

ABSTRACT
This review analyzes the relationship between microvesicles and reactive oxygen species (ROS). This relationship is bidirectional; on the one hand, the number and content of microvesicles produced by the cells are affected by oxidative stress conditions; on the other hand, microvesicles can directly and/or indirectly modify the ROS content in the extra- as well as the intracellular compartments. In this regard, microvesicles contain a pro-oxidant or antioxidant machinery that may produce or scavenge ROS: direct effect. This mechanism is especially suitable for eliminating ROS in the extracellular compartment. Endothelial microvesicles, in particular, contain a specific and well-developed antioxidant machinery. On the other hand, the molecules included in microvesicles can modify (activate or inhibit) ROS metabolism in their target cells: indirect effect. This can be achieved by the incorporation into the cells of ROS metabolic enzymes included in the microvesicles, or by the regulation of signaling pathways involved in ROS metabolism. Proteins, as well as miRNAs, are involved in this last effect.
KEYWORDS: Extracellular vesicles, microvesicles, reactive oxygen species
Microvesicles (MVs)
Extracellular vesicles are membranous subcellular structures produced by the cells; they are located in the extracellular space and are especially abundant in blood, urine, milk, saliva, semen, synovial or cerebrospinal fluid, among other liquids. According to their origin, size and biochemical composition, they are usually classified into three categories: exosomes, microvesicles (MVs) (also microparticles or ectosomes) and apoptotic bodies. The exosomes are 40–120 nm vesicles included in multi-vesicular bodies which are released to the extracellular space after fusion of these multi-vesicular bodies with the plasma membrane. MVs arise through budding and fission of the plasma membrane and are larger (50–1000 nm) than the exosomes [1]. Both types of vesicles are produced by all cells and have different functions [2]; however, it is not always easy to distinguish them [3,4]. Apoptotic bodies are the largest extracellular vesicles (1–5 um) and are formed during the late stages of apoptosis [1].
MVs represent an extraordinarily heterogeneous population of extracellular vesicles [5]. Not only are they heterogeneous in size (ranging from 0.1 to 1.5 μm) but also in their origin, biochemical content and, obviously, function. Blood MVs, in particular, are a paradigm of this heterogeneity; they can be secreted by erythrocytes, leucocytes, platelets or endothelial cells, each one with a different content and function [6]. Obtaining samples containing homogeneous populations of MVs is one of the main methodological challenges for the future.
The main function ascribed to MVs is a role in intercellular communication; however, many specific functions have also been associated with them depending on their cellular origin. Cell adhesion and migration [7,8], waste management [9], vascular function [10], coagulation [11], reticulocyte maturation [12], modulation of the immune response [13], fertilization [14], embryonic development [15], bone calcification [16] and tissue repair [17,18] are activities where MVs, as well as exosomes, have been involved. Moreover, an increase of MV production has been associated with pathological states [19,20], tumor growth, metastasis and angiogenesis [21,22]. They have also been postulated to be biomarkers and/or therapeutic targets [23], especially those derived from mesenchymal stem cells [24]. In fact, a significant therapeutic effect of mesenchymal stem cell-derived extracellular vesicles is the reduction of oxidative stress [25].
Oxidative stress and reactive oxygen species (ROS)
Under normal conditions, ROS production and ROS elimination are balanced; however, oxidative stress represents an imbalance due to an increase of ROS [26]. Thus, ROS production and ROS detoxification must be continuously adapted in order to respond to changes and alterations that occur during the cell´s lifespan. The ROS increase has been associated with stress conditions and has been shown to be the causal agent of different pathologies such as neurodegenerative disorders (Alzheimer´s disease, Parkinson´s disease and amyotrophic lateral sclerosis) [27,28], cardiovascular diseases [29,30] and carcinogenesis [31]. In general, ROS production increases with ageing [32], although it has also been proposed to be an adaptive response [33] that may be physiologically modified by age, sex (menopause), tumour growth or stress conditions.
ROS are a group of molecules and free radicals (atoms or molecules with an unpaired valence electron) derived from oxygen. They can be of exogenous (xenobiotics, radiation, pollutants …) or endogenous origin. The latter is a result of the cell’s own metabolism: mitochondrial respiration (especially ubiquinone and complex III of the electron transport chain) [34,35], oxidoreductase activities, metal-catalyzed oxidation and NADPH oxidase activity, especially in the respiratory burst of phagocytes [36].
ROS have numerous toxic and harmful effects due to their high reactivity. They are involved in different pathologies and, hence, cells have developed a complete antioxidant machinery for ROS scavenging (Figure 1). This antioxidant system includes enzymatic components, such as superoxide dismutases, catalase, peroxidases, reductases, and non-enzymatic components, such as vitamins C, E and A, glutathione, peroxiredoxins, thioredoxins, etc. It also includes enzymatic activities such as heme-oxygenase or catechol-O-methyltransferase directed at preventing ROS formation. Because the primary source of endogenous ROS is the intracellular compartment (mitochondria, peroxisomes and reticulum), cells have a stronger antioxidant machinery than the extracellular compartment.
Figure 1.
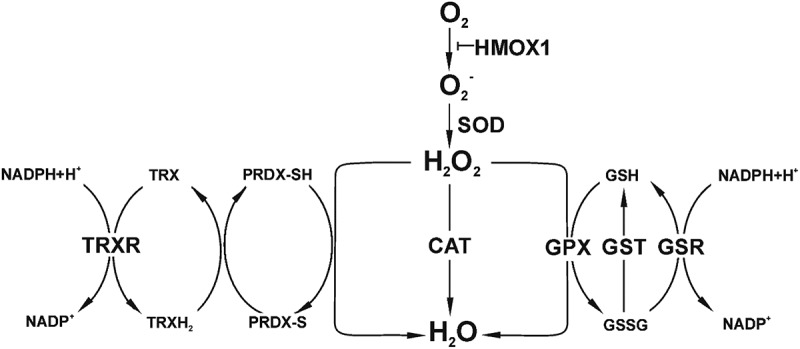
Diagram showing the two main ROS (superoxide radical, O2−, and hydrogen peroxide, H2O2) and the more essential enzymes and peptides of the antioxidant machinery. CAT: catalase; GPX: glutathione peroxidase; GSH and GSSH: reduced and oxidized glutathione; GSR: glutathione reductase; GST: glutathione S-transferase (detoxify xenobiotics); HMOX: heme oxygenase; PRDX: peroxiredoxin; SOD: superoxide dismutase; TRX: thioredoxin; TRXR: thioredoxin reductase. Other important ROS not included in the diagram are: hydroxyl (OH−), peroxyl (RO2−), alkoxyl (RO−) and hydroperoxyl (HO2−) radicals, as well as ozone (O3) and singlet oxygen (1O2) as non-radical ROS. Note that the fuel that nourishes the antioxidant machinery is NADPH.
