

Abstract
The CRISPR (clustered regularly interspaced short palindromic repeat)-Cas system is an adaptive immune system of bacteria that has furnished several RNA-guided DNA endonucleases (e.g., Cas9) that are revolutionizing the field of genome engineering. Cas9 is being used to effect genomic alterations as well as in gene drives, where a particular trait may be propagated through a targeted species population over several generations. The ease of targeting catalytically impaired Cas9 to any genomic loci has led to development of technologies for base editing, chromatin imaging and modeling, epigenetic editing, and gene regulation. Unsurprisingly, Cas9 is being developed for numerous applications in biotechnology and biomedical research and as a gene therapy agent for multiple pathologies. There is a need for precise control of Cas9 activity over several dimensions, including those of dose, time, and space in these applications. Such precision controls, which are required of therapeutic agents, are particularly important for Cas9 as off-target effects, chromosomal translocations, immunogenic response, genotoxicity, and embryonic mosaicism are observed at elevated levels and with prolonged activity of Cas9. Here, we provide a perspective on advances in the precision control of Cas9 over aforementioned dimensions using external stimuli (e.g., small molecules or light) for controlled activation, inhibition, or degradation of Cas9.
Graphical Abstract
The CRISPR-Cas system is composed of clustered regularly interspaced short palindromic DNA repeats (CRISPR) and CRISPR-associated (Cas) genes that protect bacteria against invading phages and mobile geneticelements.1–7 These Cas genes have furnished DNA endonucleases that are transforming biomedical research and gene therapies.8–11 The most studied CRISPR-associated nuclease is SpCas9 (henceforth called Cas9) from Streptococcus pyogenes, but several next-generation nucleases are rapidly emerging.8 The CRISPR-associated nucleases recognize their substrate via a protospacer adjacent motif (PAM) sequence and base pairing of the target DNA by a guide RNA (gRNA) borne by the nuclease. Upon target recognition, the two DNA strands are cleaved by distinct nuclease domains of Cas9 (Figure 1A): the HNH domain cleaves the target strand, while the RuvC domain cleaves the nontarget strand.8 The resulting double-stranded break in the DNA can be repaired by errorprone nonhomologous end joining (NHEJ) or the more precise homology-directed repair (HDR) pathways.12,13 NHEJ leads to insertion/deletion (indel) mutations, causing frameshift mutations or premature stop codons and gene knockout. HDR, on the other hand, can be used to introduce specific edits in the genome using an exogenously supplied repair template.14
Figure 1.
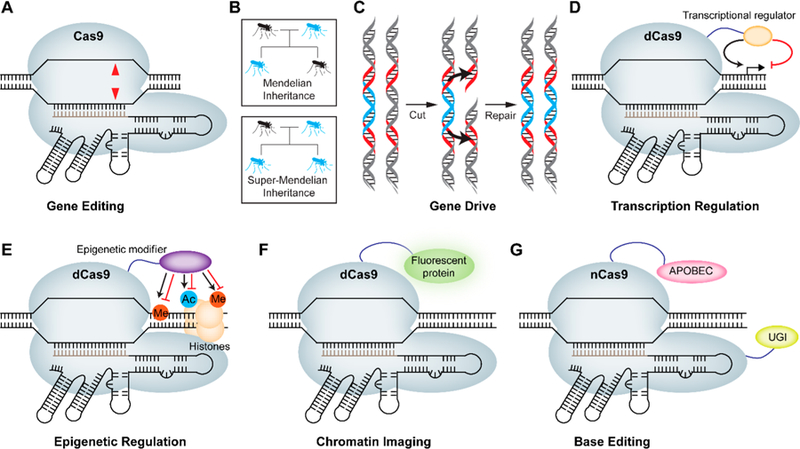
Major applications of CRISPR-Cas9. (A) Cas9 enables gene editing through its RNA-guided DNA endonuclease activity leading to double-stranded breaks (red triangles). (B) CRISPR-Cas9-based gene drive allows self-propagation of an engineered gene and/or trait in a species population. The probability of passing on an engineered gene (blue) to the progeny in Mendelian inheritance is 50%, while the same probability for gene drives approaches 100%. (C) The molecular mechanism of super-Mendelian inheritance in the gene drive involves a Cas9-induced double-stranded break on the wild type allele (gray) that is repaired by copying the drive allele (blue) from the engineered parent via HDR, causing replacement of the wild type allele with the drive element. Red regions indicate homology arms. (D) Nuclease dead Cas9 (dCas9) can be fused to an activator or repressor domain to regulate gene expression. (E) dCas9 can be fused to DNA-demethylating enzymes/methyltransferase as well as histone demethylase/methyltransferase or deacetylase/acetyltransferase domains to regulate epigenetic modifications in a sequence-specific manner. (F) dCas9 can be fused to a fluorescent protein such as GFP to enable imaging of a specific locus. (G) Nickase Cas9 (nCas9) can be fused to cytosine deaminase (APOBEC1) and uracil glycosylase inhibitor (UGI) to enable conversion of cytosine (C) to thymine (T) without double-stranded breaks.
In addition to introducing genomic alterations, CRISPRCas9-based gene drives have ushered in an era of “active genetics” where an engineered gene/trait can potentially self-propagate in the species population.15 In a normal Mendelian inheritance, the probability of passing on an engineered gene to the progeny is only 50%, while the same probability for gene drives approaches 100% (Figure 1B).16–18 Central to the molecular mechanism of super-Mendelian inheritance in the gene drive is the Cas9-induced double-stranded break on the wild type allele that is repaired by copying the allele from the engineered parent, essentially causing replacement of the wild type allele with the engineered one (Figure 1C). The super-Mendelian inheritance by gene drives has allowed ∼100% transmission of antimalarial or infertility genes in mosquitoes, enabling efficient population modification or suppression, respectively.19–22 Unsurprisingly, gene drives are under investigation for the elimination of diseases (e.g., malaria and Dengue fever) and could potentially be used to reverse pesticide resistance.16,18,19,21,22
The ease of targeting catalytically inactive Cas9 (called dCas9) to any genomic loci has led to the generation of transcriptional activators, repressors, epigenetic modifiers, and chromatin imaging systems. For instance, dCas9 has been fused to a variety of transcriptional activators to initiate transcription at a specific site (Figure 1D).23–30 Similarly, fusing dCas9 to transcriptional repression domains and targeting to gene promoters or exonic sequences has allowed for sequence-specific gene repression (Figure 1D).24,28,31–34 dCas9 has also been used to design programmable epigenetic modifiers targeting both the DNA locus of interest and the surrounding histones (Figure 1E). The alteration of DNA methylation status at a specific target locus was attained by fusing the catalytic domain of DNA methyltransferase (DNMT3A), while dCas9-TET1 fusions were used for DNA demethylation.35–42 Additionally, dCas9-LSD1 and dCas9-PRMD9 methyltransferase fusion complexes have been applied to demethylate and methylate histones, respectively, while dCas9-p300 and dCas9-HDAC3 fusions have been used to acetylate and deacetylate histones, respectively.43–46 Fluorescent protein fusions to dCas9 have allowed genomic imaging in live cells as well as probing molecular interactions at specific genomic loci (Figure 1F).32,33,47–49 Finally, a catalytically impaired “nickase” variant of Cas9 has been instrumental in base editing technologies (Figure 1F). Nickase Cas9 fusions to both APOBEC1 deaminase and uracil glycosylase inhibitor (UGI) afford a base editor that can convert C to T at the target site. Similarly, fusion of an evolved adenosine deaminase to nickase Cas9 has led to the development of a base editor capable of converting A to G.50,51
Precision control of Cas9 across multiple dimensions, including those of dose, time, and space, is critical for its several applications. Toxicology’s adage “The dose makes the poison” is particularly relevant for Cas9, as the substrate DNA is present at a concentration much lower than those of the nuclease.52–59 Unsurprisingly, significant off-target e ffects, chromosomal translocations, and genotoxicity are observed with elevated Cas9 activity.52–59 In addition, as o ff-target activity often occurs at a rate slower than that of on-target activity,60 restriction of the nuclease activity to a narrow temporal window is highly desired. Rapid degradation of Cas9 upon target editing may be required to evade the host’s immune surveillance of this bacterial protein. Following germline editing in embryos, the lingering Cas9 activity causes undesirable mosaicism at both genotypic and phenotypic levels.61–64 In the context of human germline editing, molecular safeguards are even more important for multiple reasons, including the potential to alter the human gene pool. Here, we discuss various approaches for precision control of Cas9 function over the dimensions of dose, time, and space. We describe methods for conditional activation of Cas9 using small molecules or light as well as molecules that can mediate deactivation through inhibition or degradation of Cas9.
