

Abstract
Objective
To investigate the influence of knocking down ezrin expression in combination with heat shock protein (HSP)‐induced immune killing on the apoptosis and proliferation of mouse osteosarcoma cells.
Methods
The HSP70 and ezrin‐shRNA DNA fragments cloned into the expression vector pGFP‐V‐RS and the expression vectors pGFP‐V‐RS‐shRNA and pGFP‐V‐RS‐shRNA‐HSP70 constructed and transfected into MG63 cell line, where their status was observed by fluorescent microscopy. Expression of ezrin and HSP70 was determined by RT‐PCR and western blot. Changes in cell apoptosis and proliferation were assessed by flow cytometry and MTS and changes in expression of apoptosis and cell cycle‐related proteins by western blot. Specific cytotoxic T lymphocytes (CTLs) were induced by HSP70 and its lethal effect on target MG63 tumor cells analyzed by MTS assay.
Results
The specific vector simultaneously downregulated ezrin and upregulated HSP70. Compared with ezrin knockdown alone, simultaneous HSP70 overexpression partially recovered the promoted cellular apoptosis and proliferation suppression by induced by ezrin knockdown; however, the apoptosis rate of MG63 cells was significantly greater than that of a negative control. In addition, ezrin‐shRNA and ezrin‐shRNA/HSP70 promoted expression of Bax. However, expression of these agents reduces Bcl‐2 and Cyclin D1. The cytotoxic effects of CTLs on target MG63 tumor cells were significantly greater in the CTL + IL‐2 + HSP70 group than the CTL + IL‐2 group.
Conclusions
Simultaneously knocking down ezrin and overexpressing HSP70 promotes apoptosis and inhibits proliferation of osteosarcoma cells and HSP70 induces CTL, enhancing the lethal effect on tumor cells.
Keywords: Apoptosis, Ezrin, Heat shock protein 70, Osteosarcoma, Proliferation
Introduction
Osteosarcoma is one of the most common primary malignant tumors of bone and young people have higher rates of recurrence and metastasis. The five‐year survival rate is still no more than 50% after neoadjuvant chemotherapy1. At the time of diagnosis with osteosarcoma, 80% of all patients have lung metastases2. Local recurrence occurs within 3 years in 27% of patients who have been treated with chemotherapy after surgery3. Thus, identifying new means of inhibiting proliferation and metastasis is critical for osteosarcoma therapy.
Ezrin, a cytoskeletal protein that belongs to the ezrin/Radixin/Moesin family, binds to membrane proteins or cell surface receptors to regulate local invasion and distant metastasis4. Rho or Akt signaling pathways regulate the phosphorylation of ezrin to inhibit apoptosis and promote cellular proliferation5. Meta‐analysis has shown that ezrin expression is significantly upregulated in osteosarcoma tissues and correlates negatively with five‐year survival rate6. However, the role of ezrin in osteosarcoma has not been studied.
It is currently difficult to completely remove tumor cells in vivo by targeting only one gene; how to achieve selection of complex target genes is becoming a hot research area. Heat shock protein (HSP) not only protects organism from all kinds of stress‐induced injuries, but also binds polypeptide tumor specific antigen to deliver polypeptide messages to the cell membrane to enhance the immunogenicity of this antigen. This induces T lymphocytes to develop into cytotoxic T‐lymphocytes (CTLs) and promotes specific antitumor immunity that is lethal to tumor cells7, 8. HSP70 is one of the strong inductivity proteins of the HSP family. Proteomic analysis has shown HSP70 expression is differs significantly between osteosarcoma cells and osteoblasts9. HSP70 may be an important biomarker for osteosarcoma research. In short, when the proliferation of tumor cells is suppressed and their apoptosis induced, overexpression of HSP may promote a lethal effect on tumor cells and eradicate them10.
Here, we used pGFP‐V‐RS vector with double promoters to construct vectors with HSP70 overexpression and ezrin shRNA, pGFP‐V‐RS‐shRNA and pGFP‐V‐RS‐shRNA‐HSP70. We transfected these vectors into osteosarcoma cell line MG63 to screen for stable cell lines, then used these cell lines to research the role of ezrin and HSP70 in cell proliferation and early and advanced apoptosis. Finally, we determined the activity of CTLs induced by HSP70 on MG63.
