

Abstract
The fetal myocardium is known to be sensitive to hemodynamic load, responding to systolic overload with cellular hypertrophy, proliferation, and accelerated maturation. However, the fetal cardiac growth response to primary volume overload is unknown. We hypothesized that increased venous return would stimulate fetal cardiomyocyte proliferation and terminal differentiation, particularly in the right ventricle (RV). Vascular catheters and pulmonary artery flow probes were implanted in 16 late-gestation fetal sheep: a right carotid artery-jugular vein (AV) fistula was surgically created in nine fetuses, and sham operations were performed on seven fetuses. Instrumented fetuses were studied for 1 wk before hearts were dissected for component analysis or cardiomyocyte dispersion for cellular measurements. Within 1 day of AV fistula creation, RV output was 20% higher in experimental than sham fetuses (P < 0.0001). Circulating atrial natriuretic peptide levels were elevated fivefold in fetuses with an AV fistula (P < 0.002). On the terminal day, RV-to-body weight ratios were 35% higher in the AV fistula group (P < 0.05). Both left ventricular and RV cardiomyocytes grew longer in fetuses with an AV fistula (P < 0.02). Cell cycle activity was depressed by >50% [significant in left ventricle (P < 0.02), but not RV (P < 0.054)]. Rates of terminal differentiation were unchanged. Based on these studies, we speculate that atrial natriuretic peptide suppressed fetal cardiomyocyte cell cycle activity. Unlike systolic overload, fetal diastolic load appears to drive myocyte enlargement, but not cardiomyocyte proliferation or maturation. These changes could predispose to RV dysfunction later in life.
NEW & NOTEWORTHY Adaptation of the fetal heart to changes in cardiac load allows the fetus to maintain adequate blood flow to its systemic and placental circulations, which is necessary for the well-being of the fetus. Addition of arterial-venous fistula flow to existing venous return increased right ventricular stroke volume and output. The fetal heart compensated by cardiomyocyte elongation without accelerated cellular maturation, while cardiomyocyte proliferation decreased. Even transient volume overload in utero alters myocardial structure and cardiomyocyte endowment.
Keywords: atrial natriuretic peptide, cardiomyocyte, fetus, right ventricle, volume overload
INTRODUCTION
In the adult, systolic and diastolic load are powerful modulators of cardiac hypertrophy (8, 45). Cardiac hypertrophy is a result of complex changes in structure and function of cells comprising the myocardium (5, 6, 10, 27, 29). While pressure and volume overload result in morphologically distinct forms of hypertrophy in the adult heart (5, 9, 29), less is known about the effects of pressure and volume load on the fetal heart. Segar and associates showed that increased right ventricular (RV) systolic pressure load increased heart mass primarily of the RV (40). We previously demonstrated that the increase in fetal heart mass in response to increased RV systolic pressure load results from both hyperplastic and hypertrophic growth (2). We and other researchers also showed that increased fetal arterial pressure increases both RV and left ventricular (LV) mass through intracellular signaling pathways that regulate fetal cardiomyocyte proliferation, cellular enlargement, and accelerated terminal differentiation (2, 21, 31, 32, 34, 38–42). Few studies, however, have examined the effects of increased diastolic load on fetal cardiac growth.
Understanding how increased volume load regulates fetal heart growth is clinically relevant. Fetal volume overload occurs in fetuses with sacrococcygeal teratomas, valvular defects, rhesus isoimmunization, and arteriovenous malformations (16, 44). The morbidity of these defects is severe and represents highly important clinical conditions to be better understood. Volume loading in human fetuses is associated with elevated circulating natriuretic peptides (48) and hydrops fetalis (26). Iatrogenic volume overload from transfusions can lead to fetal demise (43). Our ability to provide appropriate therapy in fetuses with these conditions is limited by our lack of understanding of the physiology of the volume-loaded fetal heart. In the 1970s, Fishman and colleagues performed the inverse experiment by obstructing fetal LV filling (15). They observed that LV growth was inhibited, reminiscent of hypoplastic left heart syndrome, and concluded that adequate filling of the ventricle is required for normal growth. However, the hypoplastic left heart condition would lead to increased volume load of the RV.
Jouannic and colleagues constructed a side-to-side carotid arterial-to-jugular vein (AV) anastomosis to increase blood flow through the right heart to the pulmonary artery (PA) at 80% gestation and found PA remodeling with associated abnormal pulmonary vascular vasoreactivity (24, 25). Despite the interesting pulmonary pathology linked to volume overload in the fetus, the effects of volume overload on the heart have not been well studied. We hypothesized that chronically increased right heart volume load would lead to increased RV stroke volume (SV), RV cardiomyocyte proliferation, RV cardiomyocyte hypertrophy, and accelerated terminal differentiation in the fetal heart. We tested this hypothesis using a side-to-side AV anastomosis to increase volume load to the near-term fetal heart in sheep.
MATERIALS AND METHODS
Animal model.
Animal experiments were carried out with approval of the Institutional Animal Care and Use Committee at Oregon Health & Science University, which is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International and compliant with the Animal Welfare Act and Public Health Service policy. Eighteen timed-bred pregnant ewes of mixed Western breeds were obtained from a local supplier. At an average of 127 days gestation (range 126–133 days, full term = 147 days), sterile surgery was conducted. Anesthesia was induced with intravenous diazepam (0.13 mg/kg) and ketamine (5 mg/kg), the ewe was intubated, and anesthesia was maintained using 1–2% isoflurane in a carrier gas mixture of oxygen and nitrous oxide (2:1). Sterile surgery was performed as described previously to expose the fetal head and chest (2, 46). The left chest was opened at the fourth intercostal space, and the pericardium overlying the upper aspect of the heart was incised. The PA was dissected free from surrounding tissues. A 1.3-mm-outer-diameter polyvinyl catheter (V-5, Bolab, Lake Havasu City, AZ) was placed in the proximal PA just beyond the pulmonic valve. An appropriately sized ultrasonic flow probe (Transonic Systems, Ithaca, NY) was then placed around the main PA distal to the PA catheter. A 1.3-mm-outer-diameter polyvinyl catheter (Bolab) was placed in the right atrium. The free edges of the pericardium were loosely approximated, and the fetal chest was closed in anatomic layers.
