

Mothers transfer immune cells via breastfeeding to provide offspring with long-term protection from parasitic infection.
Abstract
Maternal immune transfer is the most significant source of protection from early-life infection, but whether maternal transfer of immunity by nursing permanently alters offspring immunity is poorly understood. Here, we identify maternal immune imprinting of offspring nursed by mothers who had a pre-conception helminth infection. Nursing of pups by helminth-exposed mothers transferred protective cellular immunity to these offspring against helminth infection. Enhanced control of infection was not dependent on maternal antibody. Protection associated with systemic development of protective type 2 immunity in T helper 2 (TH2) impaired IL-4Rα−/− offspring. This maternally acquired immunity was maintained into maturity and required transfer (via nursing) to the offspring of maternally derived TH2-competent CD4 T cells. Our data therefore reveal that maternal exposure to a globally prevalent source of infection before pregnancy provides long-term nursing-acquired immune benefits to offspring mediated by maternally derived pathogen-experienced lymphocytes.
INTRODUCTION
Maternal transfer of immunity both in utero and via nursing provides critical sources of early-life immune education and protection from disease. Maternally acquired protection from infection is typically associated with a passive transfer to offspring of maternal innate opsonins and antibody, which provide a transient, but critical, early-life ability to counter pathogens (1, 2).
Nursing alone provides important protection to offspring against both infectious and noninfectious diseases (3, 4). While this protection is associated primarily with transfer of maternally derived antibody (5), other immunogenic components of breast milk such as cytokines and antigen can also influence offspring immunity (6). Epidemiological and experimental evidence indicates that maternal immune transfer via nursing may also provide long-lasting pathogen-specific protection from infection, despite the short half-life of substances transferred via breast milk (7, 8). However, the mechanisms underlying any such long-term protection are not well defined, and the contribution from incorporation and maintenance of maternal components by offspring has hitherto been relatively unaddressed (9).
Helminth infections are an important cause of infection and disease and leave profound immunological footprints on a host (9, 10). These infections are extremely common and have important effects on multiple components of host immunity. Evidence supports the importance of parasite-host coevolution for optimal function and regulation of host immunity (11, 12). Helminth infections before and during pregnancy are known to have significant consequences for offspring. For example, maternal infection with the nematode Heligmosomoides polygyrus (Hp) provides high levels of nursing-acquired antibody that mediates protection to offspring against this infection (13). Maternal helminth infections also have bystander effects, either driving susceptibility to or protection from allergy depending on the progression of, in this case, a maternal Schistosoma mansoni infection (14).
Clinical studies also confirm the significant influence of maternal helminth exposure on a child’s immune system. For example, children whose mothers were infected with helminths during pregnancy display populations of lymphocytes responsive to helminth antigens (15); mothers infected with helminths during pregnancy can also confer protection to their children against eczema (16). The mechanisms underlying these maternally associated effects in offspring are, however, not well defined. As helminth exposure is a huge global coordinator of our immunological footprint, understanding this influence in the context of the maternal-offspring immunological relationship is important (9). This transgenerational immunological relationship is pivotal for our understanding of how early-life events influence lifelong ability to control disease.
In this study, we address how exposure to and resolution of a helminth infection before pregnancy influences offspring cellular immunity to this infection. To address this question, we developed mouse models of maternal helminth infection effects on offspring control of infection. This approach allowed us to test how pre-conception maternal infection with the nematode Nippostrongylus brasiliensis (Nb) influenced offspring ability to control an Nb infection both early in life and when mature. Our model allowed us to identify (i) the critical importance of nursing in transfer of protective immunity, (ii) that offspring-acquired protection was cellular and not mediated by antibody, (iii) that protection could correct germline susceptibility to infection, (iv) that protection persisted into adulthood, and (v) that this protection was maintained in an allogeneic setting and (vi) required the dissemination via nursing of maternally derived T helper 2 (TH2)–competent CD4 T cells throughout the body. Our findings therefore demonstrate that maternal transfer and offspring incorporation of nursing-derived pathogen-experienced lymphocytes can provide long-lasting immunity to infection.
RESULTS
Pre-conception Nb infection transfers protective immunity against Nb infection to offspring via nursing
To identify how exposure to and resolution of a helminth infection before pregnancy influenced offspring immunity, we modeled this scenario using the murine helminth parasite Nb. Here, female BALB/c mice were infected with Nb and then treated with an anti-helminth drug 7 days later. Two weeks after treatment, these female mice were mated with naïve males, and offspring immune responses were analyzed (Fig. 1A). Total cell numbers and CD4+ T cell numbers in the spleens of a 14-day-old offspring born to Nb-exposed mothers (NbM) and those born to naïve mothers (NvM) were similar (Fig. 1, B and C), but the numbers of activated effector CD4+ T cells in NbM offspring were raised when compared to NvM offspring (Fig. 1C).
