

Abstract
The activity of 378 single neurons was recorded from areas of the parahippocampal region (PHR), including the perirhinal and lateral entorhinal cortex, as well as the subiculum, in rats performing an odor-guided delayed nonmatching-to-sample task. Nearly every neuron fired in association with some trial event, and every identifiable trial event or behavior was encoded by neuronal activity in the PHR. The greatest proportion of cells was active during odor sampling, and for many cells, activity during this period was odor selective. In addition, odor memory coding was reflected in two general ways. First, a substantial proportion of cells showed odor-selective activity throughout or at the end of the memory delay period. Second, odor-responsive cells showed odor-selective enhancement or suppression of activity during stimulus repetition in the recognition phase of the task. These data, combined with evidence that the PHR is critical for maintaining odor memories in animals performing the same task, indicate that this cortical region mediates the encoding of specific memory cues, maintains stimulus representations, and supports specific match–nonmatch judgments critical to recognition memory. By contrast, hippocampal neurons do not demonstrate evoked or maintained stimulus-specific codings, and hippocampal damage results in little if any decrement in performance on this task. Thus it becomes increasingly clear that the parahippocampal cortex can support recognition memory independent of the distinct memory functions of the hippocampus itself.
Keywords: entorhinal cortex, perirhinal cortex, subiculum, hippocampus, recognition memory, delayed nonmatching, single units
The parahippocampal region (PHR) has become a focus for anatomical, behavioral, and electrophysiological studies of the medial temporal lobe memory system (Eichenbaum et al., 1994; Brown, 1996; Murray, 1996; Suzuki, 1996). This cortical area surrounds the hippocampus and amygdala and is composed of several distinct subdivisions, including the perirhinal, entorhinal, and parahippocampal (in monkeys) or postrhinal (in rats) cortices (Witter et al., 1989;Burwell et al., 1995). The PHR receives inputs from widespread secondary or “association” cortical regions and provides the major conduit for hippocampal outputs to the same cortical association areas. This anatomical evidence indicates that the PHR occupies a pivotal position for mediating memory functions of the hippocampal region.
Neuropsychological findings indicate that the PHR indeed plays a critical role in recognition memory, independent of its role as an intermediary for cortical–hippocampal interactions (Eichenbaum et al., 1994; Murray, 1996). This evidence comes mainly from experiments examining the effects of damage to the hippocampal region on performance in a simple recognition memory test known as delayed nonmatching to sample (DNMS) (Eichenbaum et al., 1994). In the standard version of this task, originally devised by Gaffan (1974), animals are presented with a novel “sample” cue and then after a variable memory delay must show that they recognize the sample by selecting against it (nonmatching) when presented with a choice between the now familiar stimulus and a new item. Several studies using different variants of DNMS in both rats and monkeys have shown that damage to the PHR results in a severe and selective deficit in performance at long delays (Zola-Morgan et al., 1989b, 1994; Gaffan and Murray, 1992; Otto and Eichenbaum, 1992a; Meunier et al., 1993; Suzuki et al., 1993;Eacott et al., 1994; Gaffan, 1994; Mumby and Pinel, 1994), whereas ablation of the hippocampus or transection of the fornix produce relatively little deficit (Bachevalier et al., 1985; Aggleton et al., 1986, 1989; Zola-Morgan et al., 1989a, 1994; Gaffan, 1974; Gaffan et al., 1984; Mumby et al., 1992) or no deficit (Rothblat and Kromer, 1991; Otto and Eichenbaum, 1992a; Jackson-Smith et al., 1993; Kesner et al., 1993; Gaffan, 1994; Murray, 1996; Murray and Mishkin, 1996).
To mediate performance in DNMS, a specific encoding of the sample cue must be maintained throughout the memory delay. The stored representation must then be compared with choice items presented in the recognition test. In a recent study examining the firing patterns of hippocampal neurons in rats performing an odor-guided DNMS task, we found no cells that encoded specific sample cues, maintained stimulus representations during the memory delay, or fired in association with match or nonmatch comparisons for particular sample and test odors (Otto and Eichenbaum, 1992b). These findings are consistent with the neuropsychological data indicating that the hippocampus itself is not essential for odor-guided DNMS performance (Otto and Eichenbaum, 1992a). In the present study we extended our recordings to determine whether firing patterns of PHR neurons would reflect these aspects of stimulus representation. Our studies focused on the perirhinal cortex and the lateral entorhinal cortex, two parts of the PHR that receive direct olfactory inputs (Deacon et al., 1983), and on the subiculum, another retrohippocampal cortical area that is reciprocally connected with the PHR.
MATERIALS AND METHODS
Subjects. Nine male Long–Evans rats, weighing between 340 and 550 gm at the beginning of the experiment, served as subjects. The animals were housed individually, maintained on a 12 hr light/dark cycle, and given ad libitum access to food. Water access was restricted to that earned during the performance of the continuous DNMS (cDNMS) task, and to 40 min of free access per day at the end of each test session.
Electrodes, surgery, and histology. The electrode assemblies consisted of 10 25-μm-diameter Formvar-coated nichrome wires of equal length bundled into a 27 gauge cannula (Eichenbaum et al., 1977) and attached to a vertically driveable connector (Kubie, 1984). The animals were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) supplemented with methoxyflurane when necessary; atropine was administered (0.5 mg in 1 cc, i.p.) to reduce mucous secretions. With use of aseptic surgical procedures, the electrode assemblies were implanted stereotaxically, with the skull level at the following coordinates: for lateral entorhinal cortex and subiculum, 5.2 mm posterior to and 5.0 mm lateral to bregma, and 6.5 mm below the pial surface with the electrode carrier oriented at 16° from vertical in the coronal plane; for perirhinal cortex, 5.2 mm posterior to and 5.0 mm lateral to bregma, and 4.5 mm below the pial surface with the electrode carrier oriented at 17.5° from vertical in the coronal plane. At the conclusion of testing, each subject was administered a lethal dose of sodium pentobarbital (100 mg/kg), a 15 μA current was passed through three of the recording electrodes, and the animal was then perfused transcardially with normal saline followed by 10% buffered formalin. The brains were removed from the skulls and stored in 10% buffered formalin for 24 hr and then transferred to a 3% potassium ferrocyanide solution for another 24 hr. This second solution produced a Prussian blue reaction that aided the localization of the electrode tips. Finally, the brains were transferred to a 30% sucrose–formalin solution for an additional 24–48 hr, coronal sections were cut at 30 μm on a sliding microtome, and the sections were mounted and stained with thionin.
Unit recording and computerized data acquisition. The subjects were screened once per day for unit activity. If no activity was identified during screening, the electrode was advanced ∼40 μm and allowed to settle for at least 24 hr before subsequent screening. Up to four channels of neural activity were passed individually through a high-impedance JFET headstage and then to AC amplifiers (Grass, model P511K), where the signals were amplified 5000× and bandpass-filtered at 300–3000 Hz. Each channel of neural activity was recorded separately on a multi-track digital tape recorder (Vetter, model PCM400) for subsequent off-line data acquisition. Additionally, computer-generated digital pulses that coded the behavioral events were also recorded. During the off-line data analysis sessions, unit isolation was achieved using a software template-matching algorithm (Spike2) provided with a computerized data acquisition system (Cambridge Electronic Design, model 1401+). With this system, up to eight units could be isolated from each channel of neural activity. Only units with signal-to-noise ratios of at least 3:1 were included in the analysis.
Behavioral apparatus. The behavioral apparatus consisted of a 30-cm-square aluminum chamber with one wall slanted at 5° outward from the floor. Odor stimuli were presented at a conical sniff port located on the center of the slanted wall, 5 cm above the floor. A photodetector mounted in the entrance to the sniff port monitored stimulus sampling responses. A water reward cup with its own photodetector was located 2.5 cm directly below the sniff port, and two 24 V panel lamps were located 10 cm to each side and 10 cm above the water cup. Odor cues were generated by a 16-channel flow-dilution olfactometer. The stimulus set was composed of 16 arbitrarily selected volatile odors that were easily discriminable to the experimenters. Half of the odors were common food scents (e.g., anise, almond, peppermint, lemon, celery seed, cinnamon, banana, clove) and the others were pure chemical odorants or extracts (e.g., geraniol, amyl acetate, phenethyl alcohol, eugenol, terpineol, guiacol, damascone, jasmopyrane), and each was diluted to 5% in propylene glycol. Initially a clean airstream was generated from pressurized air that was dehydrated with calcium chloride, cleaned with activated charcoal, and then rehydrated with distilled water. This airstream was then split such that half of it flowed continuously at a rate of 0.5 l/min, serving to clear the odor channels during the intertrial interval (ITI). During the last 2 sec of the ITI, the other half of the airstream (0.5 l/min) was saturated with a selected odor and then added to the clean airstream, resulting in a final stimulus flow rate of 1.0 l/min. Odorized and clean airstreams were passed through a three-way solenoid valve mounted immediately outside of, and connected to, the sniff port. During the entire ITI the airstream was diverted by this valve to a vacuum dump at 2 l/min. Because the vacuum flow rate was greater than that of the input airstream, odors in the sniff port were effectively eliminated during periods when no odor was being presented. When a subject initiated an odor presentation by breaking the sniff port photobeam at an appropriate time, the solenoid valve was activated, allowing the odorized airstream to reach the sniff port; deactivation of this valve occurred immediately whenever the subject withdrew its nose from the sniff port. Additionally, a fan mounted on the top of the chamber was used to continuously exhaust air, thereby ensuring that odors did not linger in the chamber between odor presentations. All procedural events were controlled and behavioral responses were recorded by a PC-compatible computer equipped with a 32-bit digital input/output board.