The right carotid artery and right internal jugular vein were dissected free and isolated atraumatically using vessel loops. Occasionally occurring branches in the area of the anastomosis were ligated. The carotid artery and internal jugular vein were opened longitudinally and flushed with heparinized saline to remove all blood. Hemodynamic measurements were obtained in all fetuses using the methods described below before and after opening of the AV fistula. The same measurements were made in the sham (control) fetuses. A 1.5-cm side-to-side anastomosis was then fashioned using continuous 7-0 proline (Ethicon) suture. The clamps were then sequentially removed (vein, proximal artery, and distal artery), and any bleeding was controlled with thrombin-soaked gelfoam or direct suture. The carotid artery and jugular vein just distal to the anastomosis were ligated with 2-0 silk. A widely patent anastomosis was confirmed by the presence of a palpable “thrill” before closure of the neck incision. In the sham-operated animals, identical chest instrumentation and neck dissection were performed. The carotid artery and the internal jugular vein were ligated, and no fistula was created. The pressure transducers were zeroed to the right atrial level by direct inspection.
The fetus was then returned to the uterus, a multi-side-hole catheter was placed in the amniotic space, and all catheters and cables were exteriorized and secured to a cloth pouch on the ewe’s flank. The hysterotomy and abdomen were then closed in anatomic layers. Penicillin G (1 × 107 units; Bristol-Meyers Squibb, Princeton, NJ) was instilled into the amniotic space at the conclusion of the procedure, anesthesia was terminated, and the ewe was allowed to recover. Arterial blood samples for pH, blood gas, hematocrit, hemoglobin, and oxygen content analysis, as well as hemodynamic measurements, as described below, were taken from all fetuses during the surgical procedure. Plasma samples for analysis of atrial natriuretic peptide (ANP) levels were taken from a subset of fetuses (n = 5 AV fistula and 4 sham). Pressure transducers were zeroed to the right atrial level by direct inspection.
Experimental protocol.
Hemodynamic measurements were obtained in all animals during the initial surgical procedure, as well as on postoperative days 4 and 7. Hydrostatic pressures from the right atrium and PA were measured using BioPac pressure transducers (Abbott Laboratories, Chicago, IL), calibrated using a mercury manometer, and recorded using a PowerLab system (ADInstruments, Colorado Springs, CO). PA flow was also recorded to determine RV output and SV, and heart rate was calculated from the arterial waveform. All vascular pressures were referenced to amniotic fluid pressure. SV represents the integral of the RV output for each beat.
Fetal arterial blood samples were obtained in all animals during the initial surgical procedure and on postoperative days 4 and 7. Arterial pH, partial pressure of carbon dioxide, partial pressure of oxygen, oxygen content, and hematocrit were determined. Arterial blood gas determinations were made using a blood-gas analyzer (model 1306, Instrumentation Laboratory, Bedford, MA). Arterial oxygen content was determined using a CO-oximeter (model 382, Instrumentation Laboratory). Whole blood (3 ml) was separated by centrifugation, and plasma was frozen for analysis of ANP levels by radioimmunoassay (Bachem, Torrance, CA).
After study on day 7, ewes were euthanized with a pentobarbital euthanasia solution and fetal hearts were arrested in diastole with a saturated solution of potassium chloride. A standardized protocol was used for cardiac dissection (23). Hearts were used for enzymatic isolation of cardiomyocytes (n = 5 in each group) or dissected into component parts and frozen in liquid nitrogen (n = 4 in each group; control group includes two twin fetuses exposed to anesthesia but not catheterized). Fistula patency and catheter position were confirmed by dissection.
Cardiomyocyte analysis.
Fetal cardiomyocytes were dissociated and fixed with 1% formaldehyde made fresh in phosphate-buffered saline (pH 7.4), and dimensions of ≥100 cells were measured using calibrated image analysis software (Optimas, Seattle, WA) at ×400 magnification (Zeiss Axiophot, Bartels and Stout, Bellevue, WA), as previously described (2, 22, 23, 46). In a separate sample of ≥300 myocytes, the percentage of myocytes that were binucleated was measured. Myocytes in the cell cycle (≥300 counted) were labeled immunohistochemically with the nuclear antigen Ki-67 and expressed as percentage of mononucleated cardiomyocytes counted.
Statistical analysis.
Because of effects of maternal and fetal anesthesia on cardiovascular and placental function, values from surgery on day 0 were analyzed separately from values obtained on subsequent study days. Data collected during surgery were compared between groups by Student’s t-test (Prism 6.0, GraphPad Software, San Diego, CA). Physiological data collected intermittently thereafter were compared by mixed-model two-way ANOVA, followed by multiple comparison with Sidak’s multiple comparison or Bonferroni’s correction (if significant variation was detected in one or both factors, respectively). Two-way ANOVA was used to compare group variances over time. Variables measured on every study day were subjected to linear regression. Ages, weights, and cardiomyocyte values were compared by Student’s t-test. Data are presented as means ± SE. Significance was determined at P < 0.05.
RESULTS
Gestational ages were similar for the sham and AV fistula groups (135 ± 1 and 135 ± 2 days, respectively). Fetal weights were similar for the sham and AV fistula groups (4.0 ± 0.6 and 4.2 ± 0.6 kg, respectively). There were no systematic differences in blood gas or oxygen content values between groups (Table 1).
Table 1.
Arterial pH, blood gases, hematocrit, hemoglobin, and oxygen content
|
Day 0 |
Day 4 |
Day 7 |
||||
|---|---|---|---|---|---|---|
| Sham | AV fistula | Sham | AV fistula | Sham | AV fistula | |
| pH | 7.29 ± 0.02 | 7.29 ± 0.01 | 7.36 ± 0.01 | 7.36 ± 0.01 | 7.35 ± 0.01 | 7.35 ± 0.01† |
| Po2, mmHg | 22 ± 2 | 22 ± 1 | 18 ± 1 | 18 ± 1 | 17 ± 1 | 17 ± 1 |
| Pco2, mmHg | 52 ± 1 | 52 ± 2 | 50 ± 1 | 50 ± 1 | 54 ± 1† | 54 ± 2 |
| Hematocrit, % | 39 ± 2 | 39 ± 2 | 30 ± 2 | 36 ± 2* | 34 ± 1 | 34 ± 1 |
| Hemoglobin, ml/dl | 13.7 ± 1.0 | 13.7 ± 0.7 | 10.7 ± 0.6 | 10.7 ± 0.6 | 10.9 ± 0.7 | 10.9 ± 0.6 |
| Oxygen content, ml/dl | 9.3 ± 1.1 | 9.3 ± 0.4 | 7.3 ± 0.9 | 7.3 ± 0.4 | 6.4 ± 0.6 | 6.4 ± 0.8 |
Values are means ± SE. AV, right carotid artery-jugular vein.