CONTROL BY SMALL-MOLECULE ACTIVATORS
A relatively straightforward approach to control Cas9 activity is to regulate its transcription through an inducible promoter, as demonstrated in the work by Dow et al.66 and Gonzalez et al.65 in mammalian cells and mice, respectively. In addition, Kiani et al. and Aubry et al. have developed doxycycline-inducible gRNA systems for controlling Cas9-mediated genome regulation.67,68 Although robust gene editing was observed upon induction of Cas9 or gRNA in response to doxycycline treatment, these approaches have slow response times because of the lengthy time scale of transcription and translation65,66 and require additional factors (e.g., reverse tetracycline transactivator). In addition, transcriptional control is incapable of limiting Cas9 activity to the short temporal window that is necessary to maximize genome editing specificity. In contrast, strategies that post-translationally control protein activity o ffer a much higher temporal resolution.69–71
Several methods exist for post-translational control of protein function using small molecules (Figure 2A).70,71 Davis et al. inserted an evolved 4-hydroxytamoxifen (4-HT)-responsive intein at specific sites within Cas9 to disrupt enzymatic activity.69 Addition of 4-HT causes splicing of the intein to release active Cas9. Although the overall activity of this engineered Cas9 was slightly lower than that of Cas9, the ratio of on-target to o ff-target edits was approximately 6-fold higher. In another study, Liu et al. developed iCas9 by fusing the hormone binding domain of the estrogen receptor (ERT2) directly to Cas9, enabling tight temporal control of Cas9 using 4-HT.72 In the absence of 4-HT, the ERT2 domains sequester Cas9 in the cytoplasm, but upon addition of 4-HT, the fusion protein rapidly translocates to the nucleus where it can engage with genomic DNA. The cleavage activity of iCas9 was observed at multiple endogenous genomic loci as early as 4 h after 4-HT treatment. A comparison of iCas9 with the 4-HT intein-Cas9 system revealed that in the presence of inducer, iCas consistently showed a DNA cleavage efficiency higher than that of the 4-HT intein-Cas9 at all genomic loci and time points tested, although both systems had comparable background activity in the absence of an inducer. In addition, the iCas9 system was reversible and toggled between on and o ff states. The activity of iCas9 could be reversed in 72 h by removing the inducer 4-HT. Additionally, this system displayed improved specificity over wild type Cas9 at most endogenous loci and sgRNAs tested when the duration of the 4-HT treatment was restricted to 4−8 h. However, iCas9 still had residual activity in the absence of 4-HT, and the most optimal construct retained only 60% of wild type Cas9 activity.72 In another study, Oakes et al. used randomized insertional mutagenesis to insert a small domain into the Cas9 sequence and screened for active variants, thus identifiying structural “hot spots” within Cas9 that could tolerate additional protein domains.73 Insertion of the ligand binding domain of human estrogen receptor-α into position 231 of Cas9 or dCas9 a fforded a 4-HT-responsive Cas9 (arC9) or dCas9 (darC9), respectively. The authors demonstrated 4-HT dose-dependent repression by darC9 (EC50 = 440 ± 70 nM) in CRISPRi studies as well as dose-dependent control of Cas9 (arC9) with an EC50 of 1 nM. With arC9, almost no background was observed in the absence of 4-HT, but the maximum activity attained was still only 30% of that of wild type Cas9. Reversibility studies indicated that the activation of arC9 could be turned o ff by removing 4-HT from the media after 6 days. However, a small amount of residual arC9 activity remained even after ligand removal, possibly due to the high binding affinity of arC9 for 4-HT and slow dissociation of the complex.73
Figure 2.
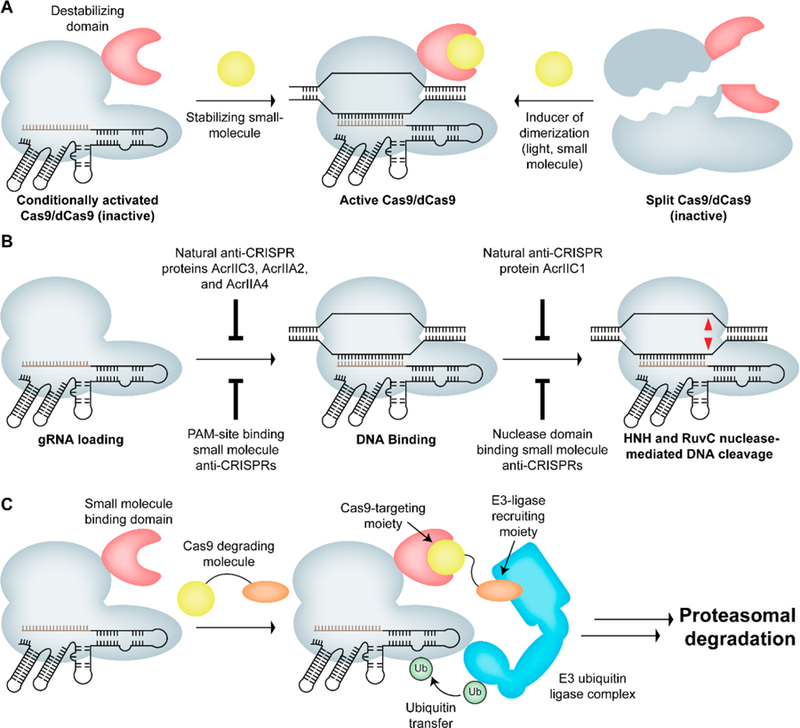
Strategies for conditional control of Cas9 activity. (A) Cas9 can be inactivated by fusing a small molecule or light-responsive domain, or it can be split into N- and C-terminal fragments that can be reconstituted in response to a small molecule or light, leading to the formation of active Cas9. (B) Steps involved in Cas9-mediated strand DNA cleavage. Any of the steps shown above can be inhibited by a protein or small-molecule inhibitor leading to loss of Cas9 activity. (C) Strategy for Cas9 degradation using a heterobifunctional small molecule such that one end of the molecule binds to the small-molecule binding domain fused to Cas9 and the other end binds to CRBN, resulting in ubiquitination and proteasomal degradation of Cas9.
Several groups have developed various small-molecule-controlled Cas9 systems based on the chemically induced dimerization of split protein fragments (Figure 2A). A well-studied example of this type of system uses the rapamycin-mediated dimerization of FK506 binding protein 12 (FKBP) and FKBP rapamycin binding domain (FRB) of the mammalian target of rapamycin (mTOR).74,75 Zetsche et al. designed a split Cas9 system in which the C-terminal fragment of Cas9 was fused to FKBP and the N-terminal fragment was fused to the FRB domain.76 Additionally, they appended a nuclear export signal (NES) to the N-terminal fragment and a nuclear localization signal (NLS) to the C-terminal fragment to prevent spontaneous reconstitution of the two fragments, reducing basal activity in the absence of rapamycin. This design produced low levels of Cas9 activity in the absence of the molecule but irreversible activation upon rapamycin addition. Furthermore, the authors demonstrated substantial indels at the intended genomic loci with no significant o ff-target e ffects upon induction of this split-Cas9 system with rapamycin.76 The availability of orthogonal small-molecule regulators that utilize multiple chemically induced dimerization systems has led to the development of orthogonal gene regulation systems. Gao et al. demonstrated dose-dependent and reversible transcriptional activation/repression using abscisic acid-inducible ABI-PYL1 and gibberellin-inducible GID1-GAI heterodimerization domains.77–80 In this case, dCas9 was fused to either ABI or GAI while the e ffector (transcription activator or repressor) domains were fused to PYL1 or GID1, respectively, allowing orthogonal and multiplexed transcription regulation without significant background. Detectable levels of transcriptional activation were observed within 24 h. Furthermore, these systems were reversible upon removal of the inducer, with the activity reaching baseline levels in 4−5 days.77,78 In another study, Bao et al. utilized the gibberellin- and rapamycin-mediated dimerization systems to demonstrate orthogonal and temporal regulation of multiple endogenous genes.77,81
A complementary approach to inducing dimerization of protein domains using small molecules is to disrupt the dimerization process using small molecules. Rose et al. used this method to develop a rapidly inducible Cas9 (ciCas9) system using the interaction between BCL-xL and BH3 peptide as an intramolecular inhibitory switch.82,83 In this case, the REC2 domain of Cas9 was replaced with BCL-xL to yield Cas9.BCL, and a BH3 peptide was attached to both N- and C-termini of Cas9.BCL.83 The intramolecular interaction between BCL-xL and the BH3 peptides keeps Cas9 in an inactive state. Addition of the small-molecule inhibitor of BCL-xL, A-385358 (A3),84 disrupted the interaction between BH3 and BCL-xL, activating Cas9. The ciCas9 system showed dose-dependent control of nuclease activity with the degree of activation being higher than that of the 4-HT-induced intein-Cas9 and iCas9 systems.69,72,83 Furthermore, ciCas9 showed lower background activity and o ff-target e ffects compared to those of Cas9 and could induce indel formation in as little as 2 h. However, the maximal activity reached by this system is still lower than that of Cas9.83
In addition to the use of small-molecule-inducible protein domains, another strategy that adds variety to the repertoire of regulated CRISPR-Cas9 systems is the fusion of conditional degrons to Cas9 (Figure 2A). Conditional degrons can be activated or deactivated by one of several external factors such as a small-molecule ligand, light, temperature, or another protein.85 We and others have adopted this approach to design destabilized Cas9 systems whose activity can be tightly regulated by a small-molecule stabilizer. Work from our laboratory demonstrated the application of destabilized Cas9 systems for both gene editing and transcription activation by fusing the destabilized domains from Escherichia coli dihydrofolate reductase (DHFR) and estrogen receptor (ER50) to Cas9 or the transcription activation domains. Transcriptional activation by these destabilized Cas9 systems could be reversed within 48 h by removing the stabilizing small molecule (TMP or 4OHT), with a rapid decrease in mRNA transcript levels seen in as little as 8 h. Furthermore, these systems showed improved specificity over that of wild type Cas9 upon administration of an optimal dose of the stabilizing small molecules.56 Senturk et al. developed a destabilized Cas9 system by fusing the FKBP-derived destabilized domain to Cas9, which rapidly degrades the protein upon its expression.86 However, the stabilizing small molecule shield-1 a ffords rapid, reversible, and temporal control of FKBP-Cas9 gene editing activity. Analysis of the protein induction kinetics revealed that destabilized Cas9 was expressed within 2 h of treatment with shield-1. This e ffect could be reversed within 2 h of withdrawal of the ligand from the media, with protein levels becoming undetectable after 12 h. Furthermore, treatment with shield-1 led to the formation of indels at the target locus within 24 h to an extent comparable to that seen for wild type Cas9. The authors also demonstrated the application of this destabilized Cas9 system in three-dimensional organoid models, indicating the utility of this system as a screening tool in primary tumor cells. They also demonstrated temporal and independent control of gene editing and Cre-mediated recombination. Therefore, this method could potentially enable interrogation of genetic interactions in various preexisting mouse models of human diseases based on the Cre-lox system.86 Kleinjan et al. utilized the auxin-inducible degron (AID) and the E. coli DHFR degron to develop conditionally degradable dCas9-based e ffector proteins. The AID-dCas9 system exhibited rapid degradation kinetics upon addition of auxin (1−2 h). Similarly, this e ffect was rapidly reversed when auxin was removed from the media, leading to the reappearance of detectable levels of the fusion protein in 30 min and maximum expression in the next 3−7 h. The AID approach was also extended to specific orthogonal Cas9 and Cpf1 proteins, a ffording orthogonal, auxin-degradable synthetic transcription factors.87 Jacobs et al. inserted stabilizable polypeptide linkages (StaPLs) based on hepatitis C virus NS3 protease domain into internal loops of dCas9 to enable conditional control of Cas9 activity. StaPLs undergo autoproteolysis to cleave the proteins to which they are attached, though the presence of a protease inhibitor prevents their cleavage to preserve protein function. Using this system, they demonstrated dose-dependent transcription activation, but the system was irreversible.88 In a similar approach, Tague et al. used the NS3 protease domain and its inhibitor BILN-2061 as a ligand-inducible connection (LInC) to control the association of DNA binding and transcription activation domains. In their design, the viral protease was incorporated into dCas9-VPR such that the protease was positioned between the DNA binding sca ffold and the C-terminal region that contained a nuclear localization sequence (NLS) and the VPR transcription activation domain (dCas9-NS3-NLS-VPR). In the absence of the protease inhibitor, NS3 autocleavage separated the transcription activation domain from dCas9, preventing transcriptional upregulation. Using the LInC-containing dCas9-VP64 system, the authors demonstrated transcription activation of the target gene in a dose-dependent manner, with activity comparable to that of the dCas9-VP64 system without LInC. They also demonstrated the application of LInC in recruiting a transcription activation domain to hairpin-modified sgRNA.89
