

Abstract
We examined the properties and regulation of Ca channels resulting from the expression of human α1B and α1E subunits stably expressed in HEK293 cells. The ancillary subunits β1B and α2/δ were also stably expressed in these cell lines. Ca currents in α1B-expressing cells had the properties of N-type currents. Ca currents in cells expressing α1E exhibited a novel profile that was similar to the properties of the “R type” Ca current. Introduction of GTP-γ-S into α1B cells greatly enhanced the extent of prepulse facilitation of the Ca current, whereas it had only a very small effect in α1E-expressing cells. Activation of somatostatin receptors endogenous to HEK293 cells or κ opioid receptors, expressed in the cells after transfection, inhibited Ca currents in α1B-expressing cells. This inhibition was blocked by pertussis toxin and was partially relieved by a depolarizing prepulse. In contrast, no inhibitory effects were noted in cells expressing α1E channels under the same circumstances. HEK293 cells normally contained G-proteins from all of the four major families. Inhibition of Ca currents by κ agonists in α1B-expressing cells was enhanced slightly by the cotransfection of several G-protein α subunits. κ agonists, however, had no effect in α1E-containing cells, even after overexpression of different G-protein α-subunits. In summary, these results demonstrate that there is a large difference in the susceptibility of α1B- and α1E-based Ca channels to regulation by G-proteins. This is so despite the fact that the two types of Ca channels show substantial similarities in their primary sequences.
Keywords: Ca channel, G-protein, κ receptor, patch clamp, HEK293, somatostatin
Regulation by receptors, G-proteins, and second messengers is one of the most widely observed characteristics of Ca channels (Hille, 1994; Dolphin, 1995). For example, the activation of L channels in the heart by cAMP-dependent phosphorylation has been studied extensively, and the physiological significance of these observations is clear (Perez-Reyes et al., 1994). In the nervous system, it has been shown frequently that activation of various G-protein-linked receptors can produce inhibition of N channels (Hille, 1994). Because it is known that N channels play a critical role in the regulation of transmitter release at many synapses (Miller, 1990;Hille, 1994), this process is believed to be a key element in the receptor regulation of synaptic communication, particularly the phenomenon of presynaptic inhibition. Rather than being mediated by a diffusible second messenger, “rapid” inhibition of N channels is thought to be attributable to direct effects of G-protein subunits on some part of the Ca channel complex (Hille, 1994). Therefore, this type of N-channel regulation has been described as “membrane delimited.” It is not yet clear, however, which subunits of the G-protein heterotrimer actually produce these effects or to which part of the N channel they bind (Sternweis, 1994; Wickman and Clapham, 1995).
N channels, which result from the expression of the α1B Ca channel subunit, are found primarily in neurons and neuroendocrine cells (Williams et al., 1992; Wheeler et al., 1995). This is also the case for at least two other types of Ca channels: the P/Q type, which probably results from the expression of α1A, and the recently described α1E subunit, which may produce a Ca current of the “R” type (Williams et al., 1994; Wheeler et al., 1995). Interestingly, the α1A, α1B, and α1E subunits form a Ca channel subfamily and exhibit a high degree of sequence homology (Dolphin, 1995; Wheeler et al., 1995). None of these Ca channels are sensitive to dihydropyridines, which block α1C, α1D, and α1S Ca channels and to which they exhibit less sequence homology (Dolphin, 1995; Wheeler et al., 1995). It is now clear that Ca currents of the P/Q type, which can also support neurotransmitter release (Wheeler et al., 1994), are regulated by G-proteins and receptors in a manner similar to N channels (Mintz and Bean, 1993; Rhim and Miller, 1994; Kanemasa et al., 1995). It is not known at this point, however, whether Ca currents resulting from the expression of α1E are also regulated in this way. Indeed, very little is known about the properties and biological functions of Ca channels resulting from α1Eexpression. We have now investigated this question and demonstrate that there seem to be considerable differences in the ability of α1B and α1E to be regulated by G-proteins.
MATERIALS AND METHODS
HEK293 cell lines. HEK293 cell lines expressing Ca channels with various subunit compositions were kindly provided by SIBIA Inc. (Williams et al., 1992; Bleakman et al., 1995). Briefly, the cell lines were developed by stable co-transfection of HEK293 cells with human α1, α2Bδ, and β Ca channel subunit expression plasmids. The G1A1 cell line consisted of α1B-1-α2Bδ-β1Bsubunits. The subunit composition of the A4A2 cells was α1B-1-α2Bδ-β1C. The E52–3 cell line expressed the α1E-type Ca channel with the α1E-3-α2Bδ-β1Bsubunit composition.
HEK293 cells stably expressing Ca2+-channels were grown in plastic Falcon dishes in DMEM (Life Technologies, Gaithersburg, MD) containing 5% defined bovine serum (HyClone, Logan, UT) plus penicillin G (100 U/ml), streptomycin sulfate (100 μg/ml), and geneticin (500 μg/ml). One day before recording, cells were dissociated by gentle trituration with a fire-polished Pasteur pipette and replated onto poly-l-lysine-coated glass coverslips.
Preparation of G-protein expression plasmids. cDNAs encoding α subunits of Gi1–3 and Go (kindly provided by Randall R. Reed, Johns Hopkins University) were subcloned into the mammalian expression vector pCMV5 (Andersson et al., 1989) and confirmed by DNA sequencing using a modification of the dideoxy-chain termination method (Sequenase 2.0; USB, Cleveland, OH). The cDNA Gsα was similarly subcloned into pCMV6b and confirmed by sequencing (Robishaw et al., 1986). The cDNAs encoding G13 and Gq in pcDNA3 were obtained from G. Babnigg (University of Chicago). κ opioid receptor cDNA in pCMV5 was a kind gift of Graeme Bell (University of Chicago).
Transfection of HEK293 cells. Monolayers of HEK293 cells not exceeding 75% confluence were dissociated and replated onto poly-l-lysine-coated (Sigma, St. Louis, MO) glass coverslips. Cells were cotransfected with plasmids containing the cDNAs for the κ receptor, G-protein, and β-galactosidase using the standard calcium-phosphate precipitation technique (Ausubel et al., 1993) or transfection kit (Mammalian Transfection Kit; Stratagene, La Jolla, CA).
Analysis of gene and cDNA expression. For patch-clamp experiments, duplicate coverslips were routinely stained for β-galactosidase expression, with an average transfection efficiency of 40–70% determined by cell counting. Currents were recorded 48–72 hr after transfection.
