudosubstrates myr-FARKGALRQ and PKC[19–31] as well as the α/β isozyme-selective Gö 6976 to antagonize slow responses to 5-HT suggests that one of the classical isozymes of PKC is likely to be responsible for the effect.
P < 0.01, compared with control. Data are mean ± SEM. n = 3–6.
Inhibition of PC-PLC antagonizes slow responses to 5-HT
The observations that the slow response of AH/type 2 neurons to 5-HT are dependent on a Go protein and PKC raise the question of how Go, after its activation by the 5-HT1P receptor, is coupled to PKC. This coupling could be dependent on the activity of a PLC; however, because the slow response to 5-HT is attributable to the inhibition of gKCa, it would seem unlikely that the activation of PKC would be linked to a mechanism that would increase intracellular Ca2+([Ca2+]i). The hypothesis was therefore tested that the slow response depends not on a phosphatidylinositol-specific isozyme of PLC but on PC-PLC. The liberation of inositol-1,4,5-trisphosphate (IP3) by a phosphatidylinositol-specific PLC would be expected to cause the release of Ca2+ from internal stores and thus to increase [Ca2+]i (Exton, 1994). In contrast, the action of PC-PLC would liberate the PKC activator, DAG, without simultaneously liberating IP3 (Schütze et al., 1992). The effect of D609 (100 μm), a specific inhibitor of PC-PLC (Schütze et al., 1992), on the slow response of AH/type 2 neurons to 5-HT was therefore tested. Although D609 did not affect the membrane potential of AH/type 2 cells, the amplitude of the 5-HT-induced slow depolarization was significantly reduced by D609 (Fig. 9A,B) (p < 0.001; n = 9).
Fig. 9.

An inhibitor of PC-PLC antagonizes slow responses to 5-HT. A, Slow responses to 5-HT are inhibited by the PC-PLC inhibitor, D609. B, The amplitudes of 5-HT-evoked slow depolarizations in the presence or absence (control) of D609 are compared.
PKA antagonism inhibits slow responses of AH/type 2 neurons to 5-HT
The data outlined above, which suggest that PKC participates in mediating slow responses of AH/type 2 neurons to 5-HT, do not imply that PKC is the only protein kinase that does so. Because the downregulation or inhibition of PKC antagonized, but did not abolish, slow responses to 5-HT, it is possible that another protein kinase contributes to the response. To test the hypothesis that PKA contributes to the slow response of AH/type 2 cells to 5-HT, the selective PKA inhibitor Rp-cAMPS was injected into AH/type 2 neurons through the recording electrode. Responses to 5-HT, applied by microejection to the surfaces of AH/type 2 neurons, were then determined. The ability of injected Rp-cAMPS to effect the slow depolarization evoked in the same cells by pressure applications of the PKC activator PDBu was assessed as an expected negative control. Forskolin and histamine were also studied as expected positive controls, because they increase cAMP and thus probably act via PKA. The resemblance of the response of AH/type 2 neurons to forskolin has been described above. When applied by microejection to the surfaces of AH/type 2 neurons, histamine evoked a similar slow depolarization (amplitude, 20 ± 1.2 mV; duration, 93 ± 14.5 sec; increase in input resistance, 100 ± 0%; n = 3). As expected, Rp-cAMPS blocked both the responses of AH/type 2 neurons to histamine and those to forskolin (data not shown). Because Rp-cAMPS rapidly diffused into impaled cells from the recording electrode, even in the absence of pressure or application of current, it was impossible to be certain that responses to 5-HT or PDBu obtained before the injection of Rp-cAMPS were not affected by the compound. The mean amplitudes of the pooled responses to 5-HT (n = 8) or PDBu (n = 4) obtained from neurons after the intracellular injection of Rp-cAMPS were thus compared with the mean amplitudes of the pooled responses to 5-HT (n = 81) or PDBu (n = 8) obtained from control neurons. Intracellular injection of Rp-cAMPS significantly (p < 0.001) reduced the amplitude of the slow response of AH/type 2 neurons to 5-HT (Fig.10A,C) but did not affect the amplitude of the slow response of these cells to PDBu (Fig.10C). In addition to the reduction in the amplitude of slow responses to 5-HT caused by the intracellular injection of Rp-cAMPS, superfusion of Rp-cAMPS antagonized the ability of 5-HT to inhibit the AH of AH/type 2 neurons (Fig. 10B).
Fig. 10.