Control of Cas9 activity has also been exercised at the level of the gRNA. Post-transcriptional control of gRNA can potentially be used to independently regulate multiple targets, because each target has its unique guide.90 Toward this end, aptamers and aptazymes have been fused with gRNA to attain ligand-dependent control. Liu et al. engineered the gRNA by incorporating tetracycline- and theophylline-responsive riboswitches within the gRNA.91 In the absence of a small-molecule trigger, the spacer portion of the gRNA is base-paired with a designed antisense strand, preventing gRNA:DNA base pairing. Upon addition of the small molecule, the aptamer undergoes a conformational change that allows gRNA to bind to the target DNA. This allowed transcriptional regulation of endogenous genes in a dose-dependent manner upon stimulation by riboswitch-responsive signals.91 Similarly, Tang et al. incorporated theophylline- and guanine-responsive aptazymes into gRNA sequences, which a fforded dose control of transcriptional and nuclease activity but were irreversible and exhibited significant background activity in the absence of small-molecule triggers.92 To overcome some of these shortcomings in the first-generation systems, Kundert et al. developed a ligand-responsive gRNA that can be activated or deactivated upon ligand addition.90 The ligand-activated gRNA (ligRNA+) was obtained by inserting the theophylline aptamer into the hairpin region while the ligand-deactivated gRNA (ligRNA−) had the aptamer inserted into the nexus. Both ligRNA+ and ligRNA− showed robust and titratable activity over a large range of ligand concentrations. Using two di fferent ligRNA− variants allowed simultaneous and reversible regulation of two genes.90 Additionally, Pu et al. utilized evolved, split RNA polymerase-based biosensors that drive the production of target gRNAs when activated using small molecules or protein−protein interacting domains. Using this system, the authors demonstrated multidimensional control of SaCas9 function.93
CONTROL BY LIGHT
Over the past decade, light has emerged as an attractive tool to control protein function. The high spatiotemporal resolution and non-invasiveness of light induction provides several advantages over other external stimuli. As such, several optically controlled Cas9 systems have been designed to enable precise genomic modifications. Nihongaki et al. developed a system based on cryptochrome 2 (CRY2) and calcium and integrin binding protein 1 (CIB1), which can heterodimerize in the presence of blue light.94 By fusing dCas9 with CIB1, and a transcriptional activator domain (VP64 or p65) with the CRY2 domain, the authors demonstrated spatiotemporal and multiplexed regulation of endogenous genes with light. The duration of blue light irradiation required to produce detectable levels of upregulation ranged from 1 h for a reporter gene to 3 h for an endogenous gene. This e ffect could be reversed by incubating the cells in the dark for 18 h.94 Polstein et al. pursued a similar strategy using multiple copies of the N-terminal fragment of CIB1 (CIBN) fused to dCas9’s N- and C-termini and CRY2 fused to VP64. CIBN-dCas9-CIBN showed robust and time-dependent upregulation of the target gene upon exposure to light, which was reversed by simply switching o ff the light source, with time scales similar to those of the system of Nihongaki et al.95
Another study by Nihongaki et al. describes a photo-activatable split Cas9 (paCas9) strategy (Figure 2A).96 Here, the N- and C-terminal fragments of Cas9 were fused to engineered, light-responsive domains called positive magnet (pMag) and negative magnet (nMag). Upon exposure to blue light, pMag and nMag dimerize, a ffording a new split system with a lower background and higher fold induction of Cas9 activity. Additionally, paCas9 exhibited similar nuclease activity and targeting specificity to the wild type Cas9 and could be used to induce random indel mutations as well as more precise genomic modifications through HDR in response to light. As with the other protein-based light-inducible systems, paCas9 activity could also be reversed by switching off the light source.96 In contrast to paCas9, the psCas9 system developed by Zhou et al. employs a single-polypeptide architecture designed by inserting the photodissociable dimeric fluorescent protein pdDronpa1 into the REC2 and PI domains of Cas9.97,98 In the dark, the dimerization of the inserted pdDronpa1 domains sterically inhibits Cas9 activity, but upon illumination with 500 nm light, pdDronpa1 dissociates and enables Cas9 to carry out gene editing functions as well as transcriptional upregulation.98 Hemphill et al. developed a genetically encoded light-activated Cas9 system by incorporation of a photocaged lysine to a residue that is critical for gRNA binding (e.g., K866). The activity of this photocaged Cas9 was restored upon a brief (120 s) exposure to 365 nm light. However, unlike some of the other light-activated systems, this system is irreversible due to the irreversible nature of the lysine chemical modification.99
As with small-molecule-inducible systems, the bulk of light-controlled Cas9 systems involve engineering of the Cas9 protein. In the only study involving photocaged gRNA, Jain et al. used photocleavable protector oligonucleotides that hybridize with the gRNA, preventing gRNA:DNA base pairing. Upon exposure to light, the protector oligo undergoes photolysis, producing short fragments with reduced binding affinity for the gRNA because of their lower melting temperature. However, once activated, the gRNA cannot be deactivated by removal of illumination, making this system irreversible.100
While several methods for precision activation of Cas9 in mammalian cell culture exist, genome editing is being used in several nonmammalian organisms (e.g., plants and insects) for which methods for precision activation are underdeveloped. In addition, methods for precision activation of Cas9 in organismal settings are still a challenge even for mammalian systems. Such controls will be useful in several therapeutic settings and within the context of gene drives.101 For example, currently switching on gene drive involves a split system in which the Cas9 gene is delivered from one parent and gRNA from another; however, this genetic method is slow and inefficient, and the output is binary (i.e., 0 or 100% super-Mendelian inheritance). Much interest exists to develop gene drives whose output can be fine-tuned to any values from 0 to 100% super-Mendelian inheritance rapidly. Such control, which can be accomplished using conditional activators of Cas9, will enable facile animal husbandry and population expansion of insects with gene drives propagating lethal traits. In addition, numerous CRISPR systems that target RNA have been discovered and have been used to develop highly sensitive diagnostic tools for diseases caused by Zika and Dengue virus.102–104 The RNA targeting Cas13a has also been used to mediate RNA base editing.105 Development of inducible RNA targeting CRISPR systems and exploration of their application for conditional control of various types of RNAs in cells will be interesting. Such programmable and inducible RNA-targeting modules could find applications in cancer diagnosis and therapy without a ffecting wild type transcripts or making any genomic alterations.106
CONTROL BY INHIBITORS
With the recent discovery of natural genetically encoded antagonists of CRISPR systems (called “anti-CRISPRs”), there has been a surge of interest in co-opting these proteins to achieve context-specific inhibition of Cas9. Protein-based anti-CRISPRs are small accessory proteins with fewer than ∼200 amino acids found in the genomes of bacteriophages that bind to and inhibit CRISPR-Cas machinery.107–109 These proteins allow phage to evade the bacterial immune response, allowing phage propagation and making anti-CRISPRs crucial components in the pervasive arms race between bacteria and phage. Indeed, this arms race has resulted in a tremendous diversity of anti-CRISPR proteins, with more than 20 di fferent anti-CRISPR families targeting type I and type II CRISPR-Cas systems currently characterized110 (including anti-CRISPRs that target SpCas9, such as AcrIIA2 and AcrIIA4).111 Anti-CRISPRs can interfere with the CRISPR-Cas machinery in a variety of ways as well, such as by binding to the gRNA-loaded CRISPR-Cas complex and preventing DNA binding,112–115 binding to Cas e ffector proteins to block their recruitment to active Cascade complexes112 (type I systems), or directly inhibiting nuclease activity of the Cas9 protein (Figure 2B).115
While anti-CRISPRs tend to show little sequence homology with other anti-CRISPR proteins, their continued discovery has been aided by their frequent association with a putative “helix−turn−helix” (HTH) regulatory protein in the phage anti-CRISPR locus (called the anti-CRISPR associated protein, or aca).116 Discovery of aca homologues in other phage genomes has allowed putative anti-CRISPRs to be identified through “guilt by association”, simplifying the characterization of these open reading frames.117,118 Recently, expansion of bioinformatic methodologies to discover novel anti-CRISPR proteins include pipelines to identify bacteria harboring “self-targeting” CRISPR spacers, indicating that survival of the bacteria depends upon the ability to self-inhibit CRISPR proteins via bacterially encoded anti-CRISPR proteins.111 This approach has recently led to the discovery of novel anti-CRISPRs targeting type V CRISPR systems (Cpf1 or Cas12).119,120 Given the important role anti-CRISPRs play for phage in counteracting bacterial defenses, it is certain similarly clever bioinformatics strategies will discover anti-CRISPRs for all remaining CRISPR-Cas types, including the RNA-targeting type VI CRISPR nuclease Cas13.