The expression of the κ receptor was detected using the specific antibody (kindly provided by Robert Elde, University of Minnesota) and ABC kit (Vector Laboratories, Burlingame, CA).
The expression of the G-protein subunits was evaluated using PCR-Southern blot and Northern blot analysis, as follows. Total RNA was prepared using the CsCl guanidinium isothiocyanate method (Sambrook et al., 1989). Poly(A+) RNA was prepared using an oligo-dT-Sepharose column or an mRNA purification kit (Pharmacia, Uppsala, Sweden); 15–20 μg of total RNA or 1–2 μg of poly(A+) RNA was loaded per lane. RNA was transferred to Hybond-N+ (Amersham) by overnight blotting. The blots were hybridized using the32P-labeled-specific oligonucleotide probes (internal sequence) for the individual G-protein α subunits as shown below. In some control experiments (not shown), cDNAs for specific G-proteins were 32P-labeled by random priming and also used as probes. After hybridization [in the solution containing 20% formamide, 5× SSC (1× SSC: 150 mm NaCl, 15 mm sodium citrate), 2.5× Denhardt’s solution] for 12–20 hr, the membranes were rinsed in 2× SSC, 0.1% SDS for 10 min at room temperature, washed in 0.5× SSC, 0.1% SDS at 42°C for 20–40 min, and then exposed to Kodak X-omat film for 12 hr.
For PCR-based detection of expression, first-strand cDNA was prepared from total RNA using random primers and Superscript reverse transcriptase (Life Technologies, Grand Island, NY). The following oligonucleotide primer pairs (forward and reverse) were used.
Giα1—Forward: 5′-CTGTGGAAGGACAGCGGTGTG-3′
Reverse: 5′-CAGCAACAGAGAATGTAGTG-3′
Internal: 5′-GCAGTGGGGTAGTAAAAATGCATT-3′
Giα2—Forward: 5′-CCTGTCGGGCGTCATCCGGAG-3′
Reverse: 5′-CCATGCTCCCTGCCTGTTCCC-3′
Internal: 5′-GATGAATCGCATGCATGAGAGC-3′
Giα3—Forward: 5′-GATTAAACGTTTATGGCGAG-3′
Reverse: 5′-GCATGACAGGACCAAGGAATG-3′
Internal: 5′-GAGGATGGCATAGTAAAAGCT-3′
Go—Forward: 5′-CACTGAACCATTCTCTGCAG-3′
Reverse: 5′-TTTGGCCTTTGTAAGACACAC-3′
Internal: 5′-CACTCAGCGGCTATGACCAG-3′
Gs—Forward: 5′-GACCAACCGCCTGCAGGAGGC-3′
Reverse: 5′-GGGCATGATTAACAAAGCAACC-3′ (bovine)
Reverse: 5′-GGGCATGATTCACACCGCAACC-3′ (rat)
Internal: 5′-CACGCAGTTGATCACCCACC-3′
Gq—Forward: 5′-CTACCCCTGGTTCCAGAACTCC-3′
Reverse: 5′-CACGCTCACAGAGTCCAGGACG-3′
Internal: 5′-GTCGACTAGGTGGGAATACATG-3′
G13—Forward: 5′-CGGGTTTTCAGCAACGTCTCC-3′
Reverse: 5′-TCAGCAGCTGTCAGCCACA-3′
Internal: 5′-CTCTTAAGCAGTGGGGGTCC-3′
The PCR was used under the following conditions: 45 sec at 94°C, 1 min at 55°C, and 1 min at 72°C for 30 cycles, followed by 10 min at 72°C. An aliquot of the PCR sample was analyzed by electrophoresis in 1% agarose. DNAs were transferred to Hybond-N+ (Amersham) and then hybridized with the specific oligonucleotide probe (internal) for each G-protein α subunit shown above, using the conditions described.
Whole-cell patch clamp. The tight-seal whole-cell configuration of the patch-clamp technique (Hamill et al., 1981) was used to record Ca currents. Recordings were made at room temperature (21–24°C). Currents were recorded using an Axopatch 1D (Axon Instruments, Foster City, CA) amplifier, filtered at 2 kHz by the built-in filter of the amplifier, and stored on the computer. Capacitative transients were canceled at 10 MHz, and their values were obtained directly, together with the series-resistance values from the settings of the Axopatch 1D amplifier. Series-resistance compensation between 40 and 80% was applied. Leak corrections were performed using a P/N protocol. Command pulses were delivered at 20 sec intervals. Soft, soda-lime capillary glass was used to make patch pipettes, which were coated with Sylgard (Dow Corning, Midland, MI) and had resistances of 1.8–3.5 MΩ when filled with internal solution. Extracellular buffer solution for whole-cell voltage-clamp experiments was composed of (in mm): 160 tetraethylammonium chloride, 5 CaCl, 1 MgCl2, 10 HEPES, 10 glucose; pH was adjusted to 7.4 with TEAOH. The standard internal solution consisted of (in mm): 100 CsCl, 37 CsOH, 1 MgCl2, 10 BAPTA, 10 HEPES, 3.6 MgATP, 1 GTP, and 14 Tris2CP, and 50 U/ml-1CPK. The pH was adjusted to 7.3 with CsOH. The osmolarity of the pipette solution was 300 mOsm/l, and the osmolarity of the extracellular solution was between 315 and 323 mOsm/l. GTP was replaced by GTP-γ-S or by GDPβS in the double-pulse and tail-current experiments.
Stock solutions of U69593 (RBI, Natick, MA) and nor-BNI (RBI) were prepared in ethanol and stored at −20°C. [d-Trp8] somatostatin (SOM) stock solutions (Bachem, Belmont, CA) were kept at −80°C. Stock solutions of PTX (100 μg/ml; RBI) were prepared in water and stored at 4°C. Cells were treated with PTX at a final concentration of 200 ng/ml overnight.
RESULTS
Effects of GTP analogs
We examined the regulation of α1B- and α1E-based Ca channels stably expressed in HEK293 cells. Ca channels in these cells also contained stably expressed α2/δ and β1B subunits (Williams et al., 1992; Bleakman et al., 1995). It has been shown that the presence of these ancillary subunits is essential for Ca channels to be expressed efficiently and display all of their normal properties (Williams et al., 1992; Bleakman et al., 1995). Ca currents could be elicited in these cells by step depolarizations to different potentials. Cells expressing α1B produced Ca currents that had properties of typical N currents (Bleakman et al., 1995). Cells expressing α1E produced currents whose properties we have described previously and which are similar to currents described in the literature as “R” type (Williams et al., 1994; Wheeler et al., 1995).
