

Abstract
Conflicting evidence exists concerning the activity of NMDA receptors (NMDARs) in cerebellar Purkinje cells and their possible functions. To investigate the activity of NMDARs, we used whole-cell recording on immunocytochemically identified Purkinje cells in primary culture. In addition, we used mice with a disrupted NMDAR1 gene that lack functional NMDARs (NR1−/−) to assess the physiological role of NMDARs. In cultures from normal mice, NMDA-mediated currents were detected in all identified Purkinje cells at 4 d in vitro (div). After 14 d, however, NMDA responses were reduced in amplitude, whereas the responses to kainate and glutamate increased steadily in amplitude. In addition, the NMDA-induced current displayed a pronounced desensitization at these later stages; peak current declined to zero during steady application of NMDA. At 7 div, the number of surviving Purkinje cells was less in cultures treated with NMDA antagonists, and their survival was dose-dependent. Purkinje cell survival was correspondingly poorer in cultures from the NR1−/− mice than in wild-type controls, suggesting that NMDAR activity enhances the survival of Purkinje cells in vitro. The addition of moderate doses of NMDA promoted the survival of wild-type Purkinje cells in the presence of tetrodotoxin. Feeder layers of cerebellar granule cells derived from wild-type or NR1−/− mice promoted survival of Purkinje cells to a similar degree, suggesting that the NMDAR in Purkinje cells, but not in other cells, is directly involved in Purkinje cell viability. The results demonstrate that NMDARs transiently produce membrane current in Purkinje cells and may serve as one of the epigenetic factors that support the survival of Purkinje cells in vitro.
Keywords: Purkinje cells, cerebellum, NMDA receptors, excitatory amino acids, dissociated cell culture, patch clamp
NMDA receptors (NMDARs) constitute cation channels gated by the excitatory transmitter glutamate in central synapses. The NMDAR has the unique properties of high Ca2+permeability and voltage-dependent blockade by Mg2+ (MacDermott et al., 1986; Mayer and Westbrook, 1987; Ascher and Nowak, 1988) and has been implicated in many developmental, pathological, and physiological processes (for reviews, see McDonald and Johnston, 1990; Bliss and Collingridge, 1993;Malenka and Nicoll, 1993). NMDARs are formed by two main classes of subunits, NMDAR1 and NMDAR2, encoded by distinct genes (for reviews, see Nakanishi, 1992; Seeburg, 1993). The presence of NMDAR1 has been shown to be necessary for the basic functional expression of the NMDAR (Moriyoshi et al., 1991; Forrest et al., 1994).
In the cerebellum, both parallel and climbing fiber inputs to Purkinje cells and mossy fiber inputs to granule cells are mediated, at least partly, by glutamate (Ito, 1984; Kaneko et al., 1989). Analysis by immunostaining (Petralia et al., 1994) and in situhybridization of mRNA (Moriyoshi et al., 1991; Monyer et al., 1992,1994; Akazawa et al., 1994; Watanabe et al., 1994) has shown that NMDAR1s are most abundant in Purkinje cells and moderately abundant in granule cells in the cerebellar cortex. There is a great deal of contradictory evidence, however, regarding the presence of functional NMDARs, i.e., receptors that open ion channels, in Purkinje cells. In neurons acutely isolated from very young animals or in neurons cultured for a short time from embryos, there is a prominent NMDA-induced current (Hockberger et al., 1989; Yuzaki et al., 1990; Sorimachi et al., 1991; Rosenmund et al., 1992; Yuzaki and Mikoshiba, 1992). In contrast, in cells maintained for longer periods in dispersed, explant, or slice cultures, there is no response (Llano et al., 1988; Joels et al., 1989; Audinat et al., 1990). Unfortunately, it has not yet been possible to isolate mature Purkinje neurons (Rosenmund et al., 1992). Some of the conflicting results can be accommodated by the hypothesis that there is a developmental downregulation of the NMDA response. Other results, however, such as those obtained by in vivorecording or in acutely isolated slice preparations, have indicated either the presence (Sekiguchi et al., 1987; Lee et al., 1988) of an NMDA current in adult animals or the absence of direct NMDA responses in neonatal rats (Farrant and Cull-Candy, 1991; Llano et al., 1991). Here, we present an analysis of the developmental changes that occur in the NMDAR-mediated currents in Purkinje cells that could explain many of these apparent discrepancies.
Analysis of receptor activity in dispersed cultures has several advantages over in vivo or slices studies. Dispersed cultures allow for better control of the external environment and easy access to neurons, thus facilitating the direct recording of drug responses. Additionally, the use of culture techniques has allowed us to follow the development of neurons from NR1−/− mice that normally die on the day of birth (Forrest et al., 1994). A major concern with dispersed cultures is the unambiguous identification of cell types (Audinat et al., 1990; Farrant and Cull-Candy, 1991). Although many studies have used immunocytochemical staining to identify Purkinje cells, the staining was not performed on the actual cells recorded. Here, we routinely marked the neurons studied with dextran-conjugated rhodamine introduced by the patch-clamp electrode, and after the electrophysiological recordings, we verified the type of neurons examined by immunocytochemical staining. Thus, we have shown responses to NMDA in identified Purkinje cells during development in culture. In addition, by studying the differentiation of Purkinje cells derived from NR1−/− mice, we suggest a potential physiological function for the transiently expressed NMDAR-mediated current.
MATERIALS AND METHODS
Culture. Primary dissociated cultures were prepared from mice embryos at 18 d postcoitum (E18) or from neonatal mice (P0), as described (Yuzaki and Mikoshiba, 1992; Forrest et al., 1994). Because this strain of mice usually give birth at E19, the difference in the developmental stage of mice was within 1 d. In addition, no differences were observed in the data obtained from cultures from mice at E18 and P0. Thus, all data were combined unless stated otherwise. Briefly, cerebella were treated with 0.1% trypsin and 0.05% DNaseI in Ca2+/Mg2+-free HBSS for 5 min at room temperature. The cells were washed with culture medium containing 10% fetal calf serum (Life Technologies, Gaithersburg, MD) and dissociated by passing through a fine-tipped pipette in Ca2+-free HBSS containing 0.05% DNaseI and 12 mm MgSO4. Dispersed cells were plated on 18-mm-round sheets of Aclar (Pro Plastics) coated with poly-l-lysine (Sigma, St. Louis, MO) at a density of 1.6 × 105cells/cm2 in Fischer’s medium (Fischer, 1982) supplemented with 10% fetal calf serum. Cells were kept in this media for 1 d, because our preliminary data suggested that the survival of neurons was generally improved by ∼10–20% by incubating cells in the medium containing serum for 1 d after plating. One day after plating, the medium was changed to serum-free Fischer’s medium, and excitatory amino acid agonists and antagonists were added at this time, unless stated otherwise. The cultures were maintained in a humidified atmosphere of 10% CO2 in air at 37°C. Half of the medium was changed weekly with the readdition of drugs.
Cerebellar cultures from NMDAR mutant mice were prepared in the same way, except that individual cerebella from P0 pups were treated with 0.05% trypsin. The tail of each pup was saved for genotype determination.
In some experiments (see Figs. 7, 8), granule-cell feeder layers were prepared in the same way as above except that they were plated at a density of 0.8 × 105cells/cm2. After 7 div, there were few Purkinje cells in the feeder layer, as confirmed by calbindin-D28k staining (see below). New cerebellar cultures were plated on top of the feeder layer. We did not use high-potassium medium for granule cell culture, which is often used to promote the maturation and survival of granule cells, but used regular Fischer’s medium, because the number of Purkinje cells was greatly reduced in high potassium (25 mm) medium (see Fig. 3D).
Fig. 7.

