

Summary
Dengue fever is a mosquito (Aedes aegypti) ‐transmitted viral disease that is endemic in more than 125 countries around the world. There are four serotypes of the dengue virus (DENV 1‐4) and a safe and effective dengue vaccine must provide protection against all four serotypes. To date, the first vaccine, Dengvaxia (CYD‐TDV), is available after many decades’ efforts, but only has moderate efficacy. More effective and affordable vaccines are hence required. Plants offer promising vaccine production platforms and food crops offer additional advantages for the production of edible human and animal vaccines, thus eliminating the need for expensive fermentation, purification, cold storage and sterile delivery. Oral vaccines can elicit humoural and cellular immunity via both the mucosal and humoral immune systems. Here, we report the production of tetravalent EDIII antigen (EDIII‐1‐4) in stably transformed lettuce chloroplasts. Transplastomic EDIII‐1‐4‐expressing lettuce lines were obtained and homoplasmy was verified by Southern blot analysis. Expression of EDIII‐1‐4 antigens was demonstrated by immunoblotting, with the EDIII‐1‐4 antigen accumulating to 3.45% of the total protein content. Immunological assays in rabbits showed immunogenicity of EDIII‐1‐4. Our in vitro gastrointestinal digestion analysis revealed that EDIII‐1‐4 antigens are well protected when passing through the oral and gastric digestion phases but underwent degradation during the intestinal phase. Our results demonstrate that lettuce chloroplast engineering is a promising approach for future production of an affordable oral dengue vaccine.
Keywords: lettuce chloroplast transformation, dengue fever, recombinant EDIII protein, immunogenicity, gastrointestinal digestion analysis, oral vaccine
Introduction
Dengue fever is caused by four antigenically distinct dengue virus serotypes (DENV‐1, DENV‐2, DENV‐3 and DENV‐4). Dengue viruses belong to the family Flaviviridae and mainly occur in the tropical and subtropical parts of the world (Calisher et al., 1989; Weaver and Vasilakis, 2009). It is estimated that about 3.9 billion people in more than 125 countries are at risk of dengue infection with an annual dengue infection scale of approximately 390 million (Bhatt et al., 2013). Of these 390 million infected people, approximately 500 000 need hospital treatment (WHO position report September 2018, Vannice et al., 2015, 2016; Wichmann et al., 2017). Primary infection with one of the mosquito‐transmitted serotypes usually causes mild dengue fever and provides lifelong immunity to that serotype (Kurane and Ennis, 1992; Simmons et al., 2012). Secondary infections with a heterologous serotype result in more life‐threatening and potentially deadly forms of the disease (WHO, 2009) due to antibody dependent enhancement. This phenomenon is associated with cross‐reactive but non‐neutralizing antibodies produced during the first infection (Halstead, 1988), which enhance the uptake of viruses into Fc‐receptor bearing cells during secondary infection (Dejnirattisai et al., 2010; Halstead, 2003). This increases the initial virus load in the cells, promotes virus replication and ultimately leads to overwhelming of the immune system, causing symptoms like fluid accumulation, plasma leaking, respiratory distress, severe bleeding and organ impairment (WHO, 2015). Recent outbreaks of dengue fever in the southern provinces of China (Xiong and Chen, 2014; Zhang et al., 2014) with over 40 000 reported cases and the occurrence of autochthonous transmissions in the non‐endemic regions of eastern China (Wang et al., 2015; Xu et al., 2007) show how serious this disease is. A safe, effective and affordable dengue vaccine against the four strains is urgently needed to control the disease and meet the WHO goal of reducing dengue morbidity by at least 25% and mortality by at least 50% by 2020 (www.who.int).
As complete eradication of the mosquito vector is impossible, vaccination seems to be the most promising protection strategy against dengue fever. The co‐circulation of the four dengue virus serotypes in most areas together with the complex pathogenesis have considerably hampered vaccine development (Ghosh and Dar, 2015; WHO, 2015). One dengue vaccine has been licensed, Dengvaxia® (CYD‐TDV, developed by Sanofi Pasteur) and approximately five additional vaccine candidates are in clinical development, with two (developed by NIH/Butantan and Takeda) now in phase III trials (Vannice et al., 2016; Wichmann et al., 2017). The CYD‐TDV vaccine is a live recombinant tetravalent dengue vaccine that is currently recommended in three doses at 0, 6 and 12 months, but it only has moderate efficacy. The other five vaccine candidates currently under evaluation in clinical trials include other live‐attenuated vaccines, as well as subunit, DNA and purified inactivated vaccine candidates (www.who.int). Development of dengue vaccine that is effective for infants and children is needed to reduce the dengue burden.
A promising alternative approach is the development of a recombinant protein‐based vaccine able to stimulate the protective immune system in a balanced way. The domain III of the dengue virus envelop protein (EDIII) protrudes from the virus surface to facilitate binding to the host cell surface receptor (Crill and Roehrig, 2001) and mediates membrane fusion (Allison et al., 2001). This approximately 100 amino acid long domain has become the focus of subunit vaccine development (Guzman et al., 2010), because it contains a number of epitopes that elicit serotype specific neutralizing antibodies (Chin et al., 2007; Megret et al., 1992). Different expression systems have been used so far to express recombinant dengue antigens based on the whole envelope protein or the EDIII domain (Batra et al., 2010a; Cardoso et al., 2013; Clements et al., 2010; Ivy et al., 2000; Martínez et al., 2010; McDonald et al., 2009; Saejung et al., 2007; Simmons et al., 1999; Srivastava et al., 2000). Combination of the EDIIIs of the four dengue virus serotypes resulted in a tetravalent fusion protein capable of stimulating the production of virus‐neutralizing antibodies against all four serotypes in mice (Batra et al., 2010b; Etemad et al., 2008). Since a simultaneous and balanced immune response against all four serotypes is essential (Hombach et al., 2005), a vaccine candidate based on the recombinant tetravalent fusion protein appears the best solution.
Genetic engineering of the plastid genome of crops has evolved over the past decades into a promising approach for the production of high‐value products such as biopharmaceuticals, industrial enzymes and diagnostic reagents (Bock, 2015; Clarke and Daniell, 2011; Maliga and Bock, 2011). The distinct characteristics of chloroplast transformation such as site‐specific transgene integration (Daniell, 2006), the absence of epigenetic gene silencing and position effects (Daniell et al., 2001; Rigano et al., 2009; Verma et al., 2010), stacking of transgenes into operons (Bock, 2013) and the excellent biosafety profile of transplastomic plants offer great potential in plant biotechnology. Significant interest in producing recombinant proteins in plastids of crop species with edible parts has resulted in transplastomic carrots (Kumar et al., 2004), tomato (Ruf et al., 2001; Zhou et al., 2008), potato (Sidorov et al., 1999), soybean (Dufourmantel et al., 2004; Moravec et al., 2007), cauliflower (Nugent et al., 2006), eggplant (Singh et al., 2010), cabbage (Liu et al., 2007) and sugar beet (De Marchis et al., 2009). Edible crops offer the potential of oral delivery of therapeutical proteins, resulting in much reduced downstream protein processing costs (Kwon et al., 2013; Streatfield, 2006).
Since the first report of transplastomic lettuce plants, the evaluation of different integration sites and transformation strategies have led to the successful expression of several recombinant proteins in lettuce plastids (Boyhan and Daniell, 2011; Davoodi‐Semiromi et al., 2010; Ichikawa et al., 2010; Kanagaraj et al., 2011; Kanamoto et al., 2006; Maldaner et al., 2013; Ruhlman et al., 2007). Most importantly, lettuce is the only system so far that has been shown to be feasible for commercial scale production of clinical grade biopharmaceuticals (Su et al., 2015). In large animal and toxicology studies (Herzog et al., 2017) biopharmaceuticals were shown to be stable for up to 30 months in lyophilized lettuce cells when stored at ambient temperature, without loss of activity. Furthermore, protection of biopharmaceuticals upon passage of the stomach by bioencapsulation within plant cells has been repeatedly demonstrated. When reaching the gut, commensal bacteria degrade plant cell walls, thereby releasing the protein drugs and facilitating delivery to the immune and/or circulatory systems (Kwon and Daniell, 2016; Xiao et al., 2016). Orally administered plant‐produced antigens can stimulate mucosal IgA and serum IgG production (Lee et al., 2015) and antigen fusion to a transmucosal carrier like the cholera toxin B subunit (CTB) improves the efficiency of antigen delivery to the immune system (Chan and Daniell, 2015; Davoodi‐Semiromi et al., 2010). A further recent example is provided by the production of a plant‐based oral booster vaccine against polio virus, for use in a routine vaccination strategy with a booster dose applied after at least 6 months. The efficacy of the vaccine in eliciting mucosal and humoural immune responses could be demonstrated (Xiao and Daniell, 2017; Xiao et al., 2016; Zhang et al., 2017).