We wished to compare the regulation of α1B and α1E channels by G-proteins. To do this, we began by examining the effect of activating the G-proteins normally found in HEK293 cells directly, using the nonhydrolyzable analog of GTP, GTP-γ-S. One interesting phenomenon was apparent immediately. Normally there was a marked increase in the amplitude of the Ca current in α1B-expressing cells in the period after the pipette broke into the cell (Fig.1A). This “run-up” of the current was not apparent in GTP-γ-S-containing cells (Fig. 1B). One of the hallmarks of rapid “membrane-delimited” N-current inhibition is that it can be relieved by a depolarizing prepulse to high voltages (Bean, 1989; Hille, 1994). If the current observed during GTP-γ-S perfusion represents an inhibited current, then a depolarizing prepulse should increase its amplitude. We therefore used a voltage-clamp protocol that consisted of two test pulses with or without a strong intervening depolarization to +80 mV (Fig.1C,D). The ratio of current integrals (P2/P1) was plotted as a function of time (Fig. 2A–C). The ratio P2/P1 became larger after the prepulse (Fig.2A). Figure 2A shows that the maximum GTP-γ-S effect developed ∼4 min after establishing whole-cell recording conditions. At this time, there was no difference in P2/P1 ratios with GDPβS in the pipette (Fig. 2B), consistent with the “inhibitory” action of GDPβS on G-protein function. The small difference in the P2/P1 ratios observed using 1 mm GTP in the patch pipette was presumably attributable to a small degree of basal G-protein activation under these conditions (Fig. 2C). Interestingly, the degree of “run-up” in α1E-expressing cells was less marked than in α1B-expressing cells. Although there was a trend suggesting that this small degree of run-up was also suppressed by GTP-γ-S, this was not significant (Fig. 1E). Furthermore, in these cells, the current evoked by a test pulse after a depolarization also was actually smaller than without the prepulse (Fig. 1F). This was presumably attributable to differences in the extent of voltage-dependent inactivation between the two types of Ca channels (Williams et al., 1994; Bleakman et al., 1995). Consistent with this finding, the “R”-type current in cerebellar granule neurons also displayed very pronounced voltage-dependent inactivation (Zhang et al., 1993). Thus, for α1E currents, the value of the P2/P1 ratio after the prepulse fell below that seen without the prepulse (Fig.2D–F). We therefore normalized the data by subtracting out the degree of voltage-dependent inactivation. This was obtained from cells perfused with GDPβS, in which we assumed that no G-protein-induced inhibition would be present. After this transformation, the data obtained with GTP or GTP-γ-S in the pipette followed a trend similar to that obtained with α1B-based channels (Fig. 2D–F,red data sets). Comparison of the data in Figure2A–C with that in D–F, however, suggests that although there is some G-protein-mediated inhibition of α1E channels, this is 7- to 10-fold less than that obtained with α1B channels. This suggests that there is a considerable difference in the ability of G-proteins to regulate α1B- and α1E-based Ca channels directly.
Fig. 1.

Ca currents in HEK293 cells expressing the α1B and α1E Ca channel subunits. A, Run-up with intracellular GTP using Ca (5 mm) as the charge carrier in HEK293 cells expressing the α1B Ca channel subunit. Currents were evoked from a holding potential of −90 mV by 200 msec depolarizing pulses to +10 mV every 20 sec. B, Average normalized current with 1 mm GTP (▪) or 0.3 mm GTP-γ-S (▴) in the patch pipette. The run-up of currents from individual experiments was normalized with respect to the first peak current obtained. The normalized values were then averaged, and the mean ± SEM was plotted (1 mm GTP, n = 11; 0.3 mm GTP-γ-S, n = 6). C, D, Relief of GTP-γ-S-induced inhibition of α1B Ca currents by an intervening prepulse depolarization. Ca currents were evoked using a double-pulse protocol without (C, lower trace) or with (D,lower trace) a depolarizing prepulse using GTP-γ-S (0.3 mm) in the patch pipette. The intervening depolarization increased the current amplitude during the second pulse (D, lower trace). Upper lines inC and D are the voltage templates (HP = −90 mV; TP = +10 mV; TP duration = 25 msec; prepulse depolarization potential = +80 mV; duration = 50 msec). P1 and P2 denote the current integrals during the first and second test pulses and are used as such in Figure 2. E, Characteristics of Ca current run-up in cells expressing α1Esubunit. Plot of averaged Ca current amplitude (mean ± SEM) in the presence of GTP-γ-S (▴, 0.3 mm;n = 15) or GTP (▪, 1 mm;n = 6). Calculations as in B. F, Superimposed Ca2+ current traces evoked by the double-pulse voltage protocol with or without a depolarizing prepulse (1 mm GTP in the patch pipette). The Ca current following the prepulse depolarization was actually smaller than without it in the cell line expressing α1Esubunit.
Fig. 2.