The influence of the NR1 granule-cell feeder genotypes on the survival of Purkinje cells. A, Cerebellar cells were prepared from individual NR1−/−, −/+, or +/+ mice at P0 and maintained for 7 d to be used as feeder cells. Cerebellar cells from individual NR1−/−, −/+, or +/+ mice were plated on top of the feeder cells and cultured for another 7 d. After determining each genotype, data from the same combination of genotypes were combined. Data from the heterozygotes (−/+) and wild-type mice (+/+) were combined to increase the n value. Data are expressed as a percentage of the number of Purkinje cells in −/+ and +/+ mice without feeder cells. Regardless of the presence or genotype of granule cell feeders, there were significant differences in the number of surviving Purkinje cells between the −/− and −/+ or +/+ culture (p < 0.05; indicated by asterisk);n = 3–6 cultures.
Fig. 8.

The influence of wild-type granule cell feeders on the survival of Purkinje cells. A, Granule cells were prepared from P5–P6 wild-type mice and maintained for 7 d to be used as feeder cells. Cerebellar cells from NR1−/−, −/+, or +/+ mice were plated on top of the feeder cells and maintained in the absence (Ctrl) or presence (+APV) ofd,l-APV (200 μm) for 6 d. B, After determining each genotype, data from the same genotype were combined and expressed as a percentage of the number of Purkinje cells in wild-type (+/+) cultures without feeder cells. There were significant differences between the −/− and −/+ or +/+ cultures (p < 0.05; indicated byasterisk) regardless of the presence or absence of granule cell feeders. The addition of APV abolished the differences;n = 5–14 cultures.
Fig. 3.

Effects of NMDAR antagonists on survival of Purkinje cells in vitro. A, Cerebellar cells were grown in the absence (left) and presence (right) of 200 μmd,l-APV for 6 d. To visualize the Purkinje cell subpopulation, cells were stained for calbindin-D28k by diaminobenzidine as a final substrate for avidin–biotin–peroxidase (Vectastain) and counterstained by hematoxylin. Scale bar, 40 μm. B, Dose-dependent effect of APV and partial recovery by addition of NMDA. Cells were treated with APV or NMDA for 6 d and stained for calbindin-D28k. The number of surviving Purkinje cells in cultures treated with 100 or 300 μmd,l-APV was significantly less than that in controls (p < 0.01; indicated by double asterisks). The viability of Purkinje cells in cultures treated with 100 μmd,l-APV was significantly increased by the addition of 300 μm NMDA (p < 0.05; indicated by asterisk);n = 6–12 cultures. C, Effect of other NMDAR antagonists on survival of Purkinje cells. Cells were treated with drugs as indicated. MK-801 (1 μm) or 7-Cl-Kyn (100 μm) significantly reduced the surviving Purkinje cells (p < 0.01; indicated bydouble asterisks); n = 6–8 cultures.D, Effect of non-NMDAR agonist/antagonist and high potassium on survival of Purkinje cells. Cells were treated with CNQX (10 μm), AMPA (1 μm), or high potassium (25 mm). AMPA or high-potassium solution significantly decreased the surviving Purkinje cells (p < 0.01; indicated by double asterisks); n = 5–6 cultures.
Immunocytochemistry and survival assay. Calbindin-D28k protein is specifically expressed in Purkinje cells as early as E14 in rats (Enderlin et al., 1987). As for mice, no data are available for the expression in embryos, but it is expressed at least from P0 (Iacopino et al., 1990). Although a minor subpopulation of Purkinje cells do not express calbindin-D28k until P4, thereafter all Purkinje cells express the protein (Wassef et al., 1985). Thus, we identified Purkinje cells in culture by staining for calbindin-D28k, as reported previously (Schilling et al., 1991; Yuzaki and Mikoshiba, 1992). Briefly, cells were fixed with 4% paraformaldehyde in 0.1 m phosphate buffer for 10 min, permeabilized with 0.01% Triton X-100 in PBS for 10 min, and blocked with 1 mg/ml bovine serum albumin for 60 min. Cells were incubated with a monoclonal antibody against calbindin-D28k (Sigma) at 1:400 dilution for 60 min. In some studies, antibodies against glial fibrillary acidic protein (GFAP) and neuron-specific enolase (NSE) were used. Antibody binding was visualized by diaminobenzidine as a final substrate for avidin–biotin–peroxidase (Vectastain) or by fluorescein isothiocyanate-labeled secondary antibody.
Calbindin-D28k-positive cells were counted in 10 fields per culture well, representing ∼45 mm2 or ∼22% of the surface area of an individual well. Results were confirmed by recounting of random samples. Data were expressed as percentages of cell numbers in parallel control wells. Figures present mean data from two or more independent experiments, each performed at least in triplicate. Statistical analysis was performed using the Student’st test.
Genotype determination of mice. DNA samples from NMDAR1 homozygous mutants (NR1−/−), heterozygotes (NR1−/+), or wild-type (NR1+/+) littermates were prepared from tail clips, and genotypes were determined either by Southern blotting, as previously described (Forrest et al., 1994), or by PCR. For PCR, we used a common antisense primer (5′-CAG CCT GCA CAC TTT AGG TCA CAT TG-3′) and two sense primers: one for the wild-type NR1 gene (5′-CCA ACG CCA TAC AGA AGA TGG CCC TGT-3′) and one for the neosubstitution (5′-GTG CCA GCG GGG CTG CTA AAG-3′). Reaction included 10% DMSO. Denaturation was at 94°C/30 sec, annealing at 65°C/30 sec, and extension at 72°C/30 sec for 35 cycles in a GeneAmp9600 thermocycler (Perkin-Elmer Cetus, Norwalk, CT). Two bands corresponding to the wild-type allele (950 bp) and disrupted allele (500 bp) were identified by agarose gel electrophoresis and staining with ethidium bromide. In some experiments, both PCR and Southern blotting were performed to verify results.
Electrophysiology. Membrane currents were measured using standard whole-cell patch-clamp methods with an EPC-7 amplifier (List, Campbell, CA). Electrodes were pulled from 1.5 mm glass (World Precision Instruments, Sarasota, FL) on a two-stage vertical puller (Narishige PP-83, Narishige, Greenvale, NY). Saline composition in the electrodes was 130 mm potassium gluconate, 10 mm KCl, 1 mmCaCl2, 2 mmMgCl2, 11 mm EGTA, 10 mm Hepes, 2 mmNa2-ATP (pH was adjusted to 7.3 with KOH). Dextran-conjugated rhodamine (Molecular Probes, Eugene, OR) was included at 0.625 mg/ml for later identification of the cells studied. The liquid junction potential was approximately −10 mV. Electrodes filled with the recording saline had resistances of ∼8–9 MΩ. Series resistances were 15–20 MΩ and partially compensated by 40–50%.
The extracellular recording solution contained 150 mm NaCl, 5 mm KCl, 2 mm CaCl2, 10 mm Hepes, and 20 mm glucose (pH was adjusted to 7.3 with NaOH). Tetrodotoxin (TTX) (1 μm) and picrotoxin (100 μm) were included in the solution to block spontaneous electrical activity and GABA channels. All drugs were dissolved in the recording solution. Glycine (5 μm) was added to and Mg2+was omitted from the recording solution to maximize the responses to NMDA and l-glutamate. The chamber was perfused continuously (1–2 ml/min) at room temperature.
Drugs were applied by the “Y-tube” method (Murase et al., 1990), improved to be controlled by a computer. Polyethylene tubing (0.92 mm diameter) was bent in a V-shape, and fine polyethylene tubing (0.1 mm diameter, 10 mm long) was connected to the curved end. One end of the V-shaped tubing was connected to the common outlet of six solenoid manifold valves (General Valve). Either control or one of the five test solutions was selected by opening one of the valves. Another end was led to the vacuum through a solenoid drainage valve (Pharmacia, Piscataway, NJ). When the drainage valve was opened, the solution in the Y-tube was replaced by new solution, and when closed, solution was released from the outlet of the Y-tube under gravity. The tip of the Y-tube was placed ∼300–400 μm from the neurons, where they were continuously exposed to the fast flow of control solution. When the shift of liquid junction potential at the tip of the electrode was measured by changing the external solution concentration from 1× to 0.1×, the time constant of solution exchange was 8–9 msec. A time constant of solution exchange surrounding a neuron was estimated to be ∼15–20 msec, judging from a rise time of kainate- and glutamate-induced currents.
