

Abstract
The notion that inflammation is a critical component of cancer has been researched extensively. Tumor-associated macrophages (TAMs) are among the inflammatory cells that greatly influence cancer. In the tumor microenvironment (TME), macrophages can either stimulate or inhibit tumorigenesis. TAMs that stimulate tumor cell proliferation (M2-phenotype) enrich the TME with growth factors and immunosuppressive molecules, whereas tumor inhibitory TAMs (M1-phenotype) initiate the immune response to dampen tumor progression. Shifting between phenotypes is controlled by several components of the TME. Targeting macrophages, specifically inhibiting M2 TAMs, has been introduced successfully in cancer immunotherapy. However, signaling mechanisms underlining TAM polarization are largely unknown. This review analyzed studies of the role of mitogen-activated protein kinase (MAPK) as a determinant of macrophage polarization. It is proposed that activation of MAPK, particularly extracellular signal-regulated kinase 1/2 and p38, might favor the differentiation into M2 TAMs. Thus, pharmacological modification of MAPK pathways will potentially offer exciting new targets in cancer immunotherapy.
Keywords: Cancer, macrophage polarization, mitogen-activated protein kinase
INTRODUCTION
Uncontrolled cell proliferation is a well-known cause of cancer. However, several factors produced from cells of the surrounding stroma or infiltrated inflammatory cells – growth factors – as well as DNA damage-promoting agents can potentiate tumorigenesis.[1,2] Understanding the tumor microenvironment (TME) reveals broader outlooks for cancer therapy. The TME is largely invaded by inflammatory cells attracted by various chemokines and cytokines produced by the tumor cells as well as active stroma. Reciprocally, inflammatory cells produce a wide array of cytotoxic mediators, selectins, proteases, and matrix metalloproteinases (MMPs) that potentiate neoplastic progression.[3,4,5] The main population of TME inflammatory cells are macrophages, dendritic cells, mast cells, natural killer cells, neutrophils, and T-cells. Macrophages, particularly, are the most versatile immune cells that produce various factors in the TME that either promote or inhibit tumor progression.[4,5,6] According to various signals in the TME, macrophages can polarize into classically activated (M1 macrophages) or alternatively activated (M2 macrophages). Studies characterizing macrophage polarization in TMEs revealed that tumor-associated macrophages (TAMs) are M2 dominant, propagating tumorigenesis by producing immunosuppressive cytokines, angiogenic factors, and extracellular matrix (ECM)-remodeling molecules.[6,7,8] In contrast, M1-TAMs are tumor inhibitors responsible for recognition and destruction of cancer cells and produce nitric oxide (NO) as well as immunostimulatory cytokines to enhance the T helper cells’ (Th1) tumourcidal response.[6]
Converting TAM from a pro-tumor M2-phenotype to an antitumor M1-phenotype, particularly the signaling mechanisms underpinning this process, is currently a subject of interest in cancer research.[8,9,10] Mitogen-activated protein kinase (MAPK) signaling pathways play a pivotal role in regulating macrophages’ activation and proliferation.[11,12,13] In several reports, members of the MAPK family have been identified as master regulators of macrophages’ pro- and anti-inflammatory cytokine production.[14,15,16] The aim of this review is to shed a light on the involvement of MAPK signaling in TAMs’ polarization. This will ultimately help to develop new therapeutic targets against TAMs to hinder cancer development.
ORIGINS OF MACROPHAGES
TAMs can be derived from primitive yolk sac precursors or differentiated from recruited bone marrow monocytes, which accounts for the majority of TAMs.[6] Colony-stimulating factor 1 is a key chemotactic factor for macrophages, whether they are tissue resident or bone marrow derived.[17] Other growth factors, such as vascular endothelial growth factor A (VEGFA), have also shown to recruit and further differentiate monocytes to TAMs.[18] In addition, blood bone marrow monocytes are infiltrated to the site of the tumor and rapidly differentiate into TAMs in response to chemoattractants such as Monocyte Chemoattractant Protein-1 (MCP-1/CCL2), produced by endothelial cells, stromal cells, and tumor cells.[6,19]
TAMs can exhibit two different phenotypes depending on the type of signals they receive from the TME. The following section discusses the main factors affecting the polarization of TAMs.