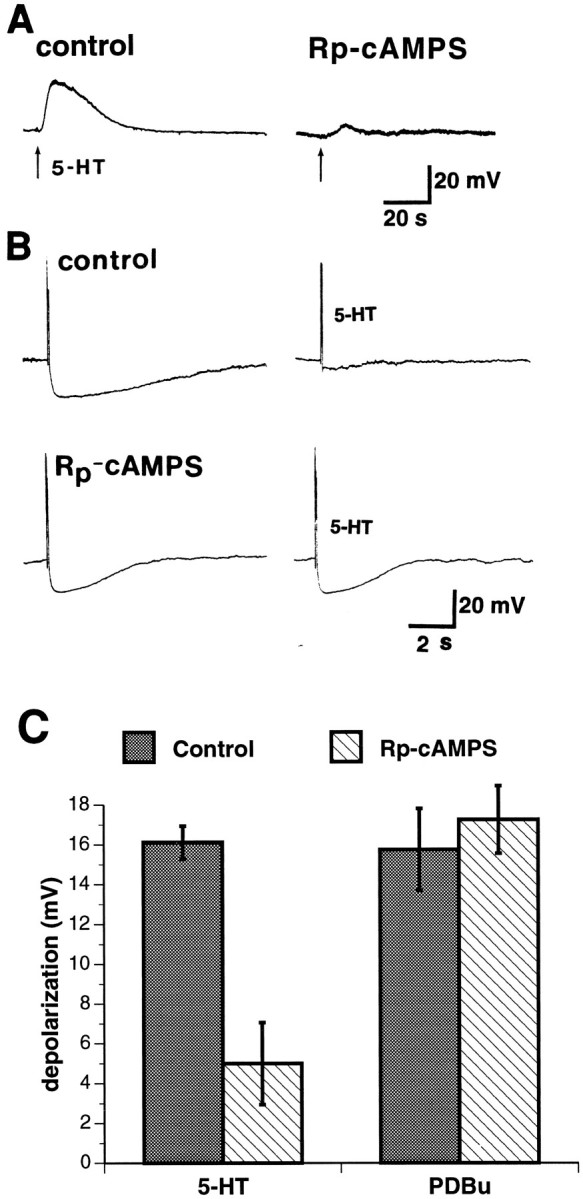
Inhibition of PKA antagonizes slow responses to 5-HT. A, Slow responses to 5-HT of an AH/type 2 neuron are antagonized by the intracellular injection of the PKA antagonist Rp-cAMPS. B, Under control conditions (top tracings), 5-HT (1.0 μm) inhibits the AH that follows the action potential in an AH/type 2 cell. Superfusion of Rp-cAMPS (200 μm; bottom tracings) does not itself affect either the action potential or the AH; however, superfusion of Rp-cAMPS prevents the inhibition of the AH by 5-HT.C, Although the slow depolarization of AH/type 2 neurons by 5-HT is inhibited by the intracellular injection of Rp-cAMPS, the slow depolarization evoked by PDBu is not affected by Rp-cAMPS.
Antagonism of adenylate cyclase inhibits slow responses of AH/type 2 neurons to 5-HT
The data obtained with Rp-cAMPS are consistent with the idea that PKA, as well as PKC, contributes to the transduction of responses mediated by the 5-HT1P receptor. The participation of PKA suggests that adenylate cyclase is also likely to play a role in the response. This hypothesis was tested by determining the effects of the specific adenylate cyclase inhibitor DDA (0.5 mm) on slow responses of AH/type 2 neurons to 5-HT. The addition of DDA to the superfusing medium did not affect the membrane potential but reduced the amplitude of the 5-HT-induced slow depolarization from 16.0 ± 3.7 mV before the addition of DDA to 4.0 ± 1.5 mV in the presence of the compound (p < 0.05; n = 5). These observations are consistent with the idea that adenylate cyclase participates in the transduction of 5-HT1P-mediated responses.
A variety of isozymes of PKC can be identified in enteric neurons
Antibodies to various isozymes of PKC were used to investigate the expression of PKC in the ENS by immunocytochemistry. Western blots were carried out with proteins extracted from isolated myenteric ganglia to confirm results obtained by immunocytochemistry. Calbindin immunoreactivity was located simultaneously with the PKC isozymes to identify probable AH/type 2 neurons in the same preparations. Of the classical (group A) isozymes, only PKC α immunoreactivity was extensively expressed in the enteric plexuses. The distribution of PKC α immunoreactivity was similar to that of Gαo. As with Gαo, the immunoreactivity of PKC α was strikingly marginal, forming rings around most myenteric neurons (Fig.11A). Virtually all of the calbindin-immunoreactive neurons (Fig. 11B) were surrounded by such rings of PKC α immunoreactivity (compare Fig.11A and B). In addition, PKC α immunoreactivity was abundant in the ganglionic neuropil and was found in processes coursing through interganglionic connectives. The immunoreactivity of PKC α was equally abundant in the submucosal plexus, where it again outlined most neurons and extended into interganglionic connectives (Fig. 11A,inset). In contrast to PKC α, almost no PKC β1 or β2 immunoreactivity could be detected in enteric neurons, although smooth muscle contained PKC β2 immunoreactivity (data not shown). Of the novel (group B) isozymes of PKC, only PKC δ immunoreactivity was present in abundance in the ENS. The pattern obtained with antibodies to PKC δ (Fig. 11C) was quite similar to that obtained with antibodies to PKC α. Again, the immunostaining of myenteric neurons was strikingly marginal and PKC δ immunoreactivity outlined myenteric nerve cell bodies, leaving the bulk of the neuronal cytoplasm unstained. All calbindin-immunoreactive neurons (Fig.11D) were surrounded by PKC δ immunoreactivity; however, PKC δ immunoreactivity was not specifically associated with calbindin-immunoreactive cells and surrounded almost all of the neurons of the myenteric plexus. PKC δ immunoreactivity was also found in the ganglionic neuropil and in axonal varicosities (Fig. 11C). The abundance of PKC δ immunoreactivity was only slightly less than that of PKC α but was more prominent than that of α in axonal varicosities. No specific immunostaining of the enteric plexuses were obtained with antibodies to another novel isozyme, PKC ε, although ε immunoreactivity was seen in the smooth muscle (data not shown). Of the atypical isozymes of PKC that were investigated, ζ, ι/λ, and η, none immunostained enteric neurons. Cells with an appearance and distribution consistent with that of peritoneal macrophages, however, were immunostained with antibodies to PKC η, which was also observed in circular muscle. The immunoreactivity of PKC γ, a classical enzyme, was observed in small numbers of myenteric axons, which were distinguished by very large varicosities (Fig. 11E). Weak PKC γ immunoreactivity was also observed in rare neurons (data not shown), which did not contain calbindin. In addition, small extraganglionic cells exhibited strong PKC γ immunoreactivity, which appeared to be located in intracellular granules (Fig.11E); these cells did not contain calbindin (comparedF and E). These observations indicate that only two isozymes of PKC, α and δ, are abundant in enteric neurons. Data obtained from Western blots of proteins obtained from preparations of isolated myenteric ganglia were consistent with the observations made by immunocytochemistry; thus, only PKC α, γ, and δ immunoreactivities were observed in isolated ganglia (data not shown).