Different within-day from sham.
Different within group from day 4.
Before construction of the AV fistula, there were no significant differences in right atrial pressure, aortic pressure, heart rate, RV output, or RV SV between the sham and the AV fistula group (Table 2). In the sham group, right atrial pressure, RV output, RV SV, and heart rate did not change after ligation of the carotid artery and the jugular vein (Table 2). Right atrial pressure in the AV fistula group was 3.2 ± 2.1 before and 4.1 ± 2.3 mmHg after the fistula was opened (Table 2). PA pressure was 44.3 ± 6.8 before and 42.2 ± 8.0 mmHg after the fistula was opened. RV output was 390 ± 130 before and 488 ± 177 ml/min after the fistula was opened (Table 2). RV stroke volume was 2.4 ± 0.8 before and 2.8 ± 0.9 ml/beat after the AV fistula was opened. Heart rate was 166 ± 24 before and 175 ± 20 beats/min after the fistula was opened. Figure 1 illustrates RV output, RV SV, and heart rate for the sham and AV fistula groups over the 7-day study period. In the AV fistula group, RV output increased 25% and RV SV increased 17% when the AV fistula was opened. There was no change in mean RV output or SV when the carotid artery and the jugular vein were ligated in the sham group. The greater RV output and SV in the AV fistula group were maintained over the 7-day study period, as indicated by the parallel nature of the lines for the sham and AV fistula groups in Fig. 1. As expected in a growing fetus, PA flow increased over the period of study (P < 0.001; Fig. 1A); the growth-related increases in RV output and RV SV were not different between the groups.
Table 2.
Hemodynamic parameters for sham and AV fistula groups at baseline, day 4, and day 7
|
Day 0 |
||||||||
|---|---|---|---|---|---|---|---|---|
| Preshunt |
Postshunt |
Day 4 |
Day 7 |
|||||
| Sham | AV fistula | Sham | AV fistula | Sham | AV fistula | Sham | AV fistula | |
| RAP, mmHg | 3.9 ± 1.8 | 3.2 ± 2.1 | 3.6 ± 2.0 | 4.1 ± 2.3 | 4.1 ± 2.2 | 3.1 ± 1.9 | 3.3 ± 1.3 | 3.9 ± 1.4 |
| PAP, mmHg | 42.7 ± 5.1 | 44.3 ± 6.8 | 42.3 ± 6.1 | 42.2 ± 8.0 | 46.1 ± 4.1 | 46.3 ± 3.5 | 46.5 ± 3.5 | 47.5 ± 6.4 |
| HR, beats/min | 167 ± 29 | 168 ± 24 | 166 ± 28 | 175 ± 20 | 170 ± 24 | 166 ± 13 | 160 ± 9 | 162 ± 9 |
| RV output, ml/min | 350 ± 138 | 390 ± 130 | 350 ± 119 | 488 ± 177 | 671 ± 233 | 919 ± 174 | 785 ± 228 | 1037 ± 184 |
| RV SV, ml/beat | 2.0 ± 0.8 | 2.4 ± 0.8 | 2.0 ± 0.6 | 2.8 ± 0.9 | 4.0 ± 5.6 | 5.6 ± 1.0 | 4.9 ± 1.2 | 6.4 ± 1.1 |
Values are means ± SE. Preshunt, immediately before right carotid artery-jugular vein (AV) fistula was opened; postshunt, immediately after shunt was opened (AV fistula group) or after carotid artery and jugular vein were tied off (sham group); RAP, right atrial pressure; PAP, mean pulmonary arterial pressure; HR, heart rate; RV, right ventricle; SV, stroke volume.
Fig. 1.
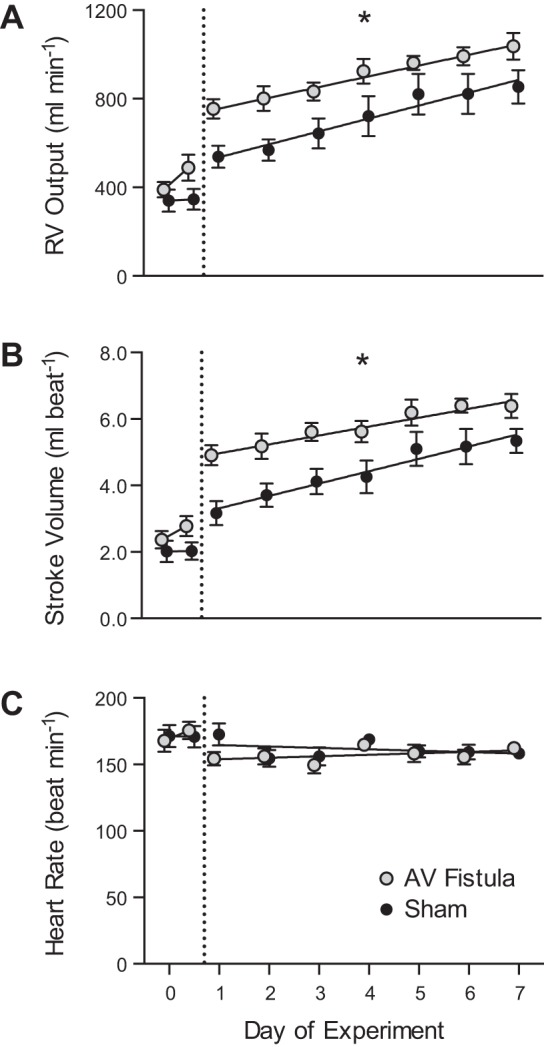
Hemodynamics in experimental fetal right carotid artery-jugular vein (AV) fistula. A–C: right ventricular (RV) output, stroke volume, and heart rate in fetuses with AV fistula and sham controls. Points to the left of the dashed vertical line represent data collected during surgery (day 0) in the anesthetized state. For the AV fistula group, the day 0 point on the left is with the fistula closed, and the point on the right is with the fistula open. For the sham group, the day 0 point on the left is with the fistula closed, and the point on the right is with the fistula ligated. Data for days 1–7 were obtained from unanesthetized fetuses. Values are means ± SE. All RV output and RV stroke volume values on days 1–7 were higher for the AV fistula than the sham group: *P < 0.0001. Comparative slopes between AV fistula and sham fetuses for the relationship for RV output, stroke volume, and heart rate study as a function of time were not different: P = 0.53, P = 0.28, and P = 0.16, respectively.