Behavioral procedures. Training on the cDNMS task proceeded in a series of three phases. First, rats were given a minimum of two 60-trial sessions of shaping. A nose poke of ∼250 msec (in the range of 220 msec to 280 msec) into the sniff port resulted in the presentation of an odor chosen on a pseudorandom basis from a set of either 8 or 16 odors selected at random from a large stock of odorants. A subsequent water port response terminated the odor presentation and was reinforced with a 0.05 ml water reward. During this phase, the odor presented on each trial always differed from that presented on the immediately preceding trial. A 3 sec delay was imposed between trials; the house lights were extinguished during the delay and subsequently reilluminated to signal the availability of the next trial. Nose pokes into the sniff port during the last 2 sec of the delay extended the delay by an additional 2 sec.
In the second phase, rats learned the cDNMS task in daily sessions of 60–200 trials, using the full set of 16 odors. On each trial the end of the ITI (memory delay) was signaled by illumination of the house light, and the rat could then perform a nose poke of ∼250 msec into the sniff port to initiate stimulus onset. On half of the trials, the odor was different from that presented on the previous trial (a nonmatch or S+ trial) and a response to the water port within 5 sec (“go” or R+) was rewarded. On other trials, the odor was the same as that presented on the previous trial (a match or S− trial), and water port responses on these trials were not reinforced; rats learned not to make the water port response (“no-go” or R−) on these trials. Errors of commission resulted in the immediate offset of the house lights. On any trial when no water port response was made within 5 sec of the odor onset, the odor and house lights were turned off simultaneously and the delay period began. Correct responses were followed by a 3 sec delay; incorrect responses were followed by a 7 sec delay. Incorrect responses on match trials were initially followed by one or two correction trials, that is, repetitions of the same match trial. Daily training was provided until the rats reached a criterion of 18 correct responses out of 20 consecutive trials. Once this criterion was met, correction trials were no longer provided.
In the third stage, daily cDNMS sessions continued while we searched for and recorded from cells. During these sessions the delay was held constant at 3 sec for five of the six rats from which perirhinal cells were recorded. In these rats, odor responses were evaluated for the full set of 16 odors. The remaining rats were presented with odors from the reduced set of eight odors and were tested with delays of both 3 and 30 sec. Each delay occurred pseudorandomly on half the trials of a cDNMS session. The actual delay between the odor offset of one trial and the odor onset of the next trial could be somewhat longer, depending on the latency of a rat to initiate a trial after the onset of the houselight signal. On each day, electrodes were surveyed for cellular activity while the animals performed the cDNMS task. If no cells were identified, the animals continued performing the task until 100 trials were completed to maintain performance. When the activity of at least one cell with a suitable signal-to-noise ratio was observed, the data recorder was started and a 300–500 trial cDNMS session was presented. After each session, the electrode was advanced 40 μm.
Data analysis. The analysis of neuronal firing patterns was performed in two stages. First, all cells were assessed for activity associated with specific trial events and behavioral acts that occurred in the same sequence on each trial. This event analysis focused on (1) the onset of the house lights signaling the beginning of a trial; (2) entry into the sniff port initiating the trial; (3) onset of the odor presentation; (4) removal of the nose from the sniff port, referred to henceforth as the “unpoke”; and (5) entry into the water port as the discriminative response to retrieve the reward. For each cell a set of graphic analyses was prepared to display unit activity associated with each of these trial events. Each analysis included a raster display of approximately 15 representative trials plus a summary histogram for all trials that displayed averaged peri-event activity accumulated in 100 msec bins for 2 sec before and after the event, plotted as a continuous line indicating average spikes/second for each bin. To compare the present results with the CA1 recordings obtained from rats performing the same task (Otto and Eichenbaum, 1992b), this analysis was used to identify three general functionally defined cell types: “cue-sampling cells,” whose activity changed maximally (either increased or decreased) while the subject was sampling an odor; “reward-approach cells,” whose activity changed during the approach to the water cup and rewar consumption; and “trial-initiation cells,” whose activity changed maximally just before or during the trial initiation. Because of our specific interest in neural activity during the delay, an additional category of cells was identified as “delay cells,” and we focused on firing during the middle of the shorter (3 sec) delay period. Cue-sampling cells were operationally defined as those neurons whose average change in activity was maximal during the period between odor onset and 500 msec after odor onset; “delay” cells were defined as those whose change in activity was maximal between 2 and 2.5 sec after the unpoke; reward-approach cells were defined as those whose change in activity was maximal 100 msec before to 400 msec after the water port entry; and trial-initiation cells were defined as those whose change in activity was maximal 250 msec before to 250 msec after the sniff port entry. The statistical significance of these changes was determined using paired t tests (two-tailed) that compared activity during the periods defined above with background activity defined as the average firing rate during the 1 sec period immediately preceding the onset of the house lights, a period that followed the defined “delay” period and preceded the “trial initiation” period. Thus this epoch is included within the overall memory delay, but it occurs near the end of that period when delay-related activity might be expected to be minimized, and so provides the best estimate of background firing rate between trials. For all cue-sampling and reward-approach units with significant (p < 0.05) changes in firing, an additional series of post hocanalyses was applied to test whether firing rates changed preferentially during trials with particular stimulus type (S+ or S−) and response type (R+ or R−) combinations as described below.
The second stage of analysis focused on the odor specificity of neural activity associated with stimulus coding, including that reflecting match–nonmatch comparisons, and during the memory delay. We first designated the stimulus period as the 500 msec interval immediately after the odor onset during which the rat was actively sampling the odor and the memory period as the 500 msec interval just before onset of the succeeding trial, that is, at the end of the effective memory delay. At this time the rat was reliably initiating the trial during which the previous odor would be compared with the subsequent odor. One-way ANOVAs were used to identify significant differences in the neuronal responses among the odor set during the sensory and memory periods. Additionally, to examine whether the activity during the memory period varied according to the length of delay, cells recorded in sessions in which both the 3 and 30 sec delays were used were subjected to a two-way factorial analysis with delay length and odor as the two factors. Newman–Keuls post hoccomparisons were used to test differences between pairs of means when necessary.
Additional analyses of differential match–nonmatch responses focused on cells that showed odor-selective activity during the stimulus period. Our aims were to determine whether odor-selective cells tended to show enhanced or suppressed responses to immediate stimulus repetition and whether the match–nonmatch response was larger for the odors to which the cell is tuned than for other odors. One analysis was focused on individual cells and used a two-way ANOVA (2 × 2) to compare responses on match versus nonmatch trials for the “best” versus “worst” odor, that is, for those odors associated with the highest and lowest mean firing during the cue-sampling period. A second analysis was focused on the entire population of cells that showed odor-selective activity during odor sampling. We first categorized activity as “enhanced” if the mean firing rate was higher on match than nonmatch trials or “suppressed” if the firing rate was lower on match than nonmatch trials. Then the difference between match and nonmatch firing rates was computed, and paired t tests were performed separately on the groups of enhanced and suppressed cells to compare the difference scores between the best versus the worst odor stimulus.
RESULTS
Electrode localizations
Reconstructions of the electrode tracks are provided in Figure1. Because of the angle of electrode penetrations, recordings in the perirhinal and entorhinal cortices were almost exclusively in superficial layers. Recordings in the perirhinal cortex were primarily within the dorsal bank of the rhinal sulcus, with some sites bordering on ectorhinal cortex and a few sites in the ventral bank of the rhinal sulcus. Recordings in the entorhinal area were restricted to the lateral entorhinal cortex. In the subiculum, recordings were within the ventral part of the subiculum near the CA1 border. Analysis of recording sites along the electrode penetrations did not reveal any systematic localization of any of the functionally characterized cell types described below; however, there was a distinct bias for neighboring cells to have similar functional correlates.
Fig. 1.

Reconstructions of the electrode tracts for each of the subjects. Thick lines indicate the extent of loci of analyzed cellular activity. Sections are identified from the Swanson (1992) atlas. ab, Angular bundle; alv, alveus; ECT, ectorhinal cortex; ENTl, lateral entorhinal cortex; m, molecular layer;PERI, perirhinal cortex; PIR, piriform cortex; rf, rhinal fissure; sp, pyramidal layer; sr, stratum radiatum; SUBv, ventral subiculum; TEv, ventral temporal association area; TR, postpiriform transition area.