Fig. 11.
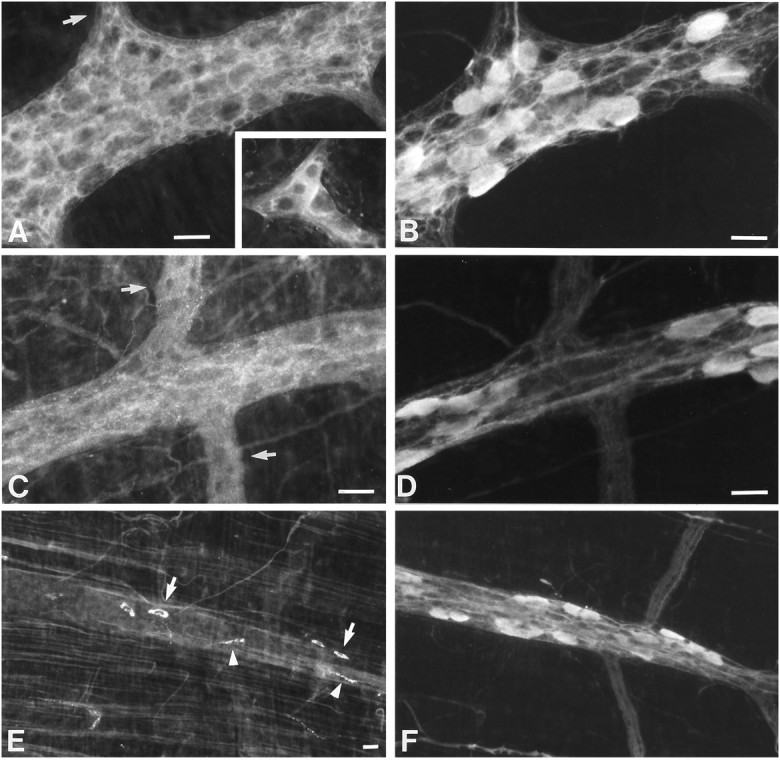
The immunoreactivities of PKC α and PKC δ are prominent in enteric neurons. A, PKC α immunoreactivity (visualized with FITC) is concentrated at the margins of neurons in both a myenteric and a submucosal ganglion (inset) and thus outline enteric neurons. Axons entering interganglionic connectives (arrows) are also PKC α-immunoreactive. B, Calbindin immunoreactivity (visualized with TRITC) is demonstrated in the same section as shown inA. Although calbindin-immunoreactive neurons are outlined by PKC α immunoreactivity, so too are virtually all of the neurons of the myenteric plexus. C, PKC δ immunoreactivity (visualized with FITC) is concentrated at the margins of neurons in a myenteric ganglion but does not outline cells quite as sharply as does the immunoreactivity of PKC α. Axons entering interganglionic connectives are also PKC δ-immunoreactive.D, Calbindin immunoreactivity (visualized with TRITC) is demonstrated in the same section as shown in C. Calbindin-immunoreactive neurons are outlined by PKC δ immunoreactivity, but again virtually all of the neurons of the myenteric plexus are ringed by PKC δ immunoreactivity.E, PKC γ immunoreactivity is seen in some highly granular cells (arrows) located outside of myenteric ganglia (above or next to the ganglia). In addition, smooth muscle cells contain PKC γ immunoreactivity, which is concentrated in their plasma membranes. The bright streaks running parallel to the nonimmunoreactive myenteric ganglion are the immunofluorescence of the membranes of circular muscle cells. Although only the appearance of PKC γ immunoreactivity in neurons is extremely rare, some very large axonal varicosities are PKC γ-immunoreactive (arrowhead). F, Calbindin immunoreactivity (visualized with TRITC) is demonstrated in the same section as shown in E. Note that neither the calbindin-immunoreactive neuronal cell bodies within the ganglion nor the calbindin-immunoreactive axons in the interganglionic connectives are PKC γ- immunoreactive. The very large PKC γ-immunoreactive varicosities are not calbindin-immunoreactive (compare withE). Scale bars, 30 μm.
DISCUSSION
The current studies confirm that the slow response of AH/type 2 neurons [inhibition of gKCa (Galligan, 1995)] to 5-HT is linked to Go and cooperatively mediated by PKA and PKC. Evidence that this response is Go-linked includes: (1) inhibition by intracellular injection of GDPβS and potentiation/prolongation by GTPγS; (2) sensitivity to PTx; (3) activation, by 5-HT1P agonists, of Go, but not of other G-proteins (Wang et al., 1996); (4) antagonism by intracellular injection of anti-Gαo Fab fragments of slow responses to 5-HT but not to histamine, or antagonism of fast, 5-HT3-mediated responses to 5-HT; and (5) the presence of Gαo immunoreactivity on the plasma membranes of many neurons in both plexuses, including all of those that express the AH/type 2 cell marker calbindin. Evidence that PKC plays a role in mediating slow responses to 5-HT includes: (1) mimicry by the PKC activators, PDBu, (−)-7-octylindolactam V, and OAG; (2) antagonism by PKC inhibitors that have little in common besides their ability to inhibit PKC, including staurosporine, K-252a, Gö6976, chelerythrine, and the pseudosubstrate peptides myr-FARKGALRQ and PKC[19–31] (by intracellular injection), as well as by (3) overnight exposure to PDBu to downregulate PKC; and (4) the concentration-dependent induction by 5-HT of PKC translocation into membranes in isolated myenteric ganglia. Evidence that PKA participates in mediating slow responses to 5-HT includes: (1) antagonism by the selective PKA inhibitor Rp-cAMPS of slow responses to 5-HT, histamine, and forskolin, but not those to PDBu; and (2) interference by Rp-cAMPS with inhibition of the AH by 5-HT.
