

Abstract
Selecting particles from digital micrographs is an essential step in single-particle electron cryomicroscopy (cryo-EM). As manual selection of complete datasets—typically comprising thousands of particles—is a tedious and time-consuming process, numerous automatic particle pickers have been developed. However, non-ideal datasets pose a challenge to particle picking. Here we present the particle picking software crYOLO which is based on the deep-learning object detection system You Only Look Once (YOLO). After training the network with 200–2500 particles per dataset it automatically recognizes particles with high recall and precision while reaching a speed of up to five micrographs per second. Further, we present a general crYOLO network able to pick from previously unseen datasets, allowing for completely automated on-the-fly cryo-EM data preprocessing during data acquisition. crYOLO is available as a standalone program under http://sphire.mpg.de/ and is distributed as part of the image processing workflow in SPHIRE.
Subject terms: Cryoelectron microscopy, Data processing
Thorsten Wagner et al. present SPHIRE-crYOLO, a particle picking software for selecting particles from digital micrographs in cryoEM data. After training, the method automatically recognizes particles with high recall and precision, simplifying data pre-processing.
Introduction
In recent years, single-particle electron cryomicroscopy (cryo-EM) has become one of the most important and versatile methods for investigating the structure and function of biological macromolecules. In single-particle cryo-EM, many images of identical but randomly oriented macromolecular particles are selected from raw cryo-EM micrographs, digitally aligned, classified, averaged, and back-projected to obtain a three-dimensional (3-D) structural model of the macromolecule.
As more than 100,000 particles often have to be selected for a near-atomic cryo-EM structure, numerous automatic particle-picking procedures, often based on heuristic approaches, have been recently developed1–7. In a popular selection approach, called template matching, cross-correlation of the micrographs is performed against pre-calculated templates of the particle of interest. However, this procedure is error-prone and the parameter optimization is often complicated. Although it performs well with well-behaved data, it often fails when dealing with non-ideal datasets where particles overlap, are compositionally and conformationally heterogeneous, or the background of the micrographs is contaminated with crystalline ice. In such cases, this basic algorithm is unable to detect particles with high enough confidence and, consequently, either particles are missed or many false positives are selected, which need to be removed again afterwards. Often, the last resort is manual selection of particles, which is laborious and time intensive.
To solve this problem, two particle selection programs, DeepEM by Wang et al.8 and DeepPicker by Zhu et al.9, have been recently published, which employ deep convolutional neural networks (CNNs). Besides these programs, Warp10 and Topaz11 were recently described in preprints. CNNs are extremely successful in processing data with a grid-like topology12. For images this is undoubtedly the state-of-the-art method for pattern recognition and object detection. Similar to learning the weights and biases of specific neurons in a multi-layer perceptron13, a CNN learns the elements of convolutional kernels. A convolution is an operation which calculates the weighted sum of the local neighborhood at a specific position in an image. The weights are the elements of the kernel which extracts specific local features of an image, such as corners or edges. In CNNs, several layers of convolutions are stacked and the output of one layer is the input of the next layer. This enables CNNs to learn hierarchies of features, thereby allowing the learning of complex patterns.
Similar to common modern object detection systems, particle selection tools employ a specific classifier for an object and evaluate it at every position. Thus, they are trained with positive examples of cropped out particles and negative examples of cropped out regions of background or contamination. After the classifier is trained, these systems slide a window over the micrograph to crop out single local regions, pass them through a CNN, and classify the extracted regions. The confidence of classification is transferred into a map and the object positions are estimated by finding the local maxima in this map. Using this approach it is possible to select particles on more challenging datasets. However, as it classifies many overlapping cut-out regions independently, this approach comes with a high computational cost. Moreover, as the classifier only sees the windowed region it is not able to learn the larger context of a particle (e.g., to not pick regions near ice contamination).
In 2016, Redmon et al.14 introduced the “You Only Look Once” (YOLO) framework as an alternative to the sliding-window approach and reformulated the classification problem into a regression task where the exact position of a particle is predicted by the network. In contrast to the sliding-window approach the YOLO framework requires only a single pass of the full image instead of multiple passes of cropped out regions. Thus the YOLO approach divides the image into a grid and predicts for each cell of the grid whether it contains the center of a bounding box enclosing an object of interest. If this is the case it applies regression to predict the relative position of the object center inside the cell, as well as the width and height of the bounding box (Fig. 1a). This simplifies the detection pipeline and reduces the number of required convolutions, which increases the speed of the network while retaining its accuracy. During training the YOLO approach only requires labeling-positive examples, whereas the sliding-window approach also requires labeling background and contaminants as negative examples. Moreover, as the crYOLO network sees the complete image it is able to learn the larger context around a particle and therefore provides an excellent framework to reliably detect single particles in electron micrographs (Fig. 1b).
Fig. 1.

Training and picking in crYOLO. a With the YOLO approach the complete micrograph is taken as the input for the CNN. When the image is passed through the network the image is spatially downsampled to a small grid. Then YOLO predicts for each grid cell if it contains the center of a particle bounding box. If this is the case, it estimates the relative position of the particle center inside the cell, as well as the width and height of the bounding box. During training, the network only needs labeled particles. Furthermore, as the network sees the complete micrograph, it learns the context of the particle. b During picking crYOLO processes up to five micrographs per second and thus outperforms the sliding-window approach
Here we present the single particle-selection procedure crYOLO, which utilizes the YOLO approach to select single particles in cryo-EM micrographs. We evaluate our procedure on simulated data, a common benchmark dataset, and three recently published high-resolution cryo-EM datasets. The results demonstrate that crYOLO is able to accurately and precisely select single particles in micrographs of varying quality with a speed of up to five micrographs per second on a single graphics processing unit (GPU). crYOLO leads to a tremendous acceleration of particle selection, simplifying the training as no negative examples have to be labeled, and improving not only the quality of the extracted particle images but also the final structure. Furthermore, we present a general model for crYOLO trained on more than 40 datasets and able to select particles of previously unseen macromolecular species, realizing automatic particle picking at expert level.