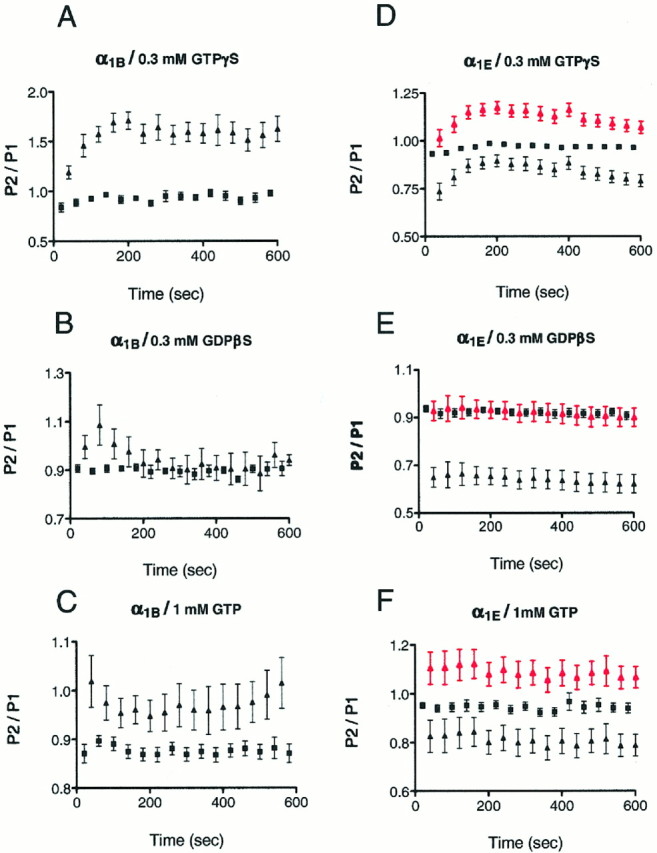
Comparison of the effect of different GTP analogs on the α1B (A–C) and α1E (D–F) Ca current using the double-pulse protocol (see Fig. 1C,D). Currents were evoked every 20 sec in HEK293 cells expressing the α1Bsubunit by applying the double-pulse voltage protocol with GTP-γ-S (A), GDPβS (B), or GTP (C) in the patch pipette. The P2/P1 ratios from individual cells were calculated, and then the isochronal values were averaged during the time course of the experiments from cells in which experiments were carried out under identical conditions. The mean ± SE is plotted; ▪ denotes the P2/P1 ratio without the prepulse, ▴ denotes the ratio with the prepulse. GTP analogs were applied in the following concentrations: GTP-γ-S (0.3 mm; n = 5); GDPβS (0.3 mm; n = 7); GTP (1 mm; n = 11). D–F, Effect of GTP analogs on α1E Ca currents using the double-pulse protocol. Plot of the average P2/P1 ratios in the presence of GTP-γ-S, 0.3 mm (n = 15; D), GDPβS, 0.3 mm(n = 7; E), and GTP, 1 mm (n = 6; F). Note that the values of P2/P1 ratios after prepulse application (▴) fell below those of P2/P1 values obtained without the prepulse depolarization (▪) (black data sets on D–F).D–F, ▴ red data sets show P2/P1 ratios with prepulse after “subtraction” of voltage-dependent inactivation obtained from data with GDPβS in the patch pipette.
Receptor regulation of Ca currents
The results presented above demonstrate that direct activation of G-proteins in these cell lines produces strong inhibition of α1B but not α1Echannels. G-proteins are normally activated by “serpentine” receptors, and this can lead to inhibition of Ca currents in neurons. We attempted to reconstitute receptor regulation of Ca channels in HEK293 cells using endogenous and exogenous receptors. There are several examples in the literature of the regulation of Ca currents by somatostatin receptors (Ikeda and Schofield, 1989; Golard and Siegelbaum, 1993; Fujii et al., 1994; Hille, 1994), and endogenous somatostatin receptors have been reported to exist in HEK293 cells (Law et al., 1993). We found that activation of these receptors with the somatostatin analog SOM produced substantial inhibition of the Ca currents in α1B-expressing cells (Fig.3A,D). Little or no inhibition, however, was observed in cells expressing α1E channels (Fig.3C,D). Inhibition of the α1B Ca current by SOM was repeatable, exhibiting modest desensitization (38.5 ± 5.5% inhibition on first application, 26.2 ± 3.8% inhibition on second; n = 6). Furthermore, inhibition of α1B currents was blocked by pretreatment of cells with PTX (Fig. 3D) and was relieved partially by a depolarizing prepulse (Fig. 3B). We also observed that SOM was equally effective in inhibiting the Ca current in a second α1B-expressing cell line that differed in the type of β subunit (β1C) expressed (Fig.3D).
Fig. 3.

Effect of SOM on α1B-type and α1E Ca currents. A, Plot of α1B-type Ca current versus time showing a typical SOM (300 nm) response. Cell was depolarized from −90 HP to +10 TP every 20 sec. Inset shows Ca currents recorded before and during SOM application. Bshows α1B-type Ca currents evoked by the double-pulse protocol in the presence of SOM (300 nm). Decreased inhibition can be seen after the prepulse. C, Plot of α1E-type Ca current versus time: SOM had little or no effect on the Ca current.D, Average responses (mean ± SEM) to SOM (300 nm) application. The number in parentheses represents the number of experiments.
Activation of κ opioid receptors in some neurons has also been shown to produce inhibition of Ca currents (Lipscombe et al., 1989; Rhim and Miller, 1994). In contrast to the effects of SOM, however, addition of a selective agonist for the κ opioid receptor U69593 produced no inhibition of Ca currents in either the α1B or α1E cell line. This was consistent with the lack of κ opioid receptors in HEK293 cells as indicated by immunostaining or Northern blot analysis (data not shown). We therefore transiently transfected the κ opioid receptor into both α1B- and α1E-expressing cell lines. Northern blots and immunostaining indicated similar degrees of transfection into the two cell lines (not shown). After transfection, U69593 robustly inhibited the Ca current in α1B- but not in α1E-containing cells (Fig. 4). Inhibition by SOM was retained in these cells (data not shown). U69593-induced inhibition was blocked by nor-BNI, a specific inhibitor of κ opioid receptors (Fig. 4). Inhibition produced by U69593 was also partially voltage-dependent and inhibited by PTX treatment (Figs.4, 5).
Fig. 4.

Effects of the κ receptor-selective agonist U69593 on κ receptor-transfected HEK293 cells expressing α1B and α1E Ca channels. Average responses (mean ± SEM) to U69593 (200 nm); n = number of cells showing response to κ receptor agonist application (left to right, bars 1, 3, and 4) or all cells tested (bars 2 and 5). In experiments in which the blocking effect of nor-BNI was examined, cells were also transfected with the α subunit of Go (see below). In the experiments examining the blocking effect of PTX (200 ng/ml overnight), cells were also transfected with the α-subunit of Gi2 (see text).
Fig. 5.

Effect of G-protein α subunit overexpression on Ca current inhibition by κ receptor activation in cells expressing α1B Ca channels. A–C, Plots of Ca current versus time. Insets show Ca current traces at the points indicated before and during the application of the κ receptor agonist U69593 (200 nm). HEK293 cells were transfected with only the κ receptor (A), κ receptor + Giα1 (B), κ receptor + Goα (C), and κ receptor + Giα3 (D). D, Superimposed control and inhibited (U69593; 200 nm) Ca current traces from a cell expressing the α1B Ca channel, κ receptor, and Giα3. The U69593 inhibition was partially relieved by a prepulse depolarization (see Fig. 2).