Go is not commonly linked to the activation of PKC and PKA. Because the selective PC-PLC inhibitor D609 antagonizes slow responses to 5-HT, it seems likely that the coupling of Go to PKC is mediated by PC-PLC. The activity of this enzyme generates DAG, which activates PKC without simultaneously releasing IP3(Schütze et al., 1992), as would a phosphatidylinositol-specific isozyme. Because the slow response to 5-HT involves the inhibition of gKCa (Galligan, 1995), the liberation of IP3would be counterproductive, because IP3 would be expected to increase intracellular Ca2+ (Exton, 1994). In fact, the P2 purinoceptor-induced elevation of intracellular Ca2+ is associated with effects, a membrane hyperpolarization and decreased input resistance, that are the converse of the slow response to 5-HT (Palmer et al., 1987a; Christofi et al., 1995). The 5-HT1P-related activation of PKA is probably stimulated by an adenylate cyclase-mediated elevation of cAMP, because slow responses are antagonized by the adenylate cyclase inhibitor DDA and 5-HT increases cAMP in isolated myenteric ganglia (Xia et al., 1991, 1994; Fiorica-Howells et al., 1993). Slow responses to 5-HT are also mimicked by forskolin (Nemeth et al., 1986) and by analogs of cAMP (Zafirov et al., 1985a; Palmer et al., 1986). Despite these observations, available evidence suggests that Gs is not responsible for the 5-HT-induced activation of adenylate cyclase. When applied to isolated myenteric ganglia, 5-HT activates Gαo, but not Gαs, (Wang et al., 1996). The adenosine-induced stimulation of Gαi does not inhibit slow responses to 5-HT (or slow EPSPs) (Palmer et al., 1987b), but it does block slow responses to forskolin, histamine, vasoactive intestinal peptide, or pituitary adenylate cyclase activating peptide (Zafirov et al., 1985b; Palmer et al., 1987a,b; Christofi and Wood, 1993). Because adenylate cyclase thus appears to be involved in mediating both the Gαi-resistant slow responses to 5-HT and the Gαi-sensitive slow responses to forskolin, histamine, vasoactive intestinal peptide, or pituitary adenylate cyclase activating peptide, the activation of adenylate cyclase associated with the response to 5-HT must differ from that associated with forskolin or compounds that probably act via receptors linked to Gs. PKC is known to be able to activate type II adenylate cyclase (Jacobowitz et al., 1993; Yoshimura and Cooper, 1993); moreover, when adenylate cyclase is activated by PKC, instead of by Gαs or forskolin, it is not inhibited by Gαi (Pieroni et al., 1993). These considerations suggest that the following signal transduction pathway may be responsible for the slow response to 5-HT: 5-HT → 5-HT1P receptor → activates Go → activates PC-PLC → liberates DAG → stimulates PKC → activates adenylate cyclase → elevates cAMP → stimulates PKA. Because the slow response to 5-HT is not abolished by inhibitors of PKA or PKC alone but by their combination, PKA and PKC probably each inhibit gKCa. Why PKC inhibitors or downregulation do not eliminate the slow response to 5-HT is not clear. Conceivably, a Ca2+-independent novel or atypical isozyme of PKC, in addition to one of the classical (group A) Ca2+-dependent PKC isozymes, becomes activated by 5-HT and is not fully inactivated by PKC inhibitors or downregulation.
Immunocytochemical studies indicated that PKC α is present in most of the neurons of both submucosal and myenteric plexuses. In fact, the distribution of the immunoreactivities of Gαo and PKC α was similar and each was found in virtually every cell that displayed calbindin immunoreactivity, investigated as an AH/type 2 cell marker (Furness et al., 1990). In contrast, enteric neurons contained no PKC β1 or β2 immunoreactivity and PKC γ immunoreactivity was observed only in rare myenteric neurons, none of which were calbindin immunoreactive. Because many neurons in each plexus and most AH/type 2 neurons exhibit the slow response to 5-HT, PKC α is the only classical (group A) isozyme of PKC that is distributed in a manner consistent with a role in transduction of 5-HT1P-mediated slow responses. Subject to the limitation that the proteins might have been present in forms or quantities that could not be detected by immunocytochemistry, the β1, β2, and γ isozymes thus are unlikely to be involved in transducing the slow response. The distribution of the immunoreactivity of the novel (group B) isozyme PKC δ, however, was found to be very similar to that of PKC α. Again, many neurons in each plexus and all calbindin-immunoreactive neurons were PKC δ immunoreactive. PKC δ is thus also a candidate to play a role in the slow response to 5-HT. In contrast, the distributions of neither PKC ε, nor any of the atypical isozymes of PKC, ζ, ι/λ, and η, were compatible with roles in 5-HT slow response signal transduction. These observations suggest that only the α and δ PKC isozyme are potential participants in transduction of slow responses to 5-HT. At least some of the PKC inhibitors that were found to antagonize slow responses to 5-HT, however, including Gö 6976 and the pseudosubstrate peptides FARKGALRQ and PKC[19–31] are specific for the α and β isozymes of PKC and have little or no activity against PKC δ. The β PKC isozymes, moreover, are evidently not present in enteric neurons; thus, these data suggest that PKC α is probably primarily responsible for transducing the 5-HT1P-mediated slow response. Although PKC α is Ca2+-sensitive and intracellular Ca2+ is unlikely to increase during slow responses to 5-HT, the constitutive level of cytosolic Ca2+ may be sufficient to support the activation of PKC α by DAG, which increases the affinity of PKC for Ca2+ (Nishizuka, 1992; Tanaka and Nishizuka, 1994). Conceivably, local elevations of intracellular Ca2+ could occur, which could enhance stimulation of PKC α without contributing to gKCa.