Results
Convolutional neural network
The program crYOLO builds upon a Python-based open-source implementation15 of YOLO and uses the deep-learning library Keras16. Beyond the basic implementation, we added patch-processing, multi-GPU support, parallel processing, preprocessing procedures, support for MRC micrographs, single channel data, RELION star files, and EMAN117 box files. CrYOLO includes a graphical tool to read and create box files for training data generation or visualization of the results in a user-friendly manner (Fig. 2). For the readers interested in the details of the YOLO architecture, please refer the online methods and the original paper18. Briefly, the YOLO network consists of 22 convolutional and 5 max-pooling layers. In addition, it contains a passthrough layer between layer 13 and 21 (similar to ResNets19) to exploit fine grain features and the network is followed by a 1 × 1 convolutional layer used as a detection layer. This architecture shows a similar performance to a 50-layer ResNet but with a much lower number of weights18. However, one limitation of the original YOLO network for particle picking is that it uses a relatively coarse grid for prediction. Under special circumstances, e.g., when particles are very small, this might lead to a lower performance due to the fact that each grid cell can only detect a single particle.
Fig. 2.
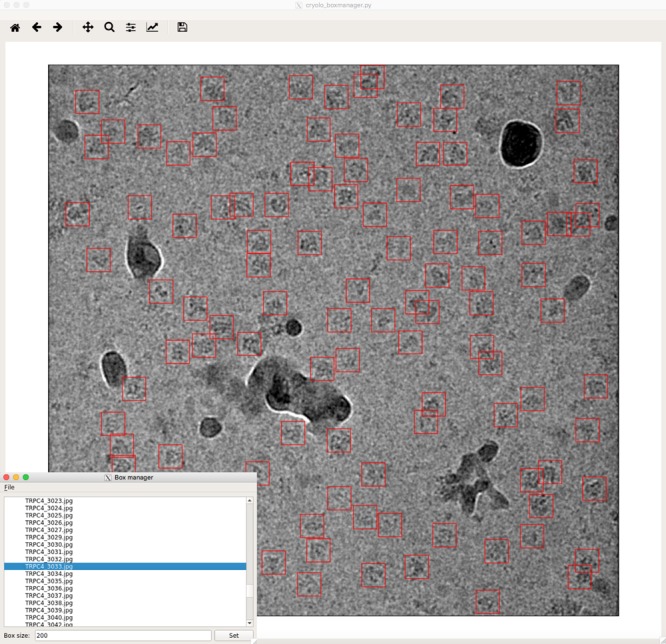
Graphical tool for creating training data and visualizing results. The tool can read images in MRC, TIFF, and JPG format and box files in EMAN1 and STAR format. The example shown is a micrograph of TRPC423 with many contaminants
In order to tackle the problem of detecting very small particles crYOLO divides the input image into a small number of overlapping patches (e.g., 2 × 2 or 3 × 3). Instead of the complete micrograph, each patch is then downscaled to the network input image size of 1024 × 1024. During prediction, all patches are classified in a single batch.
To reduce overfitting and improve the generalization capabilities of the trained network, each image is internally augmented before being passed through the network during training. During augmentation a random selection of the following methods are used: flipping, blurring, adding noise, and random contrast changes, which are described in the Methods.
Test datasets
We used crYOLO to select three different cryo-EM datasets and analyzed the results. These include the TcdA1 toxin subunit from Photorhabdus luminescens20 (EMPIAR-10089, EMDB-3645), the Drosophila transient receptor channel NOMPC21 (EMPIAR-10093, EMDB-8702), and the human peroxiredoxin-3 (Prx3)22 (EMPIAR-10050, EMDB-3233). All datasets were previously shown to produce high-resolution structures. To validate the performance of our program, we also tested it on simulated data of the canonical TRPC4 ion channel23 (EMD-4339) and on a published dataset of the keyhole limpet hemocyanin (KLH)24.
TcdA1 assembles in the soluble prepore state in a large bell-like-shaped pentamer with a molecular weight of 1.4 MDa20. TcdA1 has been our test specimen to develop the software package SPHIRE25. We previously used a dataset of 7475 particles of TcdA1 to obtain a reconstruction at a resolution of 3.5 Å25. Due to the large molecular weight of the specimen and its characteristic shape, particles are clearly discernible in the micrographs, even though a carbon support film was used. Therefore, the selection of the particles is straightforward in this case. We chose this particular test dataset because of its small size, as the quality and number of selected particles would likely have an influence on the quality of the final reconstruction.
In the case of NOMPC, which was reconstituted in nanodiscs21, the overall shape of the particle, sample concentration, and limited contrast make it difficult to accurately select the particles, despite having a molecular weight of 754 kDa. Furthermore, the density of the nanodisc is more pronounced than the density of the extramembrane domain of the protein; thus, the center of mass is not located at the center of particle but shifted toward the nanodisc density. We chose this dataset, as it is a challenge for the selection program to accurately detect the center of the particles and to avoid selection of empty nanodiscs.
Prx3 has a molecular weight of 257 kDa and has a characteristic ring-like shape. The dataset is one of the first near-atomic resolution datasets recorded using a Volta phase plate22 (VPP). The VPP introduces an additional phase shift in the unscattered beam. This increases the phase contrast, providing a boost of the signal-to-noise ratio in the low-frequency range. This makes the structural analysis of low-molecular-weight complexes at high resolution possible26. The VPP, however, enhances not only the contrast of the particles of interest but also the contrast of all weak phase objects, including smaller particles (impurities, contamination, and dissociated particle components) that would otherwise not be easily visible in conventional cryo-EM. This poses new challenges with regard to particle selection, especially for automated particle picking procedures that cross-correlate raw EM images with templates.
To obtain a simulated dataset we produced sets of 20 micrographs, each containing 250 randomly oriented TRPC4 particles, and a different noise level k. To generate realistic noise we followed a similar procedure as described in Baxter et al.27. To simulate structural noise, we first added zero mean Gaussian noise with a SD of 88*k. Then we applied a contrast transfer function (CTF) with a fixed defocus of 2.5 μm, voltage of 300 kV, amplitude contrast of 0.1, and a B-factor of 200. Finally, to simulate shot and digitalization noise, we added mean free Gaussian noise again with a standard deviation (SD) of 51*k. With this procedure we produced five sets with k = 1, 4, 6, 8, and 10. The corresponding signal to noise ratios (SNR), defined by signal power divided by noise power, are 1.25, 0.093, 0.041, 0.023, and 0.014, respectively.
A common benchmark dataset for particle picking is a published set of cryo-EM images of KLH24. Besides KLH particles, the dataset contains mitochondrial antiviral signaling (MAVS) filaments, stacked KLH particles, and broken particles. Ideally, all of these contaminants would be ignored by a particle-picking procedure and only intact and single KLH particles selected. An additional advantage of the dataset is that the micrographs were recorded as pairs with high and low defocus, which allows to evaluate the performance of a particle picker for each defocus separately.