Behavioral performance
Rats acquired the cDNMS task rapidly, with all animals reaching the performance criterion in <400 trials. Rats tested with the 16 odor set continued to perform accurately during the recording sessions, averaging 92.1% correct. Compared with the performance of rats using the 16 odor set, the performance of rats tested with the 8 odor set was significantly reduced at both delays, averaging 84.9% correct at the 3 sec delay (t(7) = 3.63; p < 0.01) and 70.3% at the 30 sec delay (t(7)= 8.89; p < 0.001). Furthermore, the performance of those rats tested with the 8 odor set was significantly higher at the 3 sec delay than at the 30 sec delay (t(6) = 4.26;p < 0.01). This pattern of decreased cDNMS performance associated with reduced odor set size (higher inter-item interference) and longer memory delays parallels the earlier findings of Otto and Eichenbaum (1992a).
Neuronal activity related to behavioral events
A total of 378 units were isolated from perirhinal cortex (n = 177), lateral entorhinal cortex (n= 128), and subiculum (n = 73). The units were recorded during the course of 56 recording sessions, averaging 6.2 sessions per animal. Table 1 summarizes the results from the analysis of neuronal activity associated with the four specific behavioral events listed above, segregating the different cell types according to whether the firing changes occurred primarily during the stimulus onset period or during the delay period. Consistent with the findings on hippocampal CA1 cells recorded from rats performing the same cDNMS task (Otto and Eichenbaum, 1992b), as well as that from odor discrimination tasks (Eichenbaum et al., 1987; Wiener et al., 1989), the activity of single neurons in these areas was correlated with each identifiable event occurring during cDNMS performance. Indeed, a large proportion of neurons from each of the regions recorded (entorhinal 91.4%, perirhinal 89.8%, subiculum 100%) displayed activity that could be statistically related to one of the four behavioral events. This behavior-related activity was in the form of an increase in some units, a decrease in others, and both an increase and decrease in still other units. Most perirhinal and entorhinal cells showed increased firing, whereas most subiculum cells showed decreases in firing. Examples of responses for each of the four categories of cells identified in the event analysis are displayed in Figure 2.
Table 1.
Percentage of cells (and n) with changes in activity associated with trial events
| Correlate | Perirhinal (n = 177) | Lateral entorhinal (n = 128) | Subiculum (n = 73) |
|---|---|---|---|
| Cue-sampling cells | 43.5 (77) | 43.0 (54) | 47.9 (35) |
| Delay cells | 2.8 (5) | 14.8 (19) | 1.4 (1) |
| Reward-approach cells | 11.3 (20) | 4.7 (6) | 5.5 (4) |
| Trial-initiation cells | 32.2 (57) | 28.9 (37) | 45.2 (33) |
| No correlate | 10.2 (18) | 9.4 (12) | 0.0 (0) |
Fig. 2.

Examples of parahippocampal neurons with activity related to different trial events. For the present figure and all subsequent figures of this format, each panel includes a raster display of representative trials and a summary histogram of peri-event activity accumulated across all trials in 100 msec bins and displayed in a continuous line showing average spikes/second for each bin during the 2 sec period before and after the synchronization event. Then shown on each panel refers to the number of cDNMS trials over which unit activity was averaged. A, A perirhinal cue-sampling cell that began firing during trial initiation and fired maximally during odor sampling. Vertical tic marks to the right of the synchronization point indicate the unpoke. B, A perirhinal reward-approach cell that fired maximally just before the water port response.Vertical tic marks to the left of the synchronization point indicate the unpoke. C, A subiculum trial-initiation cell whose firing decreased during trial initiation (the poke). D, An entorhinal delay cell that fired maximally during the intertrial (delay) interval.
Cue-sampling cells
Neuronal activity during cDNMS performance strongly reflected the cue-sampling event (Table 1). Peak changes in firing during the cue-sampling phase varied among units, with perirhinal units often showing a striking synchronization to odor onset, offset, or both of these events. Figure 3 shows a perirhinal unit that exhibited a suppression of activity after the nose poke, a subsequent marked activity increase sharply time-locked to the odor onset, and finally a return to its basal firing rate synchronized with the cessation of odor sampling (the unpoke). Much less evidence of this stimulus synchronization was observed in lateral entorhinal and subiculum units.
Fig. 3.

Example of a perirhinal cue-sampling cell that showed a rapid increase in activity after the odor onset (A) and then a decrease in activity that was less sharply time-locked to the unpoke (B).Vertical tic marks to the left andright of the synchronization point indicate the occurrence of a poke and an unpoke respectively (A); tic marks to theleft of the synchronization point indicate the occurrence of the odor onset (B).
During the cue-sampling phase of each cDNMS trial, subjects must not only store information about the current trial but also must determine whether the current cue is a “match” or a “nonmatch” for the previous sample. To assess whether the activity of the cue-sampling units identified in the preliminary event analysis reflected match–nonmatch processing across all odor comparisons, three additional statistical analyses were conducted. First, cellular activity was compared for all nonmatch (S+) versus all match trials (S−) using a two-tailed t-test. To the extent that performance was accurate, however, differences in firing on nonmatch trials versus match trials could be attributable to the match–nonmatch distinction or the difference in the associated response (R+ vs R−). To decide between these two alternatives, additional post hoc t tests were performed comparing unit activity between match and nonmatch trials that ended in the same behavioral response, that is, S+R+ versus S−R+ trials, and between match and nonmatch trials that ended in different behavioral responses, that is, S+R+ versus S−R− trials. Cells whose activity was statistically different between match and nonmatch trials and in both post hoc tests were designated “strong” match–nonmatch cells, and cells who only differed in S+ versus S− and S+R+ versus S−R+ comparisons were designated “weak” match–nonmatch cells, a distinction made in our previous analysis of hippocampal neurons (Otto and Eichenbaum, 1992b). Unfortunately, because of accurate task performance, there were very few S−R+ trials for some cells (<2% of the total trials in the most extreme case). Correspondingly, these analyses were restricted to sessions in which performance was poorest, that is, where substantial numbers of errors of commission were made. Thus, the complete set of comparisons could not be performed for the 61 cue-sampling units recorded from the perirhinal cortex of rats tested with the 16 odor set, or for 11 of the cue-sampling units recorded from the lateral entorhinal area of rats tested with the 8 odor set. Of the remaining cue-sampling cells, 7 of the 43 units recorded from lateral entorhinal cortex and 6 of the 35 units recorded from the subiculum were designated as “strong” or “weak” match–nonmatch cells (Table2). An example of a “strong” match–nonmatch cell is shown in Figure 4. This entorhinal unit showed a marked increase in activity time-locked to the unpoke on S+R+ (Fig. 4A) trials, but showed no appreciable increase in activity on S−R+ (Fig. 4B) or S−R− (Fig. 4C) trials. Match–nonmatch cells were about equally divided in their preference for either match or nonmatch trials. Additional analyses aimed to identify neural activity associated with match–nonmatch comparisons for specific sample odors are provided below.
Table 2.
Match–nonmatch cells
| Correlate | Perirhinal | Lateral entorhinal | Subiculum |
|---|---|---|---|
| Cue-sampling cells | (n = 16) | (n = 43) | (n = 35) |
| Strong match–nonmatch | – | 4 | 3 |
| Weak match–nonmatch | – | 3 | 3 |
| Response | 2 | 10 | 4 |
| Nonspecific | 14 | 26 | 25 |
| Reward-approach cells | (n = 4) | (n = 5) | (n = 4) |
| Match–nonmatch | – | 3 | – |
| Nonspecific | 4 | 2 | 4 |
Fig. 4.