It seemed possible that our inability to observe strong G-protein regulation of α1E currents was attributable to the fact that HEK293 cells lacked a particular G-protein that was required for regulation of α1E. We observed, however, that under normal conditions the cells expressed mRNAs for many different G-proteins, including representative members of all of the four major families (αi/o, αs, αq, α13) (data not shown). Nevertheless, it was still possible that the actual quantities of some of these might be limiting. We therefore overexpressed several different G-protein α subunits with and without the κ opioid receptor and examined the ability of U69593 or SOM to inhibit Ca currents in the two cell lines. Overexpression of each of the G-protein α subunits was confirmed by Northern blot analysis. Overexpression of some of the G-protein α subunits actually slightly enhanced the ability of U69593 to inhibit Ca currents in α1B-expressing cells (Figs.5B–D, 6A), although the effects of SOM were not altered (Fig. 6B). In cells overexpressing G-protein α subunits, the effects observed were still blocked by PTX (Figs. 4, 6A) and were still partially voltage-dependent (Fig. 5D). On the other hand, no substantial effect of U69593 (Fig. 6C) on α1E currents was observed, even in cells in which we overexpressed different G-protein α subunits.
Fig. 6.
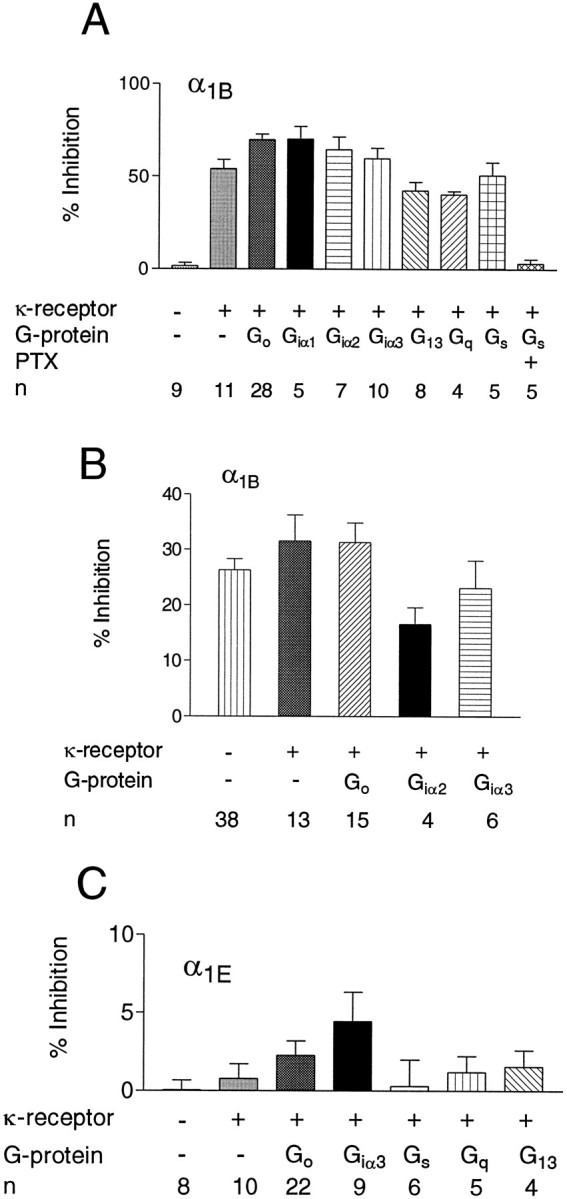
A, Average inhibition (mean ± SEM) of α1B Ca currents by U69593 (200 nm) in HEK293 cells expressing α1B Ca channels, different G-protein α subunits, and the κ opioid receptor. n denotes the number of cells showing agonist responses, except the first and last bars, where all of the responses were averaged. B, Inhibitory effects (mean ± SEM) of SOM (300 nm) in HEK293 cells expressing α1B Ca channels, κ receptors, and various G-protein α subunits. n denotes the number of responsive cells. C, Average inhibition (mean ± SEM) of the Ca current by U69593 (200 nm) in HEK293 cells expressing α1E Ca channels, κ receptors, and different G-protein α subunits. n = total number of cells.
DISCUSSION
We have used an HEK293 cell expression system to make some initial attempts at understanding the G-protein regulation of Ca channels. The two types of Ca channels investigated in the present series of experiments, α1B and α1E, are both members of the family of nondihydropyridine-sensitive channels generally found in neurons and neuroendocrine cells. These α subunits exhibit a high degree of homology with each other; they are ∼80% homologous in those regions that have been suggested as compromising the 24 transmembrane helices. The degree of homology between the two types of channels falls to ∼40%, however, in nontransmembrane-spanning regions such as the intracellular loop connecting domains 3 and 4 and the intracellular C-terminal extension (Soong et al., 1993; Schneider et al., 1994;Williams et al., 1994; Wheeler et al., 1995). Considering the high degree of overall homology between α1B and α1E and of these proteins with α1A, it is of considerable interest to note the large difference in the ability of the two channel types to be regulated by G-proteins. The regulation of N channels that we have observed in HEK293 cells closely resembles that frequently described in neurons (Hille, 1994). Little is known as yet, however, about the normal properties and functions of the Ca channels that are formed by expression of α1E, although the protein is widely expressed on the soma and dendrites of neurons throughout the brain (Williams et al., 1994; Volsen et al., 1995; Yokoyama et al., 1995). The experiments reported here suggest that direct G-protein regulation of these channels may be quite minimal. It should be noted that in our studies we found that G-proteins representative of all of the four major families were represented in HEK293 cells, and at least one member of each of these families was also overexpressed in our experiments. Thus, although it remains possible that α1E channels are normally regulated through an exceedingly restricted direct G-protein pathway that we have missed, this seems unlikely. It should be noted in this regard that although N channel inhibition tends to be mediated by PTX-sensitive G-proteins, there are several examples of non-PTX-sensitive G-proteins also producing this type of direct, membrane-delimited inhibition (Hille, 1994; Zhu and Ikeda, 1994). This implies that for N channels, many types of G-protein α subunits may be able to interact with the channel. Another interpretation of these observations would be that it is actually the β/γ subunits that produce the inhibition and that these may be released from various receptor-activated G-protein heterotrimers (Ikeda, 1996; Herlitze et al., 1996). Whichever mechanism is involved, it seems unlikely that any G-protein regulation of highly homologous α1E channels would be highly selective. In our experiments, we have also tried to produce regulation of α1E channels in several different ways. Thus, introduction of GTP-γ-S into cells would activate the entire complement of HEK293 cell G-proteins (Zhou et al., 1995; Zong et al., 1995). Furthermore, we also used somatostatin receptors (Law et al., 1993; Shapiro and Hille, 1993; Reisine and Bell, 1995) and κ opioid receptors (Shen and Crain, 1994; Avidor-Reiss et al., 1995; Ikeda et al., 1995; Lai et al., 1995; Tallent et al., 1995), both of which can activate a wide variety of G-proteins. Interestingly, all of these manipulations produced robust inhibition of α1Bchannels, which had the same ancillary subunit composition as the α1E channels. We conclude that it is unlikely that we failed to observe regulation of α1Echannels because of the fact that we failed to express the correct components for observing such regulation, if it did indeed exist.