Many characteristics of slow responses to 5-HT are similar to those that have been reported previously for slow EPSPs. For example, slow EPSPs are potentiated by GTPγS, mimicked by forskolin and PDBu, and inhibited by D609 and staurosporine (Bertrand and Galligan, 1995). No studies have yet been carried out to determine whether slow EPSPs are, like slow responses to 5-HT, affected by GDPβS, antibodies to Gαo, or inhibitors of adenylate cyclase or PKA. The one difference between slow responses to 5-HT and slow EPSPs is that only slow EPSPs have been reported to be PTx-resistant (Bertrand and Galligan, 1995). When slow EPSPs were studied, however, PTx was applied extracellularly for only 2 hr, whereas in the current investigation, at least 4 hr of exposure was found to be required before PTx inhibited slow responses to 5-HT. It is possible, therefore, that slow EPSPs might actually be PTx-sensitive but not observed to be so, because insufficient time was allowed for PTx to become effective. Both the slow response to 5-HT and slow EPSPs are resistant to inhibition by adenosine (Palmer et al., 1987b). The resistance of slow EPSPs to inhibition by adenosine implies that they are, like slow responses to 5-HT, Gαi-resistant. Thus it seems unlikely that any receptor that induces a slow EPSP could be coupled by a Gsprotein to adenylate cyclase; nevertheless, because 5-HT is only one of the transmitters that mediate slow EPSPs (Bornstein et al., 1984), the observation that PTx did not inhibit slow EPSPs (Bertrand and Galligan, 1995) could also be explained by the existence of subsets of slow EPSPs transduced by different transmitters using receptors coupled to different G-proteins. Given the many other identities between slow EPSPs and slow responses to 5-HT, however, it seems premature to postulate subsets of slow EPSPs triggered by different G-proteins.
Footnotes
This work was supported by National Institutes of Health Grant NS12969. H.P. is a fellow of the Pharmaceutical Manufacturers Association Foundation.
Correspondence should be addressed to Dr. Hui Pan, Department of Anatomy and Cell Biology, Columbia University (P&S 12-513), 630 West 168th Street, New York, NY 10032.
REFERENCES
- 1.Bertrand PP, Galligan JJ. Signal-transduction pathways causing slow synaptic excitation in guinea pig myenteric AH neurons. Am J Physiol. 1995;269:G710–G720. doi: 10.1152/ajpgi.1995.269.5.G710. [DOI] [PubMed] [Google Scholar]
- 2.Bornstein J, North RA, Costa M, Furness JB. Excitatory synaptic potentials due to activation of neurons with short projection in the myenteric plexus. Neuroscience. 1984;11:723–731. doi: 10.1016/0306-4522(84)90055-1. [DOI] [PubMed] [Google Scholar]
- 3.Branchek TA, Kates M, Gershon MD. Enteric receptors for 5-hdyroxytryptamine. Brain Res. 1984;324:107–118. doi: 10.1016/0006-8993(84)90627-9. [DOI] [PubMed] [Google Scholar]
- 4.Branchek TA, Mawe G, Gershon MD. Characterization and localization of a peripheral neural 5-hydroxytryptamine receptor subtype with a selective agonist, 3H-5-hydroxyindalpine. J Neurosci. 1988;8:2582–2595. doi: 10.1523/JNEUROSCI.08-07-02582.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Casey PJ, Gilman AG. G Protein involvement in receptor-effector coupling. J Biol Chem. 1988;263:2577–2580. [PubMed] [Google Scholar]
- 6.Christofi FL, Wood JD. Effects of PACAP on morphologically identified myenteric neurons in guinea pig small bowel. Am J Physiol. 1993;264:G414–G421. doi: 10.1152/ajpgi.1993.264.3.G414. [DOI] [PubMed] [Google Scholar]
- 7.Christofi FL, Baidan LV, Wood JD, Guan Z, Stokes BT. Multiple intracellular signaling mechanisms are linked to elevation of cytosolic calcium in myenteric multipolar neurons of the guinea-pig small bowel. Gastroenterology. 1995;108:A957. [Google Scholar]
- 8.Clarke DE, Craig DA, Fozard JR. The 5-HT4 receptor: naughty but nice. Trends Pharmacol Sci. 1989;10:385–386. doi: 10.1016/0165-6147(89)90177-6. [DOI] [PubMed] [Google Scholar]
- 9.Cohen ML, Schenck KW, Colbert W, Wittenauer L. Role of 5-HT2 receptors in serotonin-induced contractions of nonvascular smooth muscle. J Pharmacol Exp Ther. 1985;232:770–774. [PubMed] [Google Scholar]
- 10.Craig DA, Clarke DE. Pharmacological characterization of a neuronal receptor for 5-hydroxytryptamine in guinea pig ileum with properties similar to the 5-hydroxytryptamine4 receptor. J Pharmacol Exp Ther. 1990;252:1378–1386. [PubMed] [Google Scholar]