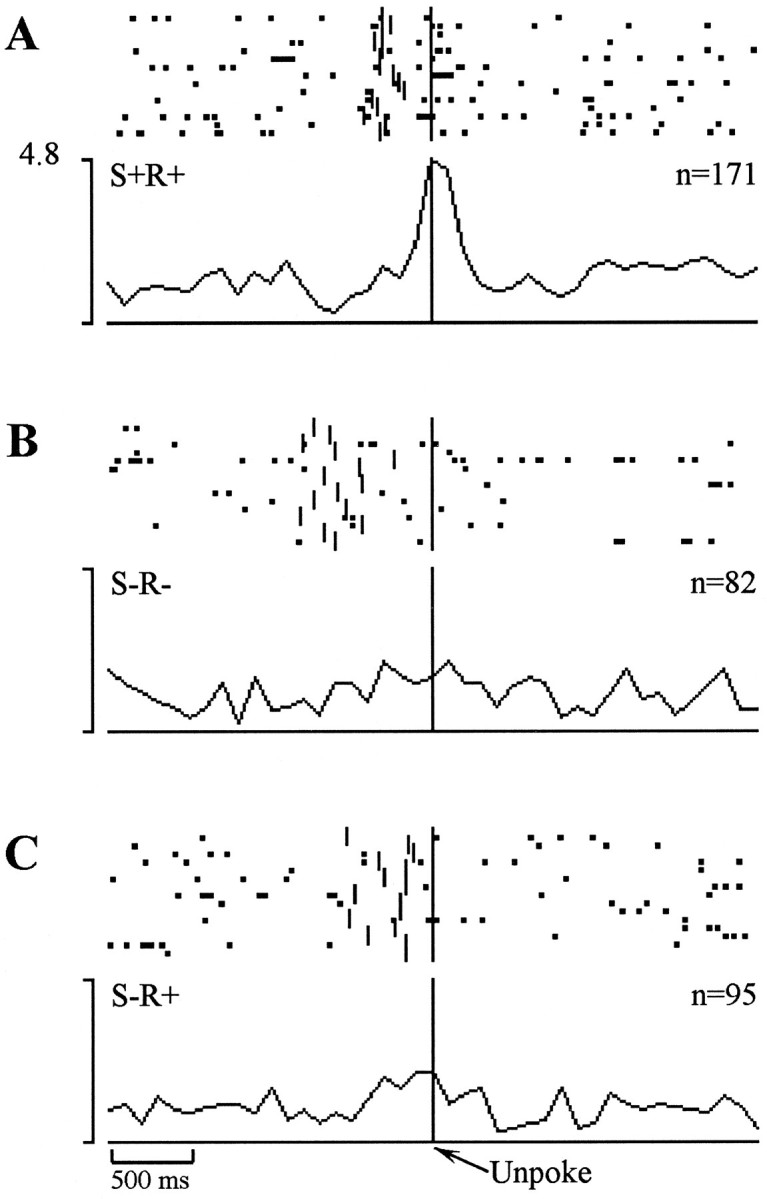
Example of a “strong nonmatch” entorhinal cell whose firing was closely synchronized to the unpoke on correct nonmatch trials (A) but showed no clear increase in firing during this period on correct match trials (B) or on errors of commission (C).Vertical tic marks to the left of the synchronization point indicate the occurrence of a sniff port poke.
Other units identified in the present analysis reflected the response (i.e., R+ or R−) that follows the test odor delivery better than the match–nonmatch comparison between two successive odors. For such cells the analyses indicated no significant difference in firing rate on S+R+ versus S−R+ trials, but differences in firing on S+R+ versus S−R− trials. These firing characteristics were identified in 16 units across all the PHR areas (Table 2). Both of the “response” cells in perirhinal cortex displayed increased activity on R+ compared with R− trials, whereas three lateral entorhinal cells and two subiculum cells preferred R+, and seven lateral entorhinal cells and two subiculum cells preferred R− trials.
Reward-approach cells
The increased firing of reward-approach cells immediately before and during the water response could reflect either the water response itself or some form of feedback associated with the outcome of the response (reinforcement or nonreinforcement). Changes in unit activity that occurred during this period on both S+R+ and S−R+ trials, or on S+R+ trials only, can be accounted for by either alternative. In contrast, if the change in activity occurred only during S−R+ trials, the firing cannot simply be associated with the water response per se, but must in some way reflect the outcome of the response. As in the analysis of the cue-sampling match–nonmatch cells, some units identified as reward-approach units were excluded from this analysis because of insufficient S−R+ trials. Of the 13 reward-approach cells that could be analyzed (Table 2), some in lateral entorhinal cortex fired significantly more on match trials, and the others exhibited no preference for either trial type. Figure 5 shows an example of a “match” reward-approach cell recorded from lateral entorhinal cortex. On both S+R+ (Fig. 5A) and S−R+ (Fig.5B) trials, firing in this cell increased ∼300 msec before the rat made the water port response, but reached a greater peak on the S−R+ trials. Note that in this example the differential increase was apparent before the water delivery, suggesting that the difference between the trial types was encoded before information (i.e., the absence of a water reward), indicating an error of commission.
Fig. 5.

Example of a “match” reward-approach cell recorded from entorhinal cortex. This cell displayed an increase in firing that was closely time-locked to the water port response but was significantly less active on correct nonmatch trials (A) than during errors of commission (B).
Trial-initiation cells
Substantial proportions of the units from all three recording sites were classified as trial initiation in the preliminary analysis (Table 1). The activity of these units might have reflected the incipient nose-poke behavior associated with the act of trial initiation or the maintenance or regeneration of an odor memory representation of the preceding sample odor. This issue will be addressed directly below in the analysis of stimulus-specific activity during the trial initiation period.
Delay cells
Units identified as delay cells in the initial event analysis (altered activity 2.0–2.5 sec after the unpoke) were most frequently recorded in the lateral entorhinal cortex, with considerably smaller proportions of this cell type in the perirhinal cortex and subiculum (Table 1). The firing pattern of these cells usually took the form of an increase in firing rate, beginning soon after the trial offset (house lights turned off), persisting for ∼1–2 sec, and then returning to baseline before the initiation of the subsequent trial. Closer examination of these cells revealed that responses usually occurred on S+R+ (i.e., rewarded) trials. Given that the trial offset occurred almost immediately after the water port response, it may be that this “delay-related” activity reflects the consummatory response rather than sensory information about the preceding sample cue. An example is presented in Figure 6. This cell displayed a marked increase in activity that occurred around the time of the trial offset on S+R+ trials, but no such increase occurred on either S−R− or S−R+ trials.
Fig. 6.
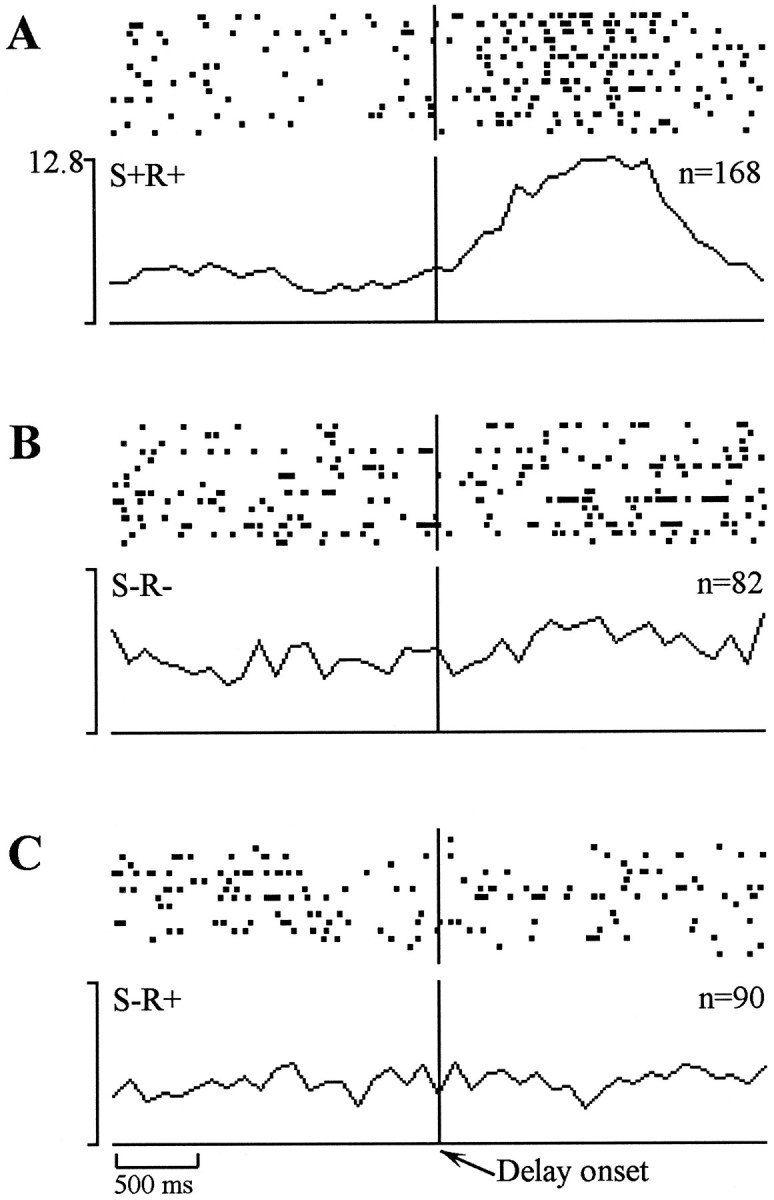
Example of an entorhinal delay cell that displayed increased firing immediately after the trial offset on correct match trials (A), but no such increase on correct nonmatch trials (B) or on errors of commission (C).
Odor-specific neuronal activity during stimulus and memory periods
Stimulus period
All units, rather than only those units classified as cue-sampling cells in event analysis, were included in the assessment of odor-selective activity during odor sampling. This inclusive approach was taken because of the possibility that a response that was highly odor-selective might be greatly diluted by averaging across a large number of odors, as was done in the event analysis, resulting in an overall nonsignificant firing rate change.