Currents were filtered at 1 kHz and digitized at 3 kHz. Data acquisition and control of the inlet and drainage solenoid valves were achieved by a custom software using AxoBasic library (Axon Instruments, Foster City, CA) and I/O board (Digidata-1200, Axon Instruments).
Drugs. NMDA, kainate, l-glutamate, glycine, d,l-2-amino-5-phosphonovalerate (APV), TTX, and picrotoxin were from Sigma. 7-Cl-kynurenate (7-Cl-Kyn), 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX), (R,S)-α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), and (5R,10S)-(+)-5-methyl-10,11-dihydro-5H- dibenzo[a,d]cyclohepten-5,10-imine hydrogen maleate (MK-801) were obtained from Research Biochemicals International (Natick, MA).
RESULTS
Transient NMDA response in Purkinje cells in vitro
The presence of NMDAR-mediated currents was examined by the patch-clamp technique during differentiation of Purkinje cells in cultures derived from mice at E18. As shown in Figure 1(left), after ∼2 weeks in vitro, Purkinje cells began to show the typical morphology, i.e., large globular cell body and thick apical dendrites with many branches, which corresponds to stage 4 Purkinje cells in vivo (Baptista et al., 1994). The cells that met the visual morphological criteria were always confirmed to be Purkinje cells by calbindin-D28k staining. At only 4 div, however, Purkinje cells did not show a characteristic morphology. Typically, they had few long processes (Fig. 1, left), which corresponds to stage 1 Purkinje cells in vivo (Baptista et al., 1994). Thus, it was often difficult to identify Purkinje cells only by morphology, i.e., 30–40% of the candidate neurons were negative for calbindin-D28k staining (cf. Audinat et al., 1990; Farrant and Cull-Candy, 1991). Therefore, we routinely performed calbindin-D28k staining for unambiguous identification of candidate Purkinje cells at any stage in culture. Neurons from which recordings were made were identified by dextran-conjugated rhodamine, which was included in the patch pipette. Figure 1 (left) shows examples of Purkinje cells stained with calbindin-D28k, whereas Figure 1 (right) shows the particular neuron studied, co-stained with rhodamine in the same field of view as in the pictures on the left.
Fig. 1.

Identification of recorded cells as Purkinje cells and their development in vitro. Three fields of Purkinje cells from three cultures of different days in vitro(div). Purkinje cells were stained for calbindin-D28k, a specific marker of Purkinje cells, and visualized with fluorescein (left). The particular neurons used for electrophysiological recordings were identified by dextran-conjugated rhodamine, which were included in whole-cell patch pipettes (right). Cells stained positively for both fluorescein and rhodamine were thus verified as recorded Purkinje cells. Scale bar, 40 μm.
Agonists were applied rapidly for 4 sec to Purkinje cells held at −60 mV. NMDA (100 μm), kainate (10 μm), and glutamate (100 μm) induced inward currents in all immunocytochemically identified Purkinje cells as early as 4 div (n = 8). The inward current evoked by NMDA was smaller than current evoked by kainate and showed the large increase in open channel noise typical of NMDA-induced current (Fig.2A, upper traces). A similar increase in open channel noise was observed in glutamate-induced currents. After 14 div, the NMDA-induced peak current became smaller, whereas kainate- and glutamate-induced currents increased (Fig.2A,B).
Fig. 2.

Changes in the responsiveness of Purkinje cells to excitatory amino acids. A, Application of NMDA (100 μm), kainate (10 μm), and glutamate (100 μm) by Y-tube method (see Materials and Methods) caused inward currents in immunocytochemically identified Purkinje cells. The upper trace was from a Purkinje cell at 7 div and the lower trace at 14 div. Holding potential was −60 mV. The current calibration of 125 pA applies to NMDA-induced currents and that of 250 pA applies to kainate- and glutamate-induced currents. B, Summary of the changes in current amplitudes in response to excitatory amino acids during maturation. C, Summary of changes in desensitization of NMDA-induced current during maturation. As an index of desensitization, the current amplitude 1 sec after the peak current was shown as a percentage of its peak amplitude. Each point in B andC represents an average of 6–11 cells (mean ± SE).
NMDA-induced currents also became very transient with increasing age of the cultures (Fig. 2A, lower traces). Because NMDAR is permeable to Ca2+, NMDAR activation could induce a Ca2+-dependent Cl− current that could contribute to the decay phase of the NMDA-induced current (Leonard and Kelso, 1990). This contamination was minimal in our recordings, however, because the equilibrium potential for Cl− was approximately −58 mV, which is close to the holding potential. The open channel noise level of glutamate-induced current also decreased, even though the amplitude increased. Thus, the decay phase of NMDA-induced current may be attributable to glycine-insensitive desensitization (Sather et al., 1990). As an index of desensitization, the current amplitude 1 sec after the peak current was shown as a percentage of its peak amplitude (Fig. 2C).
The change in desensitization of NMDARs in Purkinje cells during maturation, in addition to the reduction of peak current, may contribute to the absence of NMDA-induced currents in mature Purkinje cells in other studies. NMDA currents in granule cells in the same cultures did not decline with age in culture (data not shown). These results suggest that NMDARs are transiently functional in Purkinje cells, becoming almost inactive after 14 div.
Function of transiently active NMDARs in vitro
To determine a possible role for the transiently active NMDARs, we investigated the effect of blockade of NMDARs on the morphology and survival of Purkinje cells in vitro.
In a control experiment, we found that most of the Purkinje cell death occurred within the first 4 d in vitro, because the number of calbindin-D28k-positive cells at 4 div and at 7 div were 25 ± 5% (n = 5) and 20 ± 4% (n = 6), respectively, of that determined after 1 hrin vitro. This suggested a rationale for assessing the effect of drugs on the survival of Purkinje cells at 7 div. Thus, cells were grown in the presence of APV, a specific competitive NMDAR antagonist, from 1 div, when we changed the medium to serum-free medium (see Materials and Methods), and stained for calbindin-D28k at 7 div.
APV had little effect on the morphology of Purkinje cells but decreased the number of surviving Purkinje cells after 6 div (Fig.3A,B). The numbers of surviving Purkinje cells in control and d,l-APV (200 μm)-treated cultures were 78 ± 7 and 20 ± 4 (n = 6), respectively. The numbers of non-Purkinje cells (calbindin-D28k negative, NSE positive) and glial cells (calbindin-D28k negative, NSE negative) in control were 3100 ± 120 and 320 ± 14 (n = 5), respectively, and were not changed significantly by d,l-APV (200 μm) treatment (3300 ± 110 and 330 ± 11, respectively; n = 6). The number of surviving Purkinje cells was reduced by APV in a dose-dependent manner and was partially rescued by NMDA (Fig. 3B). The dose-dependent inhibitory effect of d,l-APV was fitted with a logistic equation by least-squares fit, with IC50 = 75 μm and Hill coefficient = 0.95. Although the antagonist equilibrium constant (Ki) or IC50 of APV for NMDAR in Purkinje cells is not known, the IC50 value we obtained seemed high compared with the reported value of Ki ofd,l-APV (16 μm) obtained by displacement of glutamate binding to NMDAR in the cerebellum (Buller et al., 1994). This may be attributable to the difference in the assay method. Because cells are exposed to drugs for 6 d in our assay, small residual responses, which may not be detected in fast assays such as binding and electrophysiological assays, may still have profound effects on neuronal survival. It is also possible that NMDARs in Purkinje cells actually have a lower affinity for APV, because NMDARs exhibit several unique characteristics, such as rapid desensitization and developmental changes (Fig. 2).
Other NMDA receptor antagonists 7-Cl-Kyn and MK-801 also reduced the number of surviving Purkinje cells in a dose-dependent manner (Fig.3C). In contrast, the number of Purkinje cells was increased slightly by a specific non-NMDA antagonist CNQX (10 μm) and decreased significantly by non-NMDA agonist AMPA (1 μm) or high potassium (25 mm) (Fig. 3D). Together, these results suggested a specific role for NMDAR activity in Purkinje cell survivalin vitro.