ROS have two faces: good and bad. The first involves low ROS levels whereas the second involves high ROS levels [37]. It is well known that ROS are associated with a high number of different pathologies (previous paragraph) because they cause damage to biomolecules (lipids, proteins and DNA) and subcellular structures; however, ROS are also involved in maintaining physiological conditions [38,39]. Reactive nitrogen species (RNS) such as nitric oxide and nitrogen dioxide, among others, can also cause oxidative damage and act as components of intracellular signaling cascades [40]. In fact, the possibility that MVs act as scavengers of nitric oxide has also been proposed [41]. Nonetheless, the relationship between MVs and RNS is not considered in this review.
Under physiological conditions, ROS are involved in cell signaling, regulating a wide variety of functions [39]. These include activation of gene transcription (directly activating transcription factors, or indirectly activating MAPK cascades) [42,43], regulation of intracellular signaling pathways [44], modulation of calcium signaling [45], apoptosis [46,47], autophagy [48,49], cellular growth [50], and embryonic development as a consequence of their role in proliferation, differentiation, and apoptosis [51]. ROS are also involved in the destruction of pathogens [9] and in inflammatory processes [52,53], blood pressure control [54] and response to physical exercise [55].
ROS are involved in a high number of essential physiological processes, but while low ROS levels are related with physiological conditions, high ROS levels and the subsequent oxidative stress are generally associated with pathological conditions [56]. Therefore, ROS levels have to be finely regulated in the intracellular as well as the extracellular compartment. The biochemical, physiological and structural differences of both compartments make it impossible for them to have identical or similar mechanisms for regulating these levels. The differential distribution and expression of SOD subtypes in these two compartments is a good example of this. SOD1 is located in the cytosol, SOD2 in mitochondria and SOD3 in the extracellular compartment. To maintain proper ROS levels, the activity of these enzymes has to be finely regulated, as can be demonstrated by the fact that a single amino acid substitution in human SOD1 is associated with familial amyotrophic lateral sclerosis whereas the knockout of the SOD2 gene is related to lethal cardiomyopathy in mice [57]. The loss of SOD3 expression has also been associated with a pancreatic ductal adenocarcinoma [58]. This differential system of ROS elimination among intra- and extracellular compartments raises a special interest in analyzing the relationship between extracellular vesicles and the adjustment of ROS levels in the extracellular space.
Microvesicles and ROS
MVs and ROS are closely interrelated, not only because MVs can produce or detoxify ROS, but also because ROS are involved in the production of MVs. Pro-oxidant conditions seem to induce extracellular vesicle release [59]; in fact, NADPH oxidase and nitric oxide synthase-2 (NOS-2) inhibitors inhibit the production of MVs in neutrophils [60]. It is worthy to point out that tumoural [61] and senescent cells [62] also produce a higher number of MVs, and both types have altered redox balances with elevated ROS levels [63,64]). MVs can also serve as an alternative mechanism to remove oxidized proteins after oxidative stress, enabling their use as biomarkers for oxidative stress [65].
The effect of MVs on ROS depends on both the conditions of the cell that originates these vesicles and those of the target cell, as well as the environmental conditions. In fact, oxidative conditions affect the content of MVs [65]; for example, MVs of ischemic muscle [66] or those produced after a high-fat diet [67] produce more ROS than controls. In addition, endothelial-derived MVs obtained after starvation or apoptotic stress have different effects on endothelial cells after hypoxic stress: the former show beneficial effects whereas the latter exhibit detrimental effects [68]. In endothelial cells, specifically human umbilical vein endothelial cells (HUVEC), high glucose conditions have been shown to induce a three-fold increase in MV production, with differences in the molecular composition of these vesicles [69], whereas neutrophils produce diverse MVs in response to different activators [70]. Moreover, direct and indirect effects can be carried out by the same MVs: T-lymphocyte-derived MVs exert a potentially beneficial effect on HUVEC, acting both as ROS scavengers (they carry SOD2 and catalase) and inducing the expression of SOD-1 in these cells [71].
As far as we know, the first evidence that MVs could be involved in ROS metabolism was the location of Cu, Zn superoxide dismutase (SOD2), a cytosolic enzyme that destroys ROS, in MVs from neuroblastoma [72]. On the other hand, tumor-derived MVs and lymphocyte-derived MVs induced the production of ROS in human monocytes [73] and endothelial cells [74]. These early studies already pointed out the different roles and mechanistic procedures that MVs can maintain with ROS: MVs can directly scavenge or produce ROS [41] but they can also act on ROS indirectly, modifying the ROS content of their target cells (Figure 2).
Figure 2.