One-way ANOVAs on mean activity during the first 0.5 sec of the stimulus sampling period revealed that more than one third (45 of 128 units; 35.2%) of lateral entorhinal neurons had significant odor-selective responses, as did somewhat lesser proportions of perirhinal (20 of 177 units; 11.3%) and subicular (18 of 73 units; 24.7%) units. Most cells showed complex patterns of differential odor responses involving varying degrees of activation across the odor set; however, in some cells, activity during the sensory period was significantly greater for one particular odor. Examples of highly selective activation in perirhinal, lateral entorhinal, and subiculum cells are shown in Figure 7. Post hoccomparisons showed that the perirhinal cell (Fig. 7A) had a significantly greater response to odor 14 than to any of the other 15 sample odors. Similarly, the lateral entorhinal cell (Fig.7B) fired more selectively to odor 6, and the subiculum cell (Fig. 7C) to odor 2. To provide a more detailed representation of the time course of neural activity in an odor-selective cell, Figure 8 shows the firing pattern of the lateral entorhinal unit displayed in the previous figure across time. This unit showed a clear increase in selective activity during presentation of odor 6, reaching its peak at ∼200 msec after odor onset, and returning to baseline after termination of the odor presentation (typically 400–700 msec after odor onset).
Fig. 7.

Examples of the odor-selective activity during 500 msec periods of stimulus sampling (see time line at top of figure) in neurons recorded from (A) perirhinal cortex (F(15,209) = 3.93; p < 0.01), (B) entorhinal cortex (F(7,216) = 14.04; p < 0.01), and (C) the subiculum (F(7,323) = 23.22;p < 0.01). All three neurons showed activity that was significantly elevated during the sampling of only one odor of the 8 or 16 odor set.
Fig. 8.

Activity of the lateral entorhinal cell from Figure 3B during the 2 sec before and after the odor onset for each of the eight odors with which the subject was tested. The firing of this cell significantly increased during the cue-sampling period only for odor 6.
Additional analyses were performed on odor-selective cells to determine whether responses during immediate stimulus repetition (match trials) were enhanced or suppressed as compared with activity when a stimulus was not preceded by itself (nonmatch trials). These analyses included all cells that showed odor-selective activity but focused on comparisons of responses to the odor associated with the highest average firing rate during stimulus sampling (best odor) versus the odor associated with the lowest average firing rate (worst odor). In cell-by-cell analyses, 17 of the 83 odor-selective cells (3 of 20 perirhinal, 12 of 45 entorhinal, 2 of 18 subiculum) had significantly different activity on match versus nonmatch trials either as a main effect along with a main effect for odor, or in the interaction of odor × trial type, or both. All three perirhinal cells showed suppression of activity on match trials, but both subiculum cells showed match enhancement. In entorhinal cortex, seven of the cells showed suppression on match trials and five showed match enhancement. Of these cells, six showed both odor and trial-type effects, or a significant interaction between these factors, indicating odor-selective match suppression or enhancement by individual cells (Fig. 9).
Fig. 9.

Examples of odor-selective match suppression and match enhancement. Solid lines, Averages for match trials; dotted lines, averages for nonmatch trials.A, An entorhinal cell that showed a clear response only to the best odor on nonmatch trials and suppression of this response on match trials (selectivity for odor: F(1,92)= 10.55, p < 0.01; selectivity for nonmatch over match trials: F(1,92) = 20.86,p < 0.01). B, An entorhinal cell that showed a clear odor-selective response only on match trials (selectivity for odor: F(1,92) = 19.56,p < 0.01; selectivity for match over nonmatch trials: F(1,92) = 8.23,p < 0.01).
In yet a further analysis that took into consideration all odor-selective cells, we first separated those cells that had greater average firing on immediate stimulus repetition, that is, showed match enhancement, from those that had lower average firing on stimulus repetition, that is, showed match suppression. Then, in separate pairedt tests, the amount of enhancement or suppression was compared for the best versus worst odor stimulus for each cell. The results of this analysis (Fig. 10) indicated that an approximately equal numbers of cells showed match enhancement (n = 44) or suppression (n = 39). The amount of match-enhancement (t(43) = 3.22;p < 0.002) and match-suppression (t(38) = 3.81; p < 0.001) was substantially greater for the best than the worst odor. Although relatively few cells individually showed statistically significant odor-specific match–nonmatch effects, when the analyses on match and nonmatch responses were combined, the cell population demonstrated robust odor-selective match enhancement and match suppression.
Fig. 10.

Differences in firing on match minus nonmatch trials for all odor-selective cells categorized as enhanced or suppressed during stimulus repetition.
Memory period
Using the same rationale as in the analysis of odor specificity during cue sampling, all units were included in the analysis of odor-selective responses during the delay. ANOVAs revealed that 14 of the 177 perirhinal units (7.9%), 14 of the 128 lateral entorhinal units (10.9%), and 9 of the 73 subicular units (12.3%) exhibited odor-selective activity at the end of the memory delay. In the examples shown in Figure 11, post hoc comparisons showed that unit activity was significantly greater when one particular odor was presented on the previous trial. The perirhinal cell fired at a greater rate at the end of the delay after presentation of odor 15 than after any other odor, the entorhinal cell fired selectively at the end of the delay after odor 7, and the subiculum cell fired selectively after odor 6.
Fig. 11.
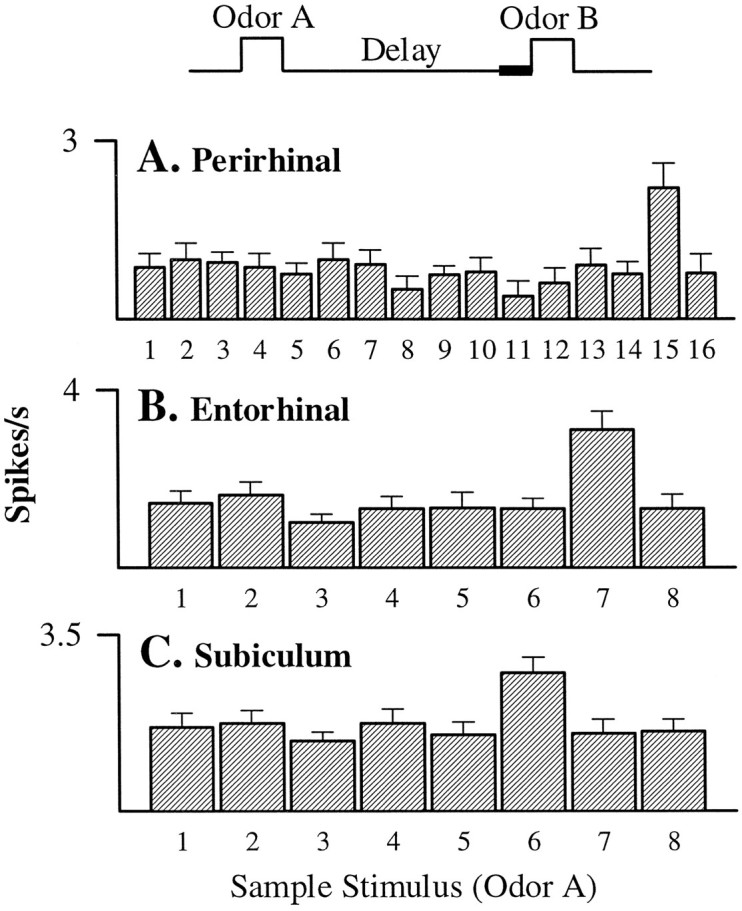
Examples of odor-selective activity during the final 500 msec of the memory delay (see time line) in neurons recorded from (A) perirhinal cortex (F(15,216) = 1.94; p < 0.05), (B) entorhinal cortex (F(7,317) = 4.57; p < 0.01), and (C) the subiculum (F(7,281) = 2.79; p< 0.01). All three neurons were maximally responsive during this period when one particular odor of the 8 or 16 odor set had been presented on the preceding trial.
To determine the duration over which odor memory representations can be maintained in the PHR, we evaluated odor specificity at the end of 3 sec delays as compared with that at the end of 30 sec delays. All units recorded from subjects tested with 3 and 30 sec delays were subjected to a two-way factorial ANOVA (odor × delay). Substantial fractions of neurons in each of the three areas displayed odor-selective responses, regardless of delay. Of the 128 lateral entorhinal cells analyzed, 10 (7.8%) exhibited odor-selective firing, as did 3 (10.0%) of the 30 perirhinal units and 8 (11.0%) of the 73 subicular units. Figure 12A shows a lateral entorhinal cell that maintained similar patterns of firing in the form of selective activation after presentation of odor 8 at the end of both 3 and 30 sec delays. Notably, we observed two distinct forms of neural activity across the delay yielding similar sensory period and subsequent memory period firing patterns. In some cells, odor-selective patterns were maintained above baseline throughout short (Fig. 13A) and in some cases long delays (not shown). In other cells, odor-selective patterns that appeared during cue sampling disappeared early in the delay but were reinstated just before the succeeding stimulus onset (Fig. 13B).
Fig. 12.

Examples of the three different categories of neurons obtained from the factorial analysis of odor and delay length.A, An entorhinal cell that displayed odor-selective activity at the end of the memory delay that was unaffected by the length of the delay (main effect of odor:F(7,221) = 9.34; p < 0.01). B, An entorhinal cell that was selectively active for one particular odor at the end of the 3 sec delay but not at the end of the 30 sec delay (main effect of odor:F(7,272) = 4.10, p < 0.01; odor × delay interaction:F(7,272) = 3.72, p < 0.01). C, A perirhinal cell that fired more at the end of 3 sec delays, but whose activity did not differ significantly across the odor set (main effect of delay: F(1,326)= 5.59; p < 0.05).