- 11.Derkach V, Surprenant A, North RA. 5-HT3 receptors are membrane ion channels. Nature. 1989;339:706–709. doi: 10.1038/339706a0. [DOI] [PubMed] [Google Scholar]
- 12.Engel G, Hoyer D, Kalkman HO, Wick MB. Identification of 5-HT2 receptors on longitudinal muscle of the guinea pig ileum. J Recept Res. 1984;4:113–126. doi: 10.3109/10799898409042543. [DOI] [PubMed] [Google Scholar]
- 13.Exton JH. Phosphoinositide phospholipases and G-proteins in hormone action. Annu Rev Physiol. 1994;56:349–369. doi: 10.1146/annurev.ph.56.030194.002025. [DOI] [PubMed] [Google Scholar]
- 14.Fiorica-Howells E, Wade PR, Gershon MD. Serotonin-induced increase in cAMP in ganglia isolated from the myenteric plexus of the guinea pig small intestine: mediation by a novel 5-HT receptor. Synapse. 1993;13:333–349. doi: 10.1002/syn.890130406. [DOI] [PubMed] [Google Scholar]
- 15.Foguet M, Hoyer D, Pardo LA, Parekh A, Kluxen FW, Kalkman HO, Stümer W, Lübbert H. Cloning and functional characterization of the rat stomach fundus serotonin receptor. EMBO J. 1992a;11:3481–3487. doi: 10.1002/j.1460-2075.1992.tb05427.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Foguet M, Nguyen H, Le H, Lübbert H. Structure of the mouse fundus serotonin receptor genes. NeuroReport. 1992b;3:345–348. doi: 10.1097/00001756-199204000-00014. [DOI] [PubMed] [Google Scholar]
- 17.Friedman E, Wang HY. The effect of age on brain cortical protein kinase C and its mediation of serotonin release. J Neurochem. 1989;52:187–192. doi: 10.1111/j.1471-4159.1989.tb10915.x. [DOI] [PubMed] [Google Scholar]
- 18.Furness JB, Trussell DC, Pompolo S, Bornstein JC, Smith TK. Calbindin neurons of the guinea-pig small intestine: quantitative analysis of their numbers and projections. Cell Tissue Res. 1990;260:261–272. doi: 10.1007/BF00318629. [DOI] [PubMed] [Google Scholar]
- 19.Galligan JJ. Electrophysiological studies of 5-hydroxytryptamine receptors on enteric neurons. In: Gaginella TS, Galligan JJ, editors. Serotonin and gastrointestinal function. CRC; Boca Raton, FL: 1995. pp. 109–126. [DOI] [PubMed] [Google Scholar]
- 20.Galligan JJ, North RA. Opioid, 5-HT1A and a2 receptors localized to subsets of guinea-pig myenteric neurons. J Auton Nerv Sys. 1991;32:1–12. doi: 10.1016/0165-1838(91)90229-v. [DOI] [PubMed] [Google Scholar]
- 21.Galligan JJ, Surprenant A, Tonini M, North RA. Differential localization of 5-HT1 receptors on myenteric and submucosal neurons. Am J Physiol. 1988;255:G603–G611. doi: 10.1152/ajpgi.1988.255.5.G603. [DOI] [PubMed] [Google Scholar]
- 22.Gershon MD. Localization and neurochemical aspects of serotonin in the gut. In: Gaginella TS, Galligan JJ, editors. Serotonin and gastrointestinal function. CRC; Boca Raton, FL: 1995. pp. 11–31. [Google Scholar]
- 23.Gershon MD, Takaki M, Tamir H, Branchek T. The enteric neural receptor for 5-hydroxtryptamine. Experentia. 1985;41:863–868. doi: 10.1007/BF01970002. [DOI] [PubMed] [Google Scholar]
- 24.Gershon MD, Wade PR, Fiorica-Howells E, Kirchgessner AL, Tamir H. 5-HT1P receptors in the bowel: G protein coupling, localization, and function. In: Fozard JR, Saxena PR, editors. Serotonin: molecular biology, receptors, and functional effects. Birkhäuser; Basel: 1991. pp. 133–143. [Google Scholar]
- 25.Gershon MD, Kirchgessner AL, Wade PR. Functional anatomy of the enteric nervous system. In: Johnson LR, Alpers DH, Jacobson ED, Walsh JH, editors. Physiology of the gastrointestinal tract, 3rd Ed, Pt 1. Raven; New York: 1994. pp. 381–422. [Google Scholar]
- 26.Grafe P, Mayer CJ, Wood JD. Synaptic modulation of calcium-dependent potassium conductance in myenteric neurones in the guinea-pig. J Physiol (Lond) 1980;305:235–248. doi: 10.1113/jphysiol.1980.sp013360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hirst GDS, Johnson SM, Van Helden DF. The slow calcium-dependent potassium current in a myenteric neurone of the guinea-pig ileum. J Physiol (Lond) 1985;361:315–337. doi: 10.1113/jphysiol.1985.sp015648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Humphrey PPA, Hartig P, Hoyer D. A proposed new nomenclature for 5-HT receptors. Trends Pharmacol Sci. 1993;14:233–236. doi: 10.1016/0165-6147(93)90016-d. [DOI] [PubMed] [Google Scholar]
- 29.Jacobowitz O, Chen J, Premont RT, Iyengar R. Stimulation of specific types of Gs-stimulated adenyl clclases by phorbol ester treatment. J Biol Chem. 1993;268:3829–3832. [PubMed] [Google Scholar]
- 30.Jiang X, Naik MU, Hrabe J, Sacktor TC. Developmental expression of the protein kinase C family in rat hippocampus. Brain Res Dev Brain Res. 1994;78:291–295. doi: 10.1016/0165-3806(94)90038-8. [DOI] [PubMed] [Google Scholar]