MACROPHAGE POLARIZATION IN TUMOR MICROENVIRONMENTS
Macrophage polarization is a diverse process because it is affected by several factors in the TME, such as apoptotic cells and immune complexes.[20,21] M2 TAMs are induced in response to transforming growth factor-beta (TGF-β), glucocorticoids, and Th2 cytokines interleukin (IL) IL4, IL10, and IL13 to express CD163 and CD206 as well as several scavenger receptors.[2,20] Moreover, pro-tumor M2 phenotypes are generated from tissue stress such as hypoxia and tumor-derived co-factor high-mobility group protein box1 (HMGB-1). Cancer-associated fibroblasts produce stromal cell-derived factor-1/chemokine (CXCL12), which is also a chemoattractant of macrophages and promotes M2 macrophage polarization.[19] M2 TAMs favor tumor growth and angiogenesis and prevent antitumor cytotoxic CD8+ T-cells’ immune response via the production of cytokines–specifically IL10; chemokines such as CCL17, CCL18, and CCL22; angiogenic factors, MMPs; and ECM-remodeling molecules such as VEGF and platelet-derived growth factor [Figure 1].[6,7,8]
Figure 1.
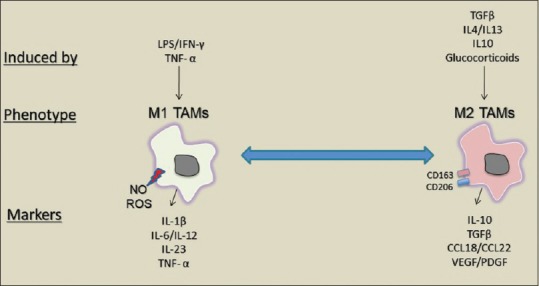
Tumor-associated macrophage activation phenotypes. Tumor-associated macrophages are activated in to (M1 phenotype) or (M2 phenotype). M1 tumor-associated macrophages produce cytokines and chemokines such as tumor necrosis factor-alpha, interleukin-1B and interleukin-12. Whereas, M2 tumor-associated macrophages express CD206 and CD163 and produce factors such as transforming growth factor-beta, interleukin-10, and CCL18
On the other hand, inhibitory M1 TAMs are activated by cytokines’ repertoire of Th1 cells, such as interferongamma (IFN-γ) and tumor necrosis factor-alpha (TNF-α) to produce reactive oxygen species (ROS) and high levels of IL-1β, IL-6, IL-12, and IL-23.. M1 TAMs play key roles in immunostimulation and antitumoral response, and accumulation of these macrophages indicates a good prognosis.[2,6,20] In the context of cancer, M1-phenotypes direct T-cells toward Th1 tumoricidal responses and play an important role in the recognition and destruction of cancer cells, and their presence indicates a good prognosis [Figure 1].[22,23]
MITOGEN-ACTIVATED PROTEIN KINASES
The MAPKs are highly conserved serine/threonine protein kinases that are involved in a wide range of macrophage activities, including proliferation, differentiation, survival, apoptosis, and immune responses.[12,24,25] Based on sequence similarity and differential activation by agonists and substrate specificity, the MAPK family exists in three components in mammalian cells. These are the extracellular signal-regulated kinases ERKs-1-4 and big MAPKs (ERK5), c-Jun N-Terminal kinases (JNK-1/2/3), and isoforms of p38 MAPKs (p38-a/b/g/d). All share the TXY-sequence, where T and Y are threonine and tyrosine and X is glutamate, proline, or glycine, in ERK, JNK, or p38 MAPKs.[25,26,27] MAPKs are activated by different stimuli such as growth environmental stresses, including osmotic shock, ultraviolet light, and cytokines.[24,28] Activation of MAPKs occurs by phosphorylation on both tyrosine and threonine residues in the Thr–X–Tyr sequence by a family of dual-specific MAP kinase kinases (MKKs or MEKs), which are in turn phosphorylated and activated by a MAPK kinase kinase (MKKK or MEKK).[29,30] Once activated, MAPKs further phosphorylate several substrates such as ELK-1, c-Jun, ATF2, and CREB,[29,31] which in turn can regulate gene expression critical for several macrophage activities such as cell growth and differentiation, inflammatory responses, and polarization.