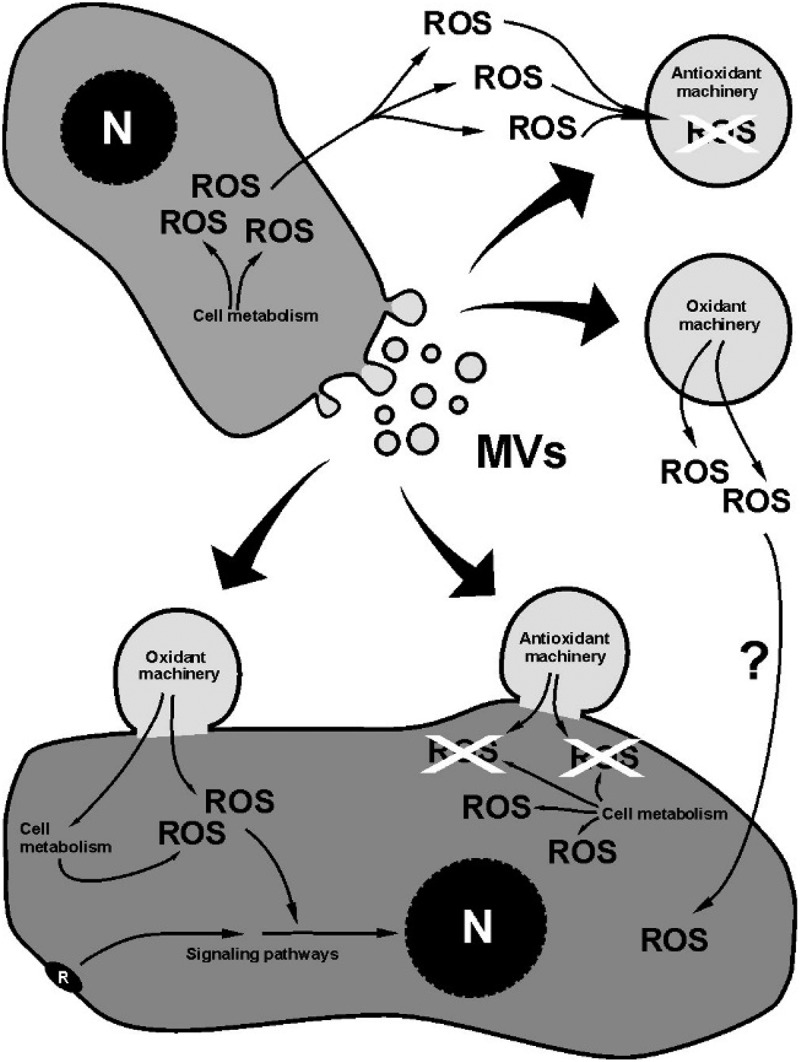
Schematic drawing showing the main effects of MVs on ROS. At the top, the direct effect of MVs; at the bottom, the indirect (cell-mediated) effect. The effect of ROS- regulated signaling pathways on the ROS content of the cell is not included.
MVs can carry different antioxidant enzymes involved in ROS scavenging: GPX, GST, PRDX, SOD2 or CAT [71,75–78]. Our group recently found a complete list of antioxidant activities and related molecules in MVs derived from HUVEC by proteomic analysis [79]. The fuel that nourishes the antioxidant machinery is NADPH. We also demonstrated that HUVEC-derived MVs contain the enzymatic machinery necessary to synthesize NADPH using blood metabolites to feed different biosynthetic pathways [80]. This last possibility seems to convert the HUVEC-derived MVs into an autonomous extracellular organelle devoted to scavenging ROS from blood and maintaining the redox status in plasma. A protective role of epididymosomes (a type of MV originated from epididymal cells) for epididymal spermatozoa against ROS released by dying cells has also been suggested [81]. On the other hand, the existence of NADPH oxidase, an enzyme that synthesizes ROS, has also been demonstrated in MVs [66,82]. Taking all this into account, it is possible to assume that a group of MVs can act as ROS scavengers in the extracellular compartment, and others as ROS producers when their content is incorporated into the target cells. As far as we know, the possibility that MVs can directly increase ROS in the extracellular compartment has not been demonstrated, although their ability to produce ROS has been assessed [82].
Although the direct effect of MVs on ROS has been analyzed in the previous paragraph, MVs, under physiological conditions, can also have an indirect effect, inducing changes in their target cells that can result in a compensatory mechanism against the effects of oxidative stress or causing detrimental effects [65]. This indirect effect can be achieved by (a) the incorporation to the cells of the enzymatic components included in MVs involved in ROS production/destruction, or (b) the incorporation of signaling molecules that can modify cellular activities and/or gene expression involved in the regulation of redox processes, which finally affect the cellular ROS content. The induction of antioxidant enzyme expression [71,83] and the ROS increase [74,82,84] have been demonstrated. The precise molecular mechanisms activated by the MVs in the target cells are not well known. However, NF-κB, JNK or PI3K/Akt-dependent signaling pathways are usually involved in ROS production in the target cells [68,74,84] whereas suppression of NOX expression [85] or inhibition of NF-κB [86] are molecular mechanisms used to decrease ROS. A common response to oxidative stress involves two transcription factors, Nrf2 and MAFG, as well as the activation of target genes via antioxidant response elements (AREs) [87]. As far as we know, however, these transcription factors have not been found in MVs, although they can be included in exosomes (see last paragraph). MicroRNAs (miRNAs) are molecules involved in the control of oxidative stress [37,88–91] and MVs represent transport vehicles for these [92]. In particular, miR-126, as well as miR-21 [93,94], miR-128 [95], miR-144 [94], miR-34a-5p [96], miR-1915-3p [97] and miR-638 [98], are all involved in oxidative stress and have been found in MVs.
Similar results have been reported for MVs derived from cells subjected to stress or under pathological conditions. MVs derived from ischemic muscles (56), plasma of patients with lupus [99] or neutrophils infected with Mycobacterium tuberculosis [70] were shown to induce ROS in mononuclear cells, neutrophils and macrophages, respectively. However, MVs derived from cells exposed to ROS induced ROS tolerance in PC12 cells [100]. MVs derived from tumour cells showed a similar behavior; they can contain antioxidant enzymes [72] or can modulate the activity of human monocytes by increasing ROS, among other effects [73].
Although MVs produced after a hypoxia/reoxygenation treatment contain ROS and may promote apoptosis and oxidative stress in the myocardium [101], the use of MVs as therapeutic tools has been demonstrated.Treatments with MVs reduced oxidative stress in injured kidneys [85,102] and in experimental colitis [86]. In addition, it has been recently demonstrated that MVs derived from mesenchymal cells downregulate oxidative stress in osteoarthritic chondrocytes [103] and that those secreted by genistein (a polyphenol)-treated cells have a protective effect against oxidative stress [104]. On the contrary, in cultured glomerular endothelial cells, platelet microparticles have been shown to induce ROS production and may contribute to glomerular endothelial injury associated with diabetic nephropathy [104].