Anti-CRISPRs have provided a natural solution for mitigating two of the main problems plaguing CRISPR-mediated genome editing: o ff-target cleavage events110 and DNA damage-induced toxicity.121 As o ff-target activity and DNA double-stranded break (DSB)-induced toxicity increase with expression time of the CRISPR-associated nuclease, handicapping or outright inhibiting Cas9 activity after the desired double-strand break would be expected to alleviate these issues. Indeed, the work by Shin et al. has shown that timed transfection of AcrIIA4 protein or plasmid can reduce the number of known o ff-target edits of Cas9 at the HBB and VEGFA loci in K562 human cells.60 Li et al. have also shown that temporally staggered delivery of Cas9/gRNA and AcrIIA2 and AcrIIA4 anti-CRISPR proteins to hematopoietic stem cells via helper-dependent adenovirus improves their eventual engraftment into host mice tissue while preserving the desired edit,122 presumably due to minimization of double-strand break genotoxicity. However, like the Cas9/gRNA complexes themselves, anti-CRISPRs su ffer from poor pharmacokinetic properties and delivery issues. The efficacy of anti-CRISPRs in selectively impairing o ff-target editing while retaining high on-target activity is highly dependent on the timing of addition: if anti-CRISPRs are added concurrently with Cas9/gRNA, the overall editing efficiency is dramatically decreased. Ideally, the addition of anti-CRISPRs follows the introduction of active Cas9/gRNA complexes, but this timing is dependent on the method of Cas9/gRNA delivery (plasmid, a ribonucleoprotein complex, or virus) and will need to be optimized for each situation. With anti-CRISPRs, staggered addition paradigms would also require multiple cycles of nucleofection or adenovirus infection, which may introduce other sources of stress and toxicity in the cells. As such, delivery of anti-CRISPRs poses a large hurdle to the routine application as part of the genome editing toolbox. Moreover, as some anti-CRISPR proteins function as a DNA mimic, the possibility of blocking certain transcription factors by anti-CRISPR proteins cannot be ruled out, and the specificity of anti-CRISPR proteins in the context of mammalian cells should be evaluated.123
Small-molecule inhibitors of Cas9 can overcome some of the challenges associated with anti-CRISPR proteins without compromising functional activity. Small-molecule inhibitors can easily be delivered to cell’s nuclei through passive di ffusion, are stable to proteases, and are generally non-immunogenic. Small-molecule inhibitors exhibit fast kinetics, in stark contrast to genetic methods,124 and can be synthesized with little batch-to-batch variability. Unsurprisingly, small molecules remain the preferred reagents for dose and temporal control of intracellular targets. However, developing small-molecule inhibitors of Cas9 is challenging for multiple reasons. First, Cas9 possesses novel protein folds limiting the application of rational design approaches.125 Second, inhibitor identification requires multiple robust, orthogonal, high-throughput assays of Cas9, which are mostly unavailable. Third, Cas9 is a single-turnover enzyme that holds on to its substrate with picomolar affinity,126 further complicating development of such high-throughput assays. Fourth, the inhibition of Cas9 activity requires the inactivation of two nuclease domains. Finally, Cas9 belongs to the class of DNA binding proteins that are often deemed chemically intractable.127
Beyond specificity enhancements, anti-CRISPR molecules will be useful for several other contexts. First, in germline editing, restricting Cas9 activity to a narrow temporal window is important, as persistent activity in dividing cells contributes to mosaicism.61,63 Timely Cas9 degradation reduced mosaicism in non-human primate embryos.62 Second, a Cas9 inhibitor will be useful in the context of gene drives that propagate lethal traits.15,16,18,128 Here, temporarily switching o ff gene drives by inhibiting Cas9 will allow facile animal husbandry and population expansion of mosquitoes for field studies.21 In addition, dose and temporal control of gene drives in a laboratory setting will allow precision population control and propel our understanding of the limits of this technology. Third, inhibition of Cas9-mediated genotoxicity to helper cells can enable efficient packaging of Cas9 in adeno-associated virus (AAV) for delivery.129 Another avenue in which the synthetic inhibitors appear to be promising is organ-specific disabling of CRISPR-Cas9. Even though AAV-mediated delivery of CRISPR/Cas has been encouraging, its accumulation in the liver, heart, and kidney remains a major concern.130 In such a scenario, small-molecule inhibitors can be administered in an organ-specific way to suppress o ff-target activity in tissues remote from the site of administration. Fourth, Cas9 inhibitors could help allay dual-use concerns from a biosafety perspective in the use of Cas9 in disease modeling.131 Fifth, Cas9 inhibitors will propel the fundamental understanding of the biological functions of endogenous Cas9 and allow application of immune response-based selection pressure on bacteria for the evolution of new CRISPR-based systems. We note that the presence of CRISPR-based defense systems within bacteria has hampered e fforts to develop phages as next-generation antibiotics. A cocktail of phage and the inhibitors of these defense systems may provide an approach for species-selective disruption while leaving beneficial bacteria una ffected, and such studies may lay the foundation for precision anti-infectives.
CONTROL BY DEGRADERS
In many scenarios, timely degradation of Cas9 may be preferred over outright inhibition. Several technologies have been developed for post-translational regulation of protein levels using small molecules.132,133 Many of these methods involve the use of heterobifunctional molecules that lead to the co-localization of the target protein and specific ubiquitin ligases that are involved in the proteasomal degradation pathway.134–136 Some of the other strategies involve fusion of a degron to the protein of interest such that degradation is induced upon addition of the small molecule or exposure to light. Examples of this include the auxin-induced degradation system, the LID domain, the B-LID domain, and the SMASh system.137–140 Another technology that leverages the ubiquitin-independent proteasomal degradation was identified in ornithine decarboxylase (ODC) and antizyme (AZ).141,142 Here, the protein of interest is fused to ODC that is degraded in a ubiquitin-independent manner upon co-expression of antizyme.141 More recently, Nabet et al. have reported a small-molecule degradation system (called dTAG system) in which the target protein is fused to the FKBP12F36V variant.143 Upon addition of a heterobifunctional molecule that can recruit specific E3/E2 ubiquitin ligase to the FKBP12F36V variant, the fusion protein is ubiquitinated and degraded. This heterobifunctional molecule (called dTAG) consists of a selective binder of FKBP12F36V and a CRBN binder. One can envision the application of similar strategies for the degradation of Cas9, wherein Cas9 is fused to one or more degron domains and degraded upon addition of the corresponding small molecule or light. Alternatively, the small-molecule inhibitors of Cas9 can be converted to heterobifunctional molecules bearing a ubiquitin ligase binder. These small-molecule degraders, which operate by “event-driven pharmacology”, are catalytic compared to Cas9 inhibitors that operate via “occupancy-driven pharmacology” and are stoichiometric (Figure 2C).144
Degradation of Cas9 might be preferred over inhibition in applications in which complete removal of the Cas9 protein is desired. For example, a recent study has pointed to the existence of antibodies against Cas9 in humans,145 and the Cas9-specific immune response is construed as a major bottleneck in the development of therapeutic applications of Cas9. Reducing the half-life of Cas9 may reduce the severity of the immune response. Cas9 degraders could also be useful for reducing Cas9-mediated genotoxicity. Cas9-induced double-strand breaks activate the p53-mediated DNA damage response leading to cell cycle arrest in the G1 phase and apoptosis.14,146,147 Such genotoxicity not only reduces the fraction of edited cells in a treated population but also selects for apoptosis-resistant clones with potential tumorigenic properties. Cas9 is known to persist for >6 h at the site of the DNA double-strand break, perhaps preventing DNA repair and/or causing stalled replication forks that might contribute to Cas9 genotoxicity.14,147 Cas9 degraders may also reduce the mosaicism in germline editing. While Cas9-mediated germline editing has revolutionized disease modeling, the appearance of mosaic mutations in the embryo with di fferent mutations in di fferent cell types in the same animal is a major problem.61–63 Such mosaic mutations are quite extensive in zebra fish, and additional crosses are required to dilute mosaicism in small animals. In large animals (e.g., non-human primates), where sexual maturity takes multiple years, mosaic mutations can be a major obstacle to obtaining genetically modified animals. Restricting the half-life of Cas9 reduced mosaicism, but the reported method involved fusing Cas9 to a constitutively active degron without temporal control.62 One can also envision that other strategies to degrade Cas9 in response to a specific stimulus would limit the duration of Cas9 activity to the early embryo stage. These approaches could have similar e ffects in reducing mosaic genotypes, thus contributing to the overall efficacy and precision of Cas9 gene editing.