The characteristics of inhibition of α1Bchannels observed in the present experiments are similar in many respects to those reported previously. For example, the inhibition displayed both voltage-dependent and -independent components (Bean, 1989; Hille, 1994; Diverse-Pierluissi and Dunlap, 1995;Diverse-Pierluissi et al., 1995). Whether this is attributable to activation of more than one G-protein-linked pathway, thereby producing more than one effect on the channels as suggested in the literature, is difficult to assess (Diverse-Pierluissi and Dunlap, 1995;Diverse-Pierluissi et al., 1995). Thus, HEK293 cells normally contained various G-proteins, even before the introduction of more of these molecules by transfection. Furthermore, both somatostatin and κ opioid receptors potentially can activate several of these G-proteins. The results, however, are also consistent with a single process. The effect of the depolarizing prepulse is assumed to lower the affinity of the interaction between the G-protein subunit(s) and the channel that allows unbinding (Boland and Bean, 1993). It is possible that rebinding of the G-protein might occur with a time course that is beyond the resolution of this study.
The present observations set some limitations on the localization of G-protein-binding to the α1Bchannel. Thus, there is a high degree of sequence identity between α1B and α1E in their 24 putative membrane-spanning domains (Soong et al., 1993; Schneider et al., 1994; Williams et al., 1994; Wheeler et al., 1995). On the other hand, other areas exhibit less homology. This is particularly true of the large intracellular loop between domains 2 and 3 and the N and C termini. Furthermore, the smaller intracellular loop between domains 1 and 2 also shows considerable divergence. It is interesting to note that this small loop is the site of interaction between the Ca channel α subunit and the small β subunit (Pragnell et al., 1994). Some studies have suggested that Ca channel β subunits and G-protein subunits may compete for a binding site on the Ca channel α1 subunit, although it is not clear whether such an interaction is competitive or allosteric (Berrow et al., 1995;Campbell et al., 1995b; Roche et al., 1995). It has been suggested further that the α subunits of Ca channels may possess GAP activity (Campbell et al., 1995a) and that the Ca channel α and β subunits cooperate in enhancing the GTPase activity of the G-protein α subunit. Such observations are interesting in light of recent studies showing that G-protein α and β/γ subunits both bind to GIRK-1, one of the G-protein-regulated K channels (Huang et al., 1995). These studies also suggested that GIRK-1 may also possess GAP activity (Slesinger et al., 1995). It is possible that a similar arrangement also applies to Ca channels. Resolution of this problem will require elucidation of precisely which G-protein subunits bind to and regulate Ca channels.
Footnotes
This work was supported by Public Health Service Grants DA02121, MH40165, DA02575, and NS33502. We thank Drs. Michael Harpold (SIBIA Inc.) and David Lodge (Eli Lilly) for providing the cell lines and for helpful discussions. We thank Dongjun Ren for her technical help.
Correspondence should be addressed to Richard J. Miller, Department of Pharmacological and Physiological Sciences, The University of Chicago, 947 East 58th Street (MC 0926), Chicago, IL 60637.
REFERENCES
- 1.Andersson S, Davis DL, Dahlback H, Jornvall H, Russell DW. Cloning, structure and expression of the mitochondrial cytochrome P-450 sterol 25-hydroxylase, a bile acid biosynthetic enzyme. J Biol Chem. 1989;264:8222–8229. [PubMed] [Google Scholar]
- 2.Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology, Wiley; New York: 1993. Transfection of DNA into eukaryotic cells. pp. 9.1.1–9.1.7. [Google Scholar]
- 3.Avidor-Reiss T, Zippel R, Levy R, Saya D, Ezra V, Barg J, Matus-Leibovitch N, Vogel Z. κ-opioid receptor transfected cell lines: modulation of adenylyl cyclase activity following acute and chronic opioid treatment. FEBS Lett. 1995;361:70–74. doi: 10.1016/0014-5793(95)00154-2. [DOI] [PubMed] [Google Scholar]
- 4.Bean BP. Neurotransmitter inhibition of neuronal calcium channels by changes in channel voltage dependence. Nature. 1989;340:153–156. doi: 10.1038/340153a0. [DOI] [PubMed] [Google Scholar]
- 5.Berrow NS, Campbell V, Fitzgerald EM, Brickley K, Dolphin AC. Antisense depletion of β subunits modulates the biophysical and pharmacological properties of neuronal calcium channels. J Physiol (Lond) 1995;482:481–491. doi: 10.1113/jphysiol.1995.sp020534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bleakman D, Bowman D, Bath CP, Brust PF, Johnson EC, Deal CR, Miller RJ, Ellis SB, Harpold MM, Hans M, Grantham CJ. Characteristics of a human N type calcium channel expressed in HEK293 cells. Neuropharmacology. 1995;34:753–765. doi: 10.1016/0028-3908(95)00078-k. [DOI] [PubMed] [Google Scholar]
- 7.Boland LM, Bean BP. Modulation of N type calcium channels in bullfrog sympathetic neurons by luteinizing hormone releasing hormone, kinetics and voltage dependence. J Neurosci. 1993;13:516–533. doi: 10.1523/JNEUROSCI.13-02-00516.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Campbell V, Berrow N, Brickley K, Page K, Wade R, Dolphin AC. Voltage dependent calcium channel β subunits in combination with α1 subunits have a GTPase activating effect to promote the hydrolysis of GTP by Gao in rat frontal cortex. FEBS Lett. 1995a;370:135–140. doi: 10.1016/0014-5793(95)00813-o. [DOI] [PubMed] [Google Scholar]