Although calbindin-D28k staining has been used widely for the identification of Purkinje cells in various culture conditions, it was theoretically possible that APV may alter the expression of calbindin-D28k protein in Purkinje cells but not the actual number of Purkinje cells. Thus, we examined APV-treated and control untreated cultures by phase-contrast microscopy to determine the percentage of all putative Purkinje cells identified by visual morphology that were calbindin-D28k-positive. Cultures were examined at 7 div when visual morphological identification was significantly more reliable than in earlier stage cultures (e.g., 4 div; see above). The percentages were 82 ± 4% and 84 ± 7% (n = 6 each) in control and APV-treated cultures, respectively, thus confirming that the numbers of calbindin-D28k-positive cells with Purkinje cell morphology did not change under APV treatment.
Next, we added APV at different culture ages to define the period of sensitivity of Purkinje cell survival to APV (Fig. 4). The effect of APV became less pronounced with later times of addition. The period of greatest sensitivity corresponds closely with the time of activity of NMDARs (Fig. 2), and the greatest loss of Purkinje cells in culture also occurred at this time. This finding suggests that the transient activity of NMDARs in Purkinje cells, but not in other types of neurons that do not show the transient activity, may support the survival of Purkinje cells in young cultures.
Fig. 4.

Sensitive period of Purkinje cells to APV. Cerebellar cells were treated with 200 μmd,l-APV for various periods in vitro, and Purkinje population was identified and counted. The number of surviving Purkinje cells was significantly less in cultures treated with APV from 1 to 7 div than in control (p < 0.01; indicated by double asterisks); n = 4–6 cultures.
NMDARs of Purkinje cells are directly involved in the survival of Purkinje cells
To exclude the possible indirect involvement of NMDARs in other cells, we conducted two studies. First, activation of NMDARs in other neurons will generate action potentials and lead to the release of neurotransmitters that may influence the survival of Purkinje cells, because neurons in dispersed culture are reported to show spontaneous synaptic activity (Ogura et al., 1987; Yuzaki et al., 1990). APV could have blocked NMDA-mediated action potentials and reduced the release of neurotransmitters. To rule out this possibility, we applied NMDA as an agonist in the presence of TTX to block the action potential-mediated release of neurotransmitters. If the activities of NMDARs in Purkinje cells are involved directly, application of NMDA should influence Purkinje cell survival regardless of the presence of TTX.
TTX (1 μm) or TTX + NMDA were applied from 1 div to 7 div. TTX itself did not have any significant effects on the survival or morphology of Purkinje cells (Fig. 5, control). The lack of effect of TTX on morphology of Purkinje cells may be attributable to the short observation period here, because it took ∼2 weeks for Purkinje cells to begin showing their typical morphologyin vitro (Fig. 1; see also Weber and Schachner, 1984;Hockberger et al., 1989; Baptista et al., 1994). Furthermore, Schilling et al. (1991) reported that TTX added to culture medium for longer than 2 weeks but not less induced morphological alternations of Purkinje cells. The addition of NMDA, in the concentration range of 10–30 μm, promoted the survival of Purkinje cells, whereas 100 μm NMDA inhibited survival (Fig.5). These effects were independent of the presence or absence of TTX, suggesting that NMDA acted directly on NMDARs in Purkinje cells.
Fig. 5.

Effects of NMDA and TTX on Purkinje cell survival. Cerebellar cells were incubated with the indicated concentration of NMDA in the absence (open bar) or presence (filled bar) of 1 μmtetrodotoxin (TTX) for 6 d. The number of Purkinje cells was expressed as a percentage of the number under control conditions in the absence of NMDA and TTX. NMDA (10 μm) significantly increased the number of surviving Purkinje cells in the presence or absence of TTX (p < 0.05; indicated by asterisk). NMDA (100 μm) significantly inhibited the survival (p < 0.05); n = 6–9 cultures.
The effect of TTX and NMDA on numbers of granule cells was studied by counting the number of small (4–5 μm diameter) round cells under phase-contrast microscopy. Although 140–300 μmNMDA has been reported to promote the survival of granule neurons (Balázs et al., 1988; Didier et al., 1990), it had no effect on the number of granule neurons here. This may be because granule cells were prepared from E18–P0 mice and maintained for 7 d in this study, whereas the critical period described for the survival effect of NMDA was 2–5 d after plating of P7 mice (Didier et al., 1990) or P9 rats (Balázs et al., 1988).
TTX cannot block the release of substances such as neurotrophic factors that may be independent of action potential. To rule out the possibility of NMDA-induced release of such substances from other cells, the survival of Purkinje cells from NR1−/− mice was investigated (Forrest et al., 1994). First, NR1−/−, −/+, or +/+ cerebellar cultures were made at P0; at 7 div, the numbers of Purkinje cells were compared. Older neonates could not be used because NR1−/− mice die on P0 as a result of respiratory failure. As shown in Figure6, the number of NR1−/− Purkinje cells was significantly less than NR1−/+ or +/+ Purkinje cells. The difference between NR1−/+ and NR1+/+ was not significant, which is consistent with the electrophysiological data showing no difference in NMDA-induced currents between neurons from NR1−/+ and +/+ mice (Forrest et al., 1994). At 4 hr in vitro, there were no differences in the number of Purkinje cells among each genotype [+/+ (190 ± 34, n = 13), −/− (201 ± 28, n = 19), and −/+ (185 ± 14, n = 30)], suggesting that most Purkinje cells died between day 0 and day 7. The difference in the number of surviving Purkinje cells between NR1−/− and −/+ or +/+ mice was less than the reduction caused by NMDA antagonists on wild-type cultures (Fig. 3). The discrepancy in the degree of survival caused by the absence of the receptor (NR1−/−) and by pharmacological antagonists could reflect, for example, a compensatory mechanism in the mutant mice (Forrest et al., 1994); however, survival of Purkinje cells was clearly inhibited in both cases. The number of granule cells was 3400 ± 120, 3200 ± 130, and 3500 ± 150 (n = 5 each) in +/+, −/+, and −/− cultures, respectively. Thus the absence of the NMDAR had no significant effect on the survival of granule cells at 7 div.
Fig. 6.
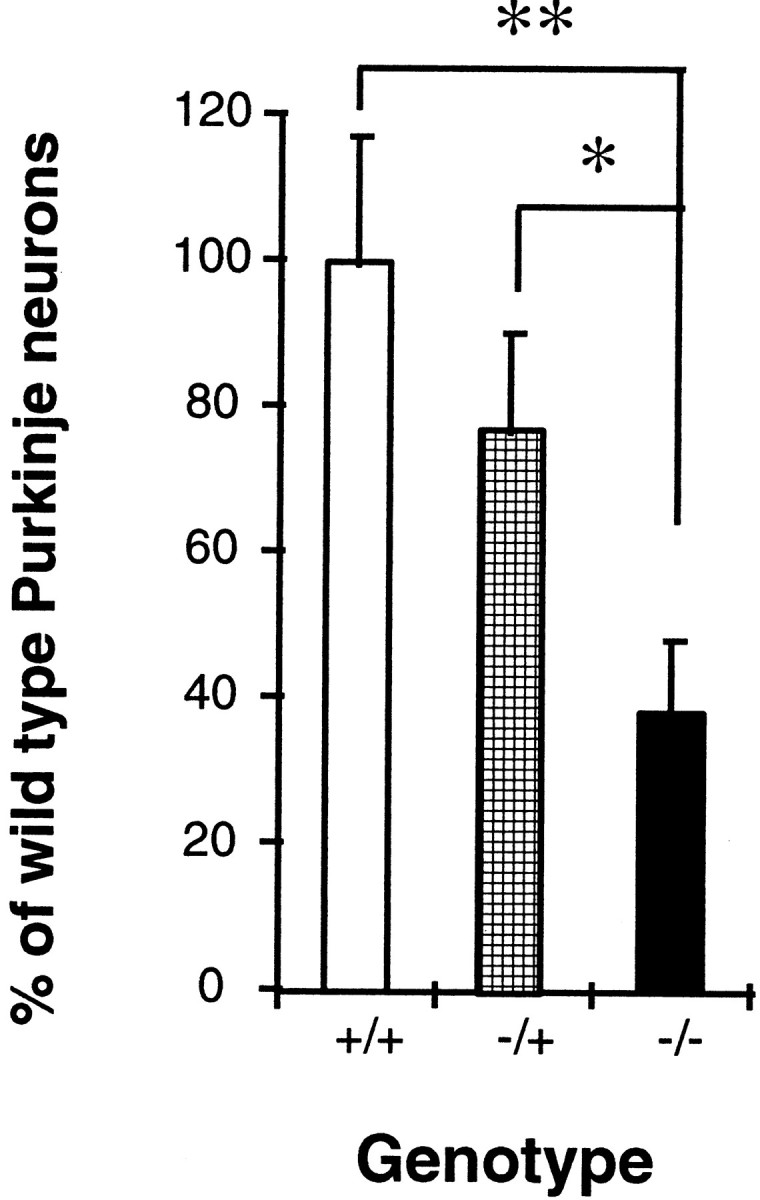
Survival of Purkinje cells in NR1−/− mutants. Cerebellar cultures were prepared from individual NR1−/−, −/+, or +/+ mice. Cells were maintained for 6 div, and the number of Purkinje cells was counted. After genotypes were determined, data from the same genotype were combined and expressed as a percentage of the number of Purkinje cells from wild-type (+/+) mice. The number of NR1−/− Purkinje cells was significantly less than the NR1−/+ (p < 0.05; indicated by asterisk) or +/+ (p < 0.01; indicated by double asterisks) Purkinje cells; n = 5–8 cultures.