Acknowledgments
Funding This work was supported by the Burroughs Wellcome Fund (Career Award at the Scientific Interface), DARPA (Brdi N66001–17-2–4055), the National Institutes of Health (1R21AI126239–01), and Army Research Office Grant W911NF1610586.
Footnotes
Notes
The authors declare the following competing financial interest(s): The Broad Institute has filed patent applications for many inventions related to CRISPR, including inventions by authors of this Perspective related to precision control of CRISPR.
REFERENCES
- (1).Makarova KS, Haft DH, Barrangou R, Brouns SJ, Charpentier E, Horvath P, Moineau S, Mojica FJ, Wolf YI, Yakunin AF, van der Oost J, and Koonin EV (2011) Evolution and classification of the crispr-cas systems. Nat. Rev. Microbiol 9, 467–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Makarova KS, Wolf YI, Alkhnbashi OS, Costa F, Shah SA, Saunders SJ, Barrangou R, Brouns SJ, Charpentier E, Haft DH, Horvath P, Moineau S, Mojica FJ, Terns RM, Terns MP, White MF, Yakunin AF, Garrett RA, van der Oost J, Backofen R, and Koonin EV (2015) An updated evolutionary classification of crispr-cas systems. Nat. Rev. Microbiol 13, 722–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Charpentier E, and Marraffini LA (2014) Harnessing crisprcas9 immunity for genetic engineering. Curr. Opin. Microbiol 19, 114–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Doudna JA, and Charpentier E (2014) Genome editing. The new frontier of genome engineering with crispr-cas9. Science 346, 1258096. [DOI] [PubMed] [Google Scholar]
- (5).Hille F, and Charpentier E (2016) Crispr-cas: Biology, mechanisms and relevance. Philos. Trans. R. Soc., B 371, 20150496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Barrangou R, Birmingham A, Wiemann S, Beijersbergen RL, Hornung V, and Smith A (2015) Advances in crispr-cas9 genome engineering: Lessons learned from rna interference. Nucleic Acids Res 43, 3407–3419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Bondy-Denomy J, and Davidson AR (2014) To acquire or resist: The complex biological effects of crispr-cas systems. Trends Microbiol 22, 218–225. [DOI] [PubMed] [Google Scholar]
- (8).Chen JS, and Doudna JA (2017) The chemistry of cas9 and its crispr colleagues. Nat. Rev. Chem 1, 0078. [Google Scholar]
- (9).Cox DB, Platt RJ, and Zhang F (2015) Therapeutic genome editing: Prospects and challenges. Nat. Med 21, 121–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Bryson TE, Anglin CM, Bridges PH, and Cottle RN (2017) Nuclease-mediated gene therapies for inherited metabolic diseases of the liver. Yale J. Biol. Med 90, 553–566. [PMC free article] [PubMed] [Google Scholar]
- (11).Saayman S, Ali SA, Morris KV, and Weinberg MS (2015) The therapeutic application of crispr/cas9 technologies for hiv. Expert Opin. Biol. Ther 15, 819–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Gallagher DN, and Haber JE (2018) Repair of a site-specific DNA cleavage: Old-school lessons for cas9-mediated gene editing. ACS Chem. Biol 13, 397–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Jasin M, and Rothstein R (2013) Repair of strand breaks by homologous recombination. Cold Spring Harbor Perspect. Biol 5, a012740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Richardson CD, Ray GJ, DeWitt MA, Curie GL, and Corn JE (2016) Enhancing homology-directed genome editing by catalytically active and inactive crispr-cas9 using asymmetric donor DNA. Nat. Biotechnol 34, 339–344. [DOI] [PubMed] [Google Scholar]
- (15).Gantz VM, and Bier E (2016) The dawn of active genetics. BioEssays 38, 50–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Esvelt KM, Smidler AL, Catteruccia F, and Church GM (2014) Concerning rna-guided gene drives for the alteration of wild populations. eLife 3, e03401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (17).Burt A, and Crisanti A (2018) Gene drive: Evolved and synthetic. ACS Chem. Biol 13, 343–346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (18).Champer J, Buchman A, and Akbari OS (2016) Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet 17, 146–159. [DOI] [PubMed] [Google Scholar]
- (19).Gantz VM, Jasinskiene N, Tatarenkova O, Fazekas A, Macias VM, Bier E, and James AA (2015) Highly efficient cas9-mediated gene drive for population modification of the malaria vector mosquito anopheles stephensi. Proc. Natl. Acad. Sci. U. S. A 112, E6736–6743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Gantz VM, and Bier E (2015) The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 348, 442–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Hammond A, Galizi R, Kyrou K, Simoni A, Siniscalchi C, Katsanos D, Gribble M, Baker D, Marois E, Russell S, Burt A, Windbichler N, Crisanti A, and Nolan T (2016) A crispr-cas9 gene drive system targeting female reproduction in the malaria mosquito vector anopheles gambiae. Nat. Biotechnol 34, 78–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (22).Kyrou K, Hammond A, Galizi R, Kranjc N, Burt A, Beaghton A, Nolan T, and Crisanti A (2018) A crispr−cas9 gene drive targeting doublesex causes complete population suppression in caged anopheles gambiae mosquitoes. Nat. Biotechnol 36, 1062–1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Mali P, Aach J, Stranges PB, Esvelt KM, Moosburner M, Kosuri S, Yang L, and Church GM (2013) Cas9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat. Biotechnol 31, 833–838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (24).Gilbert LA, Larson MH, Morsut L, Liu Z, Brar GA, Torres SE, Stern-Ginossar N, Brandman O, Whitehead EH, Doudna JA, Lim WA, Weissman JS, and Qi LS (2013) Crispr-mediated modular rna-guided regulation of transcription in eukaryotes. Cell 154, 442–451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (25).Maeder ML, Linder SJ, Cascio VM, Fu Y, Ho QH, and Joung JK (2013) Crispr rna-guided activation of endogenous human genes. Nat. Methods 10, 977–979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (26).Perez-Pinera P, Kocak DD, Vockley CM, Adler AF, Kabadi AM, Polstein LR, Thakore PI, Glass KA, Ousterout DG, Leong KW, Guilak F, Crawford GE, Reddy TE, and Gersbach CA (2013) Rna-guided gene activation by crispr-cas9-based transcription factors. Nat. Methods 10, 973–976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Cheng AW, Wang H, Yang H, Shi L, Katz Y, Theunissen TW, Rangarajan S, Shivalila CS, Dadon DB, and Jaenisch R (2013) Multiplexed activation of endogenous genes by crispr-on, an rna-guided transcriptional activator system. Cell Res 23, 1163–1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Zalatan JG, Lee ME, Almeida R, Gilbert LA, Whitehead EH, La Russa M, Tsai JC, Weissman JS, Dueber JE, Qi LS, and Lim WA (2015) Engineering complex synthetic transcriptional programs with crispr rna scaffolds. Cell 160, 339–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (29).Chavez A, Scheiman J, Vora S, Pruitt BW, Tuttle M, Iyer EPR, Lin S, Kiani S, Guzman CD, Wiegand DJ, Ter-Ovanesyan D, Braff JL, Davidsohn N, Housden BE, Perrimon N, Weiss R, Aach J, Collins JJ, and Church GM (2015) Highly efficient cas9-mediated transcriptional programming. Nat. Methods 12, 326–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (30).Konermann S, Brigham MD, Trevino AE, Joung J, Abudayyeh OO, Barcena C, Hsu PD, Habib N, Gootenberg JS, Nishimasu H, Nureki O, and Zhang F (2015) Genome-scale transcriptional activation by an engineered crispr-cas9 complex. Nature 517, 583–588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Gilbert LA, Horlbeck MA, Adamson B, Villalta JE, Chen Y, Whitehead EH, Guimaraes C, Panning B, Ploegh HL, Bassik MC, Qi LS, Kampmann M, and Weissman JS (2014) Genome-scale crispr-mediated control of gene repression and activation. Cell 159, 647–661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (32).Dominguez AA, Lim WA, and Qi LS (2016) Beyond editing: Repurposing crispr−cas9 for precision genome regulation and interrogation. Nat. Rev. Mol. Cell Biol 17, 5–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (33).Shalem O, Sanjana NE, and Zhang F (2015) High-throughput functional genomics using crispr-cas9. Nat. Rev. Genet 16, 299–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Wang H, La Russa M, and Qi LS (2016) Crispr/cas9 in genome editing and beyond. Annu. Rev. Biochem 85, 227–264. [DOI] [PubMed] [Google Scholar]