Training and application of crYOLO
To train crYOLO we manually selected particles for initial training datasets. Depending on the density of particles, the heterogeneity of the background, and the variation of defocus, more or fewer micrographs are needed. For the TcdA1, NOMPC, and Prx3 datasets, we found that 200–2500 particles from at least 5 micrographs were sufficient to properly train the networks for the 3 datasets. It should be emphasized that the picking of negative examples (including background, carbon edges, ice contamination, and bad particles) is not required at all in crYOLO, as essentially all other positions are considered to be negative examples. It is sufficient that these contaminants are present in the training images with the labeled particles. Ideally, each micrograph should be picked to completion. However, as the contrast in cryo-EM micrographs is often low, a user is typically not able to select all particles for training and often misses some of them, referred to as false negatives. Including false negatives carries a lower penalty than missing true positives during training (see Methods). This enables crYOLO to converge during training, even if only 20% of all particles in a micrograph are picked (see below).
To assess the performance of crYOLO, we calculated precision and recall scores28. The scores were calculated on 20% of the micrographs that were used for manual selection, but not for training. The recall score measures the ability of the classifier to detect positive examples and the precision score specifies the strength of the classifier to not label a true negative as a true positive. Both scores are summarized with the integral of the precision-recall curves, the so-called area under the curve (AUC), which is an aggregate measure for the quality of detection. The larger the AUC value, the better the performance; 1.0 is the maximum at perfect performance.
In order to quantify how well the particles are centered, we calculated the mean intersection over union (IOU) value of manually selected particles vs. the automatically picked boxes using crYOLO. The IOU is defined as a ratio of the intersecting area of two bounding boxes and the area of their union, and is a common measure for the localization accuracy. Picked particles with an IOU higher than 0.6 were classified as true positive. The mean IOUs for TcdA1, Prx3, and NOMPC were 0.86 ± 0.009, 0.80 ± 0.004, and 0.85 ± 0.007, respectively. This indicates a high localization accuracy of crYOLO.
To further assess the quality of particles picked by crYOLO, we additionally calculated two-dimensional (2-D) classes using the iterative stable alignment and clustering approach29 (ISAC) as implemented in the SPHIRE software package25. The 2-D clustering using ISAC relies on the concepts of stability and reproducibility, and is capable of extracting validated and homogeneous subsets of images that can be reliably processed in steps further downstream in the workflow. The number of particles in these subsets indirectly reflects the performance of the particle picker. Moreover, we calculated 3-D reconstructions and compared it with the published reconstruction.
For TcdA1, crYOLO was trained on 10 micrographs with 1100 particles and selected 10,854 particles from 98 micrographs. This is ~29% more particles than previously identified by the Gauss-Boxer in EMAN217 (Fig. 3a–c). Furthermore, the number of “good” particles after 2-D classification is higher (Fig. 3d, e), indicating that crYOLO is able to identify more true-positive particles. This results in a slightly improved resolution of the 3-D reconstruction (Fig. 3f, g).
Fig. 3.

Selection of TcdA1 particles and structural analysis. a–c Representative digital micrograph (micrograph number 0169) taken from the EMPIAR-10089 dataset. Red boxes indicate the particles selected by a Gauss-Boxer, b crYOLO, or c the generalized crYOLO network. Scale bar, 50 nm. d Summary of particle selection and structural analysis of the three datasets. All datasets were processed using the same workflow in SPHIRE. e Representative reference-free 2-D class averages of TcdA1 obtained using the ISAC and Beautifier tools (SPHIRE) from particles picked using crYOLO. Scale bar, 10 nm. f Fourier shell correlation (FSC) curves of the 3-D reconstructions calculated from the particles selected in crYOLO and Gauss-Boxer. The FSC 0.143 between the independently refined and masked half-maps indicates resolutions of ~3.4 and ~3.5 Å, respectively. g The final density map of TcdA1 obtained from particles picked by crYOLO is shown from the side and is colored by subunit. The reconstruction using particles from the generalized crYOLO network is indistinguishable
For NOMPC, the original authors picked initially 337,716 particles from 1873 micrographs using RELION7 (Fig. 4b, c) and obtained a 3-D reconstruction at a resolution of 3.55 Å from 175,314 particles after extensive cleaning of the dataset by 2-D and 3-D classification using RELION30. CrYOLO was trained on 9 micrographs with 585 particles and selected only 226,289 particles (>30% fewer) (Fig. 4a, c). After 2-D classification and the removal of bad classes (Fig. 4c, d), 171,917 particles were used for the 3-D refinement yielding a slightly improved cryo-EM structure of NOMPC of 3.4 Å (Fig. 4e–f). The reconstruction shows high-resolution features in the region of the ankyrin repeats (Fig. 4g). Although the number of initially selected particles differed tremendously from crYOLO, the number of particles used for the final reconstruction is very similar, indicating that crYOLO selects less false-positive particles than the selection tool in RELION. In this case, this also reduced the steps and overall time of image processing, as 2-D classification was performed on a much lower starting number of particles and further cleaning of the dataset by a laborious 3-D classification was unnecessary.
Fig. 4.

Selection of NOMPC particles and structural analysis. a, b Representative micrograph (micrograph number 1854) of the EMPIAR-10093 dataset. Particles picked by a crYOLO or b RELION, respectively, are highlighted by red boxes. Scale bar, 50 nm. c Summary of particle selection and structural analysis using RELION and crYOLO/SPHIRE. d Representative reference-free 2-D class averages obtained using the ISAC and Beautifier tools (SPHIRE) from particles selected by crYOLO. Scale bar, 10 nm. e FSC curves and f final 3-D reconstruction of the NOMPC dataset obtained from particles picked using crYOLO and processed with SPHIRE. The 0.143 FSC between the masked and unmasked half-maps indicates resolutions of 3.4 and 3.8 Å, respectively. The 3-D reconstruction is shown from the top and side. To allow better visualization of the nanodisc density, the unsharpened (gray, transparent) and sharpened map (colored by subunits) are overlaid. g Comparison of the density map obtained by crYOLO/SPHIRE with the deposited NOMPC 3-D reconstruction EMDB-8702
For Prx3, crYOLO was trained on 5 micrographs with 2500 particles and identified 354,581 particles from 802 digital micrographs, which is comparable to the number of particles picked in the original article using EMAN217 (Fig. 5a–c). We used two consecutive ISAC rounds to classify the particles in 2-D (Fig. 5d). In the first round, we classified the whole dataset into large classes to identify the overall orientation of the particles. In the second round, we split them into top, side, and tilted views, and ran independent classifications on each of them. This procedure is necessary to avoid centering errors in ISAC. After discarding bad classes and removing most of the preferential top view orientations, the dataset was reduced to 37,031 particles. The 3-D refinement of the remaining set of particles in SPHIRE yielded a map at a nominal resolution of 4.6 Å (Fig. 5e, f). In comparison, the final stack in the original article was composed of only 8562 particles, which were refined to a 4.4 Å resolution map in RELION. In contrast to our processing pipeline, the original authors used three rounds of 2-D and four rounds of 3-D classification in RELION7, in order to clean this dataset.