- 31.Johnson SM, Katayama Y, North RA. Multiple actions of 5-hydroxytryptamine on myenteric neurons of the guinea-pig ileum. J Physiol (Lond) 1980;304:459–479. doi: 10.1113/jphysiol.1980.sp013336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kemp BE, Faux MC, Means AR, House C, Tiganis T, Hu S-H, Michelhill KI. Structural aspects: pseudosubstrate and substrate interactions. In: Woodgett JR, editor. Protein kinases. Oxford UP; Oxford: 1994. [Google Scholar]
- 33.Kirchgessner AL, Tamir H, Gershon MD. Identification and stimulation by serotonin of intrinsic sensory neurons of the submucosal plexus of the guinea pig gut: activity-induced expression of Fos immunoreactivity. J Neurosci. 1992;12:235–249. doi: 10.1523/JNEUROSCI.12-01-00235.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kirchgessner AL, Liu M-T, Gershon MD. In situ identification and visualization of neurons that mediate enteric and enteropancreatic reflexes. J Comp Neurol. 1996;371:270–286. doi: 10.1002/(SICI)1096-9861(19960722)371:2<270::AID-CNE7>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 35.Kursar JD, Nelson DJ, Wainscott DB, Cohen ML, Baez M. Molecular cloning, functional expression, and pharmacological characterization of a novel serotonin receptor (5-hydroxytryptamine2F) from rat stomach fundus. Mol Pharmacol. 1992;42:549–557. [PubMed] [Google Scholar]
- 36.Lowry OH, Rosenbrough NJ, Fair AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 37.Mage MG, Lamoyi E. Preparation of Fab and F(ab′)2 fragments from monoclonal antibodies. Immunol Ser. 1987;33:79–97. [Google Scholar]
- 38.Martiny-Baron G, Kazanietz MG, Mischak H, Blumberg PM, Kochs G, Hug H, Marmé D, Schächtele C. Selective inhibition of protein kinase C isozymes by the indolcarbazole Gö 6976. J Biol Chem. 1993;268:9194–9197. [PubMed] [Google Scholar]
- 39.Mawe GM, Gershon MD. The 5-HT1P receptor. Drug News Perspect. 1993;6:24–30. [Google Scholar]
- 40.Mawe GM, Branchek T, Gershon MD. Peripheral neural serotonin receptors: identification and characterization with specific agonists and antagonists. Proc Natl Acad Sci USA. 1986;83:9799–9803. doi: 10.1073/pnas.83.24.9799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mawe GM, Branchek T, Gershon MD. Blockade of 5-HT-mediated enteric slow EPSPs by BRL 24924: gastrokinetic effects. Am J Physiol. 1989;257:G386–G396. doi: 10.1152/ajpgi.1989.257.3.G386. [DOI] [PubMed] [Google Scholar]
- 42.Morita K, North RA, Tokimasa T. The calcium-activated potassium conductance in guinea-pig myenteric neurones. J Physiol (Lond) 1982;333:141–149. doi: 10.1113/jphysiol.1982.sp014306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nemeth PR, Palmer JM, Wood JD, Zafirov DH. Effects of forskolin on electrical behaviour of myenteric neurones in guinea-pig small intestine. J Physiol (Lond) 1986;376:439–450. doi: 10.1113/jphysiol.1986.sp016162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nishizuka Y. Intracellular signaling by hydrolysis of phospholipids and activation of protein kinase C. Science. 1992;258:607–614. doi: 10.1126/science.1411571. [DOI] [PubMed] [Google Scholar]
- 45.North RA, Tokimasa T. Persistent calcium-sensitive potassium current and the resting properties of guinea pig myenteric neurones. J Physiol (Lond) 1987;386:333–353. doi: 10.1113/jphysiol.1987.sp016537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Palmer JM, Wood JD, Zafirov DH. Elevation of cyclic adenosine 3′,5′-phosphate mimics slow synaptic excitation in myenteric neurones. J Physiol (Lond) 1986;376:451–460. doi: 10.1113/jphysiol.1986.sp016163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Palmer JM, Wood JD, Zafirov DH. Purinergic inhibition in the small intestinal myenteric plexus of the guinea pig. J Physiol (Lond) 1987a;387:357–369. doi: 10.1113/jphysiol.1987.sp016577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Palmer JM, Wood JM, Zafirov DH. Transduction of aminergic and peptidergic signals in enteric neurones of the guinea pig. J Physiol (Lond) 1987b;387:371–383. doi: 10.1113/jphysiol.1987.sp016578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pan H, Galligan JJ. 5-HT1A and 5-HT4 receptors mediate inhibition and facilitation of fast synaptic transmission in enteric neurons. Am J Physiol. 1994;266:G230–G238. doi: 10.1152/ajpgi.1994.266.2.G230. [DOI] [PubMed] [Google Scholar]