Circulating ANP levels were not different between the groups on the day of surgery (day 0; Fig. 2). ANP levels increased in the AV fistula group, such that they were fivefold greater than in the sham group on days 4 and 7 (P < 0.05).
Fig. 2.
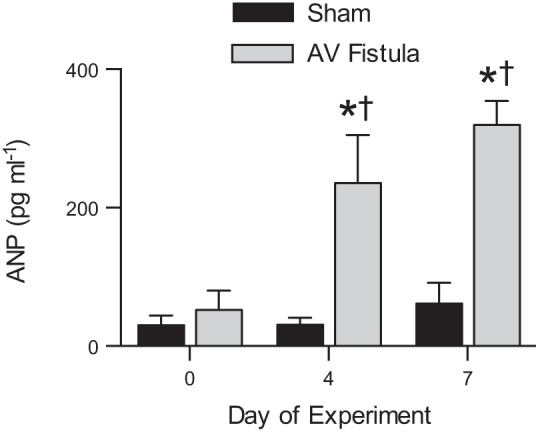
Circulating atrial natriuretic peptide (ANP) levels in experimental fetal right carotid artery-jugular vein (AV) fistula. Plasma ANP levels were not different between groups at baseline (day 0) but were 5-fold greater in the AV fistula than the sham group on days 4 and 7. Values are means ± SE. *P < 0.05 vs. same-day sham group. †P < 0.05 vs. day 0 within group.
Heart weights were not different in the sham compared with the AV fistula group (Table 3). RV free wall weight-to-fetal weight ratio (Table 3) was greater in the AV fistula than sham fetuses (P < 0.05). Mononucleated and binucleated myocytes from both ventricles were longer in the AV fistula than the sham group (P < 0.02; Table 4). In contrast to lengths, widths of myocytes from the AV fistula group were not different; binucleation was not significantly changed by volume load (Fig. 3A). The presence of a fetal AV fistula was associated with a 50% reduction in cell cycle activity in the LV (P < 0.02; Fig. 3B). Although greater in magnitude of effect, the 66% lower RV cell cycle activity in the AV fistula group did not achieve strict statistical significance (P = 0.054).
Table 3.
Fetal body and heart weights
| Sham | AV Fistula | |
|---|---|---|
| Terminal age, days gestation | 134.6 ± 0.3 | 135.2 ± 0.2 |
| Body wt, kg | 4.1 ± 0.2 | 4.0 ± 0.2 |
| Heart wt, kg | 28.8 ± 2.1 | 31.4 ± 2.0 |
| Normalized to body wt | ||
| Heart | 7.0 ± 0.3 | 7.8 ± 0.4 |
| RV free wall | 1.7 ± 0.2 | 2.3 ± 0.1* |
| LV free wall | 1.7 ± 0.1 | 1.9 ± 0.1 |
| Interventricular septum | 1.2 ± 0.0 | 1.3 ± 0.1 |
| Right atrial free wall | 0.4 ± 0.0 | 0.5 ± 0.0 |
| Left atrial free wall | 0.4 ± 0.0 | 0.5 ± 0.1 |
Values are means ± SE. AV, right carotid artery-jugular vein (fistula); RV, right ventricle; LV, left ventricle.
P < 0.05 vs. sham.
Table 4.
Cardiomyocyte dimensions
| Sham | AV Fistula | |
|---|---|---|
| Binucleate length, mm | ||
| RV | 88 ± 0 | 99 ± 3* |
| LV | 83 ± 1 | 92 ± 2* |
| Mononucleate length, mm | ||
| RV | 68 ± 1 | 75 ± 1* |
| LV | 65 ± 1 | 70 ± 2* |
| Binucleate width, mm | ||
| RV | 16 ± 1 | 16 ± 0 |
| LV | 13 ± 1 | 14 ± 1 |
| Mononucleate width, mm | ||
| RV | 14 ± 1 | 14 ± 0 |
| LV | 11 ± 0 | 13 ± 0 |
Values are means ± SE. AV, right carotid artery-jugular vein (fistula); RV, right ventricle; LV, left ventricle.
P < 0.02 vs. sham.
Fig. 3.

Cardiomyocyte maturation and cell cycle activity. A: terminal differentiation, as measured by binucleation, was not different between the right carotid artery-jugular vein (AV) and the sham group. B: left ventricular (LV) cell cycle activity, required for proliferation and binucleation, was 60% lower in AV fetuses. RV, right ventricle. Values are means ± SE. *P < 0.02 between groups.
DISCUSSION
This study was designed to test the hypothesis that chronically increased intrauterine right heart volume load leads to RV cardiomyocyte proliferation, hypertrophy, and accelerated terminal differentiation. Understanding how cardiac load affects fetal cardiac growth is important for both the fetus and the neonate as they prepare for their life ahead. The anatomic and physiological response of the fetal RV to increased diastolic load may place the heart at a disadvantage in coping with increased systolic load later in life. This is important, because hearts compromised at birth can be compromised for life (3, 4). For example, we previously showed that fetal anemia-induced augmentation of coronary blood flow persists until adulthood, even though the adult sheep has a normal hematocrit (12). We also showed that fetal anemia leads to augmented contractile response to hypoxic stress in adulthood (7). However, these changes may not always carry an advantage. Contrary to our expectation, fetal anemia results in larger myocardial infarctions in adult sheep, even though hematocrit is normal and coronary vasodilatory reserve is increased (49).
Fetal hemodynamic response to the AV fistula.