Fig. 5.

Selection of Prx3 particles and structural analysis. a, b Particles selected on a representative micrograph (micrograph number 19.22.14) of the EMPIAR-10050 dataset using either a crYOLO or b EMAN2. Scale bar, 100 nm. c Summary of particle selection and structural analysis. The resolution in parentheses is the result obtained after a 3-D refinement performed in SPHIRE using the final 8562 particles of the original dataset. d Representative 2-D class averages obtained from two rounds of classification using the crYOLO-selected particles and ISAC. Scale bar, 10 nm. Well-centered examples for all views showing high-resolution details can be readily obtained from the data. e Fourier shell correlation plots for the final 3-D reconstruction (black) using the crYOLO-selected particles or the 8562 particles from the original dataset (gray). The average resolution of our 3-D reconstruction is ~4.6 Å, whereas that one from the originally used particles is ~4.5 Å. f Top and side views of the 3-D reconstruction obtained with crYOLO/SPHIRE. For clarity, all subunits are colored differently in the reconstruction
The small differences in the final resolution of the reconstructions can be attributed to the fact that all reconstructions reach resolutions near the theoretical resolution limit; at a pixel size of 1.74 Å, 4.4 Å represents ~0.8 times Nyquist. Consistent with this observation, the FSC curves of the reconstructions from both sets of particles are practically superimposable (Fig. 5e). Interestingly, a large fraction of the particles was discarded during refinement, both by the original authors—97.6%—and by us—87.7%. In our case, this is not due to the quality of the particle selection. Indeed, 310,123 of our picks were included into the stably aligned images after ISAC. Instead, we deleted most of the particles that were found in the preferred top orientation, which represents roughly half of the dataset. The remaining discarded images correspond to correct picks in regions where the protein forms clusters. In those cases, although the particles were correctly identified by crYOLO, they were not used for single-particle reconstruction.
In order to evaluate the SNR dependency on the picking quality of crYOLO, we used the simulated data and trained a model for each noise-level set, using the 15 micrographs for training and 4 for validation for each set. Finally, we calculated the AUC for each of the sets (Fig. 6a). Up to a noise level of 6 the AUC value is >0.8 and even for a noise level of 8 the AUC values stay >0.6. In images with a noise level of ≥8 the particles are visually barely distinguishable from the background (Fig. 6b). This demonstrates the strength of crYOLO for selecting particles of near-to-focus cryo-EM datasets that naturally have a low contrast.
Fig. 6.

SNR dependence of crYOLO. a Noise-level dependency of crYOLO picking simulated TRPC4 particles (EMD-4339) measured by the area under the precision-recall curve (AUC). The AUC stays above 0.8 up to a noise level of 6 (SNR 0.041). b Example micrographs for the noise levels of 1, 4, and 8
To determine the influence of the size of the training dataset on selection quality, we trained crYOLO on the manually picked KLH dataset and on subsets of different size. We decreased the number of micrographs in each subset gradually and evaluated the results by calculating the precision and recall scores. The selection performance was excellent, for both the high defocus and the low defocus micrographs (Fig. 7a). To demonstrate the discrimination power, we trained a model to pick only the side views of KLH. The performance was comparable with picking all views (Fig. 7b). Even if the model was trained with as few as 40 KLH particles from 1 micrograph pair, crYOLO still demonstrated a high AUC value of 0.9 (Fig. 7c, Table 1). All trained models skipped MAVS filaments, stacked KLHs, and broken particles. This demonstrates that crYOLO can be easily and accurately trained with only a small number of particles.
Fig. 7.

Training of crYOLO on KLH. a One example of a particle picking result by crYOLO trained for all views with 14 micrographs of the full KLH dataset and b trained only for side views. Scale bar, 70 nm. c Precision-recall curves for the low defocus micrographs of the KLH dataset using several training set sizes (Supplementary Data 1). The curves were estimated based on 17 randomly selected test micrographs out of the full dataset. The AUC values are 0.97 (blue), 0.94 (orange), 0.9 (green)
Table 1.
Evaluation table for different subsets of the KLH micrographs
| Mic. pairs | Trained for | AUC | Precision | Recall | IOU | |
|---|---|---|---|---|---|---|
| Low defocus | 65 | All views | 0.97 | 0.92 | 0.96 | 0.94 |
| 14 | All views | 0.95 | 0.93 | 0.9 | 0.94 | |
| 7 | All views | 0.97 | 0.89 | 0.94 | 0.92 | |
| 4 | All views | 0.94 | 0.84 | 0.94 | 0.92 | |
| 2 | All views | 0.92 | 0.86 | 0.91 | 0.92 | |
| 1 | All views | 0.9 | 0.85 | 0.9 | 0.89 | |
| 14 | Side views | 0.91 | 0.89 | 0.89 | 0.92 | |
| High defocus | 65 | All views | 0.99 | 0.99 | 0.98 | 0.95 |
| 14 | All views | 0.97 | 0.91 | 0.99 | 0.95 | |
| 7 | All views | 0.98 | 0.92 | 0.96 | 0.94 | |
| 4 | All views | 0.96 | 0.98 | 0.87 | 0.94 | |
| 2 | All views | 0.9 | 0.86 | 0.95 | 0.94 | |
| 1 | All views | 0.91 | 0.85 | 0.91 | 0.91 | |
| 14 | Side views | 0.97 | 0.91 | 0.97 | 0.95 |
The AUC, Precision, Recall, and IOU were evaluated for various training sets comprising a different number of micrograph pairs. In addition, a subset with 14 micrographs was trained to pick only side views. The IOU and the AUC values of all experiments are between 0.89 and 0.97
Although it might be ideal to provide a training set where the particles were picked to completion for each micrograph, it is still often difficult or, with crowded micrographs, a time-consuming burden. To illustrate the performance of crYOLO on sparsely labeled data we used the TcdA1 dataset and randomly removed 80% of the labeled particles (Supplementary Fig. 1a,b). The training dataset where the particles were picked to completion contains 10 micrographs with 1100 particles. The sparsely labeled dataset contains the same micrographs but with only 205 particles labeled. The performance on the sparsely labeled dataset (AUC = 0.88) is still very good when compared to the training set which was picked to completion (AUC = 0.95) (Supplementary Fig. 1c).