Materials and Methods
Cell Lines and Cell Culture
Osteosarcoma cell line MG63 (Cat. No. TCHu124) was purchased from Shanghai cell bank (Chinese Academy of Sciences, Beijing, China). Cells were cultured in RPMI 1640 (Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum (Hyclone) in a 5% CO2 humidified incubator at 37 °C.
Vector Constructs
According to the sequence of ezrin mRNA (GenBank: BC013903.2) in Genbank, two oligos of shRNA containing BamH I and Hind III were synthesized, the forward oligo being 5′‐GATCCTGCTATGTTGGAATACCT‐TTCAAGAGA‐AGGTATTCCAACATAGCATTG‐3′, while the reverse oligo, 5′‐AATTCAATGCTATGTTGGAATACCT‐TCTCTTGAA‐AGGTATTCCAACATAGCAG‐3′. Forward and reverse oligos were annealed and ligated into the pGFP‐V‐RS vector (Origene, Rockville, MD, USA). The ligation mix was transformed into competent JM10 and positive clones verified by sequencing (Invitrogen, Carlsbad, CA, USA).
The sequence of HSP70 was synthesized according to HSP70 mRNA (GenBank: L12723.1). Polymerase chain reaction (PCR) was used to replace GFP with GFP‐HSP70 in the pGFP‐V‐RS vector. The ligation mix was transformed into competent DH5α and positive clones verified by performing a sequencing reaction (Invitrogen). This HSP70 overexpression vector was named pGFP‐V‐RS‐HSP70. The same method was used to construct a new pGFP‐V‐RS‐shRNA‐HSP70 vector that simultaneously overexpresses HSP70 and downregulates ezrin. The flow chart of plasmid construction is shown in Fig. 1.
Figure 1.
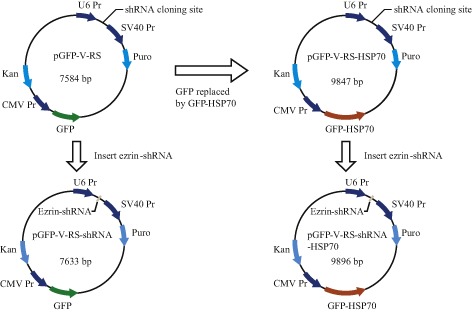
Sketch map of construction of pGFP‐V‐RS‐shRNA‐HSP70 vector.
Empty vector pGFP‐V‐RS was used as a negative control, pGFP‐V‐RS‐shRNA and pGFP‐V‐RS‐shRNA‐HSP70 were transfected into MG63 using Lipofectamine 2000 (Invitrogen). G418 (200 μg/mL) was used to generate a stable cell line.
Real Time PCR Analysis
Total RNAs were extracted using Trizol (Invitrogen) according to the manufacturer's instructions. RNAs were reverse transcribed using M‐MLV reverse transcriptase (Promega, Madison, WI, USA). Primers used for ezrin real time PCR were: ezrin‐F: 5′‐ACTCACCAGAAACCGAAAATG‐3′, ezrin‐R: 5′‐TGGAGGCCAAAGTACCACAC‐3′; the amplified DNA fragments are 159 bp. Primers used for HSP70 real time PCR were: HSP 70‐F: 5′‐AAGAGCAACAGCAGCAGACA‐3′, HSP 70‐R: 5′‐CGATTGGCAGGTCCACAGTA‐3′; the amplified DNA fragments are 150 bp. 18S RNA was used as internal control. Primers used for 18S RNA real time PCR were: 18S RNA‐F: 5′‐CCTGGATACCGCAGCTAGGA ‐3′, 18S RNA‐R: 5′‐GCGGCGCAATACGAATGCCCC‐3′; the amplified DNA fragments are 112 bp. Real‐time PCR was performed using SYBR Green qPCR SuperMix (Invitrogen) according to the manufacturer's instructions. The reaction conditions were 50 °C for 2 min, 95 °C for 2 min, 95 °C for 15 s and 60 °C for 32 s (data collection). Data presented are from three independent experiments.
Western Blot Assay
Total proteins were extracted from cells and protein samples separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes, which were blocked with for 1 h at room temperature. The membranes were then incubated with primary antibodies, followed by horseradish‐peroxidase‐linked goat anti‐Rabbit immunoglobulin G (1:20 000; Southern Biotech, Birmingham, AL, USA). Proteins were visualized using an ECL reagent and exposed to films. Anti‐ezrin, anti‐HSP70, anti‐Bcl‐2, anti‐Bax and anti‐Cyclin D1 antibody (CST, 1:1000) were used as primary antibodies and anti‐glyceraldehyde phosphate dehydrogenase antibody (Kangchen, Shanghai, China) was used as a loading control.