In the current study, we have expressed a tetravalent fusion protein EDIII‐1‐4 in lettuce chloroplasts, performed an in vitro gastrointestinal digestion study and tested the stability of the bioencapsulated recombinant protein throughout the human upper digestive system. Our results showed (i) the successful production of tetravalent EDIII antigen (EDIII1‐4) in lettuce chloroplasts; (ii) molecular analyses of transplastomic EDIII‐1‐4 ‐expressing lettuce lines; (iii) immunoblotting analysis of EDIII‐1‐4 accumulation in lettuce; (iv) immunological assays in rabbits with tetravalent EDIII‐1‐4 antigens; and (v) the results from the in vitro gastrointestinal digestion analysis including oral phase, gastric phase and intestinal phase. Our results indicate that lettuce chloroplast engineering represents a promising approach for the production of a safe and affordable oral dengue vaccine and have generated new information for the dengue vaccine research community.
Results
Generation and characterization of dengue virus EDIII‐1‐4 producing transplastomic lettuce
In order to produce a dengue antigen that covers all four dengue virus serotypes, transplastomic plants expressing the tetravalent antigen EDIII‐1‐4 (Gottschamel et al., 2016) in the chloroplast were generated. To compare expression levels and help ascertain the stability of the linker regions in the tetravalent antigen, a transplastomic line expressing only the EDIII from DENV‐1 was also generated. First, a lettuce plastid transformation vector pDEST‐PN‐L was constructed by insertion of the aadA expression cassette and the Gateway® RfA between lettuce‐specific flanking regions for integration into the plastid genome by homologous recombination. The vectors pEXP‐PN‐ediii‐1‐L and pEXP‐PN‐ediii‐1‐4‐L (Figure 1a) for lettuce plastid transformation were then obtained by Gateway® cloning of the sequences for ediii‐1 and ediii‐1‐4 into the lettuce‐specific pDEST‐PN‐L. Integration by homologous recombination into the intergenic spacer region between the trnI and trnA genes results in transplastomic plants carrying the transgene expression cassettes within the IR region of the lettuce plastid genome (Figure 1b,c).
Figure 1.

Schematic representation of the expression vectors for the generation of transplastomic lettuce plants: (a) The final lettuce‐specific plastid transformation vector pEXP‐PN‐goi‐L. (b) wild‐type lettuce plastid genome (CP). (c) lettuce plastid genome with integrated transgene expression cassettes for ediii‐1 and ediii‐1‐4, separately. The Southern blot probe (trnA) is shown as an arrow and the expected SmaI fragments are shown as arrows with their sizes indicated next to the respective goi. aadA: spectinomycin resistance gene; Amp(R): ampicillin resistance gene; attB1/attB2: Gateway® recombination sites; INSL*/INSR*: lettuce‐specific left/right flanking regions; trnI/trnA: sequences coding for tRNA‐Ile/tRNA‐Ala; EDIII 1/1‐4: transgene coding sequence including a hexa‐his‐tag; PsbA: tobacco psbA promoter (Staub and Maliga, 1993); Prrn16: tobacco rrn16 PEP+NEP promoter (Ye et al., 2001); 3′(C): 3′UTR of Chlamydomonas rbcL gene; 5′psbA: 5′UTR of tobacco psbA gene; 3′(T): 3′UTR of tobacco rbcL gene; ORI: bacterial origin of replication. p296/p297: primer used for PCR (the corresponding PCR products are shown as dotted lines and the sizes are given for both transgenes).
The two transformation constructs were introduced into plastids by particle bombardment. Antibiotic‐resistant shoots developing from callus tissue on RMOP plant regeneration medium containing spectinomycin were tested for transgene integration by PCR. Presence of the transgenic sequences in the plastid genome was shown by PCR products corresponding to ediii‐1‐4 (1841 bp) and ediii‐1 (836 bp) (Figure 2a). The transplastomic plant lines (S12‐PN‐EDIII‐1‐4 and S16‐PN‐EDIII‐1 respectively) were further characterized by Southern blot analysis. The homoplastomic state of both plant lines was verified by the presence of only the 5545 bp fragment (in S16‐PN‐EDIII‐1) or the 6533 bp fragment (in S12‐PN‐EDIII‐1‐4) in transformed plants, compared to the 3130 bp fragment diagnostic of the wild‐type plastid genome (Figure 2b) after digestion of total plant DNA with SmaI.
Figure 2.

Southern blot analysis of tetravalent EDIII‐1‐4 and monovalent EDIII‐1 lines. DNA isolated from regenerated plant lines and the wild‐type (wt) was probed using a 665 bp DIG‐labelled probe that binds inside the trnA region (INSR) of the plastid genome. The expected fragment sizes after SmaI digestion are 6533 bp (for S12‐PN‐EDIII‐1‐4), 5545 bp (for S16‐PN‐EDIII‐1) and 3130 bp (for wild‐type plants). The positions of restriction sites, probe position and the sizes of expected Southern blot bands are indicated in Figure 1. M: 1 kb DNA ladder, (NEB).
No phenotypic alterations were visible on transplastomic plants growing to maturity in the greenhouse (Figure 3a) and flower set and seed development was normal. Plants were grown to full maturity (Figure 3b) and seeds harvested from transgenic plants were germinated on spectinomycin‐containing medium. The homogenous green phenotype of the seedlings proved the absence of segregation of the antibiotic resistance gene in the F1 generation (Figure 3c) provided additional proof of transgene integration into the plastid genome and complete elimination of wild‐type copies of the (polyploid) plastid genome.
Figure 3.

Phenotype of transplastomic lettuce plants and inheritance assays. (a) Plants growing in the greenhouse. (b) Flowering plants. (c) One‐week‐old seedlings obtained from transplastomic plants and wild‐type seeds germinated on spectinomycin (30 mg/L) containing medium.
Expression of EDIII‐1‐4 and EDIII‐1 antigens
In order to assess whether the antigens were produced and accumulated stably in lettuce chloroplasts, total protein (TP) and total soluble protein (TSP) were isolated from plant lines growing in the greenhouse and quantified by BCA and Bradford assays respectively. Immunoblot analysis performed with an anti‐dengue antibody detected both the 47 kDa EDIII‐1‐4 and the 13 kDa EDIII‐1 in the respective transplastomic lettuce lines, in both the TP and TSP samples (Figure 4). The single‐domain antigen EDIII‐1 accumulated to lower levels (Figure 4b) and this line was not studied further. The tetravalent antigen EDIII‐1‐4 was only detected as a single band corresponding to the full‐length antigen, indicating that the linker regions are stable in the chloroplast and no significant proteolytic cleavage occurs. The EDIII‐1‐4 accumulation was quantified in TP extracts by using purified E. coli‐produced tetravalent antigen (Gottschamel et al., 2016) as a standard. These assays (Figure S1) established that the EDIII‐1‐4 fusion protein was present at 3.45% ± 0.05 of the TP content in the transplastomic lettuce plants.
Figure 4.
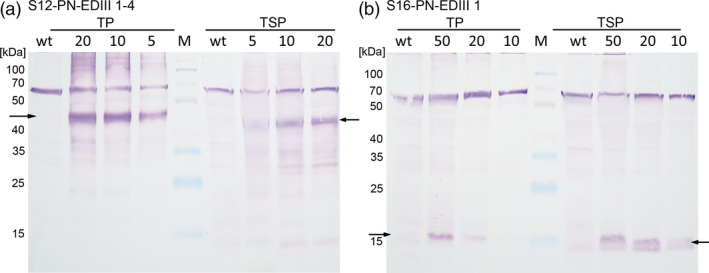
Western blot analysis of EDIII antigen accumulation in transplastomic lettuce plants. (a) TP and TSP isolated from plant line S12‐PN‐EDIII‐1‐4. (b) TP and TSP isolated from plant line S16‐PN‐EDIII‐1. The amount of TSP/TP loaded is given above the respective lane in μg. Twenty microgram TP/TSP was loaded for the wild‐type in a), while 50 μg TP/TSP was loaded for the wild‐type in b). The arrows indicate the 47 kDa EDIII‐1‐4 and the 13 kDa EDIII‐1. M: spectra multicolour broad range protein ladder (Thermo Scientific, molecular weight of the marker bands indicated in kDa). The band migrating at 70 kDa results from non‐specific binding of the antibody to a plant protein of unknown identity (Gottschamel et al., 2016).