Computational efficiency
We used a desktop computer equipped with an NVIDIA GeForce GTX 1080 graphics card with 8 GB memory and an Intel Core i7 6900K CPU to train crYOLO and select the particles using the GPU. The time needed for training was 5–6.5 min for each dataset (Fig. 8). The selection of the Prx3, TcdA1, and NOMPC datasets reached a speed of ~4.6, ~5.0, and ~4.2 micrographs per second, respectively. However, if no GPU is available, a trained model can also be used on a common CPU. The picking speed on our multi-core CPU is one second per micrograph.
Fig. 8.
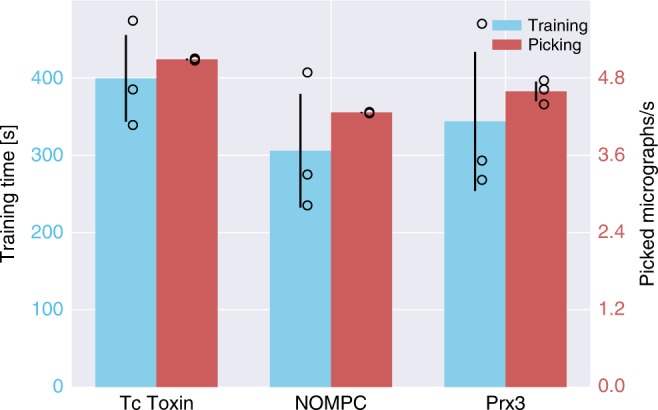
Computational efficiency statistics of training and particle selection. The crYOLO training times for TcdA1, NOMPC, and Prx3 were 400s, 300s, and 343s, respectively (blue bars). The red bars depict the number of processed micrographs per second during particle selection. Error bars represent SD, measured by training and applying crYOLO three times (black circle). All datasets were picked in less than a quarter of a second per image. For TcdA1, crYOLO needed 0.19 s per micrograph, 0.23 s for NOMPC, and 0.21 s for Prx3
In comparison the software DeepPicker8 reports 1.5 min per micrograph on a GPU and the software DeepEM9 reports 13 min on a CPU and 40s on a GPU.
Generalization to unseen datasets
Ideally, crYOLO would specifically recognize and select particles that it has not seen before. In order to reach this level of generalization, we trained the network with a combination of 840 micrographs from 45 datasets containing 26 manually picked datasets, 9 simulated datasets, and 10 particle-free datasets consisting only of contamination. The molecular weight of the complexes from the respective datasets ranged from 64 kDa to 1.1 MDa.
Using this generalized crYOLO network, we automatically selected particles of RNA polymerase31 (EMPIAR 10190) and glutamate dehydrogenase (EMPIAR 10217). Although crYOLO had not been trained on these particles, it specifically selected particles while avoiding contamination (Fig. 9a, b).
Fig. 9.

Generalized crYOLO network. a, b Particles selected on a representative micrograph of glutamate dehydrogenase (EMPIAR 10127) and RNA polymerase (EMPIAR 10190). None of the datasets were included in the set used for training the generalized crYOLO network. Scale bars, 50 nm. c AUC, recall, and precision of the datasets included into the general model evaluated for the crYOLO network architecture and the Inception-ResNet (IR) architecture (Supplementary Data 2). The box shows the lower and upper quartiles with a line as median. The whiskers represent the range of the data, whereas the points represent outliers. d Precision-recall curves for TcdA1 picked with either a network directly trained on the TcdA1 dataset (orange) and the general model but not on TcdA1 (blue) (Supplementary Data 3)
In recent years, several new network architectures have been published. These networks have a higher capacity and allow the training of more than 100 layers. One of these networks is Inception-ResNet32, which achieved high accuracy at the ImageNet competition33. To compare this network with the crYOLO network, we replaced the feature extraction part of crYOLO with the Inception-ResNet architecture and trained it on the complete combination of datasets. The Inception-ResNet architecture achieved the same performance as the crYOLO architecture (Fig. 9c). As illustrated here, the median AUC/recall/precision is approximately the same. Thus, what we conclude is that, given our current training set, increasing the capacity of the network does not result in improved performance. Bigger networks have the advantage that they have a higher capacity, i.e., can be trained to arbitrary depth. However, they tend to overfit more easily and the Inception-ResNet architecture comes with a higher computational cost. In addition, it was shown recently that many layers in modern ResNets do not learn anything34 and can be safely removed without damaging the performance of the network. This means that a network with the same prediction quality but lower number of weights, such as the one we used for crYOLO, is actually an advantage in a practical setup with limited training data. However, it is noteworthy to highlight that crYOLO offers the possibility of the integration of new architectures in a straightforward manner, which will contribute to its sustainability.
To assess the quality of the generalized crYOLO network, we compared its performance selecting TcdA1 with that of a network that was directly trained on this protein. As expected, the AUC and localization accuracy are better for the directly trained network (Fig. 9d). However, the generalized crYOLO network selected a similar number of particles (Fig. 3d); the AUC of 0.84 (Fig. 9d) and the IOU of 0.79 show that its performance is of sufficient quality to select a good set of particles. Indeed, the TcdA1 particles selected by the generalized crYOLO network resulted in a reconstruction of similar resolution and quality as the particles from the TcdA1-trained crYOLO (Fig. 3f). We expect that increasing the training set with a higher number of proteins and complexes will further expand the capabilities of crYOLO.
Discussion
In this work we present crYOLO, a particle-picking procedure for cryo-EM. CrYOLO employs a state-of-the-art deep-learning object-detection system which contains a 21-layer CNN. The excellent performance of crYOLO on several recent direct detector datasets reflects the efficiency of the program to detect good particles at high speed with an accuracy comparable to manual particle selection. CrYOLO’s speed and efficiency underlines its potential to become a crucial component in a streamlined automated workflow for single-particle analysis and thus eliminates one of the remaining bottlenecks.
To close the remaining gaps in the automated workflow between the electron microscope and data processing, such as evaluation, drift, CTF correction, and file conversion and transfer, our lab has developed an on-the-fly processing pipeline called TranSPHIRE, which is documented and freely available on www.sphire.mpg.de (manuscript in preparation). The TranSPHIRE pipeline includes crYOLO.