Apoptosis Assay
Apoptosis assays were performed using Annexin V‐PE/7‐aminoactinomycin D Apoptosis Detection Kit (Keygen, Nanjin, Jiangsu, China) according to the manufacturer's instructions. Briefly, 0.5 × 106 cells were resuspended in cold Binding Buffer, then incubated with 1.25 μL Annexin V‐PE for 15 min at room temperature, followed by incubation with 10 μL 7‐aminoactinomycin D. The samples were kept on ice and immediately subjected to flow cytometry. Data presented are averages from three independent experiments.
MTS Assay
Cell proliferation was assessed using MTS assay. Briefly, 0.1 × 104 to 0.2 × 104 cells were cultured in 96‐well plates and stained at 24, 72, 100 and 168 h with 20 μL MTS/PMS buffer for 3–4 h. The absorbance was determined at 570 nm. All experiments were performed in triplicate. Averages of absorbance are shown. Cell growth curves were drawn and the proliferation and inhibition rates calculated.
Human Spleen Lymphocyte Preparation
Human splenic tissue obtained by splenectomy for traumatically ruptured spleen was extracted under sterile conditions. The splenic tissue was cut up with sterile scissors, filtered used 100 mesh stainless steel mesh and resuspended in Hank's buffer (pH 7.2–7.6). Lymphocyte separation buffer (specific gravity 1.083) was added to cell suspensions to separate mononuclear cells (lymphocytes and monocytes) and RPMI‐1640 supplemented with 15% newly born bovine serum used to adjust the concentration of mononuclear cell to 1 × 106/mL.
Using a culture supernatant of MG63 which had been transfected with pGFP‐V‐RS‐shRNA‐HSP70 vector, 1 × 106 lymphocytes were resuspended, recombinant interleukin‐2 (rIL‐2, 2000 U/mL) added to the culture and the cells cultured in a 5% CO2 humidified incubator at 37 °C. Cells extracted after 7 days; these cells were sensitized CTLs.
Using a culture supernatant of MG63 which had been transfected with pGFP‐V‐RS‐shRNA vector, 1 × 106 lymphocytes were resuspended and rIL‐2 (2000 U/mL) added to the culture and the cells cultured in a 5% CO2 humidified incubator at 37 °C. The cells were extracted after 7 days; these cells were non‐sensitized CTLs used as negative control cells. Blank control cells were obtained by culturing 1 × 106 lymphocytes with only rIL‐2 (2000 U/mL).
Activity of CTLs Assay
The concentration of target cells (T), MG63, which were in logarithmic growth phase, was adjusted to 2 × 106/mL. Sensitized and non‐sensitized CTLs were used as effector cells (E). The number of E and T were mixed in the ratio E : T = 100:1, 50:1 and 25:1. One hundred microliter cell mixtures were added to 96‐well plates. For controls, 100 μL/well effector lymphocytes or target cells were seeded; the blank control contained medium without cells. There are 4 repeats for all samples. The cells were cultured in 5% CO2 humidified incubator at 37 °C for 20 h. MTS assay was performed to determine the absorbance. CTL activity = (1 − [ODvalue of experimental − ODvalue of CTLs control/ODvalue of target cells control]) × 100%.
Statistical Analysis
Statistical analyses were performed using SPSS 17.0. Data are presented as mean ± standard deviation (SD). ANOVA was used to evaluate the statistical significance of differences between multiple data. Student's t‐test was used for normally distributed data. Results were considered statistically significant at P < 0.05.
Results
Effects of Cell Transfection
MG63 cells were transfected into ezrin‐shRNA and ezrin‐shRNA/HSP70 vectors; the efficiency of transfection was 70%. They were screened for 1 month by adding G418, after which cells with good growth were chosen for the subsequent experiments. Cells transfected with ezrin‐shRNA and ezrin‐shRNA/HSP70 glowed green under fluorescence microscopy (Fig. 2).
Figure 2.