Immunogenicity of lettuce‐produced EDIII‐1‐4 in rabbits
Next, we wanted to test the immunogenicity of the lettuce‐produced EDIII‐1‐4 antigens. To this end, the fusion protein was isolated from the transplastomic lettuce line S12‐PN‐EDIII‐1‐4 under native conditions and used for immunization of one rabbit. Rabbit serum was analysed by an enzyme‐linked immunosorbent assay (ELISA), where binding to lettuce‐produced and E. coli‐produced EDIII‐1‐4 was assayed. Injection of lettuce‐produced dengue antigen EDIII‐1‐4 elicited antibodies detectable by ELISA at 9 weeks post vaccination (Figure 5). It is interesting to note that the rabbit serum recognized the lettuce‐produced antigen (mean antibody titre 1.0235 OD490, SD = 0.4305) and the E. coli‐produced EDIII‐1‐4 antigen (mean antibody titre 0.9598 OD490, SD = 0.2565) equally well, confirming that the antigens are specifically recognized by the serum. The results of our immunological analyses show that rabbit vaccination with lettuce‐produced EDIII‐1‐4 antigen elicits a specific immune response.
Figure 5.

Immunogenicity of lettuce‐produced tetravalent EDIII 1‐4. Sera of a rabbit immunized with lettuce‐produced EDIII‐1‐4 antigen at 63 days post vaccination (EDIII‐1‐4) were tested for binding to E. coli‐produced EDIII‐1‐4 antigen and lettuce‐produced EDIII‐1‐4 antigen. Rab neg ctrl is serum from non‐vaccinated rabbit; Rab Pos Ctrl represents a rabbit polyclonal antibody against a synthetic EDIII peptide (Gottschamel et al., 2016) used as a positive control. All sera were used in a 1 : 50 dilution.
Stability of lettuce‐encapsulated EDIII‐1‐4 in a gastrointestinal tract model
Simulated gastrointestinal digestion is widely employed in food science, nutritional studies and pharmaceutical research, as conducting human trials is costly, resource intensive and ethically disputable. The methodology is based on mimicking physiological conditions in vitro, taking into account the presence and physiological concentrations of digestive enzymes as well as the pH, ionic milieu and digestion time, among other factors (Minekus et al., 2014). Simulated digestion of lettuce chloroplast‐derived EDIII‐1‐4 tetravalent antigen through the oral, gastric and small intestinal phases (Figure 6) showed that EDIII‐1‐4 tetravalent antigen was well protected when passing through the oral phase and the gastric phase, but showed degradation after passing through the intestinal phase. The latter is consistent with the (desired) release of the antigen from the chloroplasts in the intestine, where it is intended to stimulate mucosal immunity.
Figure 6.

In vitro gastrointestinal digestion analysis of the lettuce chloroplast‐derived EDIII‐1‐4 tetravalent antigen. Western blot analysis of proteins isolated from samples taken for each phase of the gastrointestinal digestion analysis are shown. Start: untreated lettuce material; − samples treated with only buffer; + samples treated with the digestive enzymes. Each phase was analysed in three replicates with enzymes and one blank sample without enzymes as control. Thirty microgram TP was loaded for each sample. The band migrating at 70 kDa results from non‐specific binding of the antibody to a plant protein of unknown identity.
Discussion
Despite over 70 years of efforts to combat the disease, dengue fever is still a major health threat in significant parts of the world. In view of the expected further spread of mosquito‐borne diseases with climate change, the need for dengue vaccine development becomes even more pressing (Pang and Loh, 2017). Efficacy and safety remain the main challenges (Wichmann et al., 2017; www.who.int). Clinical studies with the licensed Dengvaxia® (CYD‐TDV, a live‐attenuated, recombinant tetravalent vaccine employing the attenuated YF virus 17D vaccine strain as the replication backbone) have been further investigated since 2015. The new analyses from the long‐term safety follow‐up indicated that overall the population level benefit of vaccination remains favourable, but the vaccine performs differently in seropositive versus seronegative individuals. Vaccine efficacy (VE) confirmed that symptomatic dengue was high among inferred baseline seropositive participants ≥9 years of age: 76% (95% CI: 63.9, to 84.0), but was much lower among baseline seronegative participants: 38.8% (95% CI: −0.9 to 62.9) in the first 25 months after the first dose of vaccine (www.who.int, Wichmann et al., 2017). These data demonstrated long‐term protection in seropositive individuals. However, at the same time, the studies also revealed an excess of hospitalized and severe dengue cases in seronegative vaccine recipients compared to seronegative non‐vaccinated individuals and there was an increase in the number of young vaccinated children hospitalized within 3 years after the start of vaccination (Wichmann et al., 2017). These data provide a strong incentive to look for improved vaccination strategies. A safe, effective and affordable vaccination strategy shall include careful screening pre‐vaccination, the affordability of both CYD‐TDV and screening tests and assessing the need and feasibility of oral booster dengue vaccine (Wichmann et al., 2017). Vaccination as part of an integrated dengue prevention and control strategy remains a high priority (Wichmann et al., 2017). More efforts must be invested into the development of safe, effective and affordable dengue vaccines that, ideally, should be independent of the serostatus (Wichmann et al., 2017) and employ an oral vaccine and/or an oral booster vaccine produced in an edible plant such as lettuce.
We have previously reported synthesis of an EDIII‐1‐4 tetravalent antigen‐based subunit dengue vaccine in tobacco chloroplasts (Gottschamel et al., 2016). In the present study, we have successfully expressed the tetravalent antigen in chloroplasts of the edible crop lettuce, assayed its stability upon passage through the gastrointestinal tract and determined its antigenicity in experimental animals. Further efforts in producing other dengue antigens in plants have also been reported (Kanagaraj et al., 2011; Maldaner et al., 2013). Our work adds to a growing number of studies in plant molecular farming that have demonstrated that production of vaccines and antibodies in plants is a very attractive alternative to traditional production systems. Plants offer a number of unique advantages over more traditional systems (Bock, 2015; Clarke and Daniell, 2011; Peyret and Lomonossoff, 2015), including cost‐effective production of recombinant proteins at large scale (due to the very low production costs of plant biomass and the easily scalable production levels), the low risk of contamination with human pathogens and the possibility of oral delivery. The latter eliminates the need for expensive downstream processing (usually about 80% of the production cost of pharmaceuticals and vaccines), makes vaccine administration simple and safe (Chan and Daniell, 2015; Clarke et al., 2017) and provides stability of antigens at room temperature, a highly desired property of vaccines.
Protein drugs expressed within plant cells can be protected from acids and enzymes in the stomach (Chan and Daniell, 2015). To assess the stability of the tetravalent EDIII‐1‐4 antigens produced in chloroplasts after oral delivery, we have conducted in vitro gastrointestinal tract assays. The simulated digestion of lettuce chloroplast‐derived EDIII‐1‐4 tetravalent antigen through the oral, gastric and small intestinal phases has shown that the antigen was well‐protected when passing through the oral and gastric phases and showed degradation upon passing through the intestinal phase (Figure 6). The latter is not only consistent with the expected release of the protein from the plant cells in the intestine, but also desirable to expose the antigen to the mucosal immune system.
The prime‐and‐boost based vaccination strategy has been exploited with plant‐made boost vaccines against HIV and polio virus (Chan et al., 2016; Lindh et al., 2014). The same strategy could be applied to dengue fever, given that, with the current vaccine, the immune response varies greatly among different people, age groups and regions. Moreover, the incidence of children developing dengue infection within 3 years after vaccination in one of the CYD‐TDV dengue vaccine clinical trials (Wichmann et al., 2017) suggests the potential benefit of a booster vaccine for young children. Together with a previous report (Chan and Daniell, 2015), our study suggests that the edible crop lettuce is ideal for the production of oral boost vaccines. This is not only because lettuce can be eaten raw, but also because the edible parts of the plants are the leaves. A number of studies have shown that expression of plastid genes and transgenes in non‐green tissues (including fruits and tubers) is much lower than in photosynthetically active leaf tissue (Kahlau and Bock, 2008; Valkov et al., 2009). A plant‐made dengue boost vaccine could contribute to affordable dengue treatment and reduce the burden on the health sector in developing countries. Lettuce chloroplast genome engineering could contribute to an oral boost dengue vaccine in the future.