The crYOLO package is available as a standalone program and is easily integrated in existing workflows. It will be fully integrated into the SPHIRE image processing workflow and into the Scipion framework35. In addition to a command line interface crYOLO provides an easy-to-use graphical tool for simple and direct generation of training data, as well as visual evaluation of picking results (Fig. 2). CrYOLO outputs the coordinate files in EMAN box-file and RELION STAR-file format, which can easily be imported to all available software packages for single-particle analysis. CrYOLO picks micrographs rapidly on a standard GPU. If a trained network is available, particle picking can also be performed on multi-core CPUs. However, this decreases the speed of selection to 1s per micrograph. This is still faster than other particle pickers on a GPU. Some of these differences can be attributed to the exact model of CPUs and GPUs used. However, the use of the YOLO approach clearly plays a strong role in the particle-picking time. Moreover, this approach is not only faster but has the capability to learn the context of the particle such as to avoid picking inside or in the vicinity of contamination. This feature is particularly important for specimens that pose a challenge due to the high amount of contamination present. In addition, what further contributes to the overall value of our crYOLO package includes the low number of micrographs that are required during the training process, the possibility of training with sparsely labeled data, and elimination of the need for the user to pick negative examples.
Our evaluations demonstrate that the accuracy of crYOLO yields a set of particles with a remarkably low number of false positives. In addition, the IOU values of the selected particles provide evidence for the excellent centering performance of crYOLO. These are both strengths that contribute to improving the quality of the input dataset for image processing and ultimately improve the final reconstruction in many cases. At the same time, it reduces the number of subsequent processing steps, such as computationally intensive 3-D classifications, and thereby shortens the overall processing time.
In comparison with template-based particle-picking approaches, crYOLO is not prone to template bias and therefore the danger of picking “Einstein from noise”36 is reduced. However, the user should select the particles for the training data without bias. Otherwise, the applied supervised learning method will create a biased model. For example, if a view of the particle is completely missing in the training data, crYOLO might learn to skip this view. Therefore, the micrographs in the training dataset should be representative of the full dataset with respect to range of defocus, types of contamination, etc.
Importantly, crYOLO can be trained to select previously unseen datasets. With the use of a training set containing 10–20 micrographs from 45 datasets, we have obtained a powerful general network. Adding more training datasets from different projects that were manually picked by experienced users will improve the performance of crYOLO even further. In crYOLO adding more training data is especially easy, because in contrast to the sliding-window approaches, training crYOLO only requires particles to be labeled and not the background nor any contamination. Therefore, previously processed datasets can be directly used to train crYOLO directly without manually picking background and contamination afterwards. In principle, it is also feasible to train generalized networks that are specialized on certain types of particles, such as viruses, elongated or very small particles to improve the performance of crYOLO. At the moment, the development of CNNs is moving fast and new CNN networks, which outperform earlier networks, are released frequently. The flexible software architecture of crYOLO allows an easy incorporation of new CNNs and therefore a straightforward adaptation to new developments, if necessary.
Taken together, crYOLO enables rapid fully automated on-the-fly particle selection with comparable precision as manual picking, without bias from reference templates. Furthermore, with the use of the generalized model presented here, our particle picker, crYOLO, can be used without a template or human intervention to select particles on most single-particle cryo-EM projects within a very short period of time.
Methods
CrYOLO architecture and training
CrYOLO trains a deep CNN for automated particle selection. A typical CNN consists of multiple convolutional and pooling layers, and is characterized by the depth of the network, which is the number of convolutional layers37. The input of a CNN is data with a grid-like topology, most often images that may have multiple channels (e.g., color). A convolutional layer consists of a fixed number of filters and each filter consists of multiple convolutional kernels with a fixed size (typically 1 × 1, 3 × 3 or 5 × 5). The number of convolutional kernels per filter is equal to the number of channels in the input data of the layer. During forward propagation, each filter in a convolutional layer creates a feature map, which is a channel in the final output of the layer. Each kernel is convolved with the corresponding input channel. The results are summed up along the channels, resulting in the first channel of the output. The same procedure is applied for the second filter, which results in the second channel, and so on. After all channels are calculated, the so-called batch normalization38 is applied. Batch normalization normalizes the feature map and leads to faster training. Furthermore, it has some regularization power, which makes further regularization often unnecessary. Finally, a nonlinear activation function is applied element-wise to the normalized feature map, which results in the final feature map.
In typical CNNs, max-pooling layers are inserted between some of the convolutional layers. Max-pooling layers divide the input image into equal sized tiles (e.g., 2 × 2), which are then used to calculate a condensed feature map. Therefore, for each tile a cell is created; the maximum for the tile is computed and inserted into the cell. This leads to a reduced dimensionality of the feature map and makes the network more memory-efficient and robust against small perturbations in the pixel values.
The network architecture used in crYOLO is summarized in Table 2. The feature extraction part consists of 21 convolutional layers and each convolutional layer consists either of multiple 3 × 3 filters or 1 × 1 filters. In addition, it contains a passthrough layer between layer 13 and 21, which enables the network to use low-level features during prediction. All convolutional layers use padding so that they do not reduce the size of their input. We used the leaky rectified linear unit39 (LRELU) as activation function. LRELU simply returns the element itself if the element is positive and the element multiplied by a fixed constant α (here α = 0.1), if it is negative.
Table 2.
YOLO network architecture
| Layer | Type | Filters | Size |
|---|---|---|---|
| Feature extraction | |||
| 1 | Convolutional | 32 | 3 × 3 |
| Max-pooling | |||
| 2 | Convolutional | 64 | 3 × 3 |
| Max-pooling | |||
| 3 | Convolutional | 128 | 3 × 3 |
| 4 | Convolutional | 64 | 1 × 1 |
| 5 | Convolutional | 128 | 3 × 3 |
| Max-pooling | |||
| 6 | Convolutional | 256 | 3 × 3 |
| 7 | Convolutional | 128 | 1 × 1 |
| 8 | Convolutional | 256 | 3 × 3 |
| Max-pooling | |||
| 9 | Convolutional | 512 | 3 × 3 |
| 10 | Convolutional | 256 | 1 × 1 |
| 11 | Convolutional | 512 | 3 × 3 |
| 12 | Convolutional | 256 | 1 × 1 |
| 13 | Convolutional | 512 | 3 × 3 |
| Max-pooling | |||
| 14 | Convolutional | 1024 | 3 × 3 |
| 15 | Convolutional | 512 | 1 × 1 |
| 16 | Convolutional | 1024 | 3 × 3 |
| 17 | Convolutional | 512 | 1 × 1 |
| 18 | Convolutional | 1024 | 3 × 3 |
| 19 | Convolutional | 1024 | 3 × 3 |
| 20 | Convolutional | 1024 | 3 × 3 |
| 21 | Convolutional | 1024 | 3 × 3 |
| Detection | |||
| Dropout (0.2) | |||
| 1 | Convolutional | 6 | 1 × 1 |
The CNN consists of 21 convolutional and 5 max-pooling layers for feature extraction. For detection, a single convolutional layer is used with a dropout layer in front to reduce overfitting. The dropout layer is only used during training, not during prediction
Five max-pooling layers downsample the image by a factor of 32. During training, a dropout unit in the detection part sets 20% of the entries in the feature map after layer 21 to 0. This regularizes the network and reduces overfitting. Finally, a convolution layer with six 1 × 1 filters performs the actual detection. With the YOLO18 approach, each cell in the final feature map is used to classify if the center of a particle box is inside this grid cell, and if this is the case it predicts the exact position of the box center relative to the cell, as well as the width and height of the box.