A question of interest is whether MVs having an antagonistic relationship with ROS (scavenging or production) may (1) coexist in the same place and (2) be produced by the same cell. Plasma MVs can be a paradigm to respond to these questions. We have demonstrated that cultured endothelial cells synthesize MVs that act as ROS scavengers [79,80], but it has also been suggested that MVs produce ROS as part of the signaling processes in endothelial cells [82]. This raises the possibility that MVs involved in ROS scavenging and ROS production may be found simultaneously in plasma, the first ones acting as autonomous structures independent of target cells and the second ones acting on target cells; obviously, both types of MVs should have specific mechanisms of cargo.
The underlying molecular mechanisms of aging appear to be related to increased free radical release [105]; senescence has been equally associated with an increase of oxidative stress. Two possible mechanisms may lead to this oxidative status: a malfunctioning of the antioxidant machinery or an increase of oxidative processes by metabolic alterations of the cells. As previously stated (section 3, first paragraph), senescent cells produce more MVs probably due to their high ROS levels. Senescent endothelial cell-derived MVs exhibit an inductive effect of ROS on endothelial cells [62,106]. However, we have recently demonstrated that senescent HUVEC-derived MVs have a functional and more developed antioxidant machinery, suggesting that the increase of the antioxidant machinery is not able to compensate the higher production of ROS in senescence [79]. In this context, a recent study has demonstrated that the culture time induces changes in microRNAs related to genes involved in ROS production [107]. On the other hand, MVs could modify the senescence status, a condition with high oxidative stress. In this regard, MVs of interleukin-1β-stimulated mesenchymal stem cells have been demonstrated to downregulate β-galactosidase activity, a marker of senescence [108].
In spite of their possible separate origin and functions, MVs and exosomes can have common functional mechanisms; for example, exosomes have also been involved in ROS metabolism [109]. However, the actual multivesicular body origin of the small EVs is not clearly demonstrated, and the functions described may be contained in exosomes and/or in co-isolated small MVs [3]. Thus, although this review is mainly directed at studying the interrelationship between MVs and ROS, this final paragraph is dedicated to the relationship between exosomes (and co-isolated small MVs) and ROS. Enzymes such as NADPH oxidase, involved in ROS production, have been found in platelet-derived exosomes [110]. In addition, exosomes carry cytochrome P450 [111], a protein family involved in ROS generation [112]. In eosinophils, an increase in ROS production by eosinophil-derived exosomes has been demonstrated in patients with asthma [113]. Production of ROS in axonal regeneration induced by macrophage-derived exosomes containing NADPH oxidase has also been recently demonstrated [114]; in this case, the ROS generated by NADPH oxidase served as an activator of the Akt-dependent signaling pathway involved in regeneration. In the scientific literature, however, it is easier to find studies pointing towards an antioxidant role of exosomes, although this antioxidant role can be carried out by different mechanisms. Cells treated with exosomes derived from stem cells ameliorate their oxidative stress [115] or reduce ROS production [116]. Several studies have demonstrated a protective function of exosomes against oxidative stress [117,118] as well as an increase of antioxidants in exosomes derived from cells treated with ROS, and the subsequent induction of antioxidant mechanisms in cells treated with these exosomes [119,120]. The presence of Nrf2 mRNA, a transcription factor involved in the anti-oxidant stress response, and miRNAs involved in the oxidative stress response has also been demonstrated in exosomes from granulose cells subjected to hydrogen peroxide [121]. In addition, exosomes can diminish cellular oxidative stress by secreting harmful molecules that can promote ROS elevation and induce senescence, as has been demonstrated in the case of nuclear DNA accumulation in the cytoplasm [122]. The interrelationship of ROS and exosomes is phylogenetically preserved; the existence of antioxidant molecules (SOD2, TRX, TRXR and catalase) has also been demonstrated in exosomes from the yeast, Cryptococcus neoformans [123].
Conclusions
MVs, or at least a group of them, maintain a close interrelationship with ROS and, subsequently, with oxidative stress. This interrelationship can be described as either direct or indirect. MVs, by themselves (direct effect), can act as ROS scavengers, reducing oxidative stress, since they carry antioxidant enzymes and molecules that form part of the cellular antioxidant machinery as well as enzymes involved in ROS production. After interacting with their target cells (indirect effect or horizontal transfer), MVs can also increase or reduce ROS levels by transferring enzymatic components or signaling molecules that can modify cell metabolism and/or gene expression involved in the regulation of redox processes. This direct and indirect capacity of ROS production or scavenging by the MVs can be affected by the physiological conditions of the cell that produces the MVs. Stress or pathological conditions, as well as aging or senescence, also modify the effect of MVs on ROS metabolism. Since oxidative stress is involved in different pathological conditions and aging, the use of MVs with antioxidant activity could be a useful strategy to prevent the deleterious effects of ROS. The adaptive response consisting of modifying the number and content of MVs related with ROS metabolism under these circumstances supports this possibility. Finally, the direct and indirect capacity of ROS production or scavenging implies the existence of different types of MVs. The mechanisms that regulate the synthesis of each type, their mechanisms of cargo and the mechanism that guides them to fuse with their target cells or to stay in the extracellular space are all interesting subjects for future explorations.
Funding Statement
This work was supported by the Plan Nacional Proyectos de Investigación en Salud of Instituto de Salud Carlos III (ISCIII) Fondos Feder European Grant (PI14/00806; PI17/01029; ISCIII-FEDER), P11-CTS7352, Santander/UCM PR41/17-20964, “Sociedad Española de Nefrologia”, and “Proyectos de Investigación del Programa Propio de la UAH”: UAH-GP2018-4 and CCG2018/BIO-010.
Disclosure statement
The authors report no potential conflict of interest.