In conclusion, we have demonstrated in the present study the successful production of the tetravalent EDIII dengue virus antigen (EDIII‐1‐4) in lettuce chloroplasts. Homoplastic transplastomic lettuce lines expressing EDIII‐1‐4 tetravalent dengue vaccine antigen were obtained and homoplasmy was verified by Southern blot analysis. The expressed tetravalent EDIII‐1‐4 antigen was detected in immunoblotting and quantified. Immunological assays showed immunogenicity in rabbits. Our in vitro gastrointestinal digestion analysis with EDIII‐1‐4‐producing lettuce has shown that EDIII‐1‐4 antigens were well protected when passing through the oral and gastric phases. Our results suggest that lettuce chloroplast engineering is a promising approach for the future production of a safe, affordable and effective oral dengue boost vaccine that can contribute to the control and management of dengue infection globally.
Methods
Vector construction
The lettuce‐specific plastid transformation vector was obtained by Gateway® cloning (Gottschamel and Lössl, 2016;). For the intermediary vector pMA‐lettuce, the sequences corresponding to the trnI and trnA region of the Lactuca sativum plastid genome (Ruhlman et al., 2007) flanked by KpnI and SacII restriction sites were custom synthesized and introduced into the company's standard backbone vector (GeneArt, Germany). In order to create pDEST‐PN‐L, the complete Gateway® cloning cassette together with the aadA expression cassette was excised from pDEST‐PN‐T (Gottschamel et al., 2013) and inserted into pMA‐lettuce using the restriction enzymes KpnI and SacII. The sequence of the synthetic fusion gene (ediii‐1‐4) consists of all four DENV‐EDIII sequences (order: ediii‐1, ediii‐3, ediii‐4, ediii‐2) linked by penta‐glycine linkers (Etemad et al., 2008; Gottschamel et al., 2016). The ediii‐1‐4 and ediii‐1 sequences were codon optimized for lettuce plastids and synthesized by GeneArt (Germany). The synthesized sequences have a T7g10 leader sequence and a 15 nucleotide downstream box (Herz et al., 2005), a C‐terminal 6xHis‐tag and are flanked with attB1/attB2 Gateway® recombination sites. The transgene encoding sequences were first introduced into pDONR™221 by a BP reaction yielding the intermediary vectors pEntry‐ediii‐1‐4 and pEntry‐ediii‐1 and then transferred by an LR reaction into pDEST‐PN‐L resulting in the final lettuce‐specific plastid transformation vectors pEXP‐PN‐ediii‐1‐4‐L and pEXP‐PN‐ediii‐1‐L respectively. The PCR Cloning Kit with Gateway® Technology, pDONR™ 221 and the Gateway® LR Clonase® Enzyme mix were purchased from Life Technologies (Carlsbad, CA) and the Gateway® BP and LR reactions (Karimi et al., 2002) were carried out as described in the manufacturer's protocol.
Plant growth, transformation and regeneration
Lactuca sativa cv. Barkley plants were grown in vitro from surface sterilized seeds on solid MS medium (Murashige and Skoog, 1962) containing 20 g/L sucrose. Leaves from aseptically grown lettuce plants were bombarded with 0.6 μm gold‐microcarriers coated with plasmid DNA using a Bio‐Rad Biolistic PDS‐1000/He gun (Daniell, 1997; Svab and Maliga, 1993) Several independently transformed plant lines were subjected to three additional regeneration rounds on RMOP medium (Svab and Maliga, 1993; Verma et al., 2008) containing spectinomycin. Regenerated shoots were rooted on MS medium containing spectinomycin to maintain the selection pressure. Rooted, homoplastomic plants were transferred to soil and grown to maturity in the greenhouse under standard conditions. Inheritance assays on spectinomycin‐containing MS medium were performed with the harvested seeds.
Southern blot analysis
Plant DNA was isolated by the CTAB procedure (Murray and Thompson, 1980) from wild‐type plants and transplastomic plant lines after three rounds of regeneration on spectinomycin‐containing medium. Ten microgram of plant DNA was digested with SmaI, separated by electrophoresis in a 1% agarose gel and transferred onto a positively charged nylon membrane (Carl Roth GmbH, Karlsruhe, Germany) by capillary action using the semi‐dry transfer method. The probe binding inside the trnA region was amplified from lettuce wild‐type DNA by PCR (primers 5′‐GGAGGTAGGATGGGCAGTTG‐3′ and 5′‐GGACTCGAACCGCTGACATC‐3′). The probe was purified by agarose gel electrophoresis, followed by extraction of the fragment of interest from excised gel slices with the NucleoSpin gel and PCR Clean‐up Kit (Machery‐Nagel, Düren, Germany). The probe was DIG labelled using the DIG‐High Prime DNA Labeling and Detection Starter Kit II (Roche, Basel, Switzerland), according to the manufacturer's instructions. After immobilization of the DNA to the membrane, hybridization with the corresponding DIG labelled‐probe and incubation of the membrane with the HRP conjugated anti‐DIG antibody, the chemiluminescence signal was detected by exposure to X‐ray film. One homoplastomic plant line per construct (S12‐PN‐EDIII‐1‐4 and S16‐PN‐EDIII‐1) was chosen for further analysis.
Protein extraction, Western blot analysis and protein quantification
Protein extraction and Western blot analysis was done as described previously (Gottschamel et al., 2016). Briefly, total soluble protein (TSP) was extracted by mixing frozen and grinded leaf samples in TSP extraction buffer [100 mm NaCl, 10 mm EDTA, 200 mm Tris‐HCl pH 8, 0.05% Tween‐20, 0.1% SDS, 14 mm β‐mercaptoethanol, 200 mm sucrose and 1× cOmplete protease inhibitor (Roche)] for 5 min on ice, followed by centrifugation at 15 000 g for 10 min at 4 °C and collecting the supernatant. The TSP concentration was determined with the Bradford assay (Bio‐Rad, Hercules, CA) using known concentrations of BSA as the standard. Total protein (TP) was isolated from leaf samples by the phenol extraction method (Cahoon et al., 1992). Frozen ground leaf samples were homogenized in TP extraction buffer [0.7 m sucrose, 0.5 m Tris‐HCl pH 9.4, 50 mm EDTA, 0.1 m KCl, 2% β‐mercaptoethanol, 1× cOmplete protease inhibitor (Roche, Switzerland)]. After addition of 1 vol. phenol, short vortexing and centrifugation at 13 000 r.p.m. for 10 min at 4 °C, the upper green phase was recovered and proteins were precipitated by addition of 0.1 m NH4OAc in methanol and overnight incubation at −20 °C. After centrifugation, the protein pellet was washed, air‐dried and dissolved in 1% SDS. The protein concentration was determined with the BCA Protein Assay Kit (Thermo Scientific, Waltham, MA) using known concentrations of BSA as the standard. Denatured protein samples were separated by electrophoresis in 12% SDS‐polyacrylamide gels and transferred to nitrocellulose membranes (Hybond‐ECL; GE Healthcare, Chicago, IL). The membranes were incubated with TBS‐T (20 mm Tris‐HCl pH 7.6, 137 mm NaCl, 0.1% Tween‐20,) containing 0.5% BSA as blocking buffer and subsequently treated with the primary and the secondary antibody diluted in TBS‐T. The recombinant proteins were detected with the 1:1000 diluted polyclonal anti‐dengue antibody produced in rabbits against amino acid sequence KFKVVKEIAETQHGT (by Davids Biotechnology, Regensburg, Germany), the 1:10 000 diluted anti‐rabbit‐IgG‐AP secondary antibody (Promega, Fitchburg, WI) and colorimetric reaction using the AP color development Kit (Bio‐Rad). Recombinant EDIII‐1‐4, expressed in E.coli (Gottschamel et al., 2016) and purified with the HisPur Cobalt resin (Life Technologies) under denaturing conditions served as a positive control.