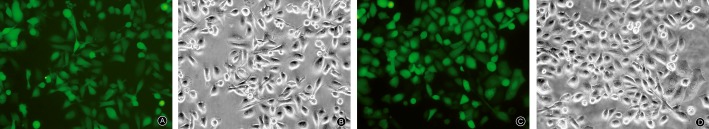
Light and fluorescence microscopy images of MG63 transfected with ezrin‐shRNA and ezrin‐shRNA/HSP70. Magnification ×100 for all images. (A) Fluorescence microscopy image of MG63 transfected with ezrin‐shRNA. (B) Light microscopy images of MG63 transfected with ezrin‐shRNA. (C) Fluorescence microscopy image of MG63 transfected with ezrin‐shRNA/HSP70. (D) Light microscopy image of MG63 transfected with ezrin‐shRNA/HSP70.
Effects of Transfected Plasmids
Real‐time PCR and western blot were used to determine expression of Ezrin and HSP70 in MG63 after transfection with plasmids. According to real‐time PCR assay, ezrin was downregulated when cells were transfected with ezrin‐shRNA and ezrin‐shRNA/HSP70 plasmids (t values were 235.30 and 109.23; P < 0.01 compared with negative control). Thus, the efficiency of RNA interference was greater than 95%. According to western blot assay, ezrin shRNA inhibited ezrin expression (Fig. 3). According to real‐time PCR assay, HSP70 expression was upregulated in cells transfected with ezrin‐shRNA/HSP70, compared with that of a transfected negative control vector or ezrin‐shRNA vector (t values were 10.85, 9.69; P < 0.01). Western blot assay also confirmed this result (Fig. 3).
Figure 3.
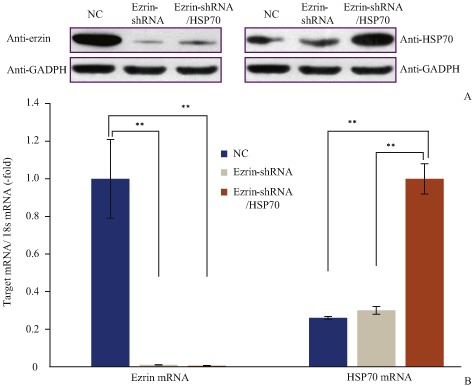
(A) Real‐time PCR and (B) western blot determination of ezrin and HSP70 expression. Ezrin expression was downregulated in cells transfected with ezrin‐shRNA or ezrin‐shRNA/HSP70 vector compared with negative control. HSP70 expression was upregulated in cells transfected with ezrin‐shRNA/HSP70 compared with cells transfected with ezrin‐shRNA or NC. NC, normal control; **, P < 0.01.
Effects of Transfected Plasmids on Cell Apoptosis
Next, whether cell apoptosis occurs when ezrin is downregulated and HSP70 simultaneously upregulated in MG63 cell was determined. Compared with the negative control, downregulation of ezrin caused apoptosis. The number of MG63 decreased from 85.71% ± 0.19% to 70.17% ± 0.58% (t = 44.15, P < 0.05). The number of apoptotic cell increased from 11.01% ± 0.22% to 27.28% ± 0.50% (t = 51.82, P < 0.05), especially the number of late apoptotic cells. Next, the effect of HSP70 overexpression on apoptosis was determined and it was found that the number of apoptotic cells decreased slightly compared with cells that had been downregulated by ezrin alone. However, compared with the negative control, the number of apoptotic cell increased from 11.01% ± 0.22% to 24.28% ± 0.50% (t = 30.06, P < 0.01). Thus, cell apoptosis was still induced when ezrin was downregulated and HSP70 simultaneously upregulated in MG63 cells (Fig. 4).
Figure 4.
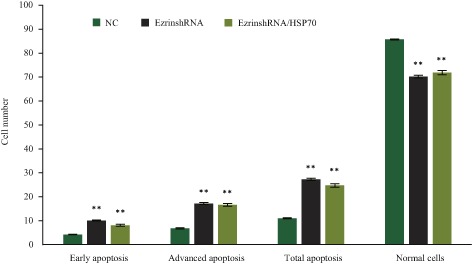
Apoptosis as assessed by flow cytometry. The number of MG63 transfected with ezrin‐shRNA was significantly fewer than for NC. The number of apoptotic cells was significantly greater, especially the number in late apoptosis. Apoptosis of MG63 transfected with Ezrin‐shRNA was also greater than for NC. NC, normal control; **, P < 0.01.