References
- [1].Yáñez-Mó M, Siljander PR, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Antonyak MA, Cerione RA.. Emerging picture of the distinct traits and functions of microvesicles and exosomes. Proc Natl Acad Sci U S A. 2015March;112(12):3589–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–289. [DOI] [PubMed] [Google Scholar]
- [4].Meldolesi J. Ectosomes and exosomes-two extracellular vesicles that differ only in some details. Biochem Mol Biol. 2016;02. [Google Scholar]
- [5].Marcoux G, Duchez AC, Cloutier N, et al. Revealing the diversity of extracellular vesicles using high-dimensional flow cytometry analyses. Sci Rep. 2016;6:35928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Tissot J-D, Canellini G, Rubin O, et al. Blood microvesicles: from proteomics to physiology. Transl Proteomics. 2013;1(1):38–52. [Google Scholar]
- [7].Niu C, Wang X, Zhao M, et al. Macrophage foam cell-derived extracellular vesicles promote vascular smooth muscle cell migration and adhesion. J Am Heart Assoc. 2016October;5(10). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Lee H, Zhang D, Wu J, et al. Lung epithelial cell-derived microvesicles regulate macrophage migration via microRNA-17/221-induced integrin β1 recycling. J Immunol. 2017;199:1453–1464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Winterbourn CC, Kettle AJ, Hampton MB. Reactive oxygen species and neutrophil function. Annu Rev Biochem. 2016June;85:765–792. [DOI] [PubMed] [Google Scholar]
- [10].Jansen F, Li Q, Pfeifer A, et al. Endothelial- and immune cell-derived extracellular vesicles in the regulation of cardiovascular health and disease. JACC Basic Transl Sci. 2017December;2(6):790–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Koshiar RL, Somajo S, Norström E, et al. Erythrocyte-derived microparticles supporting activated protein C-mediated regulation of blood coagulation. PLoS One. 2014;9(8):e104200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Griffiths RE, Kupzig S, Cogan N, et al. The ins and outs of human reticulocyte maturation: autophagy and the endosome/exosome pathway. Autophagy. 2012July;8(7):1150–1151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol. 2014March;14(3):195–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Machtinger R, Laurent LC, Baccarelli AA. Extracellular vesicles: roles in gamete maturation, fertilization and embryo implantation. Hum Reprod Update. 2016Mar-Apr;22(2):182–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Desrochers LM, Bordeleau F, Reinhart-King CA, et al. Microvesicles provide a mechanism for intercellular communication by embryonic stem cells during embryo implantation. Nat Commun. 2016;7:11958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Alique M, Ruíz-Torres MP, Bodega G, et al. Microvesicles from the plasma of elderly subjects and from senescent endothelial cells promote vascular calcification. Aging (Albany NY). 2017March;9(3):778–789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Panagiotou N, Wayne Davies R, Selman C, et al. Microvesicles as vehicles for tissue regeneration: changing of the guards. Curr Pathobiol Rep. 2016;4(4):181–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Taverna S, Pucci M, Alessandro R. Extracellular vesicles: small bricks for tissue repair/regeneration. Ann Transl Med. 2017February;5(4):83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Yuana Y, Sturk A, Nieuwland R. Extracellular vesicles in physiological and pathological conditions. Blood Rev. 2013January;27(1):31–39. [DOI] [PubMed] [Google Scholar]
- [20].Surman M, Stępień E, Hoja-Łukowicz D, et al. Deciphering the role of ectosomes in cancer development and progression: focus on the proteome. Clin Exp Metastasis. 2017April;34(3–4):273–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Muralidharan-Chari V, Clancy JW, Sedgwick A, et al. Microvesicles: mediators of extracellular communication during cancer progression. J Cell Sci. 2010May;123(Pt 10):1603–1611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Minciacchi VR, Freeman MR, Di Vizio D. Extracellular vesicles in cancer: exosomes, microvesicles and the emerging role of large oncosomes. Semin Cell Dev Biol. 2015April;40:41–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Chen Y, Li G, Liu ML. Microvesicles as emerging biomarkers and therapeutic targets in cardiometabolic diseases. Genomics Proteomics Bioinformatics. 2018February;16(1):50–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Fujita Y, Kadota T, Araya J, et al. Clinical application of mesenchymal stem cell-derived extracellular vesicle-based therapeutics for inflammatory lung diseases. J Clin Med. 2018October;7(10):355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Keshtkar S, Azarpira N, Ghahremani MH. Mesenchymal stem cell-derived extracellular vesicles: novel frontiers in regenerative medicine. Stem Cell Res Ther. 2018March;9(1):63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Sies H. On the history of oxidative stress: concept and some aspects of current development. Curr Opin Toxicol. 2018;7:122–126. [Google Scholar]
- [27].Kim GH, Kim JE, Rhie SJ, et al. The role of oxidative stress in neurodegenerative diseases. Exp Neurobiol. 2015December;24(4):325–340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Liu Z, Zhou T, Ziegler AC, et al. Oxidative stress in neurodegenerative diseases: from molecular mechanisms to clinical applications. Oxid Med Cell Longev. 2017;2017:2525967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Sugamura K, Keaney JF. Reactive oxygen species in cardiovascular disease. Free Radic Biol Med. 2011September;51(5):978–992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Panth N, Paudel KR, Parajuli K. Reactive oxygen species: a key hallmark of cardiovascular disease. Adv Med. 2016;2016:9152732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Liou GY, Storz P. Reactive oxygen species in cancer. Free Radic Res. 2010May;44(5):479–496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Davalli P, Mitic T, Caporali A, et al. ROS, cell senescence, and novel molecular mechanisms in aging and age-related diseases. Oxid Med Cell Longev. 2016;2016:3565127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Liochev SI. Reactive oxygen species and the free radical theory of aging. Free Radic Biol Med. 2013July;60:1–4. [DOI] [PubMed] [Google Scholar]
- [34].Chen Q, Vazquez EJ, Moghaddas S, et al. Production of reactive oxygen species by mitochondria: central role of complex III. J Biol Chem. 2003September;278(38):36027–36031. [DOI] [PubMed] [Google Scholar]