- 9.Campbell V, Berrow EM, Fitzgerald EM, Brickley K, Dolphin AC. Inhibition of the interaction of G-protein Go with calcium channels by the calcium channel β subunit in rat neurons. J Physiol (Lond) 1995b;485:365–372. doi: 10.1113/jphysiol.1995.sp020735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Diverse-Pierluissi M, Dunlap K. Interaction of convergent pathways that inhibit N type calcium currents in sensory neurons. Neuroscience. 1995;65:477–483. doi: 10.1016/0306-4522(94)00476-l. [DOI] [PubMed] [Google Scholar]
- 11.Diverse-Pierluissi M, Goldsmith PK, Dunlap K. Transmitter mediated inhibition of N type calcium channels in sensory neurons involves multiple GTP binding proteins and subunits. Neuron. 1995;14:191–200. doi: 10.1016/0896-6273(95)90254-6. [DOI] [PubMed] [Google Scholar]
- 12.Dolphin AC. Voltage dependent calcium channels and their modulation by neurotransmitters and G-proteins. Exp Physiol. 1995;80:1–36. doi: 10.1113/expphysiol.1995.sp003825. [DOI] [PubMed] [Google Scholar]
- 13.Ewald DA, Pang IH, Sternweis PC, Miller RJ. Differential G protein-mediated coupling of neurotransmitter receptors to Ca2+ channels in rat dorsal root ganglion neurons in vitro. Neuron. 1989;2:1185–1193. doi: 10.1016/0896-6273(89)90185-2. [DOI] [PubMed] [Google Scholar]
- 14.Fujii Y, Gonoi T, Yamada T, Chihara K, Inagaki N, Seino S. Somatostatin receptor subtype SSTR2 mediates the inhibition of high voltage activated calcium channels by somatostatin and its analogue SMS 201–995. FEBS Lett. 1994;355:117–120. doi: 10.1016/0014-5793(94)01159-1. [DOI] [PubMed] [Google Scholar]
- 15.Golard A, Siegelbaum SA. Kinetic basis for the voltage-dependent inhibition of N-type calcium current by somatostatin and norepinephrine in chick sympathetic neurons. J Neurosci. 1993;13:3884–3894. doi: 10.1523/JNEUROSCI.13-09-03884.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high resolution current recording from cells and cell-free membrane patches. Pflügers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- 17.Herlitze S, Garcia DE, Mackie K, Hille B, Scheuer T, Catterall WA. Modulation of Ca2+ channels by G-protein βγ subunits. Nature. 1996;380:258–262. doi: 10.1038/380258a0. [DOI] [PubMed] [Google Scholar]
- 18.Hille B. Modulation of ion channels by G-protein coupled receptors. Trends Neurosci. 1994;17:531–535. doi: 10.1016/0166-2236(94)90157-0. [DOI] [PubMed] [Google Scholar]
- 19.Huang CL, Slesinger PA, Casey PJ, Jan YN, Jan LY. Evidence that direct binding of Gβγ to the GIRK-1 G-protein gated inwardly rectifying K channel is important for channel activation. Neuron. 1995;15:1133–1143. doi: 10.1016/0896-6273(95)90101-9. [DOI] [PubMed] [Google Scholar]
- 20.Ikeda SR. Voltage-dependent modulation of N-type calcium channels by G-protein βγ subunits. Nature. 1996;380:255–258. doi: 10.1038/380255a0. [DOI] [PubMed] [Google Scholar]
- 21.Ikeda SR, Schofield GG. Somatostatin blocks a calcium current in rat sympathetic neurons. J Physiol (Lond) 1989;409:221–240. doi: 10.1113/jphysiol.1989.sp017494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ikeda K, Kobayashi T, Ichikawa T, Usui H, Kumanishi T. Functional couplings of the δ and the κ opioid receptors with the G-protein activated K channel. Biochem Biophys Res Commun. 1995;208:302–308. doi: 10.1006/bbrc.1995.1338. [DOI] [PubMed] [Google Scholar]
- 23.Kanemasa T, Asakura K, Ninomiya M. κ-opioid agonist U50488 inhibits P type Ca channels by two mechanisms. Brain Res. 1995;702:207–212. doi: 10.1016/0006-8993(95)01049-3. [DOI] [PubMed] [Google Scholar]
- 24.Lai HWL, Minami M, Satoh M, Wong YH. Gz coupling to the rat κ-opioid receptor. FEBS Lett. 1995;360:97–99. doi: 10.1016/0014-5793(95)00088-q. [DOI] [PubMed] [Google Scholar]
- 25.Law SF, Yasuda K, Bell GI, Reisine T. Giα3 and Goα selectively associate with the cloned somatostatin receptor subtype SSTR2. J Biol Chem. 1993;268:10721–10727. [PubMed] [Google Scholar]
- 26.Lipscombe D, Kongsamut S, Tsien RW. Alpha-adrenergic inhibition of sympathetic neurotransmitter release mediated by modulation of N-type calcium-channel gating. Nature. 1989;340:639–642. doi: 10.1038/340639a0. [DOI] [PubMed] [Google Scholar]
- 27.Miller RJ. The receptor mediated regulation of calcium channels and neurotransmitter release. FASEB J. 1990;4:3291–3300. [PubMed] [Google Scholar]
- 28.Mintz IM, Bean BP. GABA-B receptor inhibition of P type Ca channels in central neurons. Neuron. 1993;10:889–998. doi: 10.1016/0896-6273(93)90204-5. [DOI] [PubMed] [Google Scholar]
- 29.Perez-Reyes E, Yuan W, Wei X, Bers DM. Regulation of cloned L-type cardiac calcium channels by cyclic AMP dependent protein kinase. FEBS Lett. 1994;342:119–123. doi: 10.1016/0014-5793(94)80484-2. [DOI] [PubMed] [Google Scholar]
- 30.Pragnell M, De Waard M, Mori Y, Tanabe T, Snutch TP, Campbell KP. Calcium channel β subunit binds to a conserved motif in the 1–11 cytoplasmic linker of the α1subunit. Nature. 1994;368:67–70. doi: 10.1038/368067a0. [DOI] [PubMed] [Google Scholar]
- 31.Reisine T, Bell G. Molecular properties of somatostatin receptors. Neuroscience. 1995;67:777–790. doi: 10.1016/0306-4522(95)00072-q. [DOI] [PubMed] [Google Scholar]