Effects of Transfected Plasmids on Cell Proliferation
MTS was used to determine cell proliferation; cell proliferation (Fig. 5) and inhibition rates (Fig. 6) were calculated according to their absorbance. The absorbance on Day 0 was set as 100%. Compared with the negative control, on Day 3 the proliferation rate of MG63 with knockdown of ezrin was significantly downregulated, being reduced from 395.14% ± 2.24% to 256.50% ± 3.23% (t = 39.50, P < 0.01). The cell inhibition rate reached 35.09% ± 1.05% on Day 7. Proliferation of MG63 with knockdown of ezrin and HSP70 overexpression was also determined. Compared with MG63 with knockdown of ezrin alone, the cell proliferation rate was restored; however, compared with the negative control, the cell proliferation rate decreased from 395.14% ± 2.24% to 310.0% ± 2.83% (t = 25.86, P < 0.01). The cell inhibition rate reached 21.50% ± 1.74%. According to western blot assay, proapoptotic Bax upregulated and antiapoptotic Bcl‐2 and Cyclin D1 downregulated when only ezrin was downregulated. Compared to knockdown of ezrin alone, the changes in upregulation of Bax and downregulation of Bcl‐2 and Cyclin D1 were still evident in MG63 transfected ezrin‐shRNA/HSP70. The changes were significant compared with the negative control (Fig. 7). These data suggest that ezrin‐shRNA/HSP70 inhibits cell proliferation.
Figure 5.
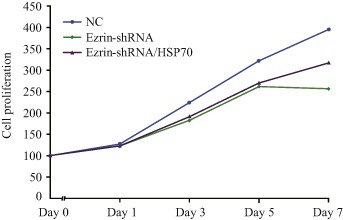
MTS analysis cell proliferation. Compared to negative control, the proliferation of MG63 transfected with ezrin‐shRNA or ezrin‐shRNA/HSP70 significantly reduced. The proliferation of MG63 transfected with ezrin‐shRNA/HSP70 slightly increased on Days 5–7 but was still significantly less than for the NC. NC, normal control.
Figure 6.
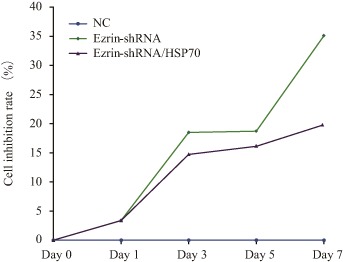
MTS analysis of cell inhibition rate. The inhibition rate of MG63 transfected with ezrin‐shRNA and ezrin‐shRNA/HSP70 significantly increased with prolongation of the culture time and the inhibition rate of MG63 transfected with ezrin‐shRNA/HSP70 was less than that of MG63 transfected with ezrin‐shRNA. However, they still inhibited cellular proliferation compared with NC. NC, normal control.
Figure 7.
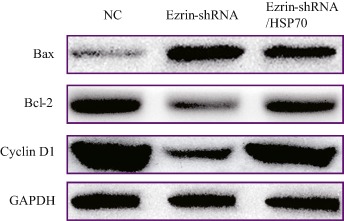
Western blot determined expression of cell cycle and apoptosis association proteins. Knockdown of ezrin promoted proapoptotic Bax expression, and inhibited Bcl‐2 and cyclin D1 expression. Simultaneous knockdown of ezrin and overexpression of HSP70 in MG63 inhibited proapoptotic Bax expression and promoted Bcl‐2 and Cyclin D1 expression, compared with MG63 downregulated by ezrin alone. However, it still differed significantly from that of NC. NC, normal control.
Effects of HSP70 on CTL Activity
MTS assays were used to determine CTL activity of MG63. It was found the CTL activity of MG63 was up to 56.33% ± 1.95%. The effects of CTL activity were significantly different among MG63 with ezrin knockdown and HSP70 overexpression, MG63 with ezrin knockdown alone and MG63 cells in different E : T ratios (t values were 23.80, 21.77, 11.53, 11.89, 12.08 and 13.18, respectively; P < 0.01). The CTL activity of MG63 with ezrin knockdown and HSP70 overexpression was the strongest when E : T was 100:1 (Fig. 8).
Figure 8.
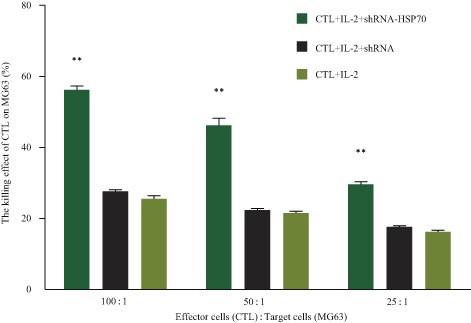
HSP70 promotes CTL activity. CTL activity significantly increased when MG63 was transected with ezrin‐shRNA/HSP70 compared with MG63 transfected with ezrin‐shRNA or NC. (**, P < 0.01 for MG63 transected with ezrin‐shRNA/HSP70 and NC). NC, normal control.