Protein quantification was done by Western blot analysis. Samples with known quantities of the purified E. coli‐produced antigen were loaded together with TP extract samples. Blotting was done as described above, but as secondary antibody 1 : 10 000 diluted anti‐rabbit‐IgG‐HRP secondary antibody (Promega) was used. Blot development was done with ECL Prime Western blotting detection reagent (GE Healthcare) and blots were imaged using an Azure C400 imaging system (Azure Biosystems, Dublin, CA) and analysed with ImageJ (NIH, Bethesda, MD). Experiments were repeated four times.
Immunological studies
The study was performed at the Section for Experimental Biomedicine at The Norwegian University of Life Sciences, Norway. The unit is licensed by the Norwegian Animal Research Authority (NARA) (http://www.mattilsynet.no/dyr_og_dyrehold/dyrevelferd/forsoksdyr/) and accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (www.aaalac.org). The study was approved by the unit's animal ethics committee (Institutional Animal Care and Use Committee/IACUC) and NARA.
Animal model
The rabbit used in this study was a female New Zeeland White SPF (Harlan Laboratories, Horst, Netherlands). It had 3 weeks of acclimation at the animal facility before starting the immunization. The rabbit was housed and taken care of according to the requirements in the European Union Directive 2010/63/EU and Norway's own regulation based on the EU directive ‘The regulation on use of animals in research FOR‐2015‐06‐18‐761’.
Tetravalent dengue antigen EDIII‐1‐4 purification and immunization
The EDIII‐1‐4 tetravalent antigen was purified from lettuce line S12‐PN‐EDIII‐1‐4. Frozen leaves were ground to powder, mixed with an equal volume of extraction buffer [0.05 m Tris pH 9.2, 0.5 m NaCl, 0.1% (v/v) Tween 20, 15 mm β‐mercaptoethanol] and incubated on ice for 20 min. The mixture was filtered through four layers of Miracloth (Merck, Darmstadt, Germany) and the filtrate was centrifuged at 25 000 g for 40 min at 4 °C. Imidazole pH 9 to a final concentration of 20 mm was added to the supernatant, which was then mixed with NiNTA agarose beads (Qiagen, Hilden, Germany) and incubated under mixing for 3 h at 22 °C. The beads were collected by low‐speed centrifugation and washed with buffer (20 mm imidazole pH 8.6, 0.5 m NaCl, 0.1% Tween 20) and subsequently eluted with elution buffer (0.3 m imidazole pH 8.6, 0.5 m NaCl, 0.1% Tween 20). The buffer was exchanged to 20 mm Tris pH 9, 0.15 m NaCl, 0.1% Tween 20 and the protein was concentrated to 0.2 mg/mL by ultracentrifugation using a 10 kDa MWCO Microsep device (Pall, Westborough, MA).
Injections were done by a veterinarian with FELASA C certification. The rabbit was given one immunization injection and three booster injections. The first immunization was a mixture of 0.5 mL solution containing 0.1 mg antigen and 0.5 mL Freund`s Adjuvant Complete, whereas all three booster injections contained the same amount of antigen but were instead prepared with 0.5 mL Freund`s Adjuvant Incomplete (both from Sigma‐Aldrich, Oslo, Norway). The first booster was given 2 months after the first immunization. All subsequent booster injections were given in 2‐week intervals. The solution was mixed right before injection in two coupled 2 mL Omnifix Luer Lock syringes and then injected with a 23 G Terumo needle (all from Jan F. Andersen, Jevnaker, Norway). All injections were given divided on 5‐10 places. Blood samples were collected before the first and after the last injection, to analyse the antibodies. The blood was taken from the ear artery with a 4 mL Vacuette Serum container and a 0.7 × 25 mm needle (Jan F. Andersen, Jevnaker, Norway). Both the injections and the blood samples were done while the rabbits were sedated with Hypnorm (Fentanyl 0.315 mg/mL and Fluanison 10 mg/mL) 0.03 mL/kg iv. Two weeks after the last booster injection both rabbits were terminally bled under full anaesthesia. They were premedicated with Domitor (Medetomidin 1 mg/mL) 0.1 mL/kg im and anaesthetized with a Zoletil Mix [Zoletil dry matter, 10 mL Rompun (Xylazin 20 mg/mL) and 0.75 mL Torbugesic (Butorphanol 10 mg/mL)] (all from VESO Pharmacy, Oslo, Norway) 0.1 mL/kg im. The blood was taken via heart puncture with a 10 mL Vacuette Serum container and a 0.7 × 25 mm needle (Jan F. Andersen, Jevnaker, Norway). After the bleeding, the rabbits were sacrificed with Penthobarbithal injection to the heart and opening of the thorax.
Enzyme‐linked immunosorbent assay (ELISA)
ELISA was carried out by coating two plates with a commercial anti‐dengue mouse monoclonal antibody (3H5 against dengue type 2, Temecula, CA) at a dilution of 1 : 1000. The coated plates (NUNC, Denmark) were incubated at 4 °C overnight. After incubation, the plates were washed three times with phosphate buffered saline containing 1% Tween 20 (PBST). This was followed by blocking each well with 5% dry milk and incubation at room temperature (RT) for 2 h. After washing three times with PBST, the denatured EDIII‐1‐4 produced in E. coli was added to plate 1, while lettuce‐produced EDIII‐1‐4 was added to plate 2 and incubated at RT for 2 h. After washing three times with PBST, the vaccinated rabbit serum was added to each plate. For the positive control, the polyclonal rabbit antiserum raised against EDIII peptide KFKVVKEIAETQHGT (Gottschamel et al., 2016) was used while for the negative control, serum obtained from an unvaccinated rabbit was used. All serum samples were applied as a dilution series, starting from a 1 : 50 dilution, then 1 : 100, 1 : 200, etc. The plates were incubated at RT for 2 h followed by washing three times with PBST. Thereafter, a goat anti‐rabbit polyclonal antibody conjugated to horse radish peroxidase (HRP) (DAKO; Glostrup, Denmark) was added to each well at a dilution of 1 : 1000. The plates were incubated for 1 h followed by washing. This was followed by adding the substrate made of OPD tablets (O‐Phenylenediamine dihydrochloride, DAKO; Glostrup, Denmark) plus 5% hydrogen peroxide (H2O2). The plates were observed for 15 min for colour change followed by adding 0.05 μL of the stop solution (1 mm H2SO4). The plates were read using a spectrophotometer (TECAN, Genios, Boston, MA) at an optical density of OD490. A Student's t‐test was used in statistical analysis.
In vitro gastrointestinal digestion analysis
The harmonized, static in vitro digestion model (Minekus et al., 2014) was performed with fresh leaves from the lettuce plant line S12‐PN‐EDIII‐1‐4. The plant material was cut into pieces of 5 mm2 and a total amount of 2 g plant material per reaction was incubated under shaking in a water bath at 37 °C. The model consists of three stages: stage I ‐ oral phase: buffer (simulated salivary fluid: 15.1 mm KCl, 3.7 mm KH2PO4, 13.6 mm NaHCO3, 0.15 mm MgCl2(H2O)6, 0.06 mm (NH4)2CO3, 1.5 mm CaCl2(H2O)2, 1.1 mm HCl, pH 7), 0.44 U salivary α‐amylase (Sigma‐Aldrich, St. Louis, MO), 2 min reaction time; stage II ‐ gastric phase: buffer (simulated gastric fluid: 6.9 mm KCl, 0.9 mm KH2PO4, 25 mm NaHCO3, 47.2 mm NaCl, 0.12 mm MgCl2(H2O)6, 0.5 mm (NH4)2CO3, 0.15 mm CaCl2(H2O)2, 15.6 mm HCl, pH 3), 90 U porcine pepsin (Sigma‐Aldrich), 2 h incubation time; and stage III ‐ duodenal stage: buffer (simulated intestinal fluid (6.8 mm KCl, 0.8 mm KH2PO4, 85 mm NaHCO3, 38.4 mm NaCl, 0.33 mm MgCl2(H2O)6, 0.6 mm CaCl2(H2O)2, 8.4 mm HCl, pH 7), porcine pancreatin (4340 U protease; Sigma‐Aldrich) and 10 mm porcine bile extract (Sigma‐Aldrich), 2 h incubation time. Reactions in samples from all phases were stopped and neutralized by mixing with a TP extraction buffer containing protease inhibitors, as described above and immediately snap frozen in liquid nitrogen. Total protein was extracted as described above and subjected to Western Blot analysis.