- 50.Pieroni JP, Jacobowitz O, Chen J, Iyengar R. Signal recognition and integration by Gs-stimulated adenyl cyclases. Curr Opin Neurobiol. 1993;3:345–351. doi: 10.1016/0959-4388(93)90127-k. [DOI] [PubMed] [Google Scholar]
- 51.Pompolo S, Furness JB. Ultrastructure and synaptic relationships of calbindin-reactive, Dogiel type II neurons, in the myenteric ganglia of guinea-pig small intestine. J Neurocytol. 1988;17:771–782. doi: 10.1007/BF01216705. [DOI] [PubMed] [Google Scholar]
- 52.Schütze S, Potthoff K, Machleidt T, Berkovic D, Wiegmann K, Krönke M. TNF activates NF-kB by phosphatidylcholine-specific sphingomyelin breakdown. Cell. 1992;71:765–776. doi: 10.1016/0092-8674(92)90553-o. [DOI] [PubMed] [Google Scholar]
- 53.Takaki M, Branchek T, Tamir H, Gershon MD. Specific antagonism of enteric neural serotonin receptors by dipeptides of 5-hydroxytryptophan: evidence that serotonin is a mediatory of slow synaptic excitation in the myenteric plexus. J Neurosci. 1985a;5:1769–1780. doi: 10.1523/JNEUROSCI.05-07-01769.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Takaki M, Mawe GM, Barasch J, Gershon MD. Physiological responses of guinea-pig myenteric neurons secondary to the release of endogenous serotonin by tryptamine. Neuroscience. 1985b;16:223–240. doi: 10.1016/0306-4522(85)90059-4. [DOI] [PubMed] [Google Scholar]
- 55.Tanaka C, Nishizuka Y. The protein kinase C family for neuronal signaling. Annu Rev Neurosci. 1994;17:551–567. doi: 10.1146/annurev.ne.17.030194.003003. [DOI] [PubMed] [Google Scholar]
- 56.Wade PR, Branchek TA, Mawe GM, Gershon MD. Use of stereoisomers of zacopride to analyze actions of 5-hydroxytryptamine on enteric neurons. Am J Physiol. 1991;260:G80–G90. doi: 10.1152/ajpgi.1991.260.1.G80. [DOI] [PubMed] [Google Scholar]
- 57.Wade PR, Tamir H, Kirchgessner AL, Gershon MD. Analysis of the role of 5-HT in the enteric nervous system using anti-idiotypic antibodies to 5-HT receptors. Am J Physiol. 1994;266:G403–G416. doi: 10.1152/ajpgi.1994.266.3.G403. [DOI] [PubMed] [Google Scholar]
- 58.Wade PR, Chen J, Jaffe B, Kassem IS, Blakely RD, Gershon MD. Localization and function of a 5-HT transporter in crypt epithelia of the gastrointestinal tract. J Neurosci. 1996;16:2352–2364. doi: 10.1523/JNEUROSCI.16-07-02352.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang H-Y, Fiorica-Howells E, Pan H, Gershon MD, Friedman E. Myenteric ganglionic 5-hydroxytryptamine1P signal transmission is mediated via Go protein. J Pharmacol Exp Ther. 1996;277:518–524. [PubMed] [Google Scholar]
- 60.Wood JD. Electrical and synaptic behavior of enteric neurons. In: Wood JD, editor. Handbook of physiology. The gastrointestinal system motility and circulation, Sec 6, Vol 1, Pt 1. American Physiological Society; Bethesda, MD: 1989. pp. 465–517. [Google Scholar]
- 61.Wood JD. Physiology of the enteric nervous system. In: Johnson LR, Alpers DH, Jacobson ED, Walsh JH, editors. Physiology of the gastrointestinal tract, 3rd Ed. Raven; New York: 1994. pp. 423–482. [Google Scholar]
- 62.Wood JD, Mayer CJ. Serotonergic activation of tonic-type enteric neurons in guinea pig small bowel. J Neurophysiol. 1979;422:582–593. doi: 10.1152/jn.1979.42.2.582. [DOI] [PubMed] [Google Scholar]
- 63.Xia Y, Baidan LV, Fertel RH, Wood JD. Determination of levels of cAMP in the myenteric plexus of guinea-pig small intestine. Eur J Pharmacol. 1991;206:231–236. doi: 10.1016/s0922-4106(05)80023-9. [DOI] [PubMed] [Google Scholar]
- 64.Xia Y, Fertel RH, Wood JD. Stimulation of formation of cAMP by 5-hydroxytryptamine in myenteric ganglia isolated from guinea pig small intestine. Life Sci. 1994;55:685–692. doi: 10.1016/0024-3205(94)00675-x. [DOI] [PubMed] [Google Scholar]
- 65.Yau WM, Dorsett JA, Parr EL. Characterization of acetylcholine release from enzyme-dissociated myenteric ganglia. Am J Physiol. 1989;19:G233–G239. doi: 10.1152/ajpgi.1989.256.1.G233. [DOI] [PubMed] [Google Scholar]
- 66.Yoshimura M, Cooper D, MF. Type specific stimulation of adenyl cyclase by protein kinase C. J Biol Chem. 1993;268:4604–4607. [PubMed] [Google Scholar]
- 67.Zafirov DH, Palmer JM, Nemeth PR, Wood JD. Cyclic 3′,5′-adenosine monophosphate mimics slow synaptic excitation in myenteric plexus neurons. Brain Res. 1985a;347:368–371. doi: 10.1016/0006-8993(85)90201-x. [DOI] [PubMed] [Google Scholar]
- 68.Zafirov DH, Palmer JM, Wood JD. Adenosine inhibits forskolin-induced excitation in myenteric neurons. Eur J Pharmacol. 1985b;113:143–144. doi: 10.1016/0014-2999(85)90358-9. [DOI] [PubMed] [Google Scholar]