Discussion
A stable cell line transfected with pGFP‐V‐RS‐shRNA‐HSP70 was constructed successfully. Both knockdown of ezrin using siRNA or shRNA and dephosphorylation of ezrin inhibit the function of ezrin11; however, regulation of ezrin activity using siRNA‐mediated RNA inference or inhibitor‐mediated dephosphorylation is transient. Zhang and his colleagues used pGenesil‐1 vector to silence ezrin expression in MG63; however, the efficiency of RNA inference was only 60%12. Ghafouri‐Fard et al. used pGFP‐V‐RS vector to silence GHOXF1 expression in two breast cancer cell lines; the efficiency of RNA inference was up to 80%13. In addition, Zhou et al. found pGFP‐V‐RS vector significantly downregulates Sox4 expression in adenocarcinoma14.
In the present study, we used pGFP‐V‐RS vector to silence ezrin expression. Both real‐time PCR and western blot confirmed ezrin was downregulated in MG63, the efficiency being up to 95%. The amount of ezrin mRNA was downregulated after 36 h, but its protein expression was downregulated after 48 h, this difference may be attributable to the time sequence from gene transcription to protein translation. pGFP‐V‐RS vector contains CMV and pU6 promoter: after cloning ezrin shRNA into a vector, we replaced GFP with HSP70‐GFP to construct a new vector that not only inhibited ezrin, but also simultaneously overexpressed HSP70. We observed green fluorescence 1 month after transfecting MG63with the new vector. We also found ezrin was downregulated and HSP70 upregulated by using real‐time PCR and western blot. These findings suggest the vector and stable cell line were constructed successfully.
Some signaling pathways regulate HSP70 and ezrin to affect cell apoptosis and proliferation. We used flow cytometry to determine early and advanced apoptosis and found their rates in MG63 were 4.3% and 6.8%, respectively15. This is consistent with previous reports. Knockdown of ezrin induces early, advanced and total apoptosis, antiapoptotic Bcl‐2 is downregulated and proapoptotic Bcl homologous protein Bax significantly upregulated. Interaction between Bcl‐2 and Bax can form a specific homodimer that is an important molecular switch for apoptosis signaling. When expression of Bcl‐2 is greater than that of Bax in the homodimer, apoptosis is suppressed, whereas when Bax expression is greater than Bcl‐2 in the homodimer, apoptosis is activated16. This study supports this phenotype. With coordinate HSP70 overexpression, early and total apoptosis rates are slightly less than with knockdown of ezrin alone, which upregulates Bax expression. The ratio of Bcl‐2/Bax in the homodimer also changes. These findings suggest that HSP70 coordinates ezrin to regulate cell apoptosis, especially early apoptosis. However, the signaling pathways that regulate Bcl‐2/Bax are very complicated. Lévy et al. found that ezrin activates PI3K‐Akt and NF‐κB pathways to induce colonic carcinoma apoptosis mediated by Bcl‐2 and Bax17. Wick et al. found that ezrin interacts with CD44 and HSP70 in gliomas. In the absence of transforming growth factor‐β2, Bcl‐2 not only induces apoptosis, but also activates matrix metalloproteinases (MMPs) to promote migration18.
Generally, cell apoptosis often accompanies inhibition of cell proliferation. We found cell proliferation reached its peak on Day 7 after knockdown by ezrin alone. However, we did not determine the expression of MMPs.
p38, MAPK or ROCK signaling pathways regulate expression or phosphorylation of ezrin to suppress MMPs expression19, and inhibit adenocarcinoma proliferation and metastasis20. Heiska et al. found mutation of Tyr477 in ezrin results in ezrin losing its phosphorylation and inhibits Src‐mTOR signaling; cyclin D1 is also inhibited. Cell arrests in G0/G121 in our study also demonstrate that cyclin D1 may participate in the mechanism by which ezrin regulates cell proliferation.
In this study, we did not exhaustively investigate the mechanism of synergy between HSP70 and ezrin on cell proliferation and apoptosis. We plan to use inhibitors of the signaling pathway to further identify the co‐targets of HSP70 and ezrin and the molecular basis for clinical targets of gene therapy.