MITOGEN-ACTIVATED PROTEIN KINASE PATHWAYS IN M1–M2 POLARIZATION
As mentioned above, macrophages are functionally heterogeneous cells because they are affected by several signals in the TME, including cytokines, chemokines, and growth factors. Understanding the possible molecular mechanism responsible for macrophage polarization opens a new potential therapeutic area for cancer. The evidence that MAPK cascades might be implicated in the modulation of macrophage polarization has been illustrated recently. The MAPK/ERK pathway has shown to be involved in M2 TAMs’ repolarization. Chakraborty et al. showed that copper N-(2-hydroxy acetophenone) glycinate repolarizes M2-TAMs toward the inhibitory M1 phenotype via p38 MAPK and ERK1/2 activation as well as intracellular glutathione upregulation. Activation of p38 MAPK enhances IL-12 production, and ERK1/2 increases IFN-γ generation and, most importantly, downregulates TGFβ.[32] On the other hand, the p38/MAPKAP kinase 2 (MK2) has been shown to promote tumorigenesis in a murine model of colitis-associated cancer. Azoxymethane/dextran sodium sulphate-treated MK2 KO mice showed a marked decrease in macrophage infiltration in colon tissues compared with wild-type (WT) mice.[33] In a comparable work, MK2 knockout (KO) mice showed significantly smaller tumors and reduced total tumor burden when compared to WT mice. Specifically, MK2 depletion impaired the recruitment of arginase-1-positive M2 macrophages, whereas inducible nitric oxide synthase (iNOS)-positive M1 macrophages were enhanced in MK2 KO mouse tumors compared to WT counterparts. Upon IL-4 stimulation to induce an M2 phenotype, expression of CD206 and IL-10, common M2 markers were significantly lower in bone marrow macrophages derived from KO mice, suggesting the role of MK2 in M2 polarization.[34] Furthermore, Zhang et al. reported a critical role of ROS in ERK activation and the differentiation of monocytes toward M2 TAMs. Inhibition of ROS generation by butylated hydroxyanisole (BHA) specifically altered macrophage polarization to M2, but not M1 TAMs. BHA administration blocked ERK activation and furthered the expression of M2 macrophage marker CD163, IL-10, and chemokines CCL17 and CCL18.[35] In another study, blocking ERK5 reprogramed macrophages toward tumourcidal M1 phenotypes, evidenced by upregulation of M1 markers (e.g., iNos and Il12 β) and downregulated Tgfβ, Il10, and VEGFA messenger RNA expression. The same work reported that the growth of carcinoma grafts was inhibited in ERK5-deficient mice.[36]
The above-mentioned observations, summarized in Table 1, underline the importance of the MAPK pathway in M2 polarization and tumor progression. It is clear that interfering with this critical axis reprograms macrophages toward inhibitory M1 TAMs, favoring the production of pro-inflammatory mediators and the reduction in M2 markers. Interestingly, these studies indicated that targeting MAPK not only controlled M2 TAM density, but also led to the progression of tumors using different mice models.
Table 1.
Studies that targeted mitogen-activated protein kinase pathway in tumor-associated macrophage polarization
| Mechanism | Action | Effect on cytokines | References |
|---|---|---|---|
| Activation of p38 and ERK | Promotes repolarization of M2-TAMs towards M1 TAMs | 1 - Enhances IL12 and IFN-γ generation 2 - Downregulates TGFβ |
[32] |
| Inhibition of p38/MK2 | Impairs M2 polarization and M2 macrophage-induced angiogenesis | Decreases the expression of CD206, arginase-1 and IL-10 | [34] |
| Inhibition of ERK | Impairs the differentiation of monocytes to M2 TAMs | Decreases the expression of CD163, IL-10, and chemokines CCL17 and CCL18 | [35] |
| Blocking ERK5 | Promotes repolarization of M2-TAMs towards M1 TAMs | 1 - Upregulates iNos and IL12β 2 - Downregulates TGFβ, IL10 and VEGFA mRNA expression |
[36] |
MAPK: Mitogen-activated protein kinase, TAMs: Tumour-associated macrophages, ERK: Extracellular signal-regulated kinase, MK2: MAPKAP kinase 2, IL: Interleukin, IFN: Interferon, TGF: Transforming growth factor, VEGFA: Vascular endothelial growth factor A, CCL: CC chemokine ligand, iNOS: Inducible nitric oxide synthase, mRNA: Messenger RNA
CONCLUSION