- [35].Murphy MP. How mitochondria produce reactive oxygen species. Biochem J. 2009January;417(1):1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Robinson JM. Reactive oxygen species in phagocytic leukocytes. Histochem Cell Biol. 2008August;130(2):281–297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Banerjee J, Khanna S, Bhattacharya A. MicroRNA regulation of oxidative stress. Oxid Med Cell Longev. 2017;2017:2872156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Valko M, Leibfritz D, Moncol J, et al. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39(1):44–84. [DOI] [PubMed] [Google Scholar]
- [39].Di Meo S, Reed TT, Venditti P, et al. Role of ROS and RNS sources in physiological and pathological conditions. Oxid Med Cell Longev. 2016;2016:1245049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Weidinger A, Kozlov AV. Biological activities of reactive oxygen and nitrogen species: oxidative stress versus signal transduction. Biomolecules. 2015April;5(2):472–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Larson MC, Hillery CA, Hogg N. Circulating membrane-derived microvesicles in redox biology. Free Radic Biol Med. 2014August;73:214–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Turpaev KT. Reactive oxygen species and regulation of gene expression. Biochemistry (Mosc). 2002March;67(3):281–292. [DOI] [PubMed] [Google Scholar]
- [43].Mikhed Y, Görlach A, Knaus UG, et al. Redox regulation of genome stability by effects on gene expression, epigenetic pathways and DNA damage/repair. Redox Biol. 2015;5:275–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Finkel T. Signal transduction by reactive oxygen species. J Cell Biol. 2011July;194(1):7–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Görlach A, Bertram K, Hudecova S, et al. Calcium and ROS: a mutual interplay. Redox Biol. 2015;6:260–271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Circu ML, Aw TY. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic Biol Med. 2010March;48(6):749–762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Redza-Dutordoir M, Averill-Bates DA. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim Biophys Acta. 2016December;1863(12):2977–2992. [DOI] [PubMed] [Google Scholar]
- [48].Azad MB, Chen Y, Gibson SB. Regulation of autophagy by reactive oxygen species (ROS): implications for cancer progression and treatment. Antioxid Redox Signal. 2009April;11(4):777–790. [DOI] [PubMed] [Google Scholar]
- [49].Filomeni G, De Zio D, Cecconi F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ. 2015March;22(3):377–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Sauer H, Wartenberg M, Hescheler J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell Physiol Biochem. 2001;11(4):173–186. [DOI] [PubMed] [Google Scholar]
- [51].Dennery PA. Effects of oxidative stress on embryonic development. Birth Defects Res C Embryo Today. 2007September;81(3):155–162. [DOI] [PubMed] [Google Scholar]
- [52].Mittal M, Siddiqui MR, Tran K, et al. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal. 2014March;20(7):1126–1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Forrester SJ, Kikuchi DS, Hernandes MS, et al. Reactive oxygen species in metabolic and inflammatory signaling. Circ Res. 2018March;122(6):877–902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Lassègue B, Griendling KK. Reactive oxygen species in hypertension; an update. Am J Hypertens. 2004September;17(9):852–860. [DOI] [PubMed] [Google Scholar]
- [55].He F, Li J, Liu Z, et al. Redox mechanism of reactive oxygen species in exercise. Front Physiol. 2016;7:486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014May;24(10):R453–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Zelko IN, Mariani TJ, Folz RJ. Superoxide dismutase multigene family: a comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic Biol Med. 2002August;33(3):337–349. [DOI] [PubMed] [Google Scholar]
- [58].O’Leary BR, Fath MA, Bellizzi AM, et al. Loss of SOD3 (EcSOD) expression promotes an aggressive phenotype in human pancreatic ductal adenocarcinoma. Clin Cancer Res. 2015April;21(7):1741–1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Benedikter BJ, Weseler AR, Wouters EFM, et al. Redox-dependent thiol modifications: implications for the release of extracellular vesicles. Cell Mol Life Sci. 2018July;75(13):2321–2337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Thom SR, Bhopale VM, Yang M. Neutrophils generate microparticles during exposure to inert gases due to cytoskeletal oxidative stress. J Biol Chem. 2014July;289(27):18831–18845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Huber V, Fais S, Iero M, et al. Human colorectal cancer cells induce T-cell death through release of proapoptotic microvesicles: role in immune escape. Gastroenterology. 2005June;128(7):1796–1804. [DOI] [PubMed] [Google Scholar]
- [62].Burger D, Kwart DG, Montezano AC, et al. Microparticles induce cell cycle arrest through redox-sensitive processes in endothelial cells: implications in vascular senescence. J Am Heart Assoc. 2012June;1(3):e001842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Kong H, Chandel NS. Regulation of redox balance in cancer and T cells. J Biol Chem. 2018May;293(20):7499–7507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Chandrasekaran A, Idelchik MDPS, Melendez JA. Redox control of senescence and age-related disease. Redox Biol. 2017April;11:91–102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Yarana C, St Clair DK. Chemotherapy-induced tissue injury: an insight into the role of extracellular vesicles-mediated oxidative stress responses. Antioxidants (Basel). 2017September;6(4):75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Leroyer AS, Ebrahimian TG, Cochain C, et al. Microparticles from ischemic muscle promotes postnatal vasculogenesis. Circulation. 2009June;119(21):2808–2817. [DOI] [PubMed] [Google Scholar]
- [67].Heinrich LF, Andersen DK, Cleasby ME, et al. Long-term high fat feeding of rats results in increased numbers of circulating microvesicles with pro-inflammatory effects on endothelial cells. Br J Nutr. 2015June;113(11):1704–1711. [DOI] [PubMed] [Google Scholar]