The reduced numbers of Purkinje cells in NR1−/− cultures can be explained in two ways: the absence of functional NMDARs in non-Purkinje cells leads to the loss of Purkinje cells, or the absence of NMDARs in Purkinje cells leads directly to the loss of Purkinje cells. To investigate which NMDARs are critical for the survival of Purkinje cells, we prepared Purkinje cell-depleted cultures, which we refer to as “granule-cell feeder” cultures. When cerebellar cultures were prepared from wild-type newborn mice (P4–P7) or at low plating density (0.8 × 105/cm2), few Purkinje cells survived and 90–95% of total cells were granule cells, as was reported previously (Fischer, 1982; Weber and Schachner, 1984). Thus, when we plated new cells on top of this preconditioned granule-cell feeder, all Purkinje cells in this culture were considered to be derived from cells plated on top of feeder cells. Therefore, by comparing the survival of Purkinje cells in different combinations of genotypes of feeder cells and top cells, we tested NMDARs expressed in the cells that were critical for the survival of Purkinje cells.
First, the effect of the presence of NMDARs in feeder cultures was studied by preparing feeder cells at a plating density of 0.8 × 105 cells/cm2 from mice of different genotypes (Fig. 7A). Feeder cells from −/− mice promoted the survival of Purkinje cells to the same degree as those from −/+ or +/+ mice (Fig. 7B). The difference in the number of Purkinje cells was dependent only on the genotype of the cells plated on top of the feeder culture, indicating that the activity of NMDARs in Purkinje cells themselves directly promotes their survival, and that the effect of the granule cell feeder is independent of NMDAR activity.
In this experiment, we used P0 mice to prepare feeder cells, because NR1−/− mice die on P0. Thus, only a small number of Purkinje cells could be derived from the feeder cultures even when we plated cells at low density. Therefore, we next prepared feeder cells from P5–P6 wild-type mice at low plating density and plated top cells from mice of different genotypes (Fig. 8A). In this condition, we confirmed that all Purkinje cells were derived from cells plated on top of the feeder cells. There were significant differences in the survival of Purkinje cells between the −/− and −/+ or +/+ cultures even on wild-type feeder granule cells expressing NMDARs (Fig.8B), suggesting again that the activity of NMDARs in Purkinje cells affects the survival.
The effect of the granule cell feeder layer was also shown to be independent of NMDAR activity, because the addition of APV did not inhibit the effect of the feeder layer, and the difference between each genotype disappeared (Fig. 7B). Taken together, these findings suggest that although non-Purkinje cells significantly support the survival of Purkinje cells, the effect is independent of NMDARs, and that the activity of NMDARs in Purkinje cells constitutes an additional survival-promoting factor.
DISCUSSION
In this study, we have shown that functional NMDARs are present in unambiguously identified Purkinje cells in culture. With use of a rapid drug application method, we have found that the NMDAR-mediated current not only decreases in amplitude but also shows progressive desensitization during development in culture. We have also demonstrated that the NMDARs in Purkinje cells, but not in non-Purkinje cells, have a direct role in the survival of Purkinje cells in vitro
Interestingly, the period during which NMDAR-mediated currents were most prominent corresponds to the time when the cerebellum is in the initial stages of wiring and the inputs to Purkinje neurons that are dominant in the mature cerebellum are scarce. Parallel fibers, one of the two classes of glutaminergic synaptic inputs to Purkinje cells, begin innervation only after the first postnatal week (Ito, 1984). Moreover, granule neurons, the source of parallel fibers, are still proliferating and migrating from the external germinal layer until about the third postnatal week (Ito, 1984). Smeyne and Goldowitz (1989)reported that the majority of granule cells in vivo are generated postnatally with a peak at P6 and then stay at the external granule layer before final cell division. Climbing fibers, another class of glutaminergic synaptic inputs to Purkinje cells, begin immature multiple innervation as early as P3 in the rat (Crepel, 1971); however, there was no NMDAR component to the climbing fiber response in Purkinje cells from young rats at P8 (Llano et al., 1991). Therefore, the expression of the NMDA response that we show here in the Purkinje cells is greatest when synaptic input is the weakest.
We postulate that during the early stage of the period we studied, NMDARs in postnatal Purkinje cells may be activated by ambient glutamate in a nonsynaptic manner because TTX, which blocks neurotransmission at synapses, had no effect on Purkinje cell survival in this study. Furthermore, spontaneous synaptic activity did not occur until 10–14 div in cerebellar dispersed cultures (Yuzaki et al., 1990). The finding that Purkinje neurons from NR1−/− mice had lower background current noise and higher input resistance than those from wild-type mice (Forrest et al., 1994) is consistent with this view. The concentration of glutamate in culture medium containing glutamine is reported to be between 1 and 70 μm (Didier et al., 1990; Driscoll et al., 1991). Although no glutamate was added to our medium, glutamate could be generated by spontaneous release from presynaptic terminals, contamination of glutamate in glutamine added to the medium, or conversion of glutamine to glutamate by extracellular glutaminase released from damaged cells (Sah et al., 1989; Driscoll et al., 1991). Therefore, it is possible that the NMDARs may be activated tonically by ambient glutamate, because NMDAR has a high affinity for glutamate (EC50 = 2.3 μm;Patneau and Mayer, 1990).
Tonic activation of NMDAR by ambient glutamate has also been reported in cerebral cortex in vivo (Blanton et al., 1990), in hippocampal slices (Sah et al., 1989), and in cerebellar slices (Komuro and Rakic, 1993). Glutamate is also present in cerebrospinal fluid at concentrations of 2–26 μm (Perry et al., 1975;McGale et al., 1977). We consider it likely that such tonic nonsynaptic activation of NMDARs by ambient glutamate may also occur in vivo in Purkinje cells as well as in vitro, as we have shown here. The nonsynaptic activation of transient functional NMDARs, through a unique property such as high Ca2+permeability, may provide an important signal that is required for the expression of necessary functions in early postnatal Purkinje cells. These may include the survival of Purkinje cells in vivo as we have shown in vitro. During the later phase of the period studied, however, transiently active NMDARs may also be essential in synaptic competition in vivo, because the window of activity found here coincides with early stages of synapse formation and migration of mossy (Takeda and Maekawa, 1989) and climbing (Crepel, 1971) fiber synapses on Purkinje cells. Indeed, NMDAR activation is reported to be a critical step in the regression of supernumerary climbing fiber synapses during development (Rabacchi et al., 1992). The role of active NMDARs in synaptic competition and in the survival of target neurons might be mediated by a common mechanism such as an increase in neurotrophic factor expression or an increase in the binding of neurotrophic factors to their receptors, both of which have been demonstrated in activity-dependent survival and synaptic competition in other preparations (Ghosh, 1996).