- 32.Rhim H, Miller RJ. Opioid receptors modulate diverse types of calcium channels in the nucleus tractus solitarius of the rat. J Neurosci. 1994;14:7608–7615. doi: 10.1523/JNEUROSCI.14-12-07608.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Robishaw JD, Russell DW, Harris BA, Smigel MD, Gilman AG. Deduced primary structure of the α-subunit of the GTP-binding stimulatory protein of adenylate cyclase. Proc Natl Acad Sci USA. 1986;83:1251–1255. doi: 10.1073/pnas.83.5.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Roche JP, Anantharam V, Treistman SN. Abolition of G-protein inhibition of α1A and α1B calcium channels by coexpression of the β3 subunit. FEBS Lett. 1995;371:43–46. doi: 10.1016/0014-5793(95)00860-c. [DOI] [PubMed] [Google Scholar]
- 35.Sambrook J, Fritsch EF, Maniatis T. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: 1989. Molecular cloning: a laboratory manual. . [Google Scholar]
- 36.Schneider T, Wei X, Olcese R, Costantin JL, Neely A, Palade P, Perez-Reyes E, Qin N, Zhou J, Crawford GD, Smith RG, Appel SH, Stefani E, Birnbaumer L. Molecular analysis and functional expression of the human type E neuronal Ca channel α1 subunit. Recept Channels. 1994;2:255–270. [PubMed] [Google Scholar]
- 37.Shapiro MS, Hille B. Substance P and somatostatin inhibit calcium channels in rat sympathetic neurons via different G protein pathways. Neuron. 1993;10:11–20. doi: 10.1016/0896-6273(93)90237-l. [DOI] [PubMed] [Google Scholar]
- 38.Shen KF, Crain SM. Nerve growth factor rapidly prolongs the action potential of mature sensory ganglion neurons in culture, and this effect requires activation of Gs coupled excitatory κ-opioid receptors on these cells. J Neurosci. 1994;14:5570–5579. doi: 10.1523/JNEUROSCI.14-09-05570.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Slesinger PA, Reuveny E, Jan YN, Jan LY. Identification of structural elements involved in G-protein gating of the GIRK-1 potassium channel. Neuron. 1995;15:1145–1156. doi: 10.1016/0896-6273(95)90102-7. [DOI] [PubMed] [Google Scholar]
- 40.Soong TW, Stea A, Hodson CD, Dubel SJ, Vincent SR, Snutch TP. Structure and functional expression of a member of the low voltage activated calcium channel family. Science. 1993;260:1133–1136. doi: 10.1126/science.8388125. [DOI] [PubMed] [Google Scholar]
- 41.Sternweis PC. The active role of β/γ by in signal transduction. Curr Opin Cell Biol. 1994;6:198–203. doi: 10.1016/0955-0674(94)90136-8. [DOI] [PubMed] [Google Scholar]
- 42.Tallent M, Dichter MA, Bell GI, Reisine T. The cloned kappa opioid receptor couples to an N-type calcium current in undifferentiated PC-12 cells. Neuroscience. 1995;63:1033–1040. doi: 10.1016/0306-4522(94)90570-3. [DOI] [PubMed] [Google Scholar]
- 43.Volsen SG, Day NC, McCormack AC, Smith W, Craig PJ, Beattie R, Incep G, Show PJ, Ellis SB, Gillespie A, Harpold MM, Lodge D. The expression of neuronal voltage dependent Ca channels in human cerebellum. Mol Brain Res. 1995;34:217–232. doi: 10.1016/0169-328x(95)00234-j. [DOI] [PubMed] [Google Scholar]
- 44.Wheeler DB, Randall A, Tsien RW. Roles of N type and Q type Ca channels in supporting hippocampal synaptic transmission. Science. 1994;264:107–111. doi: 10.1126/science.7832825. [DOI] [PubMed] [Google Scholar]
- 45.Wheeler DB, Randall A, Sather WA, Tsien RW. Neuronal calcium channels encoded by the α1A subunit and their contribution to excitatory synaptic transmission in the CNS. Prog Brain Res. 1995;105:65–78. doi: 10.1016/s0079-6123(08)63284-7. [DOI] [PubMed] [Google Scholar]
- 46.Wickman K, Clapham DE. Ion channel regulation by G-proteins. Physiol Rev. 1995;75:865–885. doi: 10.1152/physrev.1995.75.4.865. [DOI] [PubMed] [Google Scholar]
- 47.Williams ME, Brust PF, Feldman DH, Patthi S, Simerson S, Maroufi A, McCue AF, Velicelebi G, Ellis SB, Harpold MM. Structure and functional expression of an ω-conotoxin sensitive human N-type calcium channel. Science. 1992;257:389–395. doi: 10.1126/science.1321501. [DOI] [PubMed] [Google Scholar]
- 48.Williams SM, Marubio LM, Deal CR, Hans M, Brust PF, Philipson LH, Miller RJ, Johnson EC, Harpold MM, Ellis SB. Structure and functional characterization of neuronal α1Ecalcium channels subtypes. J Biol Chem. 1994;269:22347–22357. [PubMed] [Google Scholar]
- 49.Yokoyama CT, Westenbroek RE, Hell JW, Soong T-W, Snutch TP, Catterall WA. Biochemical properties and subcellular distribution of the neuronal class E calcium channel α1subunit. J Neurosci. 1995;15:6419–6432. doi: 10.1523/JNEUROSCI.15-10-06419.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang J-F, Randall AD, Ellinor PT, Horne WA, Sather WA, Tanabe Schwarz TL, Tsien RW. Distinctive pharmacology and kinetics of cloned neuronal Ca2+ channels and their possible counterparts in mammalian CNS neurons. Neuropharmacology. 1993;32:1075–1088. doi: 10.1016/0028-3908(93)90003-l. [DOI] [PubMed] [Google Scholar]
- 51.Zhou J, Zong S, Tanabe T. Modulation of cloned neuronal calcium channels through membrane delimited pathway. Biochem Biophys Res Commun. 1995;208:485–491. doi: 10.1006/bbrc.1995.1364. [DOI] [PubMed] [Google Scholar]
- 52.Zhu Y, Ikeda SR. VIP inhibits N type Ca channels of sympathetic neurons via a pertussis toxin insensitive but cholera toxin sensitive pathway. Neuron. 1994;13:657–669. doi: 10.1016/0896-6273(94)90033-7. [DOI] [PubMed] [Google Scholar]
- 53.Zong S, Yassin M, Tanabe T. G-protein modulation of α1A (P/Q) type calcium channel expressed in GH3 cells. Biochem Biophys Res Commun. 1995;215:302–308. doi: 10.1006/bbrc.1995.2466. [DOI] [PubMed] [Google Scholar]