The network was trained using backpropagation with the stochastic optimization procedure ADAM40. Backpropagation applies the chain rule to compute the gradient values in every layer. The gradient determines how the kernel elements in each convolutional layer should be updated to get a lower loss. The optimizer determines how the gradient of the loss is used to update the network parameters. For YOLO, the loss function to be minimized is given by:
where is 1 if the center of a particle is in cell i and 0 otherwise, λcoord, λobj, and λnoobj constant weights, (xi,yi) the centrum coordinates of the training boxes, and the predicted coordinates. The width and height of the box with their estimates are given by (wi,hi)) and () respectively. The confidence that a cell contains a particle is Ci.
The first term of the loss function penalizes bad localization of particle boxes. The second term penalizes inaccurate estimates for the width and height of the boxes. The third term attempts to increase the confidence for cells with a particle inside. The last term decreases the confidence of those cells containing no particle center. The loss function slightly differs from the one used in Redmon et al.14, as we only have a single class to predict and also only one reference box (anchor box in Redmon et al.14).
Data augmentation
During training, each image is augmented before passing it through the network. This means that it is slightly altered by random selection methods instead of passing the original image through the network. Each image is passed multiple times through the network, randomly modified in different ways. This helps the network to reduce overfitting and also the amount of training data needed. The applied methods are as follows:
Gaussian blurring: a random SD between 0 and 3 is selected and then a corresponding filter mask is created. This mask is then convolved with the input image. Average blurring: a random mask size between 2 and 7 is chosen. This mask is shifted over the image. At each position, the central element is replaced with the mean values of its neighbors. Flip: the image is mirrored along the horizontal and vertical axes. Noise: Gaussian noise with a randomly selected SD proportional to the image SD is added to the image. Dropout: randomly replaces 1–10% of the image pixels with the image mean value. Contrast normalization: the contrast is changed by subtracting the median pixel value from each pixel, multiplying them by a random constant and finally adding the median value again.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Supplementary information
Acknowledgements
We thank Yifan Cheng and Radostin Danev for kindly providing us the original coordinates of the NOMPC and Prx3 particles, respectively. This work was supported by the Max Planck Society (S.R.) and the European Council under the European Union’s Seventh Framework Programme (FP7/2007–2013) (Grant 615984 to S.R.).
Author contributions
S.R. designed the project. T.W. conceived the approach and wrote the software. T.W., C.G., M.S. and T.M. optimized the approach. T.W., C.G. and F.M. evaluated the performance. T.W., C.A., A.A., P.H., O.S., T.R., T.R.S., D.P., D.R., D.Q., S.T., B.S., E.S. and P.L. picked datasets for the general model. T.W. and S.R. wrote the manuscript with input from all authors. All authors discussed the results and the manuscript.
Data availability
The training datasets for this study are available from the corresponding author upon reasonable request. CrYOLO—along with a detailed practical manual—is available for download under http://sphire.mpg.de.
Code availability
The source code is contained in the crYOLO software package (http://sphire.mpg.de) and its use is restricted by the end user license agreement: http://sphire.mpg.de/wiki/doku.php?id=cryolo_license.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information accompanies this paper at 10.1038/s42003-019-0437-z.
References
- 1.Voss NR, Yoshioka CK, Radermacher M, Potter CS, Carragher B. DoG Picker and TiltPicker: software tools to facilitate particle selection in single particle electron microscopy. J. Struct. Biol. 2009;166:205–213. doi: 10.1016/j.jsb.2009.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ogura T, Sato C. An automatic particle pickup method using a neural network applicable to low-contrast electron micrographs. J. Struct. Biol. 2001;136:227–238. doi: 10.1006/jsbi.2002.4442. [DOI] [PubMed] [Google Scholar]
- 3.Volkmann N. An approach to automated particle picking from electron micrographs based on reduced representation templates. Journal of Structural Biology. 2004;145(1-2):152–156. doi: 10.1016/j.jsb.2003.11.026. [DOI] [PubMed] [Google Scholar]
- 4.Nicholson WV, Glaeser RM. Review: automatic particle detection in electron microscopy. J. Struct. Biol. 2001;133:90–101. doi: 10.1006/jsbi.2001.4348. [DOI] [PubMed] [Google Scholar]
- 5.Huang Z, Penczek PA. Application of template matching technique to particle detection in electron micrographs. J. Struct. Biol. 2004;145:29–40. doi: 10.1016/j.jsb.2003.11.004. [DOI] [PubMed] [Google Scholar]
- 6.Roseman AM. FindEM—a fast, efficient program for automatic selection of particles from electron micrographs. J. Struct. Biol. 2004;145:91–99. doi: 10.1016/j.jsb.2003.11.007. [DOI] [PubMed] [Google Scholar]
- 7.Scheres SHW. Semi-automated selection of cryo-EM particles in RELION-1.3. J. Struct. Biol. 2015;189:114–122. doi: 10.1016/j.jsb.2014.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang F, et al. DeepPicker: a deep learning approach for fully automated particle picking in cryo-EM. J. Struct. Biol. 2016;195:325–336. doi: 10.1016/j.jsb.2016.07.006. [DOI] [PubMed] [Google Scholar]
- 9.Zhu Y, Ouyang Q, Mao Y. A deep convolutional neural network approach to single-particle recognition in cryo-electron microscopy. BMC Bioinformatics. 2017;18:29. doi: 10.1186/s12859-017-1757-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tegunov, D. & Cramer, P. Real-time cryo-EM data pre-processing with Warp. bioRxiv 338558 (2018). 10.1101/338558.