We observed a small increase in RV SV and RV output immediately on opening the AV fistula in the fetuses in the AV fistula group. On day 1 the difference between RV output and SV for the AV fistula group compared with the sham group reached statistical significance (Fig. 1, A and B). The increase in RV SV was superimposed on the normal daily growth of the RV, as illustrated by the parallel nature of the data comparing the AV fistula with the sham group in Fig. 1B. Differences between the AV fistula and the sham group were about the same for days 2–7. This suggests that the volume-loaded fetal RV is capable of continued growth at the expected rate as gestation continues. It is likely that a continuous increase in cardiac output is required to keep up with increasing demands due to somatic growth. These processes allowed RV growth, which led to a parallel increase in SV of fetuses in the AV fistula and sham groups.
The mechanisms that caused the small initial increase in RV SV at the time of surgery are not exactly known, since we did not measure RV end-diastolic and end-systolic volumes because of technical challenges under sterile conditions. In the unanesthetized fetus, RV and LV function are near the break point for the filling pressure-SV relationship, suggesting little if any preload reserve in the unanesthetized fetus (18, 30, 36, 47). In the anesthetized fetus, a very small undetected preload effect, too small to distinguish in our pressure measurements due to the steepness of the ascending limb of the fetal right atrial pressure-SV relationship, could play a role (35). The anesthetized fetal heart could have preload reserve, such that the increase in right heart filling associated with opening of the AV fistula would allow a larger end-diastolic volume and account for the accompanying increased SV. The anesthetized fetal heart could also have contractile reserve, allowing a smaller end-systolic volume and, thus, a greater SV, given the same end-diastolic volume, accounting for the increase in SV in the anesthetized fetus during surgery. Further study is needed to address these speculations.
However, an increase in preload is supported by our finding of a fivefold increase in ANP by experimental day 7. Because the heart ventricles in the fetus are able to produce ANP, we suspect that both the atria and the ventricles (11) increased their ANP production rates due to increased diastolic loading conditions. It is likely that the kinetic energy, a nontrivial force in the fetal heart (1), also contributes to the findings of myocyte and RV enlargement. It is important to remember that the energy imparted to the walls of the atria and ventricles is not derived solely from hydrostatic pressure. Rather, it is the sum total of energy in the blood comprising hydrostatic pressure and kinetic energy. Our previous studies showed that where hydrostatic pressures are low and blood velocities are high, as in the normal foramen ovale, kinetic energy is the dominant form of energy that is sensed by atrial and ventricular regions of the myocardium (1). In the case of an AV shunt, blood flow velocities are increased substantially and can be detected by palpation of the resulting thrill in the vein downstream from the shunt. Thus, because the velocities were increased downstream of the anastomosis relative to increases is mean atrial pressure, we surmise that the source of the energy in the blood that stimulated RV cardiomyocyte elongation was derived largely from kinetic energy (1). PA pressure, a surrogate for RV afterload, was the same for the sham and AV fistula groups. The increase in pulmonary flow occurred without a measurable increase in right atrial or PA pressure. Jouannic and colleagues found increased PA pressures in fetuses with AV fistulas (24, 25). Unlike our experiments, their experiments were carried out in acutely exteriorized fetuses under the influence of maternal and fetal halothane anesthesia immediately following fetal thoracic dissection and direct cannulation of the PA (24, 25). We observed at the time of our surgeries that slight manipulation of the ductus arteriosus can cause substantial constriction, which would increase PA pressure in an acute preparation, but not in the chronic in utero preparation, where the ductus arteriosus is fully relaxed, as in the present study.
Cardiac growth in response to the AV fistula.
Overall, cardiac mass was not increased in the AV fistula compared with the sham group. However, RV free wall weight was greater in the AV fistula group (Table 3). The increase in RV free wall weight following chronically elevated RV venous return is consistent with the finding of increased RV weight reported by Jouannic and colleagues (24). Chronic fetal anemia is another condition of increased volume load with increased cardiac output (13, 14). Chronically anemic fetuses have heavier hearts, with both RV and LV chamber enlargement and thicker walls with accelerated maturation of the RV cardiomyocytes (28). Conversely, Fishman and associates, using a left atrial balloon to reduce fetal LV inflow, found that LV inflow obstruction inhibited free wall growth in the LV (15), similar to the hypoplastic left heart syndrome. Together, these observations show that diastolic filling of the heart can either up- or downregulate fetal ventricular growth. Growth factors could also play a role, as our laboratory has shown (17, 46).
In normally growing fetal sheep, the fetal heart grows by both myocyte hyperplasia and hypertrophy (21–23). As birth approaches, hyperplastic growth slows, with the number of mononucleated myocytes decreasing and binucleated myocytes increasing. After birth, hyperplastic growth is at a very low level. Before and after birth, LV myocytes, growing by hypertrophy, grow more rapidly in their short axis (diameter) than long axis (length) (21, 22). In contrast, RV myocytes growing by hypertrophy remain relatively more elongated (21, 22), especially after birth, when RV systolic load decreases with the hemodynamic changes associated with breathing after birth. Compared with normal growth in the sham group, both mononucleated and binucleated LV and RV myocytes were longer in the AV fistula group. RV and LV cardiomyocyte widths were not different between the two groups (Table 4). The myocyte elongation in this study is thus likely a response to increased preload in the absence of increased afterload.
Cardiomyocyte size and cell cycle activity changed in both ventricles, but only RV free wall weight was increased. The signal for the increased LV myocyte size and why LV mass was not increased are not known. It is likely that the greatest portion of the flow from the AV fistula coursed through the superior vena cava, through the RA to the RV and into the PA, with most of this PA flow through the ductus arteriosus into the descending thoracic aorta (1, 37). It is likely that the increase in volume flow contributed by the AV fistula was the stimulus for the hypertrophy in the RV. It is possible that a portion of the AV fistula flow, along with flow from the placenta and fetal lower body, would course through the foramen ovale to the left atrium and then into the LV. This additional flow could have been enough to stimulate elongation of the LV cardiomyocytes without significantly increasing LV free wall mass during the 7-day experimental period, or perhaps myocyte elongation was offset by reduced LV myocyte proliferation. RV and LV myocyte elongation suggests a high level of mechanical sensitivity to diastolic load. We speculate that the increase in circulating ANP could account for the depressed cell cycle activity in both ventricles, as we previously found is a dose-dependent inhibition of angiotensin II-stimulated proliferation by ANP in fetal cardiomyocytes (33). The cellular changes in the hearts of fetuses with fistulas cannot be explained as accelerated maturation. Myocytes of fetuses with fistula did not grow wider, nor did they become more terminally differentiated as a population; both of these outcomes would be expected with accelerated maturation. The presence of a fetal AV fistula was associated with a 50% reduction in cell cycle activity in the LV (P < 0.02; Fig. 3B). Although greater in magnitude of effect, RV cell cycle activity was 66% lower in the AV fistula group (P = 0.054).