Author contributions
AvE: wrote the manuscript, helped with experiments. JG: this study was part of her PhD project; she carried out plant‐related experiments and the GI tract assay and wrote the manuscript. RB: was an external co‐supervisor to JG and involved in project planning and trouble‐shooting and contributed to writing. KEAH: performed the rabbit study, sample collection and contributed to the writing. HMM: conducted immunogenicity analysis and contributed to the writing. HD: gave advice about plant‐made oral vaccines to JLC and contributed to manuscript writing. JLC: was the project leader and co‐supervisor to JG. JLC designed the study, followed up the study through the whole project period and contributed considerably to the manuscript preparation.
Conflict of interest
There is no conflict of interest for the current study.
Supporting information
Figure S1 Quantification of EDIII‐1‐4 accumulation in transplastomic lettuce plants.
Acknowledgements
This work was supported by the GLOBVAC program (Project 192510) of the Research Council of Norway (RCN), NIBIO core funding and the Max Planck Society. The authors thank Hege Særvold Steen and Sissel Haugslien for technical support and fruitful discussions, Professor Øystein Evensen, Norwegian University of Life Sciences, for his kind help with the animal study and for reading the manuscript, Dr Anna‐Marja Aura, VTT, Finland for support on GI Tract analysis and Dr Andreas Lössl, BOKU, Austria, for valuable discussions.
References
- Allison, S.L. , Schalich, J. , Stiasny, K. , Mandl, C.W. and Heinz, F.X. (2001) Mutational evidence for an internal fusion peptide in flavivirus envelope protein E. J. Virol. 75, 4268–4275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Batra, G. , Raut, R. , Dahiya, S. , Kamran, N. , Swaminathan, S. and Khanna, N. (2010a) Pichia pastoris‐expressed dengue virus type 2 envelope domain III elicits virus‐neutralizing antibodies. J. Virol. Methods, 167, 10–16. [DOI] [PubMed] [Google Scholar]
- Batra, G. , Talha, S.M. , Nemani, S.K. , Dhar, N. , Swaminathan, S. and Khanna, N. (2010b) Expression, purification and characterization of in vivo biotinylated dengue virus envelope domain III based tetravalent antigen. Protein Expr. Purif. 74, 99–105. [DOI] [PubMed] [Google Scholar]
- Bhatt, S. , Gething, P.W. , Brady, O.J. , Messina, J.P. , Farlow, A.W. , Moyes, C.L. , Drake, J.M. et al. (2013) The global distribution and burden of dengue. Nature, 496, 504–507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bock, R. (2013) Strategies for metabolic pathway engineering with multiple transgenes. Plant Mol. Biol. 83, 21–31. [DOI] [PubMed] [Google Scholar]
- Bock, R. (2015) Engineering plastid genomes: methods, tools, and applications in basic research and biotechnology. Annu. Rev. Plant Biol. 66, 211–241. [DOI] [PubMed] [Google Scholar]
- Boyhan, D. and Daniell, H. (2011) Low‐cost production of proinsulin in tobacco and lettuce chloroplasts for injectable or oral delivery of functional insulin and C‐peptide. Plant Biotechnol. J. 9, 585–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahoon, E.B. , Shanklin, J. and Ohlrogge, J.B. (1992) Expression of a coriander desaturase results in petroselinic acid production in transgenic tobacco. Proc. Natl Acad. Sci. U S A, 89, 11184–11188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calisher, C.H. , Karabatsos, N. , Dalrymple, J.M. , Shope, R.E. , Porterfield, J.S. , Westaway, E.G. and Brandt, W.E. (1989) Antigenic relationships between flaviviruses as determined by cross‐neutralization tests with polyclonal antisera. J. Gen. Virol. 70(Pt 1), 37–43. [DOI] [PubMed] [Google Scholar]
- Cardoso, S.A. , Paixão, V.F. , Oliveira, M.D. , Honda, E.R. , Oliveira, L.L. , da Silva, C.C. and De Paula, S.O. (2013) Dengue‐1 envelope protein domain III produced in Pichia pastoris: potential use for serological diagnosis. Protein Expr. Purif. 92, 9–13. [DOI] [PubMed] [Google Scholar]
- Chan, H.T. and Daniell, H. (2015) Plant‐made oral vaccines against human infectious diseases‐Are we there yet? Plant Biotechnol. J. 13, 1056–1070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan, H.T. , Xiao, Y. , Weldon, W.C. , Oberste, S.M. , Chumakov, K. and Daniell, H. (2016) Cold chain and virus‐free chloroplast‐made booster vaccine to confer immunity against different poliovirus serotypes. Plant Biotechnol. J. 14, 2190–2200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chin, J.F. , Chu, J.J. and Ng, M.L. (2007) The envelope glycoprotein domain III of dengue virus serotypes 1 and 2 inhibit virus entry. Microbes Infect. 9, 1–6. [DOI] [PubMed] [Google Scholar]
- Clarke, J.L. and Daniell, H. (2011) Plastid biotechnology for crop production: present status and future perspectives. Plant Mol. Biol. 76, 211–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke, J.L. , Paruch, L. , Dobrica, M.O. , Caras, I. , Tucureanu, C. , Onu, A. , Ciulean, S. et al. (2017) Lettuce‐produced hepatitis C virus E1E2 heterodimer triggers immune responses in mice and antibody production after oral vaccination. Plant Biotechnol. J. 15, 1611–1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements, D.E. , Coller, B.A. , Lieberman, M.M. , Ogata, S. , Wang, G. , Harada, K.E. , Putnak, J.R. et al. (2010) Development of a recombinant tetravalent dengue virus vaccine: immunogenicity and efficacy studies in mice and monkeys. Vaccine, 28, 2705–2715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crill, W.D. and Roehrig, J.T. (2001) Monoclonal antibodies that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. J. Virol. 75, 7769–7773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniell, H. (1997) Transformation and foreign gene expression in plants by microprojectile bombardment. Methods Mol. Biol. 62, 463–489. [DOI] [PubMed] [Google Scholar]
- Daniell, H. (2006) Henry Daniell: chloroplast genetic engineering. Biotechnol. J. 1, 31–33. [DOI] [PubMed] [Google Scholar]
- Daniell, H. , Lee, S.B. , Panchal, T. and Wiebe, P.O. (2001) Expression of the native cholera toxin B subunit gene and assembly as functional oligomers in transgenic tobacco chloroplasts. J. Mol. Biol. 311, 1001–1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davoodi‐Semiromi, A. , Schreiber, M. , Nalapalli, S. , Verma, D. , Singh, N.D. , Banks, R.K. , Chakrabarti, D. et al. (2010) Chloroplast‐derived vaccine antigens confer dual immunity against cholera and malaria by oral or injectable delivery. Plant Biotechnol. J. 8, 223–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Marchis, F. , Wang, Y. , Stevanato, P. , Arcioni, S. and Bellucci, M. (2009) Genetic transformation of the sugar beet plastome. Transgenic Res. 18, 17–30. [DOI] [PubMed] [Google Scholar]
- Dejnirattisai, W. , Jumnainsong, A. , Onsirisakul, N. , Fitton, P. , Vasanawathana, S. , Limpitikul, W. , Puttikhunt, C. et al. (2010) Cross‐reacting antibodies enhance dengue virus infection in humans. Science, 328, 745–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dufourmantel, N. , Pelissier, B. , Garcon, F. , Peltier, G. , Ferullo, J.M. and Tissot, G. (2004) Generation of fertile transplastomic soybean. Plant Mol. Biol. 55, 479–489. [DOI] [PubMed] [Google Scholar]
- Etemad, B. , Batra, G. , Raut, R. , Dahiya, S. , Khanam, S. , Swaminathan, S. and Khanna, N. (2008) An envelope domain III‐based chimeric antigen produced in Pichia pastoris elicits neutralizing antibodies against all four dengue virus serotypes. Am. J. Trop. Med. Hyg. 79, 353–363. [PubMed] [Google Scholar]
- Ghosh, A. and Dar, L. (2015) Dengue vaccines: challenges, development, current status and prospects. Indian J. Med. Microbiol. 33, 3–15. [DOI] [PubMed] [Google Scholar]
- Gottschamel, J. and Lössl, A. (2016) Chloroplast‐based expression of recombinant proteins by gateway® cloning technology. Methods Mol. Biol. 1385, 3–27. [DOI] [PubMed] [Google Scholar]
- Gottschamel, J. , Waheed, M.T. , Clarke, J.L. and Lossl, A.G. (2013) A novel chloroplast transformation vector compatible with the Gateway® recombination cloning technology. Transgenic Res. 22, 1273–1278. [DOI] [PubMed] [Google Scholar]
- Gottschamel, J. , Lössl, A. , Ruf, S. , Wang, Y. , Skaugen, M. , Bock, R. and Clarke, J.L. (2016) Production of dengue virus envelope protein domain III‐based antigens in tobacco chloroplasts using inducible and constitutive expression systems. Plant Mol. Biol. 91, 497–512. [DOI] [PubMed] [Google Scholar]
- Guzman, M.G. , Hermida, L. , Bernardo, L. , Ramirez, R. and Guillén, G. (2010) Domain III of the envelope protein as a dengue vaccine target. Expert Rev. Vaccines, 9, 137–147. [DOI] [PubMed] [Google Scholar]
- Halstead, S.B. (1988) Pathogenesis of dengue: challenges to molecular biology. Science 239, 476–481. [DOI] [PubMed] [Google Scholar]
- Halstead, S.B. (2003) Neutralization and antibody‐dependent enhancement of dengue viruses. Adv. Virus Res. 60, 421–467. [DOI] [PubMed] [Google Scholar]
- Herz, S. , Füßl, M. , Steiger, S. and Koop, H.U. (2005) Development of novel types of plastid transformation vectors and evaluation of factors controlling expression. Transgenic Res. 14, 969–982. [DOI] [PubMed] [Google Scholar]
- Herzog, R.W. , Nichols, T.C. , Su, J. , Zhang, B. , Sherman, A. , Merricks, E.P. , Raymer, R. et al. (2017) Oral tolerance induction in hemophilia B dogs fed with transplastomic lettuce. Mol. Ther. 25, 512–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hombach, J. , Barrett, A.D. , Cardosa, M.J. , Deubel, V. , Guzman, M. , Kurane, I. , Roehrig, J.T. et al. (2005) Review on flavivirus vaccine development. Proceedings of a meeting jointly organised by the World Health Organization and the Thai Ministry of Public Health, 26‐27 April 2004, Bangkok, Thailand. Vaccine, 23, 2689–2695. [DOI] [PubMed] [Google Scholar]
- Ichikawa, Y. , Tamoi, M. , Sakuyama, H. , Maruta, T. , Ashida, H. , Yokota, A. and Shigeoka, S. (2010) Generation of transplastomic lettuce with enhanced growth and high yield. GM Crops, 1, 322–326. [DOI] [PubMed] [Google Scholar]
- Ivy, J. , Nakano, E. and Clements, D. (2000) Subunit immunogenic composition against dengue infection. In: US6165477.