- [68].Wang J, Chen S, Ma X, et al. Effects of endothelial progenitor cell-derived microvesicles on hypoxia/reoxygenation-induced endothelial dysfunction and apoptosis. Oxid Med Cell Longev. 2013;2013:572729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Burger D, Turner M, Xiao F, et al. High glucose increases the formation and pro-oxidative activity of endothelial microparticles. Diabetologia. 2017September;60(9):1791–1800. [DOI] [PubMed] [Google Scholar]
- [70].Alvarez-Jiménez VD, Leyva-Paredes K, García-Martínez M, et al. Extracellular vesicles released from Mycobacterium tuberculosis-infected neutrophils promote macrophage autophagy and decrease intracellular mycobacterial survival. Front Immunol. 2018;9:272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Soleti R, Lauret E, Andriantsitohaina R, et al. Internalization and induction of antioxidant messages by microvesicles contribute to the antiapoptotic effects on human endothelial cells. Free Radic Biol Med. 2012December;53(11):2159–2170. [DOI] [PubMed] [Google Scholar]
- [72].Mondola P, Ruggiero G, Serù R, et al. The Cu,Zn superoxide dismutase in neuroblastoma SK-N-BE cells is exported by a microvesicles dependent pathway. Brain Res Mol Brain Res. 2003January;110(1):45–51. [DOI] [PubMed] [Google Scholar]
- [73].Baj-Krzyworzeka M, Szatanek R, Weglarczyk K, et al. Tumour-derived microvesicles modulate biological activity of human monocytes. Immunol Lett. 2007November;113(2):76–82. [DOI] [PubMed] [Google Scholar]
- [74].Mostefai HA, Meziani F, Mastronardi ML, et al. Circulating microparticles from patients with septic shock exert protective role in vascular function. Am J Respir Crit Care Med. 2008December;178(11):1148–1155. [DOI] [PubMed] [Google Scholar]
- [75].Jin M, Drwal G, Bourgeois T, et al. Distinct proteome features of plasma microparticles. Proteomics. 2005May;5(7):1940–1952. [DOI] [PubMed] [Google Scholar]
- [76].Peterson DB, Sander T, Kaul S, et al. Comparative proteomic analysis of PAI-1 and TNF-alpha-derived endothelial microparticles. Proteomics. 2008June;8(12):2430–2446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Little KM, Smalley DM, Harthun NL, et al. The plasma microparticle proteome. Semin Thromb Hemost. 2010November;36(8):845–856. [DOI] [PubMed] [Google Scholar]
- [78].Povero D, Eguchi A, Li H, et al. Circulating extracellular vesicles with specific proteome and liver microRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS One. 2014;9(12):e113651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Bodega G, Alique M, Bohórquez L, et al. The antioxidant machinery of young and senescent human umbilical vein endothelial cells and their microvesicles. Oxid Med Cell Longev. 2017;2017:7094781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Bodega G, Alique M, Bohórquez L, et al. Young and especially senescent endothelial microvesicles produce nadph: the fuel for their antioxidant machinery. Oxid Med Cell Longev. 2018;2018:3183794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Sullivan R. Epididymosomes: a heterogeneous population of microvesicles with multiple functions in sperm maturation and storage. Asian J Androl. 2015Sep-Oct;17(5):726–729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Burger D, Turner M, Munkonda MN, et al. Endothelial microparticle-derived reactive oxygen species: role in endothelial signaling and vascular function. Oxid Med Cell Longev. 2016;2016:5047954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Mahmoud AM, Wilkinson FL, McCarthy EM, et al. Endothelial microparticles prevent lipid-induced endothelial damage via Akt/eNOS signaling and reduced oxidative stress. Faseb J. 2017October;31(10):4636–4648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Wang H, Wang ZH, Kong J, et al. Oxidized low-density lipoprotein-dependent platelet-derived microvesicles trigger procoagulant effects and amplify oxidative stress. Mol Med. 2012;18:159–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Zhang G, Zou X, Miao S, et al. The anti-oxidative role of micro-vesicles derived from human Wharton-Jelly mesenchymal stromal cells through NOX2/gp91(phox) suppression in alleviating renal ischemia-reperfusion injury in rats. PLoS One. 2014;9(3):e92129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Yang J, Liu XX, Fan H, et al. Extracellular vesicles derived from bone marrow mesenchymal stem cells protect against experimental colitis via attenuating colon inflammation, oxidative stress and apoptosis. PLoS One. 2015;10(10):e0140551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Katsuoka F, Motohashi H, Engel JD, et al. Nrf2 transcriptionally activates the mafG gene through an antioxidant response element. J Biol Chem. 2005February;280(6):4483–4490. [DOI] [PubMed] [Google Scholar]
- [88].Bu H, Wedel S, Cavinato M, et al. MicroRNA regulation of oxidative stress-induced cellular senescence. Oxid Med Cell Longev. 2017;2017:2398696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Cheleschi S, De Palma A, Pascarelli NA, et al. Could oxidative stress regulate the expression of microRNA-146a and microRNA-34a in human osteoarthritic chondrocyte cultures? Int J Mol Sci. 2017December;18(12). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Matsuzaki J, Ochiya T. Extracellular microRNAs and oxidative stress in liver injury: a systematic mini review. J Clin Biochem Nutr. 2018July;63(1):6–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Wang Z, Liu Y, Han N, et al. Profiles of oxidative stress-related microRNA and mRNA expression in auditory cells. Brain Res. 2010;1346:14–25. [DOI] [PubMed] [Google Scholar]
- [92].Diehl P, Fricke A, Sander L, et al. Microparticles: major transport vehicles for distinct microRNAs in circulation. Cardiovasc Res. 2012March;93(4):633–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Hulsmans M, Holvoet P. MicroRNA-containing microvesicles regulating inflammation in association with atherosclerotic disease. Cardiovasc Res. 2013October;100(1):7–18. [DOI] [PubMed] [Google Scholar]