Fig. 13.

Examples of cells that showed odor-selective delay activity. A, An entorhinal cell that showed odor-selective activity above baseline throughout the short but not the long delay (odor × delay interaction:F(7,272) = 8.84; p < 0.01). B, An entorhinal cell that “recalled” the odor-selective sensory-evoked activity pattern at the end of the delay (main effect of odor: F(7,221) = 7.83;p < 0.01).
Fewer units recorded in lateral entorhinal cortex (6 of 128 units; 4.7%) and subiculum (3 of 73 units; 4.1%), and none of 30 units in the perirhinal cortex, exhibited an odor-specific change in activity that was also dependent on the length of the delay. Furthermore, examination of the magnitude of the odor-specific and delay length-dependent entorhinal and perirhinal cells revealed no clear tendency for cells to exhibit better specificity at either of the delays. Thus only three of the six entorhinal cells and one of the three subiculum cells that showed odor- and delay-selective activity displayed greater specificity at the 3 sec delay than at the 30 sec delay, as might be expected if these cells “forgot” the sample stimulus. Figure 12B shows an example of an entorhinal unit that displayed strong odor selectivity for odor 5 at the end of 3 sec delay but not at the end of the 30 sec delay.
Other cells in each area, however, fired differentially at the short versus long delays, with or without odor-selective responses. Substantial proportions of lateral entorhinal units (21 of 128 units; 16.4%) and perirhinal units (5 of 30 units; 16.7%) exhibited a significant delay effect, as did some subiculum units (3 of 73 units; 4.1%). Examination of the magnitude of these responses revealed that the firing of entorhinal units was not preferentially associated with either delay, with 11 of the 21 neurons displaying greater activity during the 3 sec delay. By contrast, all three subiculum units whose activity varied with the delay showed greater firing at the end of the 3 sec delay, and the activity of four of the five perirhinal units was greatest at the end of the 30 sec delay. An example of a perirhinal unit that fired significantly more at the end of 3 sec delays, but did not show statistically significant odor selectivity, is displayed in Figure 12C. When the data across these analyses are combined, the main finding is that firing patterns frequently vary with the length of the delay, but the pattern of differential odor-selective activity is largely maintained even across a long delay.
DISCUSSION
Behavioral correlates of neuronal activity in the PHR
Neuronal activity in the PHR reflected all identifiable behavioral events in the DNMS task. Many of the behavioral correlates likely reflect specific behaviors, e.g., odor investigation, reward consumption, and movements into or out of the sniff port. Other responses that were closely time-locked to stimulus onset and offset may reflect sensory-evoked responses. Yet other firing patterns that closely followed the match or nonmatch stimulus relations seem to reflect some general cognitive operation associated with all recognition judgments. More compelling evidence that the PHR is involved in stimulus-specific memory processing comes from comparisons of neural responses across the stimulus set described below.
The broad range of responses observed in the PHR is not surprising from the perspective of the anatomical inputs from widespread cortical regions, including all unimodal and polymodal sensory association areas, sensorimotor cortex, and multiple prefrontal and limbic cortical areas (Deacon et al., 1983; Burwell et al., 1995). The finding that nearly all cells reflected sensory, behavioral, and cognitive events within this task is consistent with the massive convergence of cortical inputs onto the PHR and the importance of this region to performance on the DNMS task.
Stimulus-specific “sensory” responses and “memory” correlates in the PHR of rats and monkeys
Sustained stimulus-selective neuronal activity has been observed in cells recorded from several cortical association areas, including the inferotemporal (Fuster and Jervey, 1981; Miyashita and Chang, 1988;Fuster, 1990) and prefrontal cortices (Goldman-Rakic et al., 1990) in monkeys and the auditory cortex in rats (Sakurai, 1990a). There have also been several reports of sensory- and memory-related activity in the PHR (Sakurai, 1990b; Miller et al., 1991, 1993; Riches et al., 1991; Quirk et al., 1992; Fahy et al., 1993; Li et al., 1993; Miller and Desimone, 1994; Zhu and Brown, 1995; Zhu et al., 1995). In particular, Brown and colleagues (Brown, 1996) and Miller and colleagues (Miller et al., 1991, 1993; Miller and Desimone, 1994) observed selective visually driven activity in the perirhinal and entorhinal areas of monkeys performing visually guided match and nonmatch to sample tasks. In those studies the predominant memory correlate was reduced activation on repetition of a sample cue, and this response was stimulus specific; however, Miller and colleagues (Miller et al., 1991, 1993; Miller and Desimone, 1994) also noted cells with sustained delay activity, which ended on immediate presentation of another cue, as well as enhanced activity when a match stimulus was repeated after intervening stimuli. Recently, Zhu and colleagues (Zhu and Brown, 1995; Zhu et al., 1995) also observed lasting stimulus-specific decremental sensory responses in the rat perirhinal and entorhinal cortex, and these responses were also sustained through intervening stimulus presentations.
In the present study, cells throughout the PHR of rats demonstrated sustained stimulus-selective activity during the memory delay. In some cases the odor-specific representation was maintained above baseline activity throughout the delay, and in other cases the activity pattern during odor sampling disappeared and then was “recalled” at the end of the delay. These observations suggest the existence of explicitly sustained sensory representations and subthreshold encodings that can be enhanced in anticipation of the matching event. In addition, equivalent proportions of parahippocampal neurons showed stimulus-selective match enhancement or match suppression of activity during a repeated stimulus. One possible explanation for both types of responses is that these firing patterns reflect the separate outcomes of equally frequent “match” and “nonmatch” judgments. Alternatively, the combination of enhancement and suppression could reflect competitive cellular interactions during reestablishment of a familiar stimulus representation. Regardless of the basis of these correlates, the capacity of parahippocampal neurons to identify specific match and nonmatch comparisons for a preceding cue indicates that this area contains sufficient information to support the recognition judgment.
Observations on the memory correlates of neuronal activity in the hippocampus compared with that in the PHR
The characteristics of PHR cells described above differ in important ways with observations on hippocampal neurons recorded in animals performing DNMS tasks. In the most directly comparable study,Otto and Eichenbaum (1992b) recorded the activity of CA1 neurons from rats performing the same odor-guided cDNMS task used here. Some aspects of PHR and hippocampal neuronal activity patterns are similar. In both the PHR and the hippocampus, cellular activity reflects virtually all identifiable trial events, including trial initiation, stimulus sampling, discriminative responses, and appetitive behaviors. This observation is consistent with the close and bidirectional connections between these areas and likely reflects their interactions as an interconnected system; however, the two areas differ strikingly in the extent to which neural activity reflects some of the critical aspects of stimulus coding relevant to recognition memory. In particular, unlike cells in the PHR, CA1 neurons exhibited none of the three characteristics identified above as important for the recognition memory demands of the cDNMS task. A subset of the CA1 cells was active during stimulus sampling, but these cells did not show odor-selective activity or persistent firing throughout the delay or stimulus-specific enhancement or suppression on stimulus repetition. Rather, the CA1 cells fired briefly during some part of the delay, perhaps reflecting ongoing behaviors that occurred at that time, and their activity reflected the abstract match–nonmatch relations between sample and choice stimuli. These findings are similar to results from other studies of hippocampal cellular activity in animals performing DNMS tasks. Thus, previous experiments that involved recording from hippocampal neurons in rats (Sakurai, 1990b) and monkeys (Brown et al., 1987; Riches et al., 1991) found no evidence of stimulus-specific encoding during the performance of DNMS tasks. Instead, similar to theOtto and Eichenbaum (1992b) findings, each of these studies described cellular activity related to the match and nonmatch judgments and related behavioral responses.
Two functional components of the hippocampal system
The present data, combined with the results from other recording and neuropsychological studies, indicate that the PHR contains the necessary coding elements for identifying individual stimuli, for maintaining individual stimulus representations across long delays, and for mediating specific match–nonmatch comparisons. By contrast, the hippocampus is not required for recognition of individual stimuli, and hippocampal neurons do not encode specific cues during recognition performance, nor do they show delay-related activity for specific memory cues. Studies using other tasks that require more elaborate memory processing, however, do indicate a role for the hippocampus, albeit one that may be qualitatively different from that of the PHR. Considerable evidence from studies on rats indicates the importance of the hippocampus in spatial memory processing (O’Keefe and Nadel, 1978; Nadel, 1991; Jarrard, 1993). These studies distinguish a critical role for the hippocampus whenever the animal must learn spatial relations among environmental stimuli and use the spatial layout of these cues to guide behavior. Correspondingly, hippocampal neuronal activity reflects the relevant spatial configuration of cues in animals exploring open fields (O’Keefe, 1976). Eichenbaum and colleagues (Eichenbaum et al., 1992; Cohen and Eichenbaum, 1993) have argued that the involvement of the hippocampus in learning stimulus organizations and using representations of relations among items should be extended to a broader scope of relational dimensions rather than only those of physical space. Indeed, recent evidence shows that selective damage to the hippocampus results in impairments in the flexible expression of learned odor organizations (Bunsey and Eichenbaum, 1996; Dusek and Eichenbaum, 1997). Correspondingly, hippocampal neuronal activity reflects a broad range of conjunctions or relations among cues that are relevant to performance in various tasks, even in DNMS tasks in which relational coding is not critical to task performance (Eichenbaum, 1996).