- Kahlau, S. and Bock, R. (2008) Plastid transcriptomics and translatomics of tomato fruit development and chloroplast‐to‐chromoplast differentiation: chromoplast gene expression largely serves the production of a single protein. Plant Cell, 20, 856–874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanagaraj, A.P. , Verma, D. and Daniell, H. (2011) Expression of dengue‐3 premembrane and envelope polyprotein in lettuce chloroplasts. Plant Mol. Biol. 76, 323–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanamoto, H. , Yamashita, A. , Asao, H. , Okumura, S. , Takase, H. , Hattori, M. , Yokota, A. et al. (2006) Efficient and stable transformation of Lactuca sativa L. cv. Cisco (lettuce) plastids. Transgenic Res. 15, 205–217. [DOI] [PubMed] [Google Scholar]
- Karimi, M. , Inzé, D. and Depicker, A. (2002) GATEWAY vectors for Agrobacterium‐mediated plant transformation. Trends Plant Sci. 7, 193–195. [DOI] [PubMed] [Google Scholar]
- Kumar, S. , Dhingra, A. and Daniell, H. (2004) Plastid‐expressed betaine aldehyde dehydrogenase gene in carrot cultured cells, roots, and leaves confers enhanced salt tolerance. Plant Physiol. 136, 2843–2854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurane, I. and Ennis, F.E. (1992) Immunity and immunopathology in dengue virus infections. Semin. Immunol. 4, 121–127. [PubMed] [Google Scholar]
- Kwon, K.C. and Daniell, H. (2016) Oral delivery of protein drugs bioencapsulated in plant cells. Mol. Ther. 24, 1342–1350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon, K.C. , Verma, D. , Singh, N.D. , Herzog, R. and Daniell, H. (2013) Oral delivery of human biopharmaceuticals, autoantigens and vaccine antigens bioencapsulated in plant cells. Adv. Drug Deliv. Rev. 65, 782–799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, G. , Na, Y.J. , Yang, B.G. , Choi, J.P. , Seo, Y.B. , Hong, C.P. , Yun, C.H. et al. (2015) Oral immunization of haemaggulutinin H5 expressed in plant endoplasmic reticulum with adjuvant saponin protects mice against highly pathogenic avian influenza A virus infection. Plant Biotechnol. J. 13, 62–72. [DOI] [PubMed] [Google Scholar]
- Lindh, I. , Bråve, A. , Hallengärd, D. , Hadad, R. , Kalbina, I. , Strid, A. and Andersson, S. (2014) Oral delivery of plant‐derived HIV‐1 p24 antigen in low doses shows a superior priming effect in mice compared to high doses. Vaccine, 32, 2288–2293. [DOI] [PubMed] [Google Scholar]
- Liu, C.W. , Lin, C.C. , Chen, J.J. and Tseng, M.J. (2007) Stable chloroplast transformation in cabbage (Brassica oleracea L. var. capitata L.) by particle bombardment. Plant Cell Rep. 26, 1733–1744. [DOI] [PubMed] [Google Scholar]
- Maldaner, F.R. , Aragao, F.J. , dos Santos, F.B. , Franco, O.L. , da Rocha Queiroz Lima, M. , de Oliveira Resende, R. , Vasques, R.M. et al. (2013) Dengue virus tetra‐epitope peptide expressed in lettuce chloroplasts for potential use in dengue diagnosis. Appl. Microbiol. Biotechnol. 97, 5721–5729. [DOI] [PubMed] [Google Scholar]
- Maliga, P. and Bock, R. (2011) Plastid biotechnology: food, fuel, and medicine for the 21st century. Plant Physiol. 155, 1501–1510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martínez, C.A. , Topal, E. , Giulietti, A.M. , Talou, J.R. and Mason, H. (2010) Exploring different strategies to express Dengue virus envelope protein in a plant system. Biotechnol. Lett. 32, 867–875. [DOI] [PubMed] [Google Scholar]
- McDonald, W. , Powell, T. , Price, A. and Becker, R. (2009) Compositions of dengue viral proteins and methods of use. In: WO2009128949.