The cardiomyocyte growth pattern in the current study contrasts with that observed in fetuses experiencing increased systolic pressure load. When the fetal heart is exposed to increased systolic load, cardiomyocytes proliferate in the first several days of increased systolic load, increasing cell number (2, 21), but proliferative activity is diminished with prolonged arterial load (21, 31). Fetal arterial hypertension stimulates increases in myocyte width and length, as well as increases in terminal differentiation (2, 21, 31). The elongation and widening of fetal cardiomyocytes are the underpinnings of concentric and eccentric hypertrophy described by Grossman and associates (19, 20).
Conclusion.
We hypothesized that chronically increased intrauterine right heart volume load would lead to RV cardiomyocyte proliferation, hypertrophy, and accelerated terminal differentiation. We found that addition of AV fistula flow to existing venous return increased RV SV and output. The fetal heart compensated by cardiomyocyte elongation, a classical response to volume load in the adult heart. In contrast to systolic overload, this study suggests that fetal diastolic load does not drive accelerated cellular maturation and that proliferation is actually decreased. These results indicate that fetuses experiencing even transient volume overload states in utero likely have altered myocardial structure and cardiomyocyte endowment. Based on these studies, we speculate that ANP suppresses fetal cardiomyocyte cell cycle activity in vivo (in contrast to its antihypertrophic effect in mature cardiomyocytes).
Perspectives
There is clear evidence that prenatal conditions impact risk for long-term cardiovascular health. For example, even brief periods of fetal anemia result in detrimental long-term changes in the heart and vasculature in adults. Prior to this study, the role of diastolic load in regulating fetal heart growth was unknown. Our findings show that volume overload stimulates RV hypertrophic growth while suppressing hyperplastic cardiac growth. Based on fetal cardiac adaptation to the volume load, we speculate that fetal volume load will predispose the heart to long-term RV diastolic dysfunction in later life.
GRANTS
This research was supported by National Heart, Lung, and Blood Institute Grant R01-HL-102763 and Eunice Kennedy Shriver National Institute of Child Health and Human Development Grants R01-HD-071068 and P01-HD-034430.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
T.B.K., G.D.G., D.F.M., I.S., and K.L.T. performed experiments; T.B.K., G.D.G., and S.S.J. analyzed data; T.B.K., G.D.G., D.F.M., S.S.J., and K.L.T. interpreted results of experiments; T.B.K. and G.D.G. drafted manuscript; T.B.K., G.D.G., D.F.M., S.S.J., I.S., R.M.U., and K.L.T. edited and revised manuscript; T.B.K., G.D.G., D.F.M., S.S.J., I.S., R.M.U., and K.L.T. approved final version of manuscript; G.D.G. and K.L.T. conceived and designed research; S.S.J. prepared figures.
ACKNOWLEDGMENTS
We thank Robert Webber and Loni Socha for excellent technical assistance.
REFERENCES
- 1.Anderson DF, Faber JJ, Morton MJ, Parks CM, Pinson CW, Thornburg KL. Flow through the foramen ovale of the fetal and new-born lamb. J Physiol 365: 29–40, 1985. doi: 10.1113/jphysiol.1985.sp015757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Barbera A, Giraud GD, Reller MD, Maylie J, Morton MJ, Thornburg KL. Right ventricular systolic pressure load alters myocyte maturation in fetal sheep. Am J Physiol Regul Integr Comp Physiol 279: R1157–R1164, 2000. doi: 10.1152/ajpregu.2000.279.4.R1157. [DOI] [PubMed] [Google Scholar]
- 3.Barker DJ, Osmond C, Golding J, Kuh D, Wadsworth ME. Growth in utero, blood pressure in childhood and adult life, and mortality from cardiovascular disease. BMJ 298: 564–567, 1989. doi: 10.1136/bmj.298.6673.564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Barker DJ, Osmond C, Winter PD, Margetts B, Simmonds SJ. Weight in infancy and death from ischaemic heart disease. Lancet 334: 577–580, 1989. doi: 10.1016/S0140-6736(89)90710-1. [DOI] [PubMed] [Google Scholar]
- 5.Belenkie I, Baumber JS, Rademaker A. Changes in left ventricular dimensions and performance resulting from acute and chronic volume overload in the conscious dog. Can J Physiol Pharmacol 61: 1274–1280, 1983. doi: 10.1139/y83-184. [DOI] [PubMed] [Google Scholar]
- 6.Braunwald E. Heart Disease: A Textbook of Cardiovascular Medicine (3rd ed.). Philadelphia: Saunders, 1988, p. 214. [Google Scholar]
- 7.Broberg CS, Giraud GD, Schultz JM, Thornburg KL, Hohimer AR, Davis LE. Fetal anemia leads to augmented contractile response to hypoxic stress in adulthood. Am J Physiol Regul Integr Comp Physiol 285: R649–R655, 2003. doi: 10.1152/ajpregu.00656.2002. [DOI] [PubMed] [Google Scholar]
- 8.Carabello BA. Concentric versus eccentric remodeling. J Card Fail 8, Suppl: S258–S263, 2002. doi: 10.1054/jcaf.2002.129250. [DOI] [PubMed] [Google Scholar]
- 9.Carabello BA, Nakano K, Corin W, Biederman R, Spann JF Jr. Left ventricular function in experimental volume overload hypertrophy. Am J Physiol Heart Circ Physiol 256: H974–H981, 1989. doi: 10.1152/ajpheart.1989.256.4.H974. [DOI] [PubMed] [Google Scholar]