HSP70 overexpression promotes CTL activity. The cell inhibition rate was 35.1% when ezrin was downregulated alone in MG63. This suggests that knockdown of only one gene cannot kill tumor cells completely in vivo: combined tumor biotherapy may become a new approach to therapy. CTLs are the most important immune surveillance cells and may be ideal effect cells in adoptive immunotherapy. Shen et al. found that tumor cell lysate extracted from mouse colorectal cancer contains HSP70 polypeptide complex; HSP70 can activate specific CTLs to kill tumor cells8. In our study, we found that cocultured overexpression of HSP70 in MG63 and human spleen lymphocytes in vitro in the right proper proportion activates CTLs: when E : T is 100:1, CTLs kill tumor cells with good specificity.
Cytotoxic T‐lymphocytes, which are activated by HSP70 antigen peptide, kill target cells through the following two pathways: (i) HSP70 induces CTLs to secrete IL‐6 and IL‐17, which makes them more lethal22; and (ii) CTLs selectively bind cancer cells with Toll‐like receptor 2/4, cluster of differentiation (CD)40 and CD144 to induce inflammatory signaling, and kill tumor cells23. The question remains: how does HSP overexpression enhance CTL activity on MG3? The answer may be found by analyzing the components of culture supernatants of CTL activity assays or by inhibiting the specificity of CTLs.
In summary, we constructed pCFP‐V‐RS‐shRNA‐HSP70 vector, which when transfected into NG63, not only induced Bcl‐2/Bax‐mediated cell apoptosis and reduced cell proliferation, but also activated CTL activity by combining HSP70. However, we did not ascertain how HSP promotes CTL activity. We only used a cell line to determine cell proliferation, apoptosis and CTL activity; we did not perform animal experiments. Some cytokines do not participate in the lethal effect of CTL in vitro and this may reduce the tumor suppression effect. We therefore plan to study the mechanism of HSP on CTL activity, construct an animal model with ezrin‐shRNA‐HSP70 and evaluate the effects of therapy in vivo. We hope to establish a theoretical and experimental basis for use of gene therapy in the clinic.
Disclosure: This work was supported by the Natural Science Foundation of Fujian Province Youth Innovation Project (2010D017).
References
- 1. Shi X, Xu M, Liu YX, et al Preliminary report of a randomized controlled multicenter study of first‐line chemotherapy regimen combined with angiogenesis inhibitor for osteosarcoma of the extremities. Zhonghua Gu Ke Za Zhi, 2012, 32: 1027–1031 (in Chinese). [Google Scholar]
- 2. Choi LE, Healey JH, Kuk D, Brennan MF. Analysis of outcomes in extraskeletal osteosarcoma: a review of fifty‐three cases. J Bone Joint Surg Am, 2014, 96: e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Lin N, Ye ZM, Li P, Li WX, Tao HM, Yang DS, et al The diuleal features and prognostic factors of osteosarcoma in adult. Zhonghua Gu Ke Za Zhi, 2009, 29: 717–722 (in Chinese). [Google Scholar]
- 4. Donatello S, Babina IS, Hazelwood LD, Hill AD, Nabi IR, Hopkins AM. Lipid raft association restricts CD44‐ezrin interaction and promotion of breast cancer cell migration. Am J Pathol, 2012, 181: 2172–2187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Agarwal E, Chaudhuri A, Leiphrakpam PD, Haferbier KL, Brattain MG, Chowdhury S. Akt inhibitor MK‐2206 promotes anti‐tumor activity and cell death by modulation of AIF and Ezrin in colorectal cancer. BMC Cancer, 2014, 14: 145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Lun DX, Hu YC, Xu ZW, Xu LN, Wang BW. The prognostic value of elevated ezrin in patients with osteosarcoma. Tumour Biol, 2014, 35: 1263–1266. [DOI] [PubMed] [Google Scholar]