Although the granule neuron feeder layer had a major effect on Purkinje neuron survival (Figs. 7, 8), it is difficult to assess the in vivo relevance of the finding. First, granule neuron numbers are low in the first postnatal week (see above), and second, the feeder layer used was more mature than the Purkinje neurons (also see Baptista et al., 1994). Our data indicate that this trophic effect probably has nothing to do with functional NMDA receptors, because survival was enhanced in both wild-type and NR1−/− cultures.
NMDAR activity has also been reported to affect the morphological differentiation of several types of cells, including hippocampal neurons (Mattson et al., 1988; Brewer and Cotman, 1989) and cerebellar granule cells (Balzs et al., 1988). None of the treatments used here to alter NMDAR activity, however, caused noticeable morphological effects on the early stages of differentiation of Purkinje cells. This is consistent with our previous study of the cerebellar anatomy of neonatal mice lacking NMDAR1, where no obvious morphological defect was observed (Forrest et al., 1994).
The characteristics of NMDAR current and its changes with development described here go a long way toward reconciling the previous mixed reports regarding the presence of functional NMDARs in Purkinje cells (see introductory remarks). Although there is reasonable consensus on several points, a major unreconciled finding is the reported absence of functional NMDARs in Purkinje cells of postnatal animals. It has been reported that there were no direct NMDA responses in Purkinje cells from P8–P12 (Farrant and Cull-Candy, 1991) and P8–P22 (Llano et al., 1991) rats. Earlier reports, however, showed the presence of functional NMDARs in acutely isolated Purkinje cell from P0–P4 rats (Rosenmund et al., 1992), and other studies showed that NMDARs are transiently active in the first few weeks of postnatal development, as shown here (Fig. 2) and in previous studies (Dupont et al., 1987; Garthwaite et al., 1987;Krupa and Crepel, 1990; Yuzaki and Mikoshiba, 1992). Our data now suggest that any NMDA current in slice preparations from animals older than 1 week would be small, and if the method of agonist application was slow, as is generally the case in slice preparations (Farrant and Cull-Candy, 1991; Llano et al., 1991), the response would be desensitized without producing a large current. It should be noted thatSekiguchi et al. (1987) showed NMDA-induced current in Purkinje cells in adult rat slice preparations with rapid, local application of NMDA to the proximal dendrite of Purkinje cells by iontophoresis. When synaptic activation is used in the slice (Perkel et al., 1990; Llano et al., 1991), delivery of the agonist is very rapid, but it may not address the location of the receptors.
Mount et al. (1993) reported that rat Purkinje cell survival was promoted by APV (100 μm) or MK-801 (10 μm) by ∼40–60%. The discrepancy with our results might be attributable to the difference in local density of neurons and in glial cells, in addition to the different species and culture technique used. NMDA applied to the medium promoted the survival of Purkinje cells in a limited range (10–30 μm) (Fig. 5), and concentrations higher than 100 μm decreased survival. Similar neurotoxic and neurotrophic effects of NMDA on granule cells, which depend on the duration and concentration of NMDA and the concentration of Mg2+, have been reported (Didier et al., 1990). Thus, when the concentration of glutamate in the local medium is relatively high because of cell crowding, NMDA antagonists could exert a Purkinje cell-supporting effect. In addition, the relative age of the rats from which cultures were established and analyzed also differed from ours. This could be critical, because we showed that the NMDARs are only transiently functional during a limited window of development.
Although NMDAR1 mRNA and protein are expressed abundantly in mature Purkinje neurons (Moriyoshi et al., 1991; Akazawa et al., 1994; Monyer et al., 1994; Petralia et al., 1994; Watanabe et al., 1994) and are essential for functional NMDARs (Forrest et al., 1994), NMDAR1 by itself produces only small currents when analyzed using in vitro expression in Xenopus oocytes (Moriyoshi et al., 1991). Coexpression of NMDAR 1 and NMDAR2 subunits, however, yielded larger currents (Kutsuwada et al., 1992; Meguro et al., 1992; Monyer et al., 1992). In mouse Purkinje cells, NMDAR1 mRNA was reported to be expressed from E13 through adult, whereas NMDAR2B mRNA was seen transiently from E13 to P1 (Watanabe et al., 1994). On the other hand, in rat Purkinje cells, there is no consensus regarding the expression of NMDAR2 mRNAs. Monyer et al. (1994) showed no signal for NMDAR2 mRNAs from E17 through adult, whereas Akazawa et al. (1994) showed delayed expression of NMDA2A from P11 to adult and transient expression of NMDAR2C mRNA from P14 to adult. Thus, it is still not clear whether changes in expression of NMDAR2 genes account for the changes in NMDAR sensitivity in Purkinje cells. The culture system we used here will be useful to elucidate the mechanisms responsible for the changes in NMDA current and also to elucidate the signal that induces such changes.
Footnotes
This work was supported in part by the International Human Frontier Science Program Organization (LT-663/93), the National Institutes of Health Cancer Center Support CORE Grant PBOCA 21765, and the American Lebanese Syrian Associated Charities.
Correspondence should be addressed to Michisuke Yuzaki, St. Jude Children’s Research Hospital, Department of Developmental Neurobiology, 332 North Lauderdale, Memphis, TN 38105-2794.
REFERENCES
- 1.Akazawa C, Shigemoto R, Bessho Y, Nakanishi S, Mizuno N. Differential expression of five N -methyl-d-aspartate receptor subunit mRNAs in the cerebellum of developing and adult rats. J Comp Neurol. 1994;347:150–160. doi: 10.1002/cne.903470112. [DOI] [PubMed] [Google Scholar]
- 2.Ascher P, Nowak L. The roles of divalent cations in the N -methyl-d-aspartate responses of mouse central neurones in culture. J Physiol (Lond) 1988;399:247–266. doi: 10.1113/jphysiol.1988.sp017078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Audinat E, Knöpfel T, Gähwiler BH. Responses to excitatory amino acids of Purkinje cells and neurones of the deep nuclei in cerebellar slice cultures. J Physiol (Lond) 1990;430:297–313. doi: 10.1113/jphysiol.1990.sp018292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Balázs R, Jorgensen OS, Hack N. N -methyl-d-aspartate promotes the survival of cerebellar granule cells in culture. Neuroscience. 1988;27:437–451. doi: 10.1016/0306-4522(88)90279-5. [DOI] [PubMed] [Google Scholar]
- 5.Baptista CA, Hatten ME, Blazeski R, Mason CA. Cell–cell interactions influence survival and differentiation of purified Purkinje cells in vitro . Neuron. 1994;12:243–260. doi: 10.1016/0896-6273(94)90268-2. [DOI] [PubMed] [Google Scholar]