These neuropsychological and neurophysiological findings are consistent with our proposal (Cohen and Eichenbaum, 1993; Eichenbaum et al., 1994;Eichenbaum et al., 1995) that the hippocampal system can be divided into at least two functionally distinct components: a PHR component that supports persistent representations of individual items and a hippocampal component that mediates representations of relevant stimulus relationships. During the course of most memory performances, these functions of the PHR and hippocampus likely operate cooperatively and interactively. The PHR may store single items and episodes and maintain persistent representations that are then accessed by the hippocampus for interleaving into relational organizations that are then stored in the cortex (Alvarez and Squire, 1994; McClelland et al., 1995).
Finally, the present data speak to the issue of what specific areas should be included within these functional components of the hippocampal region. The current observations indicated similar firing patterns of cells in the perirhinal cortex, lateral entorhinal cortex, and subiculum, suggesting that all of these areas might be considered functionally related. Other studies, however, have indicated that the subiculum should be considered part of the hippocampus, both from an anatomical view and from evidence for common function (Eichenbaum et al., 1994). When combined with the present data, and consistent with its intermediate position between the entorhinal cortex and hippocampus, these findings show that the subiculum may contribute to both functional mechanisms.
Footnotes
This work was supported by Grant MH51570 from the National Institute of Mental Health. We thank Emma Wood and Pablo Alvarez for their comments on an earlier version of this manuscript.
Correspondence should be addressed to Dr. Howard Eichenbaum, Laboratory of Cognitive Neurobiology, Department of Psychology, Boston University, 64 Cummington Street, Boston, MA 02215. recognize the sample by selecting
REFERENCES
- 1.Aggleton J, Hunt PR, Rawlins JNP. The effects of hippocampal lesions upon spatial and nonspatial tests of working memory. Behav Brain Res. 1986;19:133–146. doi: 10.1016/0166-4328(86)90011-2. [DOI] [PubMed] [Google Scholar]
- 2.Aggleton J, Blint HS, Rawlins JNP. Effects of amygdaloid and amygdaloid-hippocampal lesions on object recognition and spatial working memory in rats. Behav Neurosci. 1989;103:962–974. doi: 10.1037//0735-7044.103.5.962. [DOI] [PubMed] [Google Scholar]
- 3.Alvarez P, Squire LR. Memory consolidation and the medial temporal lobe: a simple network model. Proc Natl Acad Sci USA. 1994;91:7041–7045. doi: 10.1073/pnas.91.15.7041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bachevalier J, Parkinson JK, Mishkin M. Visual recognition in monkeys: effects of separate vs. combined transection of fornix and amygdalofugal pathways. Exp Brain Res. 1985;57:554–561. doi: 10.1007/BF00237842. [DOI] [PubMed] [Google Scholar]
- 5.Brown MW. Neuronal responses and recognition memory. Semin Neurosci. 1996;8:23–32. [Google Scholar]
- 6.Brown MW, Wilson FA, Riches IP. Neuronal evidence that inferomedial temporal cortex is more important than hippocampus in certain processes underlying recognition memory. Brain Res. 1987;409:158–162. doi: 10.1016/0006-8993(87)90753-0. [DOI] [PubMed] [Google Scholar]
- 7.Bunsey M, Eichenbaum H. Conservation of hippocampal memory function in rats and humans. Nature. 1996;379:255–257. doi: 10.1038/379255a0. [DOI] [PubMed] [Google Scholar]
- 8.Burwell RD, Witter MP, Amaral DG. Perirhinal and postrhinal cortices in the rat: a review of the neuroanatomical literature and comparison with findings from the monkey brain. Hippocampus. 1995;5:390–408. doi: 10.1002/hipo.450050503. [DOI] [PubMed] [Google Scholar]
- 9.Cohen NJ, Eichenbaum H. Memory, amnesia, and the hippocampal system. MIT; Cambridge, MA: 1993. [Google Scholar]
- 10.Deacon TW, Eichenbaum E, Rosenberg P, Eckmann K. Afferent connections of the perirhinal cortex in the rat. J Comp Neurol. 1983;220:168–190. doi: 10.1002/cne.902200205. [DOI] [PubMed] [Google Scholar]
- 11.Dusek J, Eichenbaum H (1997) The hippocampus and memory for orderly stimulus relations. Proc Natl Acad Sci USA, in press. [DOI] [PMC free article] [PubMed]
- 12.Eacott MJ, Gaffan D, Murray EA. Preserved recognition memory for small sets, and impaired stimulus identification for large sets, following rhinal cortex ablations in monkeys. Eur J Neurosci. 1994;6:1466–1478. doi: 10.1111/j.1460-9568.1994.tb01008.x. [DOI] [PubMed] [Google Scholar]
- 13.Eichenbaum H. Is the rodent hippocampus just for “place”? Curr Opin Neurobiol. 1996;6:187–195. doi: 10.1016/s0959-4388(96)80072-9. [DOI] [PubMed] [Google Scholar]
- 14.Eichenbaum H, Pettijohn D, Deluca AM, Chorover SL. Compact miniature microelectrode-telemetry system. Physiol Behav. 1977;18:1175–1178. doi: 10.1016/0031-9384(77)90026-9. [DOI] [PubMed] [Google Scholar]
- 15.Eichenbaum H, Kuperstein M, Fagan A, Nagode J. Cue-sampling and goal-approach correlates of hippocampal unit activity in rats performing an odor discrimination task. J Neurosci. 1987;7:716–732. doi: 10.1523/JNEUROSCI.07-03-00716.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Eichenbaum H, Otto T, Cohen NJ. The hippocampus: what does it do? Behav Neural Biol. 1992;57:2–36. doi: 10.1016/0163-1047(92)90724-i. [DOI] [PubMed] [Google Scholar]
- 17.Eichenbaum H, Otto T, Cohen NJ. Two functional components of the hippocampal memory system. Behav Brain Sci. 1994;17:449–472. [Google Scholar]
- 18.Eichenbaum H, Young BJ, Bunsey MD. Persisting questions about hippocampal function in memory. In: McGaugh JL, Bermudez-Rattoni F, Prado-Alcala R, editors. Plasticity in the central nervous system: learning and memory. Gilford; New York: 1995. pp. 129–148. [Google Scholar]
- 19.Fahy FL, Riches IP, Brown MW. Neuronal activity related to visual recognition memory: long term memory and the encoding of recency and familiarity information in the primate anterior and medial inferior temporal and rhinal cortex. Exp Brain Res. 1993;96:457–472. doi: 10.1007/BF00234113. [DOI] [PubMed] [Google Scholar]
- 20.Fuster JM. Inferotemporal units in selective visual attention and short-term memory. J Neurophysiol. 1990;64:681–697. doi: 10.1152/jn.1990.64.3.681. [DOI] [PubMed] [Google Scholar]
- 21.Fuster JM, Jervey JP. Inferotemporal neurons distinguish and retain behaviourally relevant features of visual stimuli. Science. 1981;219:952–955. doi: 10.1126/science.7233192. [DOI] [PubMed] [Google Scholar]
- 22.Gaffan D. Recognition impaired and association intact in the memory of monkeys after transection of the fornix. J Comp Physiol Psychol. 1974;86:1100–1109. doi: 10.1037/h0037649. [DOI] [PubMed] [Google Scholar]
- 23.Gaffan D. Dissociated effects of perirhinal cortex ablation, fornix transection, and amygdalectomy: evidence for multiple memory systems in the primate temporal lobe. Exp Brain Res. 1994;99:411–422. doi: 10.1007/BF00228977. [DOI] [PubMed] [Google Scholar]
- 24.Gaffan D, Murray EA. Monkeys (Macaca fascicularis) with rhinal cortex ablations succeed in object discrimination learning despite 24-hr intervals and fail at matching to sample despite double sample presentations. Behav Neurosci. 1992;106:30–38. doi: 10.1037//0735-7044.106.1.30. [DOI] [PubMed] [Google Scholar]
- 25.Gaffan D, Gaffan EA, Harrison S. Effects of fornix transection on spontaneous and trained non-matching by monkeys. Q J Exp Psychol. 1984;36:285–303. doi: 10.1080/14640748408402209. [DOI] [PubMed] [Google Scholar]
- 26.Goldman-Rakic PS, Funahashi S, Bruce CJ. Cold Spring Harbor symposia on quantitative biology. LV. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: 1990. Neocortical memory circuits. pp. 1025–1038. [DOI] [PubMed] [Google Scholar]