- (35).Liu XS, Wu H, Ji X, Stelzer Y, Wu X, Czauderna S, Shu J, Dadon D, Young RA, and Jaenisch R (2016) Editing DNA methylation in the mammalian genome. Cell 167, 233–247.e217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (36).Amabile A, Migliara A, Capasso P, Biffi M, Cittaro D, Naldini L, and Lombardo A (2016) Inheritable silencing of endogenous genes by hit-and-run targeted epigenetic editing. Cell 167, 219–232.e214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (37).Vojta A, Dobrinic P, Tadic V, Bockor L, Korac P, Julg B, Klasic M, and Zoldos V (2016) Repurposing the crispr-cas9 system for targeted DNA methylation. Nucleic Acids Res 44, 5615–5628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (38).McDonald JI, Celik H, Rois LE, Fishberger G, Fowler T, Rees R, Kramer A, Martens A, Edwards JR, and Challen GA (2016) Reprogrammable crispr/cas9-based system for inducing site-specific DNA methylation. Biol. Open 5, 866–874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (39).Lei Y, Zhang X, Su J, Jeong M, Gundry MC, Huang YH, Zhou Y, Li W, and Goodell MA (2017) Targeted DNA methylation in vivo using an engineered dcas9-mq1 fusion protein. Nat. Commun 8, 16026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (40).Xiong T, Meister GE, Workman RE, Kato NC, Spellberg MJ, Turker F, Timp W, Ostermeier M, and Novina CD (2017) Targeted DNA methylation in human cells using engineered dcas9-methyltransferases. Sci. Rep 7, 6732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (41).Morita S, Noguchi H, Horii T, Nakabayashi K, Kimura M, Okamura K, Sakai A, Nakashima H, Hata K, Nakashima K, and Hatada I (2016) Targeted DNA demethylation in vivo using dcas9-peptide repeat and scfv-tet1 catalytic domain fusions. Nat. Biotechnol 34, 1060–1065. [DOI] [PubMed] [Google Scholar]
- (42).Xu X, Tao Y, Gao X, Zhang L, Li X, Zou W, Ruan K, Wang F, Xu G, and Hu R (2016) A crispr-based approach for targeted DNA demethylation. Cell Discovery 2, 16009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (43).Kearns NA, Pham H, Tabak B, Genga RM, Silverstein NJ, Garber M, and Maehr R (2015) Functional annotation of native enhancers with a cas9-histone demethylase fusion. Nat. Methods 12, 401–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (44).Cano-Rodriguez D, Gjaltema RA, Jilderda LJ, Jellema P, Dokter-Fokkens J, Ruiters MH, and Rots MG (2016) Writing of h3k4me3 overcomes epigenetic silencing in a sustained but context-dependent manner. Nat. Commun 7, 12284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (45).Hilton IB, D’Ippolito AM, Vockley CM, Thakore PI, Crawford GE, Reddy TE, and Gersbach CA (2015) Epigenome editing by a crispr-cas9-based acetyltransferase activates genes from promoters and enhancers. Nat. Biotechnol 33, 510–517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (46).Kwon DY, Zhao YT, Lamonica JM, and Zhou Z (2017) Locus-specific histone deacetylation using a synthetic crispr-cas9-based hdac. Nat. Commun 8, 15315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (47).Chen B, Gilbert LA, Cimini BA, Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS, Qi LS, and Huang B (2013) Dynamic imaging of genomic loci in living human cells by an optimized crispr/cas system. Cell 155, 1479–1491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (48).Ma H, Naseri A, Reyes-Gutierrez P, Wolfe SA, Zhang S, and Pederson T (2015) Multicolor crispr labeling of chromosomal loci in human cells. Proc. Natl. Acad. Sci. U. S. A 112, 3002–3007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (49).Fujita T, and Fujii H (2013) Efficient isolation of specific genomic regions and identification of associated proteins by engineered DNA-binding molecule-mediated chromatin immunoprecipitation (enchip) using crispr. Biochem. Biophys. Res. Commun 439, 132–136. [DOI] [PubMed] [Google Scholar]
- (50).Komor AC, Badran AH, and Liu DR (2018) Editing the genome without double-stranded DNA breaks. ACS Chem. Biol 13, 383–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (51).Rees HA, and Liu DR (2018) Base editing: Precision chemistry on the genome and transcriptome of living cells. Nat. Rev. Genet 19, 770–788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (52).Davis KM, Pattanayak V, Thompson DB, Zuris JA, and Liu DR (2015) Small molecule-triggered cas9 protein with improved genome-editing specificity. Nat. Chem. Biol 11, 316–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (53).Frock RL, Hu J, Meyers RM, Ho YJ, Kii E, and Alt FW (2015) Genome-wide detection of DNA double-stranded breaks induced by engineered nucleases. Nat. Biotechnol 33, 179–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (54).Fu Y, Foden JA, Khayter C, Maeder ML, Reyon D, Joung JK, and Sander JD (2013) High-frequency off-target mutagenesis induced by crispr-cas nucleases in human cells. Nat. Biotechnol 31, 822–826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (55).Hsu PD, Scott DA, Weinstein JA, Ran FA, Konermann S, Agarwala V, Li Y, Fine EJ, Wu X, Shalem O, Cradick TJ, Marraffini LA, Bao G, and Zhang F (2013) DNA targeting specificity of rna-guided cas9 nucleases. Nat. Biotechnol 31, 827–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (56).Maji B, Moore CL, Zetsche B, Volz SE, Zhang F, Shoulders MD, and Choudhary A (2017) Multidimensional chemical control of crispr-cas9. Nat. Chem. Biol 13, 9–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (57).Pattanayak V, Guilinger JP, and Liu DR (2014) Determining the specificities of talens, cas9, and other genome-editing enzymes. Methods Enzymol 546, 47–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (58).Pattanayak V, Lin S, Guilinger JP, Ma E, Doudna JA, and Liu DR (2013) High-throughput profiling of off-target DNA cleavage reveals rna-programmed cas9 nuclease specificity. Nat. Biotechnol 31, 839–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (59).Tsai SQ, Zheng Z, Nguyen NT, Liebers M, Topkar VV, Thapar V, Wyvekens N, Khayter C, Iafrate AJ, Le LP, Aryee MJ, and Joung JK (2015) Guide-seq enables genome-wide profiling of off-target cleavage by crispr-cas nucleases. Nat. Biotechnol 33, 187–197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (60).Shin J, Jiang F, Liu JJ, Bray NL, Rauch BJ, Baik SH, Nogales E, Bondy-Denomy J, Corn JE, and Doudna JA (2017) Disabling cas9 by an anti-crispr DNA mimic. Sci. Adv 3, No. e1701620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (61).Yen S-T, Zhang M, Deng JM, Usman SJ, Smith CN, Parker-Thornburg J, Swinton PG, Martin JF, and Behringer RR (2014) Somatic mosaicism and allele complexity induced by crispr/cas9 rna injections in mouse zygotes. Dev. Biol 393, 3–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (62).Tu Z, Yang W, Yan S, Yin A, Gao J, Liu X, Zheng Y, Zheng J, Li Z, Yang S, Li S, Guo X, and Li X-J (2017) Promoting cas9 degradation reduces mosaic mutations in non-human primate embryos. Sci. Rep 7, 42081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (63).Wang H, Yang H, Shivalila CS, Dawlaty MM, Cheng AW, Zhang F, and Jaenisch R (2013) One-step generation of mice carrying mutations in multiple genes by crispr/cas-mediated genome engineering. Cell 153, 910–918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (64).Mehravar M, Shirazi A, Nazari M, and Banan M (2019) Mosaicism in crispr/cas9-mediated genome editing. Dev. Biol 445, 156. [DOI] [PubMed] [Google Scholar]
- (65).Gonzalez F, Zhu Z, Shi ZD, Lelli K, Verma N, Li QV, and Huangfu D (2014) An icrispr platform for rapid, multiplexable, and inducible genome editing in human pluripotent stem cells. Cell stem cell 15, 215–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (66).Dow LE, Fisher J, O’Rourke KP, Muley A, Kastenhuber ER, Livshits G, Tschaharganeh DF, Socci ND, and Lowe SW (2015) Inducible in vivo genome editing with crispr-cas9. Nat. Biotechnol 33, 390–394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (67).Kiani S, Beal J, Ebrahimkhani MR, Huh J, Hall RN, Xie Z, Li Y, and Weiss R (2014) Crispr transcriptional repression devices and layered circuits in mammalian cells. Nat. Methods 11, 723–726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (68).Aubrey BJ, Kelly GL, Kueh AJ, Brennan MS, O’Connor L, Milla L, Wilcox S, Tai L, Strasser A, and Herold MJ (2015) An inducible lentiviral guide rna platform enables the identification of tumor-essential genes and tumor-promoting mutations in vivo. Cell Rep 10, 1422–1432. [DOI] [PubMed] [Google Scholar]
- (69).Davis KM, Pattanayak V, Thompson DB, Zuris JA, and Liu DR (2015) Small molecule-triggered cas9 protein with improved genome-editing specificity. Nat. Chem. Biol 11, 316–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (70).Buskirk AR, and Liu DR (2005) Creating small-molecule-dependent switches to modulate biological functions. Chem. Biol 12, 151–161. [DOI] [PubMed] [Google Scholar]