- 6.Blanton MG, Lo Turco JJ, Kriegstein AR. Endogenous neurotransmitter activates N -methyl-d-aspartate receptors on differentiating neurons in embryonic cortex. Proc Natl Acad Sci USA. 1990;87:8027–8030. doi: 10.1073/pnas.87.20.8027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bliss T, Collingridge GD. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. doi: 10.1038/361031a0. [DOI] [PubMed] [Google Scholar]
- 8.Brewer GJ, Cotman CW. NMDA receptor regulation of neuronal morphology in cultured hippocampal neurons. Neurosci Lett. 1989;99:268–273. doi: 10.1016/0304-3940(89)90458-8. [DOI] [PubMed] [Google Scholar]
- 9.Buller AL, Larson HC, Schneider E, Beaton JA, Morrisett RA, Monaghan DT. The molecular basis of NMDA receptor subtypes: native receptor diversity is predicted by subunit composition. J Neurosci. 1994;14:5471–5484. doi: 10.1523/JNEUROSCI.14-09-05471.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Crepel F. Maturation of climbing fiber responses in the rat. Brain Res. 1971;35:272–276. doi: 10.1016/0006-8993(71)90617-2. [DOI] [PubMed] [Google Scholar]
- 11.Didier M, Heaulme M, Soubrié P, Bockart J, Pin J-P. Rapid, sensitive, and simple method for quantification of both neurotoxic and neurotrophic effects of NMDA on cultured cerebellar granule cells. J Neurosci Res. 1990;27:25–35. doi: 10.1002/jnr.490270105. [DOI] [PubMed] [Google Scholar]
- 12.Driscoll BF, Law MJ, Crane AM. Cell damage associated with changing the medium of mesencephalic cultures in serum-free medium is mediated via N -methyl-d-aspartate receptors. J Neurochem. 1991;56:1201–1206. doi: 10.1111/j.1471-4159.1991.tb11411.x. [DOI] [PubMed] [Google Scholar]
- 13.Dupont JL, Gardette R, Crepel F. Postnatal development of the chemosensitivity of rat cerebellar Purkinje cells to excitatory amino acids: an in vitro study. Dev Brain Res. 1987;34:59–68. doi: 10.1016/0165-3806(87)90195-7. [DOI] [PubMed] [Google Scholar]
- 14.Enderlin S, Norman AW, Celio MR. Ontogeny of the calcium binding protein calbindin D-28k in the rat nervous system. Anat Embryol (Berl) 1987;177:15–28. doi: 10.1007/BF00325286. [DOI] [PubMed] [Google Scholar]
- 15.Farrant M, Cull-Candy SG. Excitatory amino acid receptor-channels in Purkinje cells in thin cerebellar slices. Proc R Soc Lond [Biol] 1991;244:179–184. doi: 10.1098/rspb.1991.0067. [DOI] [PubMed] [Google Scholar]
- 16.Fischer G. Cultivation of mouse cerebellar cells in serum free, hormonally defined media: survival of neurons. Neurosci Lett. 1982;28:325–329. doi: 10.1016/0304-3940(82)90079-9. [DOI] [PubMed] [Google Scholar]
- 17.Forrest D, Yuzaki M, Soares HD, Ng L, Luk DC, Sheng M, Stewart CL, Morgan JI, Connor JA, Curran T. Targeted disruption of NMDA receptor 1 gene abolishes NMDA response and results in neonatal death. Neuron. 1994;13:325–338. doi: 10.1016/0896-6273(94)90350-6. [DOI] [PubMed] [Google Scholar]
- 18.Garthwaite G, Yamini B, Jr, Garthwaite J. Selective loss of Purkinje and granule cell responsiveness to N -methyl-d-aspartate in rat cerebellum during development. Dev Brain Res. 1987;36:288–292. doi: 10.1016/0165-3806(87)90034-4. [DOI] [PubMed] [Google Scholar]
- 19.Ghosh A. Cortical development: with an eye on neurotrophins. Curr Biol. 1996;6:130–133. doi: 10.1016/s0960-9822(02)00442-6. [DOI] [PubMed] [Google Scholar]
- 20.Hockberger PE, Tseng HY, Connor JA. Fura-2 measurements of cultured rat Purkinje cells show dendritic localization of Ca2+ influx. J Neurosci. 1989;9:2272–2284. doi: 10.1523/JNEUROSCI.09-07-02272.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Iacopino AM, Rhoten WB, Christakos S. Calcium binding protein (calbindin-D28k) gene expression in the developing and aging mouse cerebellum. Mol Brain Res. 1990;8:283–290. doi: 10.1016/0169-328x(90)90041-b. [DOI] [PubMed] [Google Scholar]
- 22.Ito M. Raven; New York: 1984. The cerebellum and neural control. . [Google Scholar]
- 23.Joels M, Yool AJ, Gruol DL. Unique properties of non-N -methyl-d-aspartate excitatory responses in cultured Purkinje cells. Proc Natl Acad Sci USA. 1989;86:3404–3408. doi: 10.1073/pnas.86.9.3404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kaneko T, Itoh K, Sigemoto R, Mizuno N. Glutamate-like immunoreactivity in the lower brain stem and cerebellum of the adult rat. Neuroscience. 1989;32:79–98. doi: 10.1016/0306-4522(89)90109-7. [DOI] [PubMed] [Google Scholar]
- 25.Komuro H, Rakic P. Modulation of neuronal migration by NMDA receptors. Science. 1993;260:95–97. doi: 10.1126/science.8096653. [DOI] [PubMed] [Google Scholar]
- 26.Krupa M, Crepel F. Transient sensitivity of rat cerebellar Purkinje cells to N -methyl-d-aspartate during development: a voltage clamp study in in vitro slices. Eur J Neurosci. 1990;2:312–316. doi: 10.1111/j.1460-9568.1990.tb00423.x. [DOI] [PubMed] [Google Scholar]
- 27.Kutsuwada T, Kashiwabuchi N, Mori H, Sakimura K, Kushiya E, Araki K, Meguro H, Masaki H, Kumanishi T, Arakawa M, Mishina M. Molecular diversity of the NMDA receptor channel. Nature. 1992;358:36–41. doi: 10.1038/358036a0. [DOI] [PubMed] [Google Scholar]
- 28.Lee M, Strahlendorf HK, Strahlendorf JC. Differential effects of N -methyl-d-aspartic acid andl-homocysteic acid on cerebellar Purkinje neurons. Brain Res. 1988;456:104–112. doi: 10.1016/0006-8993(88)90351-4. [DOI] [PubMed] [Google Scholar]
- 29.Leonard JP, Kelso SR. Apparent desensitization of NMDA responses in Xenopus oocytes involves calcium-dependent chloride current. Neuron. 1990;2:53–60. doi: 10.1016/0896-6273(90)90443-j. [DOI] [PubMed] [Google Scholar]
- 30.Llano I, Marty A, Johnson JW, Ascher P, Gähwiler BH. Patch-clamp recording of amino acid-activated responses in “organotypic” slice cultures. Proc Natl Acad Sci USA. 1988;85:3221–3225. doi: 10.1073/pnas.85.9.3221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Llano I, Marty A, Armstrong CM, Konnerth A. Synaptic- and agonist-induced excitatory currents of Purkinje cells in rat cerebellar slices. J Physiol (Lond) 1991;434:183–213. doi: 10.1113/jphysiol.1991.sp018465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.MacDermott AB, Mayer ML, Westbrook GL, Smith SJ, Barker JL. NMDA-receptor activation increases calcium concentration in cultured spinal cord neurons. Nature. 1986;321:519–522. doi: 10.1038/321519a0. [DOI] [PubMed] [Google Scholar]
- 33.Malenka R, Nicoll RA. NMDA receptor-dependent synaptic plasticity: multiple forms and mechanisms. Trends Neurosci. 1993;16:521–527. doi: 10.1016/0166-2236(93)90197-t. [DOI] [PubMed] [Google Scholar]
- 34.Mattson MP, Lee RE, Adams ME, Guthrie PB, Kater SB. Interactions between entorhinal axons and target hippocampal neurons: a role for glutamate in the development of hippocampal circuitry. Neuron. 1988;1:865–876. doi: 10.1016/0896-6273(88)90134-1. [DOI] [PubMed] [Google Scholar]
- 35.Mayer M, Westbrook GL. The physiology of excitatory amino acids in the central nervous system. Prog Neurobiol. 1987;28:197–276. doi: 10.1016/0301-0082(87)90011-6. [DOI] [PubMed] [Google Scholar]