- 27.Jackson-Smith P, Kesner RP, Chiba AA. Continuous recognition of spatial and nonspatial stimuli in hippocampal-lesioned rats. Behav Neural Biol. 1993;59:107–119. doi: 10.1016/0163-1047(93)90821-x. [DOI] [PubMed] [Google Scholar]
- 28.Jarrard LE. Review: on the role of the hippocampus in learning and memory in the rat. Behav Neural Biol. 1993;60:9–26. doi: 10.1016/0163-1047(93)90664-4. [DOI] [PubMed] [Google Scholar]
- 29.Kesner RP, Boland BL, Davis M. Memory for spatial locations, motor responses, and objects: a triple dissociation among the hippocampus caudate nucleus, and extrastriate visual cortex. Exp Brain Res. 1993;93:462–470. doi: 10.1007/BF00229361. [DOI] [PubMed] [Google Scholar]
- 30.Kubie JL. A driveable bundle of microwires for collecting single-unit data from freely moving rats. Physiol Behav. 1984;32:115–118. doi: 10.1016/0031-9384(84)90080-5. [DOI] [PubMed] [Google Scholar]
- 31.Li L, Miller EK, Desimone R. The representation of stimulus familiarity in anterior inferior temporal cortex. J Neurophysiol. 1993;69:1918–1929. doi: 10.1152/jn.1993.69.6.1918. [DOI] [PubMed] [Google Scholar]
- 32.McClelland JL, McNaughton BL, O’Reilly RC. Why there are complementary learning systems in the hippocampus and neocortex: insights from successes and failures of connectionist models of learning and memory. Psychol Rev. 1995;102:419–457. doi: 10.1037/0033-295X.102.3.419. [DOI] [PubMed] [Google Scholar]
- 33.Meunier M, Bachevalier J, Mishkin M, Murray EA. Effects on visual recognition of combined and separate ablations of the entorhinal and perirhinal cortex in rhesus monkeys. J Neurosci. 1993;13:5418–5432. doi: 10.1523/JNEUROSCI.13-12-05418.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Miller EK, Desimone R. Parallel neuronal mechanisms for short-term memory. Science. 1994;263:520–522. doi: 10.1126/science.8290960. [DOI] [PubMed] [Google Scholar]
- 35.Miller EK, Li L, Desimone R. A neural mechanism for working and recognition memory in inferior temporal cortex. Science. 1991;254:1377–1379. doi: 10.1126/science.1962197. [DOI] [PubMed] [Google Scholar]
- 36.Miller EK, Li L, Desimone R. Activity of neurons in anterior inferior temporal cortex during a short-term memory task. J Neurosci. 1993;13:1460–1478. doi: 10.1523/JNEUROSCI.13-04-01460.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Miyashita Y, Chang HS. Neuronal correlate of pictorial short-term memory in the primate temporal cortex. Nature. 1988;331:68–70. doi: 10.1038/331068a0. [DOI] [PubMed] [Google Scholar]
- 38.Mumby DG, Pinel JPJ. Rhinal cortex lesions and object recognition in rats. Behav Neurosci. 1994;108:11–18. doi: 10.1037//0735-7044.108.1.11. [DOI] [PubMed] [Google Scholar]
- 39.Mumby DG, Wood ER, Pinel JPJ. Object-recognition memory is only mildly impaired in rats with lesions of the hippocampus and amygdala. Psychobiology. 1992;20:18–27. [Google Scholar]
- 40.Murray EA. What have ablation studies told us about the neural substrates of stimulus memory? Semin Neurosci. 1996;8:13–22. [Google Scholar]
- 41.Murray EA, Mishkin M. 40-minute visual recognition memory in rhesus monkeys with hippocampal lesions. Soc Neurosci Abstr. 1996;22:281. [Google Scholar]
- 42.Nadel L. The hippocampus and space revisited. Hippocampus. 1991;1:221–229. doi: 10.1002/hipo.450010302. [DOI] [PubMed] [Google Scholar]
- 43.O’Keefe JA. Place units in the hippocampus of the freely moving rat. Exp Neurol. 1976;51:78–109. doi: 10.1016/0014-4886(76)90055-8. [DOI] [PubMed] [Google Scholar]
- 44.O’Keefe JA, Nadel L. The hippocampus as a cognitive map. Oxford UP; Oxford: 1978. [Google Scholar]
- 45.Otto T, Eichenbaum H. Complementary roles of orbital prefrontal cortex and the perirhinal-entorhinal cortices in an odor-guided delayed non-matching to sample task. Behav Neurosci. 1992a;106:763–776. doi: 10.1037//0735-7044.106.5.762. [DOI] [PubMed] [Google Scholar]
- 46.Otto T, Eichenbaum H. Neuronal activity in the hippocampus during delayed non-match to sample performance in rats: evidence for hippocampal processing in recognition memory. Hippocampus. 1992b;2:323–334. doi: 10.1002/hipo.450020310. [DOI] [PubMed] [Google Scholar]
- 47.Quirk GJ, Muller RU, Kubie JL, Ranck JB. The positional firing properties of medial entorhinal neurons: description and comparison with hippocampal place cells. J Neurosci. 1992;12:1945–1963. doi: 10.1523/JNEUROSCI.12-05-01945.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Riches IP, Wilson FAW, Brown MW. The effects of visual stimulation and memory on neurons of the hippocampal formation and the neighboring parahippocampal gyrus and inferior temporal cortex of the primate. J Neurosci. 1991;11:1763–1779. doi: 10.1523/JNEUROSCI.11-06-01763.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rothblat LA, Kromer LF. Object recognition memory in the rat: the role of the hippocampus. Behav Brain Res. 1991;42:25–32. doi: 10.1016/s0166-4328(05)80036-1. [DOI] [PubMed] [Google Scholar]
- 50.Sakurai Y. Cells in the rat auditory system have sensory-delay correlates during the performance of an auditory working memory task. Behav Neurosci. 1990a;104:856–868. doi: 10.1037//0735-7044.104.6.856. [DOI] [PubMed] [Google Scholar]
- 51.Sakurai Y. Hippocampal cells have behavioral correlates during the performance of an auditory working memory task in the rat. Behav Neurosci. 1990b;104:253–263. doi: 10.1037//0735-7044.104.2.253. [DOI] [PubMed] [Google Scholar]
- 52.Suzuki WA. The anatomy, physiology and functions of the perirhinal cortex. Curr Opin Neurobiol. 1996;6:179–186. doi: 10.1016/s0959-4388(96)80071-7. [DOI] [PubMed] [Google Scholar]
- 53.Suzuki WA, Zola-Morgan S, Squire LA, Amaral DG. Lesions of the perirhinal and parahippocampal cortices in the monkey produce long-lasting memory impairment in the visual and tactual modalities. J Neurosci. 1993;13:2430–2451. doi: 10.1523/JNEUROSCI.13-06-02430.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Swanson LW. Brain maps: Structure of the rat brain. Elsevier; Amsterdam: 1992. [Google Scholar]
- 55.Wiener SI, Paul CA, Eichenbaum H. Spatial and behavioral correlates of hippocampal neuronal activity. J Neurosci. 1989;9:2737–2763. doi: 10.1523/JNEUROSCI.09-08-02737.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Witter MP, Groenewegen HJ, Lopes da Silva Lohman AHM. Functional organization of the extrinsic and intrinsic circuitry of the parahippocampal region. Prog Neurobiol. 1989;33:161–254. doi: 10.1016/0301-0082(89)90009-9. [DOI] [PubMed] [Google Scholar]
- 57.Zhu XO, Brown MW. Changes in neuronal activity related to the repetition and relative familiarity of visual stimuli in rhinal and adjacent cortex of the anaesthetised rats. Brain Res. 1995;689:101–110. doi: 10.1016/0006-8993(95)00550-a. [DOI] [PubMed] [Google Scholar]
- 58.Zhu XO, Brown MW, Aggleton JP. Neuronal signaling of information important to visual recognition memory in rat rhinal and neighbouring cortices. Eur J Neurosci. 1995;7:753–765. doi: 10.1111/j.1460-9568.1995.tb00679.x. [DOI] [PubMed] [Google Scholar]
- 59.Zola-Morgan S, Squire LR, Amaral DG. Lesions of the hippocampal formation but not lesions of the fornix or mamillary nuclei produce long-lasting memory impairment in the monkey. J Neurosci. 1989a;9:898–913. doi: 10.1523/JNEUROSCI.09-03-00898.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zola-Morgan S, Squire LR, Amaral DG, Suzuki WA. Lesions of perirhinal and parahippocampal cortex that spare the amygdala and hippocampal formation produce severe memory impairment. J Neurosci. 1989b;9:4355–4370. doi: 10.1523/JNEUROSCI.09-12-04355.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Zola-Morgan S, Squire LR, Ramus SJ. Severity of memory impairment in monkeys as a function of locus and extent of damage within the medial temporal lobe memory system. Hippocampus. 1994;4:483–495. doi: 10.1002/hipo.450040410. [DOI] [PubMed] [Google Scholar]