- 7. Calderwood SK, Stevenson MA, Murshid A. Heat shock proteins, autoimmunity, and cancer treatment. Autoimmune Dis, 2012, 2012: 486069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Shen YM, Shan BE, Liu LH, Zhang C, Zhou KX, Jiao YQ, Jiao WJ. The anti‐tumor effects of HSP70 peptide complex. Carcinog Teratog Mutag, 2013, 25: 262–266 (in Chinese). [Google Scholar]
- 9. Liu X, Zeng B, Ma J, Wan C. Comparative proteomic analysis of osteosarcoma cell and human primary cultured osteoblastic cell. Cancer Invest, 2009, 27: 345–352. [DOI] [PubMed] [Google Scholar]
- 10. Wang J, Zhang YZ, Pan LL, et al Effect of rhHSP70 on anti‐tumor activity of CTL induced by human cord blood dendritic cells transfect with RNA of esophageal carcinoma cells. Chin J Diffic and Compl Cas, 2011, 10: 45–47 (in Chinese). [Google Scholar]
- 11. Ren L, Hong SH, Chen QR, et al Dysregulation of ezrin phosphorylation prevents metastasis and alters cellular metabolism in osteosarcoma. Cancer Res, 2012, 72: 1001–1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Zhang J, Zuo J, Lei M, Wu S, Zang X, Zhang C. Ezrin promotes invasion and migration of the MG63 osteosarcoma cell. Chin Med J (Engl), 2014, 127: 1954–1959. [PubMed] [Google Scholar]
- 13. Ghafouri‐Fard S, Abdollahi DZ, Omrani M, Azizi F. shRNA mediated RHOXF1 silencing influences expression of BCL2 but not CASP8 in MCF‐7 and MDA‐MB‐231 cell lines. Asian Pac J Cancer Prev, 2012, 13: 5865–5869. [DOI] [PubMed] [Google Scholar]
- 14. Zhou Y, Wang X, Chen Y, Huang Y, Liu X, Jin C, Chen X, et al Target Sox4 gene‐knock‐down suppresses xenograft growth of Xuanwei female lung cancer cell line XWLC‐05 cells in nude mice. Chin J Oncol, 2014, 36: 11–16 (in Chinese). [PubMed] [Google Scholar]
- 15. Zhang H, Wang K, Han Y, Yang J, Wang Y. Effects of polyamine analogues tetrabutyl propanediamine on proliferation, apoptosis and migration of human MG63 myeloma cells. Zhongguo Yao Li Tong Bao, 2012, 28: 974–977 (in Chinese). [Google Scholar]
- 16. Lv XG, Ji MY, Dong WG, et al EBP50 gene transfection promotes 5‐fluorouracil‐induced apoptosis in gastric cancer cells through Bax‐ and Bcl‐2‐triggered mitochondrial pathways. Mol Med Rep, 2012, 5: 1220–1226. [DOI] [PubMed] [Google Scholar]
- 17. Lévy P, Robin H, Bertrand F, Kornprobst M, Capeau J. Butyrate‐treated colonic Caco‐2 cells exhibit defective integrin‐mediated signaling together with increased apoptosis and differentiation. J Cell Physiol, 2003, 197: 336–347. [DOI] [PubMed] [Google Scholar]
- 18. Wick W, Grimmel C, Wild‐Bode C, Platten M, Arpin M, Weller M. Ezrin‐dependent promotion of glioma cell clonogenicity, motility, and invasion mediated by BCL‐2 and transforming growth factor‐beta2. J Neurosci, 2001, 21: 3360–3368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Luo XH, Liao EY, Zhou HD. Differentiation and gene expression of human osteosarcoma cell line MG‐63. Journal of Central South University (Medical Science), 2001, 26: 107–110 (in Chinese). [PubMed] [Google Scholar]
- 20. Liang H, Gu M, Yang C, Wang H, Wen X, Zhou Q. Sevoflurane inhibits invasion and migration of lung cancer cells by inactivating the p38 MAPK signaling pathway. J Anesth, 2012, 26: 381–392. [DOI] [PubMed] [Google Scholar]
- 21. Heiska L, Melikova M, Zhao F, Saotome I, McClatchey AI, Carpén O. Ezrin is key regulator of Src‐induced malignant phenotype in three‐dimensional environment. Oncogene, 2011, 30: 4953–4962. [DOI] [PubMed] [Google Scholar]
- 22. Kottke T, Sanchez‐Perez L, Diaz RM, et al Induction of hsp70‐mediated Th17 autoimmunity can be exploited as immunotherapy for metastatic prostate cancer. Cancer Res, 2007, 67: 11970–11979. [DOI] [PubMed] [Google Scholar]
- 23. Calderwood SK, Murshid A, Gong J. Heat shock proteins: conditional mediators of inflammation in tumor immunity. Front Immunol, 2012, 3: 75. [DOI] [PMC free article] [PubMed] [Google Scholar]