- 36.McDonald JW, Johnston MV. Physiological and pathophysiological roles of excitatory amino acids during central nervous system development. Brain Res Rev. 1990;15:41–70. doi: 10.1016/0165-0173(90)90011-c. [DOI] [PubMed] [Google Scholar]
- 37.McGale EH, Pye IF, Stonier C, Hutchinson EC, Aber GM. Studies of the inter-relationship between cerebrospinal fluid and plasma amino acid concentrations in normal individuals. J Neurochem. 1977;29:291–297. doi: 10.1111/j.1471-4159.1977.tb09621.x. [DOI] [PubMed] [Google Scholar]
- 38.Meguro H, Mori H, Araki K, Kushiya E, Kutsuwada T, Yamazaki M, Kumanishi T, Arakawa M, Sakimura K, Mishina M. Functional characterization of a heteromeric NMDA receptor channel expressed from cloned cDNAs. Nature. 1992;357:70–74. doi: 10.1038/357070a0. [DOI] [PubMed] [Google Scholar]
- 39.Monyer H, Sprengel R, Schoepfer R, Herb A, Higuchi M, Lomeli H, Burnashev N, Sakmann B, Seeburg PH. Heteromeric NMDA receptors: molecular and functional distinction of subtypes. Science. 1992;256:1217–1221. doi: 10.1126/science.256.5060.1217. [DOI] [PubMed] [Google Scholar]
- 40.Monyer H, Burnashev N, Laurie DJ, Sakmann B, Seeburg PH. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron. 1994;12:529–540. doi: 10.1016/0896-6273(94)90210-0. [DOI] [PubMed] [Google Scholar]
- 41.Moriyoshi K, Masu M, Ishii T, Shigemoto R, Mizuno N, Nakanishi S. Molecular cloning and characterization of the rat NMDA receptor. Nature. 1991;354:31–37. doi: 10.1038/354031a0. [DOI] [PubMed] [Google Scholar]
- 42.Mount HTJ, Dreyfus CF, Black IB. Purkinje cell survival is differentially regulated by metabotropic and ionotropic excitatory amino acid receptors. J Neurosci. 1993;13:3173–3179. doi: 10.1523/JNEUROSCI.13-07-03173.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Murase K, Randic M, Shirasaki T, Nakagawa T, Akaike N. Serotonin suppresses N -methyl-d-aspartate responses in acutely isolated spinal dorsal horn neurons of the rat. Brain Res. 1990;525:84–91. doi: 10.1016/0006-8993(90)91323-9. [DOI] [PubMed] [Google Scholar]
- 44.Nakanishi S. Molecular diversity of glutamate receptors and implications for brain function. Science. 1992;258:597–603. doi: 10.1126/science.1329206. [DOI] [PubMed] [Google Scholar]
- 45.Ogura A, Iijima T, Amano T, Kudo Y. Optical monitoring of excitatory synaptic activity between cultured hippocampal neurons by a multi-site Ca2+ fluorometry. Neurosci Lett. 1987;78:69–74. doi: 10.1016/0304-3940(87)90563-5. [DOI] [PubMed] [Google Scholar]
- 46.Patneau DK, Mayer ML. Structure-activity relationships for amino acid transmitter candidates acting at N -methyl-d-aspartate and quisqualate receptors. J Neurosci. 1990;10:2385–2399. doi: 10.1523/JNEUROSCI.10-07-02385.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Perkel DJ, Hestrin S, Sah P, Nicoll RA. Excitatory synaptic currents in Purkinje cells. Proc R Soc Lond [Biol] 1990;241:116–121. doi: 10.1098/rspb.1990.0074. [DOI] [PubMed] [Google Scholar]
- 48.Perry TL, Hansen S, Kennedy J. CSF amino acids and plasma-CSF amino acid ratios in adults. J Neurochem. 1975;24:587–589. doi: 10.1111/j.1471-4159.1975.tb07680.x. [DOI] [PubMed] [Google Scholar]
- 49.Petralia RS, Yokotani N, Wenthold RJ. Light and electron microscope distribution of the NMDA receptor subunit NMDAR1 in the rat central nervous system using a selective anti-peptide antibody. J Neurosci. 1994;14:667–696. doi: 10.1523/JNEUROSCI.14-02-00667.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rabacchi S, Bailly Y, Delhaye-Bouchaud N, Marianti J. Involvement of the N -methyl-d-aspartate receptor in synapse elimination during cerebellar development. Science. 1992;256:1823–1825. doi: 10.1126/science.1352066. [DOI] [PubMed] [Google Scholar]
- 51.Rosenmund C, Legendre P, Westbrook GL. Expression of NMDA channels on cerebellar Purkinje cells acutely dissociated from newborn rats. J Neurophysiol. 1992;68:1901–1905. doi: 10.1152/jn.1992.68.5.1901. [DOI] [PubMed] [Google Scholar]
- 52.Sah P, Hestrin S, Nicoll RA. Tonic activation of NMDA receptors by ambient glutamate enhances excitability of neurons. Science. 1989;246:815–818. doi: 10.1126/science.2573153. [DOI] [PubMed] [Google Scholar]
- 53.Sather W, Johnson JW, Henderson G, Ascher P. Glycine-insensitive desensitization of NMDA responses in cultured mouse embryonic neurons. Neuron. 1990;4:725–731. doi: 10.1016/0896-6273(90)90198-o. [DOI] [PubMed] [Google Scholar]
- 54.Schilling K, Dickinson MH, Connor JA, Morgan JI. Electrical activity in cerebellar cultures determines Purkinje cell dendritic growth patterns. Neuron. 1991;7:891–902. doi: 10.1016/0896-6273(91)90335-w. [DOI] [PubMed] [Google Scholar]
- 55.Seeburg PH. The molecular biology of mammalian glutamate receptor channels. Trends Neurosci. 1993;16:359–365. doi: 10.1016/0166-2236(93)90093-2. [DOI] [PubMed] [Google Scholar]
- 56.Sekiguchi M, Okamoto K, Sakai Y. NMDA receptors on Purkinje cell dendrites in guinea pig cerebellar slices. Brain Res. 1987;437:402–406. doi: 10.1016/0006-8993(87)91661-1. [DOI] [PubMed] [Google Scholar]
- 57.Smeyne RJ, Goldowitz D. Development and death of external granular layer cells in the weaver mouse cerebellum: a quantitative study. J Neurosci. 1989;9:1608–1620. doi: 10.1523/JNEUROSCI.09-05-01608.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sorimachi M, Nishimura S, Morita Y. N -methyl-d-aspartate and aspartate raise the cytosolic free calcium concentration by acting upon receptors transiently expressed on immature cerebellar Purkinje cells. Neurosci Lett. 1991;132:89–92. doi: 10.1016/0304-3940(91)90440-5. [DOI] [PubMed] [Google Scholar]
- 59.Takeda T, Maekawa K. Transient direct connection of vestibular mossy fibers to the vestibulocerebellar Purkinje cells in early postnatal development of kittens. Neurosci. 1989;32:99–111. doi: 10.1016/0306-4522(89)90110-3. [DOI] [PubMed] [Google Scholar]
- 60.Wassef M, Zanetta JP, Brehier A, Sotelo C. Transient biochemical compartmentalization of Purkinje cells during early cerebellar development. Dev Biol. 1985;111:129–137. doi: 10.1016/0012-1606(85)90441-5. [DOI] [PubMed] [Google Scholar]
- 61.Watanabe M, Mishina M, Inoue Y. Distinct spatiotemporal expressions of five NMDA receptor channel subunit mRNAs in the cerebellum. J Comp Neurol. 1994;343:513–519. doi: 10.1002/cne.903430402. [DOI] [PubMed] [Google Scholar]
- 62.Weber A, Schachner M. Maintenance of immunocytologically identified Purkinje cells from mouse cerebellum in monolayer culture. Brain Res. 1984;311:119–130. doi: 10.1016/0006-8993(84)91404-5. [DOI] [PubMed] [Google Scholar]
- 63.Yuzaki M, Mikoshiba K. Pharmacological and immunocytochemical characterization of the metabotropic glutamate receptors in cultured Purkinje cells. J Neurosci. 1992;12:4253–4263. doi: 10.1523/JNEUROSCI.12-11-04253.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Yuzaki M, Kudo Y, Akita K, Miyawaki A, Mikoshiba K. MK-801 blocked the functional NMDA receptors in identified cerebellar neurons. Neurosci Lett. 1990;119:19–22. doi: 10.1016/0304-3940(90)90745-u. [DOI] [PubMed] [Google Scholar]