- [94].Pu C, Huang H, Wang Z, et al. Extracellular vesicle-associated mir-21 and mir-144 are markedly elevated in serum of patients with hepatocellular carcinoma. Front Physiol. 2018;9:930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Bollati V, Angelici L, Rizzo G, et al. Microvesicle-associated microRNA expression is altered upon particulate matter exposure in healthy workers and in A549 cells. J Appl Toxicol. 2015January;35(1):59–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Rani A, O’Shea A, Ianov L, et al. miRNA in circulating microvesicles as biomarkers for age-related cognitive decline. Front Aging Neurosci. 2017;9:323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Carmona A, Guerrero F, Buendia P, et al. Microvesicles derived from indoxyl sulfate treated endothelial cells induce endothelial progenitor cells dysfunction. Front Physiol. 2017;8:666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Zhang J, Zhao A, Sun L, et al. Selective surface marker and miRNA profiles of CD34. Oncol Lett. 2017August;14(2):1866–1874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Winberg LK, Jacobsen S, Nielsen CH. Microparticles from patients with systemic lupus erythematosus induce production of reactive oxygen species and degranulation of polymorphonuclear leukocytes. Arthritis Res Ther. 2017October;19(1):230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Grindheim AK, Vedeler A. Extracellular vesicles released from cells exposed to reactive oxygen species increase annexin A2 expression and survival of target cells exposed to the same conditions. Commun Integr Biol. 2016Jul-Aug;9(4):e1191715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Zhang Q, Shang M, Zhang M, et al. Microvesicles derived from hypoxia/reoxygenation-treated human umbilical vein endothelial cells promote apoptosis and oxidative stress in H9c2 cardiomyocytes. BMC Cell Biol. 2016June;17(1):25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Zhang G, Zou X, Huang Y, et al. Mesenchymal stromal cell-derived extracellular vesicles protect against acute kidney injury through anti-oxidation by enhancing Nrf2/ARE activation in rats. Kidney Blood Press Res. 2016;41(2):119–128. [DOI] [PubMed] [Google Scholar]
- [103].Tofiño-Vian M, Pérez Del Caz MD, Silvestre A, et al. FRI0014 Antioxidant role of microvesicles from adipose tissue-derived mesenchymal stem cells in human osteoarthritic chondrocytes. Ann Rheumatic Dis. 2017;76:485. [Google Scholar]
- [104].Gimeno-Mallench L, Mas-Bargues C, Inglés M, et al. Antioxidant role of microvesicles, activated by genistein, in stem cells. Free Radic Biol Med. 2018;120 (suppl. 1):S83–S84. [Google Scholar]
- [105].Haigis MC, Yankner BA. The aging stress response. Mol Cell. 2010October;40(2):333–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Abbas M, Jesel L, Auger C, et al. Endothelial microparticles from acute coronary syndrome patients induce premature coronary artery endothelial cell aging and thrombogenicity: role of the Ang II/AT1 receptor/NADPH oxidase-mediated activation of MAPKs and PI3-kinase pathways. Circulation. 2017January;135(3):280–296. [DOI] [PubMed] [Google Scholar]
- [107].Lange-Consiglio A, Lazzari B, Pizzi F, et al. Different culture times affect microrna cargo in equine amniotic mesenchymal cells and their microvesicles. Tissue Eng Part C Methods. 2018October;24(10):596–604. [DOI] [PubMed] [Google Scholar]
- [108].Tofiño-Vian M, Guillén MI, Pérez Del Caz MD, et al. Extracellular vesicles from adipose-derived mesenchymal stem cells downregulate senescence features in osteoarthritic osteoblasts. Oxid Med Cell Longev. 2017;2017:7197598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Chettimada S, Lorenz DR, Misra V, et al. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci Rep. 2018May;8(1):7227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Janiszewski M, Do Carmo AO, Pedro MA, et al. Platelet-derived exosomes of septic individuals possess proapoptotic NAD(P)H oxidase activity: A novel vascular redox pathway. Crit Care Med. 2004March;32(3):818–825. [DOI] [PubMed] [Google Scholar]
- [111].Kumar S, Sinha N, Gerth KA, et al. Specific packaging and circulation of cytochromes P450, especially 2E1 isozyme, in human plasma exosomes and their implications in cellular communications. Biochem Biophys Res Commun. 2017September;491(3):675–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Caro AA, Cederbaum AI. Oxidative stress, toxicology, and pharmacology of CYP2E1. Annu Rev Pharmacol Toxicol. 2004;44:27–42. [DOI] [PubMed] [Google Scholar]
- [113].Cañas JA, Sastre B, Mazzeo C, et al. Exosomes from eosinophils autoregulate and promote eosinophil functions. J Leukoc Biol. 2017May;101(5):1191–1199. [DOI] [PubMed] [Google Scholar]
- [114].Hervera A, De Virgiliis F, Palmisano I, et al. Reactive oxygen species regulate axonal regeneration through the release of exosomal NADPH oxidase 2 complexes into injured axons. Nat Cell Biol. 2018March;20(3):307–319. [DOI] [PubMed] [Google Scholar]
- [115].Zhou Y, Xu H, Xu W, et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem Cell Res Ther. 2013April;4(2):34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Alcayaga-Miranda F, González PL, Lopez-Verrilli A, et al. Prostate tumor-induced angiogenesis is blocked by exosomes derived from menstrual stem cells through the inhibition of reactive oxygen species. Oncotarget. 2016July;7(28):44462–44477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Eldh M, Ekström K, Valadi H, et al. Exosomes communicate protective messages during oxidative stress; possible role of exosomal shuttle RNA. PLoS One. 2010December;5(12):e15353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Ezquer F, Quintanilla ME, Morales P, et al. Intranasal delivery of mesenchymal stem cell-derived exosomes reduces oxidative stress and markedly inhibits ethanol consumption and post-deprivation relapse drinking. Addict Biol. 2018Sep-Adb.12675 [Epub Ahead of Print]. [DOI] [PubMed] [Google Scholar]
- [119].Patel GK, Khan MA, Bhardwaj A, et al. Exosomes confer chemoresistance to pancreatic cancer cells by promoting ROS detoxification and miR-155-mediated suppression of key gemcitabine-metabolising enzyme, DCK. Br J Cancer. 2017February;116(5):609–619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Shi B, Wang Y, Zhao R, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-21 protects C-kit+ cardiac stem cells from oxidative injury through the PTEN/PI3K/Akt axis. PLoS One. 2018;13(2):e0191616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].Saeed-Zidane M, Linden L, Salilew-Wondim D, et al. Cellular and exosome mediated molecular defense mechanism in bovine granulosa cells exposed to oxidative stress. PLoS One. 2017;12(11):e0187569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Takahashi A, Okada R, Nagao K, et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat Commun. 2017;8:15287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Rodrigues ML, Nakayasu ES, Oliveira DL, et al. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryot Cell. 2008January;7(1):58–67. [DOI] [PMC free article] [PubMed] [Google Scholar]