- 11.Bepler, T. et al. Positive-unlabeled convolutional neural networks for particle picking in cryo-electron micrographs. Res. Comput. Mol. Biol. 10812, 245–247 (2018). [PMC free article] [PubMed]
- 12.Rawat W, Wang Z. Deep convolutional neural networks for image classification: a comprehensive review. Neural Comput. 2017;29:2352–2449. doi: 10.1162/neco_a_00990. [DOI] [PubMed] [Google Scholar]
- 13.Schmidhuber J. Deep learning in neural networks: an overview. Neural Netw. 2015;61:85–117. doi: 10.1016/j.neunet.2014.09.003. [DOI] [PubMed] [Google Scholar]
- 14.Redmon, J., Divvala, S., Girshick, R. & Farhadi, A. In 2016 IEEE Conf. Comput. Vis. Pattern Recognition (CVPR). 779–788 (IEEE, 2016). 10.1109/CVPR.2016.91.
- 15.Anh, H. N., Suppa, M., Sirotenko, M. & Schoen, A. J. experiencor/basic-yolo-keras v0.1. https://doi.org/10.5281/zenodo.1168755 (2018).
- 16.Chollet, F. C. O. et al. Keras (GitHub, 2015).
- 17.Tang G, et al. EMAN2: an extensible image processing suite for electron microscopy. J. Struct. Biol. 2007;157:38–46. doi: 10.1016/j.jsb.2006.05.009. [DOI] [PubMed] [Google Scholar]
- 18.Redmon, J. & Farhadi, A. In 2017 IEEE Conf. Comput. Vis. Pattern Recognition (CVPR) 6517–6525 (IEEE, 2017). 10.1109/CVPR.2017.690.
- 19.He, K., Zhang, X., Ren, S. & Sun, J. 2016 IEEE Conf. Comput. Vis. Pattern Recognition (CVPR) (IEEE, 2016).
- 20.Gatsogiannis C, et al. A syringe-like injection mechanism in Photorhabdus luminescens toxins. Nature. 2013;495:520–523. doi: 10.1038/nature11987. [DOI] [PubMed] [Google Scholar]
- 21.Jin P, et al. Electron cryo-microscopy structure of the mechanotransduction channel NOMPC. Nature. 2017;547:118–122. doi: 10.1038/nature22981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Khoshouei M, et al. Volta phase plate cryo-EM of the small protein complex Prx3. Nat. Commun. 2016;7:10534. doi: 10.1038/ncomms10534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vinayagam D, et al. Electron cryo-microscopy structure of the canonical TRPC4 ion channel. eLife. 2018;7:213. doi: 10.7554/eLife.36615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhu Y, et al. Automatic particle selection: results of a comparative study. J. Struct. Biol. 2004;145:3–14. doi: 10.1016/j.jsb.2003.09.033. [DOI] [PubMed] [Google Scholar]
- 25.Moriya, T. et al. High-resolution single particle analysis from electron cryo-microscopy images using SPHIRE. J. Vis. Exp. 1–11 (2017). 10.3791/55448. [DOI] [PMC free article] [PubMed]
- 26.Khoshouei M, Radjainia M, Baumeister W, Danev R. Cryo-EM structure of haemoglobin at 3.2 Å determined with the Volta phase plate. Nat. Commun. 2017;8:16099. doi: 10.1038/ncomms16099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Baxter WT, Grassucci RA, Gao H, Frank J. Determination of signal-to-noise ratios and spectral SNRs in cryo-EM low-dose imaging of molecules. J. Struct. Biol. 2009;166:126–132. doi: 10.1016/j.jsb.2009.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Goutte, C. & Gaussier, E. in Advances in Information Retrieval3408, 345–359 (Springer, Berlin, Heidelberg, 2005).
- 29.Yang Z, Fang J, Chittuluru J, Asturias FJ, Penczek PA. Iterative stable alignment and clustering of 2D transmission electron microscope images. Structure. 2012;20:237–247. doi: 10.1016/j.str.2011.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Scheres SHW. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 2012;180:519–530. doi: 10.1016/j.jsb.2012.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vorländer MK, Khatter H, Wetzel R, Hagen WJH, Müller CW. Molecular mechanism of promoter opening by RNA polymerase III. Nature. 2018;553:295–300. doi: 10.1038/nature25440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Szegedy, C., Ioffe, S., Vanhoucke, V. & Alemi, A. Inception-v4, inception-ResNet and the impact of residual connections on learning. arXiv:1602.07261 (2016).
- 33.Russakovsky O, et al. ImageNet large scale visual recognition challenge. Int J. Comput Vis. 2015;115:211–252. doi: 10.1007/s11263-015-0816-y. [DOI] [Google Scholar]
- 34.Huang, G., Liu, Z., Maaten, L. V. D. & Weinberger, K. Q. In 2017 IEEE Conf. Comput. Vis. Pattern Recognition (CVPR) 2261–2269 (IEEE, 2017). 10.1109/CVPR.2017.243.
- 35.de la Rosa-Trevín JM, et al. Scipion: a software framework toward integration, reproducibility and validation in 3D electron microscopy. J. Struct. Biol. 2016;195:93–99. doi: 10.1016/j.jsb.2016.04.010. [DOI] [PubMed] [Google Scholar]
- 36.Henderson R. Avoiding the pitfalls of single particle cryo-electron microscopy: Einstein from noise. Proc. Natl. Acad. Sci. USA. 2013;110:18037–18041. doi: 10.1073/pnas.1314449110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lecun Y, Bottou L, Bengio Y, Haffner P. Gradient-based learning applied to document recognition. Proc. IEEE. 1998;86:2278–2324. doi: 10.1109/5.726791. [DOI] [Google Scholar]
- 38.Ioffe, S. & Szegedy, C. Batch normalization - accelerating deep network training by reducing internal covariate shift. arXiv:1502.03167 (2015).
- 39.Maas, A. L., Hannun, A. Y. & Ng, A. Y. Rectifier nonlinearities improve neural network acoustic models. 30 (2013).
- 40.Kingma, D. P. & Ba, J. Adam: a method for stochastic optimization. arXiv:1412.6980 (2014).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The training datasets for this study are available from the corresponding author upon reasonable request. CrYOLO—along with a detailed practical manual—is available for download under http://sphire.mpg.de.
The source code is contained in the crYOLO software package (http://sphire.mpg.de) and its use is restricted by the end user license agreement: http://sphire.mpg.de/wiki/doku.php?id=cryolo_license.