- 10.Chancey AL, Brower GL, Peterson JT, Janicki JS. Effects of matrix metalloproteinase inhibition on ventricular remodeling due to volume overload. Circulation 105: 1983–1988, 2002. doi: 10.1161/01.CIR.0000014686.73212.DA. [DOI] [PubMed] [Google Scholar]
- 11.Cheung CY, Roberts VJ. Developmental changes in atrial natriuretic factor content and localization of its messenger ribonucleic acid in ovine fetal heart. Am J Obstet Gynecol 169: 1345–1351, 1993. doi: 10.1016/0002-9378(93)90303-Z. [DOI] [PubMed] [Google Scholar]
- 12.Davis L, Roullet JB, Thornburg KL, Shokry M, Hohimer AR, Giraud GD. Augmentation of coronary conductance in adult sheep made anaemic during fetal life. J Physiol 547: 53–59, 2003. doi: 10.1113/jphysiol.2002.023283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Davis LE, Hohimer AR. Hemodynamics and organ blood flow in fetal sheep subjected to chronic anemia. Am J Physiol Regul Integr Comp Physiol 261: R1542–R1548, 1991. doi: 10.1152/ajpregu.1991.261.6.R1542. [DOI] [PubMed] [Google Scholar]
- 14.Davis LE, Hohimer AR, Giraud GD, Reller MD, Morton MJ. Right ventricular function in chronically anemic fetal lambs. Am J Obstet Gynecol 174: 1289–1294, 1996. doi: 10.1016/S0002-9378(96)70673-7. [DOI] [PubMed] [Google Scholar]
- 15.Fishman NH, Hof RB, Rudolph AM, Heymann MA. Models of congenital heart disease in fetal lambs. Circulation 58: 354–364, 1978. doi: 10.1161/01.CIR.58.2.354. [DOI] [PubMed] [Google Scholar]
- 16.Frawley GP, Dargaville PA, Mitchell PJ, Tress BM, Loughnan P. Clinical course and medical management of neonates with severe cardiac failure related to vein of Galen malformation. Arch Dis Child Fetal Neonatal Ed 87: F144–F149, 2002. doi: 10.1136/fn.87.2.F144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Giraud GD, Louey S, Jonker S, Schultz J, Thornburg KL. Cortisol stimulates cell cycle activity in the cardiomyocyte of the sheep fetus. Endocrinology 147: 3643–3649, 2006. doi: 10.1210/en.2006-0061. [DOI] [PubMed] [Google Scholar]
- 18.Giraud GD, Morton MJ, Reid DL, Reller MD, Thornburg KL. Effects of ductus arteriosus occlusion on pulmonary artery pressure during in utero ventilation in fetal sheep. Exp Physiol 80: 129–139, 1995. doi: 10.1113/expphysiol.1995.sp003828. [DOI] [PubMed] [Google Scholar]
- 19.Grossman W, Jones D, McLaurin LP. Wall stress and patterns of hypertrophy in the human left ventricle. J Clin Invest 56: 56–64, 1975. doi: 10.1172/JCI108079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grossman W, Paulus WJ. Myocardial stress and hypertrophy: a complex interface between biophysics and cardiac remodeling. J Clin Invest 123: 3701–3703, 2013. doi: 10.1172/JCI69830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jonker SS, Faber JJ, Anderson DF, Thornburg KL, Louey S, Giraud GD. Sequential growth of fetal sheep cardiac myocytes in response to simultaneous arterial and venous hypertension. Am J Physiol Regul Integr Comp Physiol 292: R913–R919, 2007. doi: 10.1152/ajpregu.00484.2006. [DOI] [PubMed] [Google Scholar]
- 22.Jonker SS, Louey S, Giraud GD, Thornburg KL, Faber JJ. Timing of cardiomyocyte growth, maturation, and attrition in perinatal sheep. FASEB J 29: 4346–4357, 2015. doi: 10.1096/fj.15-272013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jonker SS, Zhang L, Louey S, Giraud GD, Thornburg KL, Faber JJ. Myocyte enlargement, differentiation, and proliferation kinetics in the fetal sheep heart. J Appl Physiol (1985) 102: 1130–1142, 2007. doi: 10.1152/japplphysiol.00937.2006. [DOI] [PubMed] [Google Scholar]
- 24.Jouannic JM, Martinovic J, Roussin R, Laborde F, Dumez Y, Dinh-Xuan AT. The effect of a systemic arteriovenous fistula on the pulmonary arterial blood pressure in the fetal sheep. Prenat Diagn 22: 48–51, 2002. doi: 10.1002/pd.255. [DOI] [PubMed] [Google Scholar]
- 25.Jouannic JM, Roussin R, Hislop AA, Lanone S, Martinovic J, Boczkowski J, Dumez Y, Dinh-Xuan AT. Systemic arteriovenous fistula leads to pulmonary artery remodeling and abnormal vasoreactivity in the fetal lamb. Am J Physiol Lung Cell Mol Physiol 285: L701–L709, 2003. doi: 10.1152/ajplung.00369.2002. [DOI] [PubMed] [Google Scholar]
- 26.Knilans TK. Cardiac abnormalities associated with hydrops fetalis. Semin Perinatol 19: 483–492, 1995. doi: 10.1016/S0146-0005(05)80055-2. [DOI] [PubMed] [Google Scholar]
- 27.Levy D, Garrison RJ, Savage DD, Kannel WB, Castelli WP. Prognostic implications of echocardiographically determined left ventricular mass in the Framingham Heart Study. N Engl J Med 322: 1561–1566, 1990. doi: 10.1056/NEJM199005313222203. [DOI] [PubMed] [Google Scholar]
- 28.Martin C, Yu AY, Jiang BH, Davis L, Kimberly D, Hohimer AR, Semenza GL. Cardiac hypertrophy in chronically anemic fetal sheep: increased vascularization is associated with increased myocardial expression of vascular endothelial growth factor and hypoxia-inducible factor 1. Am J Obstet Gynecol 178: 527–534, 1998. doi: 10.1016/S0002-9378(98)70433-8. [DOI] [PubMed] [Google Scholar]
- 29.Modesti PA, Vanni S, Bertolozzi I, Cecioni I, Polidori G, Paniccia R, Bandinelli B, Perna A, Liguori P, Boddi M, Galanti G, Serneri GG. Early sequence of cardiac adaptations and growth factor formation in pressure- and volume-overload hypertrophy. Am J Physiol Heart Circ Physiol 279: H976–H985, 2000. doi: 10.1152/ajpheart.2000.279.3.H976. [DOI] [PubMed] [Google Scholar]