- Megret, F. , Hugnot, J.P. , Falconar, A. , Gentry, M.K. , Morens, D.M. , Murray, J.M. , Schlesinger, J.J. et al. (1992) Use of recombinant fusion proteins and monoclonal antibodies to define linear and discontinuous antigenic sites on the dengue virus envelope glycoprotein. Virology, 187, 480–491. [DOI] [PubMed] [Google Scholar]
- Minekus, M. , Alminger, M. , Alvito, P. , Ballance, S. , Bohn, T. , Bourlieu, C. , Carrière, F. et al. (2014) A standardised static in vitro digestion method suitable for food ‐ an international consensus. Food Funct. 5, 1113–1124. [DOI] [PubMed] [Google Scholar]
- Moravec, T. , Schmidt, M.A. , Herman, E.M. and Woodford‐Thomas, T. (2007) Production of Escherichia coli heat labile toxin (LT) B subunit in soybean seed and analysis of its immunogenicity as an oral vaccine. Vaccine, 25, 1647–1657. [DOI] [PubMed] [Google Scholar]
- Murashige, T. and Skoog, F. (1962) A revised medium for rapid growth and bio‐assays with tobacco tissue cultures. Physiol. Plant. 15, 473–497. [Google Scholar]
- Murray, M.G. and Thompson, W.F. (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 8, 4321–4325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nugent, G.D. , Coyne, S. , Nguyen, T.T. , Kavanagh, T.A. and Dix, P.J. (2006) Nuclear and plastid transformation of Brassica oleracea var. botrytis (cauliflower) using PEG‐mediated uptake of DNA into protoplasts. Plant Sci. 170, 135–142. [Google Scholar]
- Pang, E.L. and Loh, H.S. (2017) Towards development of a universal dengue vaccine ‐ How close are we? Asian Pac. J. Trop. Med. 10, 220–228. [DOI] [PubMed] [Google Scholar]
- Peyret, H. and Lomonossoff, G.P. (2015) When plant virology met Agrobacterium: the rise of the deconstructed clones. Plant Biotechnol. J. 13, 1121–1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rigano, M.M. , Manna, C. , Giulini, A. , Pedrazzini, E. , Capobianchi, M. , Castilletti, C. , Di Caro, A. et al. (2009) Transgenic chloroplasts are efficient sites for high‐yield production of the vaccinia virus envelope protein A27L in plant cells. Plant Biotechnol. J. 7, 577–591. [DOI] [PubMed] [Google Scholar]
- Ruf, S. , Hermann, M. , Berger, I.J. , Carrer, H. and Bock, R. (2001) Stable genetic transformation of tomato plastids and expression of a foreign protein in fruit. Nat. Biotechnol. 19, 870–875. [DOI] [PubMed] [Google Scholar]
- Ruhlman, T. , Ahangari, R. , Devine, A. , Samsam, M. and Daniell, H. (2007) Expression of cholera toxin B‐proinsulin fusion protein in lettuce and tobacco chloroplasts–oral administration protects against development of insulitis in non‐obese diabetic mice. Plant Biotechnol. J. 5, 495–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saejung, W. , Fujiyama, K. , Takasaki, T. , Ito, M. , Hori, K. , Malasit, P. , Watanabe, Y. et al. (2007) Production of dengue 2 envelope domain III in plant using TMV‐based vector system. Vaccine, 25, 6646–6654. [DOI] [PubMed] [Google Scholar]
- Sidorov, V.A. , Kasten, D. , Pang, S.Z. , Hajdukiewicz, P.T. , Staub, J.M. and Nehra, N.S. (1999) Technical Advance: stable chloroplast transformation in potato: use of green fluorescent protein as a plastid marker. Plant J. 19, 209–216. [DOI] [PubMed] [Google Scholar]
- Simmons, M. , Hayes, C. and Porter, K. (1999) Recombinant dengue virus envelop protein/maltose‐binding protein antigens and subunit vaccine compositions containing said antigens. In: US5895651.
- Simmons, C.P. , Farrar, J.J. , Nguyen, V. and Wills, B. (2012) Dengue. N. Engl. J. Med. 366, 1423–1432. [DOI] [PubMed] [Google Scholar]
- Singh, A.K. , Verma, S.S. and Bansal, K.C. (2010) Plastid transformation in eggplant (Solanum melongena L.). Transgenic Res. 19, 113–119. [DOI] [PubMed] [Google Scholar]
- Srivastava, A. , Putnak, J. , Hoke, C. and Warren, R. (2000) Recombinant vaccine made in E.coli against dengue virus. In: US6117640.
- Staub, J.M. and Maliga, P. (1993) Accumulation of D1 polypeptide in tobacco plastids is regulated via the untranslated region of the psbA mRNA. EMBO J. 12, 601–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streatfield, S.J. (2006) Mucosal immunization using recombinant plant‐based oral vaccines. Methods, 38, 150–157. [DOI] [PubMed] [Google Scholar]
- Su, J. , Zhu, L. , Sherman, A. , Wang, X. , Lin, S. , Kamesh, A. , Norikane, J.H. et al. (2015) Low cost industrial production of coagulation factor IX bioencapsulated in lettuce cells for oral tolerance induction in hemophilia B. Biomaterials, 70, 84–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svab, Z. and Maliga, P. (1993) High‐frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proc. Natl Acad. Sci. U S A, 90, 913–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valkov, V.T. , Scotti, N. , Kahlau, S. , Maclean, D. , Grillo, S. , Gray, J.C. , Bock, R. et al. (2009) Genome‐wide analysis of plastid gene expression in potato leaf chloroplasts and tuber amyloplasts: transcriptional and posttranscriptional control. Plant Physiol. 150, 2030–2044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vannice, K.S. , Roehrig, J.T. and Hombach, J. (2015) Next generation dengue vaccines: a review of the preclinical development pipeline. Vaccine, 33, 7091–7099. [DOI] [PubMed] [Google Scholar]
- Vannice, K.S. , Durbin, A. and Hombach, J. (2016) Status of vaccine research and development of vaccines for dengue. Vaccine, 34, 2934–2938. [DOI] [PubMed] [Google Scholar]
- Verma, D. , Samson, N.P. , Koya, V. and Daniell, H. (2008) A protocol for expression of foreign genes in chloroplasts. Nat. Protoc. 3, 739–758. [DOI] [PubMed] [Google Scholar]
- Verma, D. , Kanagaraj, A. , Jin, S. , Singh, N.D. , Kolattukudy, P.E. and Daniell, H. (2010) Chloroplast‐derived enzyme cocktails hydrolyse lignocellulosic biomass and release fermentable sugars. Plant Biotechnol. J. 8, 332–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, W. , Yu, B. , Lin, X.D. , Kong, D.G. , Wang, J. , Tian, J.H. , Li, M.H. et al. (2015) Reemergence and autochthonous transmission of dengue virus, Eastern China, 2014. Emerg. Infect. Dis. 21, 1670–1673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weaver, S.C. and Vasilakis, N. (2009) Molecular evolution of dengue viruses: contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect Genet. Evol. 9, 523–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO . (2009) World Health Organization and the Special Program for Research and Training in Tropical Diseases: Dengue: guidelines for diagnosis, treatment, prevention and control. Geneva, Switzerland. [PubMed] [Google Scholar]
- WHO (2015) Factsheet on Dengue and Dengue haemorrhagic fever. http://www.who.int/mediacentre/factsheets/fs117/en/. World Health Organization.
- Wichmann, O. , Vannice, K. , Asturias, E.J. , de Albuquerque Luna, E.J. , Longini, I. , Lopez, A.L. , Smith, P.G. et al. (2017) Live‐attenuated tetravalent dengue vaccines: the needs and challenges of post‐licensure evaluation of vaccine safety and effectiveness. Vaccine, 35, 5535–5542. [DOI] [PubMed] [Google Scholar]
- Xiao, Y. and Daniell, H. (2017) Long‐term evaluation of mucosal and systemic immunity and protection conferred by different polio booster vaccines. Vaccine, 35, 5418–5425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao, Y. , Kwon, K.C. , Hoffman, B.E. , Kamesh, A. , Jones, N.T. , Herzog, R.W. and Daniell, H. (2016) Low cost delivery of proteins bioencapsulated in plant cells to human non‐immune or immune modulatory cells. Biomaterials, 80, 68–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong, Y. and Chen, Q. (2014) Epidemiology of dengue fever in China since 1978. Nan Fang Yi Ke Da Xue Xue Bao, 34, 1822–1825. [PubMed] [Google Scholar]
- Xu, G. , Dong, H. , Shi, N. , Liu, S. , Zhou, A. , Cheng, Z. , Chen, G. et al. (2007) An outbreak of dengue virus serotype 1 infection in Cixi, Ningbo, People's Republic of China, 2004, associated with a traveler from Thailand and high density of Aedes albopictus. Am. J. Trop. Med. Hyg. 76, 1182–1188. [PubMed] [Google Scholar]
- Ye, G.N. , Hajdukiewicz, P.T. , Broyles, D. , Rodriguez, D. , Xu, C.W. , Nehra, N. and Staub, J.M. (2001) Plastid‐expressed 5‐enolpyruvylshikimate‐3‐phosphate synthase genes provide high level glyphosate tolerance in tobacco. Plant J. 25, 261–270. [DOI] [PubMed] [Google Scholar]
- Zhang, F.C. , Zhao, H. , Li, L.H. , Jiang, T. , Hong, W.X. , Wang, J. , Zhao, L.Z. et al. (2014) Severe dengue outbreak in Yunnan, China, 2013. Int. J. Infect. Dis. 27, 4–6. [DOI] [PubMed] [Google Scholar]
- Zhang, B. , Shanmugaraj, B. and Daniell, H. (2017) Expression and functional evaluation of biopharmaceuticals made in plant chloroplasts. Curr. Opin. Chem. Biol. 38, 17–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou, F. , Badillo‐Corona, J.A. , Karcher, D. , Gonzalez‐Rabade, N. , Piepenburg, K. , Borchers, A.M.I. , Maloney, A.P. et al. (2008) High‐level expression of human immunodeficiency virus antigens from the tobacco and tomato plastid genomes. Plant Biotechnol. J. 6, 897–913. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 Quantification of EDIII‐1‐4 accumulation in transplastomic lettuce plants.