- (71).Rakhit R, Navarro R, and Wandless TJ (2014) Chemical biology strategies for posttranslational control of protein function. Chem. Biol 21, 1238–1252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (72).Liu KI, Ramli MN, Woo CW, Wang Y, Zhao T, Zhang X, Yim GR, Chong BY, Gowher A, Chua MZ, Jung J, Lee JH, and Tan MH (2016) A chemical-inducible crispr-cas9 system for rapid control of genome editing. Nat. Chem. Biol 12, 980–987. [DOI] [PubMed] [Google Scholar]
- (73).Oakes BL, Nadler DC, Flamholz A, Fellmann C, Staahl BT, Doudna JA, and Savage DF (2016) Profiling of engineering hotspots identifies an allosteric crispr-cas9 switch. Nat. Biotechnol 34, 646–651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (74).Fegan A, White B, Carlson JC, and Wagner CR (2010) Chemically controlled protein assembly: Techniques and applications. Chem. Rev 110, 3315–3336. [DOI] [PubMed] [Google Scholar]
- (75).DeRose R, Miyamoto T, and Inoue T (2013) Manipulating signaling at will: Chemically-inducible dimerization (cid) techniques resolve problems in cell biology. Pfluegers Arch 465, 409–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (76).Zetsche B, Volz SE, and Zhang F (2015) A split-cas9 architecture for inducible genome editing and transcription modulation. Nat. Biotechnol 33, 139–142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (77).Lo A, and Qi L (2017) Genetic and epigenetic control of gene expression by crispr-cas systems. F1000Research 6, 747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (78).Gao Y, Xiong X, Wong S, Charles EJ, Lim WA, and Qi LS (2016) Complex transcriptional modulation with orthogonal and inducible dcas9 regulators. Nat. Methods 13, 1043–1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (79).Miyamoto T, DeRose R, Suarez A, Ueno T, Chen M, Sun T, Wolfgang MJ, Mukherjee C, Meyers DJ, and Inoue T (2012) Rapid and orthogonal logic gating with a gibberellin-induced dimerization system. Nat. Chem. Biol 8, 465–470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (80).Liang FS, Ho WQ, and Crabtree GR (2011) Engineering the aba plant stress pathway for regulation of induced proximity. Sci. Signaling 4, rs2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (81).Bao Z, Jain S, Jaroenpuntaruk V, and Zhao H (2017) Orthogonal genetic regulation in human cells using chemically induced crispr/cas9 activators. ACS Synth. Biol 6, 686–693. [DOI] [PubMed] [Google Scholar]
- (82).Rose JC, Huang PS, Camp ND, Ye J, Leidal AM, Goreshnik I, Trevillian BM, Dickinson MS, Cunningham-Bryant D, Debnath J, Baker D, Wolf-Yadlin A, and Maly DJ (2017) A computationally engineered ras rheostat reveals ras-erk signaling dynamics. Nat. Chem. Biol 13, 119–126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (83).Rose JC, Stephany JJ, Valente WJ, Trevillian BM, Dang HV, Bielas JH, Maly DJ, and Fowler DM (2017) Rapidly inducible cas9 and dsb-ddpcr to probe editing kinetics. Nat. Methods 14, 891–896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (84).Shoemaker AR, Oleksijew A, Bauch J, Belli BA, Borre T, Bruncko M, Deckwirth T, Frost DJ, Jarvis K, Joseph MK, Marsh K, McClellan W, Nellans H, Ng S, Nimmer P, O’Connor JM, Oltersdorf T, Qing W, Shen W, Stavropoulos J, Tahir SK, Wang B, Warner R, Zhang H, Fesik SW, Rosenberg SH, and Elmore SW (2006) A small-molecule inhibitor of bcl-xl potentiates the activity of cytotoxic drugs in vitro and in vivo. Cancer Res 66, 8731–8739. [DOI] [PubMed] [Google Scholar]
- (85).Natsume T, and Kanemaki MT (2017) Conditional degrons for controlling protein expression at the protein level. Annu. Rev. Genet 51, 83–102. [DOI] [PubMed] [Google Scholar]
- (86).Senturk S, Shirole NH, Nowak DG, Corbo V, Pal D, Vaughan A, Tuveson DA, Trotman LC, Kinney JB, and Sordella R (2017) Rapid and tunable method to temporally control gene editing based on conditional cas9 stabilization. Nat. Commun 8, 14370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (87).Kleinjan DA, Wardrope C, Nga Sou S, and Rosser SJ (2017) Drug-tunable multidimensional synthetic gene control using inducible degron-tagged dcas9 effectors. Nat. Commun 8, 1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (88).Jacobs CL, Badiee RK, and Lin MZ (2018) Stapls: Versatile genetically encoded modules for engineering drug-inducible proteins. Nat. Methods 15, 523–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (89).Tague EP, Dotson HL, Tunney SN, Sloas DC, and Ngo JT (2018) Chemogenetic control of gene expression and cell signaling with antiviral drugs. Nat. Methods 15, 519–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (90).Kundert K, Lucas JE, Watters KE, Fellmann C, Ng AH, Heineike BM, Fitzsimmons CM, Oakes BL, Savage DF, El-Samad H, Doudna JA, and Kortemme T (2018) Controlling crispr-cas9 with ligand-activated and ligand-deactivated sgrnas. bioRxiv, n/a [DOI] [PMC free article] [PubMed]
- (91).Liu Y, Zhan Y, Chen Z, He A, Li J, Wu H, Liu L, Zhuang C, Lin J, Guo X, Zhang Q, Huang W, and Cai Z (2016) Directing cellular information flow via crispr signal conductors. Nat. Methods 13, 938–944. [DOI] [PubMed] [Google Scholar]
- (92).Tang W, Hu JH, and Liu DR (2017) Aptazyme-embedded guide rnas enable ligand-responsive genome editing and transcriptional activation. Nat. Commun 8, 15939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (93).Pu J, Kentala K, and Dickinson BC (2018) Multidimensional control of cas9 by evolved rna polymerase-based biosensors. ACS Chem. Biol 13, 431–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (94).Nihongaki Y, Yamamoto S, Kawano F, Suzuki H, and Sato M (2015) Crispr-cas9-based photoactivatable transcription system. Chem. Biol 22, 169. [DOI] [PubMed] [Google Scholar]
- (95).Polstein LR, and Gersbach CA (2015) A light-inducible crispr-cas9 system for control of endogenous gene activation. Nat. Chem. Biol 11, 198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (96).Nihongaki Y, Kawano F, Nakajima T, and Sato M (2015) Photoactivatable crispr-cas9 for optogenetic genome editing. Nat. Biotechnol 33, 755–760. [DOI] [PubMed] [Google Scholar]
- (97).Nihongaki Y, Otabe T, and Sato M (2018) Emerging approaches for spatiotemporal control of targeted genome with inducible crispr-cas9. Anal. Chem 90, 429–439. [DOI] [PubMed] [Google Scholar]
- (98).Zhou XX, Zou X, Chung HK, Gao Y, Liu Y, Qi LS, and Lin MZ (2018) A single-chain photoswitchable crispr-cas9 architecture for light-inducible gene editing and transcription. ACS Chem. Biol 13, 443–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (99).Hemphill J, Borchardt EK, Brown K, Asokan A, and Deiters A (2015) Optical control of crispr/cas9 gene editing. J. Am. Chem. Soc 137, 5642–5645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (100).Jain PK, Ramanan V, Schepers AG, Dalvie NS, Panda A, Fleming HE, and Bhatia SN (2016) Development of light-activated crispr using photocleavable protectors of guide rnas. Angew. Chem., Int. Ed 55, 12440–12444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (101).Marshall JM, and Akbari OS (2018) Can crispr-based gene drive be confined in the wild? A question for molecular and population biology. ACS Chem. Biol 13, 424–430. [DOI] [PubMed] [Google Scholar]
- (102).Gootenberg JS, Abudayyeh OO, Kellner MJ, Joung J, Collins JJ, and Zhang F (2018) Multiplexed and portable nucleic acid detection platform with cas13, cas12a, and csm6. Science 360, 439–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (103).Myhrvold C, Freije CA, Gootenberg JS, Abudayyeh OO, Metsky HC, Durbin AF, Kellner MJ, Tan AL, Paul LM, Parham LA, Garcia KF, Barnes KG, Chak B, Mondini A, Nogueira ML, Isern S, Michael SF, Lorenzana I, Yozwiak NL, MacInnis BL, Bosch I, Gehrke L, Zhang F, and Sabeti PC (2018) Field-deployable viral diagnostics using crispr-cas13. Science 360, 444–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (104).Hille F, Richter H, Wong SP, Bratovic M, Ressel S, and Charpentier E (2018) The biology of crispr-cas: Backward and forward. Cell 172, 1239–1259. [DOI] [PubMed] [Google Scholar]
- (105).Cox DB, Gootenberg JS, Abudayyeh OO, Franklin B, Kellner MJ, Joung J, and Zhang F (2017) Rna editing with crispr-cas13. Science 358, 1019–1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (106).Granados-Riveron JT, and Aquino-Jarquin G (2018) Crispr-cas13 precision transcriptome engineering in cancer. Cancer Res 78, 4107–4113. [DOI] [PubMed] [Google Scholar]
- (107).Pawluk A, Davidson AR, and Maxwell KL (2017) Anti-crispr: Discovery, mechanism and function. Nat. Rev. Microbiol 16, 12–17. [DOI] [PubMed] [Google Scholar]
- (108).Bondy-Denomy J (2018) Protein inhibitors of crispr-cas9. ACS Chem. Biol 13, 417–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (109).Maxwell KL (2017) The anti-crispr story: A battle for survival. Mol. Cell 68, 8–14. [DOI] [PubMed] [Google Scholar]