- 30.Morton MJ, Thornburg KL. The pericardium and cardiac transmural filling pressure in the fetal sheep. J Dev Physiol 9: 159–168, 1987. [PubMed] [Google Scholar]
- 31.Norris AW, Bahr TM, Scholz TD, Peterson ES, Volk KA, Segar JL. Angiotensin II-induced cardiovascular load regulates cardiac remodeling and related gene expression in late-gestation fetal sheep. Pediatr Res 75: 689–696, 2014. doi: 10.1038/pr.2014.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.O’Tierney PF, Anderson DF, Faber JJ, Louey S, Thornburg KL, Giraud GD. Reduced systolic pressure load decreases cell-cycle activity in the fetal sheep heart. Am J Physiol Regul Integr Comp Physiol 299: R573–R578, 2010. doi: 10.1152/ajpregu.00754.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.O’Tierney PF, Chattergoon NN, Louey S, Giraud GD, Thornburg KL. Atrial natriuretic peptide inhibits angiotensin II-stimulated proliferation in fetal cardiomyocytes. J Physiol 588: 2879–2889, 2010. doi: 10.1113/jphysiol.2010.191098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Olson AK, Protheroe KN, Segar JL, Scholz TD. Mitogen-activated protein kinase activation and regulation in the pressure-loaded fetal ovine heart. Am J Physiol Heart Circ Physiol 290: H1587–H1595, 2006. doi: 10.1152/ajpheart.00984.2005. [DOI] [PubMed] [Google Scholar]
- 35.Pinson CW, Morton MJ, Thornburg KL. An anatomic basis for fetal right ventricular dominance and arterial pressure sensitivity. J Dev Physiol 9: 253–269, 1987. [PubMed] [Google Scholar]
- 36.Reller MD, Morton MJ, Thornburg KL. Right ventricular function in the hypoxaemic fetal sheep. J Dev Physiol 8: 159–166, 1986. [PubMed] [Google Scholar]
- 37.Rudolph AM, Heymann MA. The circulation of the fetus in utero. Methods for studying distribution of blood flow, cardiac output and organ blood flow. Circ Res 21: 163–184, 1967. doi: 10.1161/01.RES.21.2.163. [DOI] [PubMed] [Google Scholar]
- 38.Samson F, Bonnet N, Heimburger M, Rücker-Martin C, Levitsky DO, Mazmanian GM, Mercadier JJ, Serraf A. Left ventricular alterations in a model of fetal left ventricular overload. Pediatr Res 48: 43–49, 2000. doi: 10.1203/00006450-200007000-00010. [DOI] [PubMed] [Google Scholar]
- 39.Sandgren J, Scholz TD, Segar JL. ANG II modulation of cardiac growth and remodeling in immature fetal sheep. Am J Physiol Regul Integr Comp Physiol 308: R965–R972, 2015. doi: 10.1152/ajpregu.00034.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Segar JL, Dalshaug GB, Bedell KA, Smith OM, Scholz TD. Angiotensin II in cardiac pressure-overload hypertrophy in fetal sheep. Am J Physiol Regul Integr Comp Physiol 281: R2037–R2047, 2001. doi: 10.1152/ajpregu.2001.281.6.R2037. [DOI] [PubMed] [Google Scholar]
- 41.Segar JL, Scholz TD, Bedell KA, Smith OM, Huss DJ, Guillery EN. Angiotensin AT1 receptor blockade fails to attenuate pressure-overload cardiac hypertrophy in fetal sheep. Am J Physiol Regul Integr Comp Physiol 273: R1501–R1508, 1997. doi: 10.1152/ajpregu.1997.273.4.R1501. [DOI] [PubMed] [Google Scholar]
- 42.Segar JL, Volk KA, Lipman MH, Scholz TD. Thyroid hormone is required for growth adaptation to pressure load in the ovine fetal heart. Exp Physiol 98: 722–733, 2013. doi: 10.1113/expphysiol.2012.069435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Selbing A, Stangenberg M, Westgren M, Rahman F. Intrauterine intravascular transfusions in fetal erythroblastosis: the influence of net transfusion volume on fetal survival. Acta Obstet Gynecol Scand 72: 20–23, 1993. doi: 10.3109/00016349309013343. [DOI] [PubMed] [Google Scholar]
- 44.Silverman NH, Schmidt KG. Ventricular volume overload in the human fetus: observations from fetal echocardiography. J Am Soc Echocardiogr 3: 20–29, 1990. doi: 10.1016/S0894-7317(14)80295-2. [DOI] [PubMed] [Google Scholar]
- 45.Sugden PH. Mechanotransduction in cardiomyocyte hypertrophy. Circulation 103: 1375–1377, 2001. doi: 10.1161/01.CIR.103.10.1375. [DOI] [PubMed] [Google Scholar]
- 46.Sundgren NC, Giraud GD, Schultz JM, Lasarev MR, Stork PJ, Thornburg KL. Extracellular signal-regulated kinase and phosphoinositol-3 kinase mediate IGF-1 induced proliferation of fetal sheep cardiomyocytes. Am J Physiol Regul Integr Comp Physiol 285: R1481–R1489, 2003. doi: 10.1152/ajpregu.00232.2003. [DOI] [PubMed] [Google Scholar]
- 47.Thornburg KL, Morton MJ. Filling and arterial pressures as determinants of left ventricular stroke volume in fetal lambs. Am J Physiol Heart Circ Physiol 251: H961–H968, 1986. doi: 10.1152/ajpheart.1986.251.5.H961. [DOI] [PubMed] [Google Scholar]
- 48.Walther T, Stepan H, Faber R. Dual natriuretic peptide response to volume load in the fetal circulation. Cardiovasc Res 49: 817–819, 2001. doi: 10.1016/S0008-6363(00)00303-5. [DOI] [PubMed] [Google Scholar]
- 49.Yang Q, Hohimer AR, Giraud GD, Van Winkle DM, Underwood MJ, He GW, Davis LE. Effect of fetal anaemia on myocardial ischaemia-reperfusion injury and coronary vasoreactivity in adult sheep. Acta Physiol (Oxf) 194: 325–334, 2008. doi: 10.1111/j.1748-1716.2008.01892.x. [DOI] [PMC free article] [PubMed] [Google Scholar]