The notion of the importance of the MAPK cascade with respect to the inhibition of ERK1/2 and p38 could clarify the molecular basis of TAMs polarization. Further studies regarding the role of MAPK phosphatases, for example, the negative regulator of MAPK members, will undoubtedly answer questions regarding TAMs’ differentiation and polarization. With the previous attempts to dissect the mechanism by which MAPK regulate TAMs’ functions, it is expected that, in the future, works will be expanded to produce selective MAPK signaling modifiers that can benefit cancer treatment.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
REFERENCES
- 1.Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19:1423–37. doi: 10.1038/nm.3394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wang M, Zhao J, Zhang L, Wei F, Lian Y, Wu Y, et al. Role of tumor microenvironment in tumorigenesis. J Cancer. 2017;8:761–73. doi: 10.7150/jca.17648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420:860–7. doi: 10.1038/nature01322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hanahan D, Weinberg RA. Hallmarks of cancer: The next generation. Cell. 2011;144:646–74. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 5.Hainaut P, Plymoth A. Targeting the hallmarks of cancer: Towards a rational approach to next-generation cancer therapy. Curr Opin Oncol. 2013;25:50–1. doi: 10.1097/CCO.0b013e32835b651e. [DOI] [PubMed] [Google Scholar]
- 6.Yang L, Zhang Y. Tumor-associated macrophages: From basic research to clinical application. J Hematol Oncol. 2017;10:58. doi: 10.1186/s13045-017-0430-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Olson OC, Kim H, Quail DF, Foley EA, Joyce JA. Tumor-associated macrophages suppress the cytotoxic activity of antimitotic agents. Cell Rep. 2017;19:101–13. doi: 10.1016/j.celrep.2017.03.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ngambenjawong C, Gustafson HH, Pun SH. Progress in tumor-associated macrophage (TAM)-targeted therapeutics. Adv Drug Deliv Rev. 2017;114:206–21. doi: 10.1016/j.addr.2017.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yuan R, Li S, Geng H, Wang X, Guan Q, Li X, et al. Reversing the polarization of tumor-associated macrophages inhibits tumor metastasis. Int Immunopharmacol. 2017;49:30–7. doi: 10.1016/j.intimp.2017.05.014. [DOI] [PubMed] [Google Scholar]
- 10.Müller-Quernheim UC, Potthast L, Müller-Quernheim J, Zissel G. Tumor-cell co-culture induced alternative activation of macrophages is modulated by interferons in vitro. J Interferon Cytokine Res. 2012;32:169–77. doi: 10.1089/jir.2011.0020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pixley FJ, Stanley ER. CSF-1 regulation of the wandering macrophage: Complexity in action. Trends Cell Biol. 2004;14:628–38. doi: 10.1016/j.tcb.2004.09.016. [DOI] [PubMed] [Google Scholar]
- 12.Rao KM. MAP kinase activation in macrophages. J Leukoc Biol. 2001;69:3–10. [PubMed] [Google Scholar]
- 13.Kim HS, Asmis R. Mitogen-activated protein kinase phosphatase 1 (MKP-1) in macrophage biology and cardiovascular disease. A redox-regulated master controller of monocyte function and macrophage phenotype. Free Radic Biol Med. 2017;109:75–83. doi: 10.1016/j.freeradbiomed.2017.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chi H, Barry SP, Roth RJ, Wu JJ, Jones EA, Bennett AM, et al. Dynamic regulation of pro- and anti-inflammatory cytokines by MAPK phosphatase 1 (MKP-1) in innate immune responses. Proc Natl Acad Sci U S A. 2006;103:2274–9. doi: 10.1073/pnas.0510965103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lim MX, Png CW, Tay CY, Teo JD, Jiao H, Lehming N, et al. Differential regulation of proinflammatory cytokine expression by mitogen-activated protein kinases in macrophages in response to intestinal parasite infection. Infect Immun. 2014;82:4789–801. doi: 10.1128/IAI.02279-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Raza A, Crothers JW, McGill MM, Mawe GM, Teuscher C, Krementsov DN, et al. Anti-inflammatory roles of p38α MAPK in macrophages are context dependent and require IL-10. J Leukoc Biol. 2017;102:1219–27. doi: 10.1189/jlb.2AB0116-009RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.De I, Steffen MD, Clark PA, Patros CJ, Sokn E, Bishop SM, et al. CSF1 overexpression promotes high-grade glioma formation without impacting the polarization status of glioma-associated microglia and macrophages. Cancer Res. 2016;76:2552–60. doi: 10.1158/0008-5472.CAN-15-2386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Riabov V, Gudima A, Wang N, Mickley A, Orekhov A, Kzhyshkowska J, et al. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front Physiol. 2014;5:75. doi: 10.3389/fphys.2014.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zheng X, Turkowski K, Mora J, Brüne B, Seeger W, Weigert A, et al. Redirecting tumor-associated macrophages to become tumoricidal effectors as a novel strategy for cancer therapy. Oncotarget. 2017;8:48436–52. doi: 10.18632/oncotarget.17061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Weagel E, Smith C, Liu PG, Robison R, O’Neill K. Macrophage polarization and its role in cancer. J Clin Cell Immunol. 2015;6:338. [Google Scholar]
- 21.Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, Goerdt S, et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity. 2014;41:14–20. doi: 10.1016/j.immuni.2014.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Franklin RA, Liao W, Sarkar A, Kim MV, Bivona MR, Liu K, et al. The cellular and molecular origin of tumor-associated macrophages. Science. 2014;344:921–5. doi: 10.1126/science.1252510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wyckoff JB, Wang Y, Lin EY, Li JF, Goswami S, Stanley ER, et al. Direct visualization of macrophage-assisted tumor cell intravasation in mammary tumors. Cancer Res. 2007;67:2649–56. doi: 10.1158/0008-5472.CAN-06-1823. [DOI] [PubMed] [Google Scholar]
- 24.Kyriakis JM, Avruch J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol Rev. 2001;81:807–69. doi: 10.1152/physrev.2001.81.2.807. [DOI] [PubMed] [Google Scholar]
- 25.Cargnello M, Roux PP. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol Mol Biol Rev. 2011;75:50–83. doi: 10.1128/MMBR.00031-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.McCain J. The MAPK (ERK) pathway: Investigational combinations for the treatment of BRAF-mutated metastatic melanoma. PT. 2013;38:96–108. [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002;12:9–18. doi: 10.1038/sj.cr.7290105. [DOI] [PubMed] [Google Scholar]
- 28.Torii S, Nakayama K, Yamamoto T, Nishida E. Regulatory mechanisms and function of ERK MAP kinases. J Biochem. 2004;136:557–61. doi: 10.1093/jb/mvh159. [DOI] [PubMed] [Google Scholar]
- 29.Turjanski AG, Vaqué JP, Gutkind JS. MAP kinases and the control of nuclear events. Oncogene. 2007;26:3240–53. doi: 10.1038/sj.onc.1210415. [DOI] [PubMed] [Google Scholar]
- 30.Chang L, Karin M. Mammalian MAP kinase signalling cascades. Nature. 2001;410:37–40. doi: 10.1038/35065000. [DOI] [PubMed] [Google Scholar]
- 31.Widmann C, Gibson S, Jarpe MB, Johnson GL. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol Rev. 1999;79:143–80. doi: 10.1152/physrev.1999.79.1.143. [DOI] [PubMed] [Google Scholar]
- 32.Chakraborty P, Chatterjee S, Ganguly A, Saha P, Adhikary A, Das T, et al. Reprogramming of TAM toward proimmunogenic type through regulation of MAP kinases using a redox-active copper chelate. J Leukoc Biol. 2012;91:609–19. doi: 10.1189/jlb.0611287. [DOI] [PubMed] [Google Scholar]
- 33.Ray AL, Castillo EF, Morris KT, Nofchissey RA, Weston LL, Samedi VG, et al. Blockade of MK2 is protective in inflammation-associated colorectal cancer development. Int J Cancer. 2016;138:770–5. doi: 10.1002/ijc.29716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Suarez-Lopez L, Sriram G, Kong YW, Morandell S, Merrick KA, Hernandez Y, et al. MK2 contributes to tumor progression by promoting M2 macrophage polarization and tumor angiogenesis. Proc Natl Acad Sci U S A. 2018;115:E4236–E4244. doi: 10.1073/pnas.1722020115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang Y, Choksi S, Chen K, Pobezinskaya Y, Linnoila I, Liu ZG, et al. ROS play a critical role in the differentiation of alternatively activated macrophages and the occurrence of tumor-associated macrophages. Cell Res. 2013;23:898–914. doi: 10.1038/cr.2013.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Giurisato E, Xu Q, Lonardi S, Telfer B, Russo I, Pearson A, et al. Myeloid ERK5 deficiency suppresses tumor growth by blocking protumor macrophage polarization via STAT3 inhibition. Proc Natl Acad Sci U S A. 2018;115:E2801–10. doi: 10.1073/pnas.1707929115. [DOI] [PMC free article] [PubMed] [Google Scholar]