- (110).Dong C, Hao GF, Hua HL, Liu S, Labena AA, Chai G, Huang J, Rao N, and Guo FB (2018) Anti-crisprdb: A comprehensive online resource for anti-crispr proteins. Nucleic Acids Res 46, D393–D398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (111).Rauch BJ, Silvis MR, Hultquist JF, Waters CS, McGregor MJ, Krogan NJ, and Bondy-Denomy J (2017) Inhibition of crispr-cas9 with bacteriophage proteins. Cell 168, 150–158.e110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (112).Bondy-Denomy J, Garcia B, Strum S, Du M, Rollins MF, Hidalgo-Reyes Y, Wiedenheft B, Maxwell KL, and Davidson AR (2015) Multiple mechanisms for crispr-cas inhibition by anti-crispr proteins. Nature 526, 136–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (113).Dong D, Guo M, Zhu Y, Wang S, Wang S, Xiong Z, Yang J, Xu Z, and Huang Z (2017) Structural basis of crispr-spycas9 inhibition by an anti-crispr protein. Nature 546, 436–439. [DOI] [PubMed] [Google Scholar]
- (114).Yang H, and Patel DJ (2017) Inhibition mechanism of an anti-crispr suppressor acriia4 targeting spycas9. Mol. Cell 67, 117–127.e115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (115).Harrington LB, Doxzen KW, Ma E, Liu JJ, Knott GJ, Edraki A, Garcia B, Amrani N, Chen JS, Cofsky JC, Kranzusch PJ, Sontheimer EJ, Davidson AR, Maxwell KL, and Doudna JA (2017) A broad-spectrum inhibitor of crispr-cas9. Cell 170, 1224–1233.e1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (116).Bondy-Denomy J, Pawluk A, Maxwell KL, and Davidson AR (2012) Bacteriophage genes that inactivate the crispr/cas bacterial immune system. Nature 493, 429–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (117).Pawluk A, Amrani N, Zhang Y, Garcia B, Hidalgo-Reyes Y, Lee J, Edraki A, Shah M, Sontheimer EJ, Maxwell KL, and Davidson AR (2016) Naturally occurring off-switches for crispr-cas9. Cell 167, 1829–1838.e1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (118).Pawluk A, Staals RH, Taylor C, Watson BN, Saha S, Fineran PC, Maxwell KL, and Davidson AR (2016) Inactivation of crispr-cas systems by anti-crispr proteins in diverse bacterial species. Nat. Microbiol 1, 16085. [DOI] [PubMed] [Google Scholar]
- (119).Marino ND, Zhang JY, Borges AL, Sousa AA, Leon LM, Rauch BJ, Walton RT, Berry JD, Joung JK, Kleinstiver BP, and Bondy-Denomy J (2018) Discovery of widespread type i and type v crispr-cas inhibitors. Science 362, 240–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (120).Watters KE, Fellmann C, Bai HB, Ren SM, and Doudna JA (2018) Systematic discovery of natural crispr-cas12a inhibitors. Science 362, 236–239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (121).Haapaniemi E, Botla S, Persson J, Schmierer B, and Taipale J (2018) Crispr-cas9 genome editing induces a p53-mediated DNA damage response. Nat. Med 24, 927–930. [DOI] [PubMed] [Google Scholar]
- (122).Li C, Psatha N, Gil S, Wang H, Papayannopoulou T, and Lieber A (2018) Hdad5/35(++) adenovirus vector expressing anti-crispr peptides decreases crispr/cas9 toxicity in human hematopoietic stem cells. Mol. Ther.–Methods Clin. Dev 9, 390–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (123).Shin J, Jiang F, Liu J-J, Bray NL, Rauch BJ, Baik SH, Nogales E, Bondy-Denomy J, Corn JE, and Doudna JA (2017) Disabling cas9 by an anti-crispr DNA mimic. Sci. Adv 3, No. e1701620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (124).Weiss WA, Taylor SS, and Shokat KM (2007) Recognizing and exploiting differences between rnai and small-molecule inhibitors. Nat. Chem. Biol 3, 739–744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (125).Nishimasu H, Ran FA, Hsu PD, Konermann S, Shehata SI, Dohmae N, Ishitani R, Zhang F, and Nureki O (2014) Crystal structure of cas9 in complex with guide rna and target DNA. Cell 156, 935–949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (126).Sternberg SH, Redding S, Jinek M, Greene EC, and Doudna JA (2014) DNA interrogation by the crispr rna-guided endonuclease cas9. Nature 507, 62–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (127).Koehler AN (2010) A complex task? Direct modulation of transcription factors with small molecules. Curr. Opin. Chem. Biol 14, 331–340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (128).Esvelt KM, Smidler AL, Catteruccia F, and Church GM (2014) Concerning rna-guided gene drives for the alteration of wild populations. eLife 3, No. e03401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (129).Neve RL, Neve KA, Nestler EJ, and Carlezon WA Jr, (2005) Use of herpes virus amplicon vectors to study brain disorders. BioTechniques 39, 381–391. [DOI] [PubMed] [Google Scholar]
- (130).Zincarelli C, Soltys S, Rengo G, and Rabinowitz JE (2008) Analysis of aav serotypes 1−9 mediated gene expression and tropism in mice after systemic injection. Mol. Ther 16, 1073–1080. [DOI] [PubMed] [Google Scholar]
- (131).Wegrzyn RD, Lee AH, Jenkins AL, Stoddard CD, and Cheever AE (2018) Genome editing: Insights from chemical biology to support safe and transformative therapeutic applications. ACS Chem. Biol 13, 333. [DOI] [PubMed] [Google Scholar]
- (132).Banaszynski LA, and Wandless TJ (2006) Conditional control of protein function. Chem. Biol 13, 11–21. [DOI] [PubMed] [Google Scholar]
- (133).Raina K, and Crews CM (2010) Chemical inducers of targeted protein degradation. J. Biol. Chem 285, 11057–11060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (134).Bondeson DP, and Crews CM (2017) Targeted protein degradation by small molecules. Annu. Rev. Pharmacol. Toxicol 57, 107–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (135).Gu S, Cui D, Chen X, Xiong X, and Zhao Y (2018) Protacs: An emerging targeting technique for protein degradation in drug discovery. BioEssays 40, No. 1700247. [DOI] [PubMed] [Google Scholar]
- (136).Neklesa TK, Winkler JD, and Crews CM (2017) Targeted protein degradation by protacs. Pharmacol. Ther 174, 138–144. [DOI] [PubMed] [Google Scholar]
- (137).Nishimura K, Fukagawa T, Takisawa H, Kakimoto T, and Kanemaki M (2009) An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nat. Methods 6, 917–922. [DOI] [PubMed] [Google Scholar]
- (138).Bonger KM, Chen L, Liu CW, and Wandless TJ (2011) Small molecule displacement of a cryptic degron causes conditional protein degradation. Nat. Chem. Biol 7, 531–537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (139).Chung HK, Jacobs CL, Huo Y, Yang J, Krumm SA, Plemper RK, Tsien RY, and Lin MZ (2015) Tunable and reversible drug control of protein production via a self-excising degron. Nat. Chem. Biol 11, 713–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (140).Bonger KM, Rakhit R, Payumo AY, Chen JK, and Wandless TJ (2014) A general method for regulating protein stability with light. ACS Chem. Biol 9, 111–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (141).Matsuzawa S, Cuddy M, Fukushima T, and Reed JC (2005) Method for targeting protein destruction by using a ubiquitin-independent, proteasome-mediated degradation pathway. Proc. Natl. Acad. Sci. U. S. A 102, 14982–14987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (142).Coffino P (2001) Regulation of cellular polyamines by antizyme. Nat. Rev. Mol. Cell Biol 2, 188–194. [DOI] [PubMed] [Google Scholar]
- (143).Nabet B, Roberts JM, Buckley DL, Paulk J, Dastjerdi S, Yang A, Leggett AL, Erb MA, Lawlor MA, Souza A, Scott TG, Vittori S, Perry JA, Qi J, Winter GE, Wong KK, Gray NS, and Bradner JE (2018) The dtag system for immediate and target-specific protein degradation. Nat. Chem. Biol 14, 431–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (144).Lai AC, and Crews CM (2017) Induced protein degradation: An emerging drug discovery paradigm. Nat. Rev. Drug Discovery 16, 101–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (145).Charlesworth CT, Deshpande PS, Dever DP, Dejene B, Gomez-Ospina N, Mantri S, Pavel-Dinu M, Camarena J, Weinberg KI, and Porteus MH (2018) Identification of pre-existing adaptive immunity to cas9 proteins in humans. bioRxiv, n/a
- (146).Ihry RJ, Worringer KA, Salick MR, Frias E, Ho D, Theriault K, Kommineni S, Chen J, Sondey M, Ye C, Randhawa R, Kulkarni T, Yang Z, McAllister G, Russ C, Reece-Hoyes J, Forrester W, Hoffman GR, Dolmetsch R, and Kaykas A (2018) P53 inhibits crispr-cas9 engineering in human pluripotent stem cells. Nat. Med 24, 939–946. [DOI] [PubMed] [Google Scholar]
- (147).Haapaniemi E, Botla S, Persson J, Schmierer B, and Taipale J (2018) Crispr-cas9 genome editing induces a p53-mediated DNA damage response. Nat. Med 24, 927–930. [DOI] [PubMed] [Google Scholar]