Fig. 1. Maternal helminth infection confers systemic protective immunity against Nb to pups via nursing.
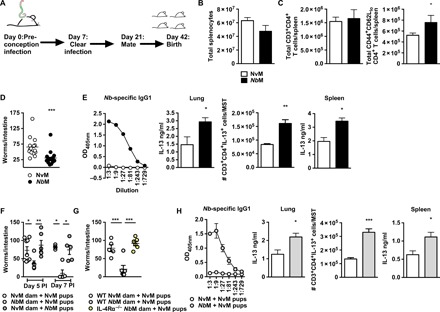
(A) Female BALB/c mice were infected with 500 × Nb L3 and, 7 days PI, were cleared of Nb by a 5-day oral treatment with ivermectin; at day 21 PI, mice were mated and immunity in subsequent pups was established at various times after birth and/or Nb infection. (B) Total splenocyte cellularity in 14-day-old offspring. (C) Total numbers of CD+CD4+ T cells and activated T effector CD4 T cell populations (CD3+C0D4+CD44+CD62Llo) in 14-day-old offspring. (D) Intestinal worm burdens at day 5 PI of pups born to NvM or NbM mothers and infected with 250 × Nb L3 when 14 days old. (E) NvM and NbM offspring anti-CD3 restimulated splenocyte IL-13 secretion, lung tissue homogenate levels of IL-13 and IL-13+CD3+CD4+ T cells/MST, and Nb-specific serum immunoglobulin G1 (IgG1) at day 5 after Nb infection. OD405nm, optical density at 405 nm. (F) Worm burdens of NvM and NbM offspring infected when 14 days old and euthanized at day 5 PI following fostering on NvM or NbM dams at 3 days old. (G) Worm burdens of NvM and NbM offspring infected when 14 days old and euthanized at day 5 PI following fostering on WT or IL-4Rα−/− NvM and NbM dams. (H) NvM pups fostered by either NvM or NbM anti-CD3 restimulated splenocyte IL-13 secretion, lung tissue homogenate levels of IL-13 and IL-13+CD3+CD4+ T cells/MST, and Nb-specific serum IgG1 at day 5 after Nb infection. All data are representative of a minimum of two experimental repeats. *P < 0.05, **P < 0.01, and ***P < 0.001.
Increased T cell activation suggested that NbM offspring may acquire an enhanced ability to control Nb infection. We therefore tested whether NbM transferred an enhanced interleukin-4 receptor α (IL-4Rα)–driven type 2 immunity against Nb to their offspring. Reduced intestinal worm burdens were found at day 5 post-infection (PI) in 19-day-old NbM pups in comparison to NvM pups (Fig. 1D). In addition to raised anti-Nb immunoglobulin G1 (IgG1) in NbM pups (Fig. 1E), we also found enhanced levels of the TH2 cytokine IL-13 (essential for resolving Nb infection) in the lung, mediastinal lymph nodes (MST), and spleen, demonstrating that maternal transfer of immunity resulted in a systemic TH2 priming in NbM offspring reflective of acquired host-protective pulmonary TH2 immunity to Nb (17).
We next investigated whether protection in NbM offspring was conferred in utero or via nursing. Here, we fostered 3-day-old pups born to NvM or NbM onto either NbM or NvM foster dams. Subsequent infection of fostered pups resulted in pups nursed by NbM mothers having reduced worm burdens 5 days PI and displaying early clearance of Nb by day 7 PI when compared to pups nursed on NvM (Fig. 1F). This protection was dependent on maternal TH2 competency: Wild-type (WT) offspring nursed by IL-4Rα−/− NbM did not display reduced intestinal worm burdens when compared to WT offspring nursed by WT NbM (Fig. 1G). Analysis of nursing-dependent transfer of protection in pups born to WT NvM but nursed on WT NbM also revealed both elevated Nb-specific IgG1 and systemically raised IL-13 levels associated with enhanced maternal transfer of protection from infection (Fig. 1H).
Nursing-derived protection is dependent on maternal B cells but not antibody
Nursing-derived protective immunity in offspring is often attributed to passive antibody transfer (18). Our findings also demonstrated associations between raised Nb-specific antibody in offspring and enhanced protection from Nb. Moreover, milk from NbM had higher Nb-specific IgG1 titers than milk from NvM (Fig. 2A). However, existing literature has shown that immunity to Nb is largely antibody independent (10, 19). Nevertheless, studies using other helminth infection models show that transfer of serum from infected mice to pups confers high levels of protection to offspring (13). In support of antibody being redundant in providing maternally derived protection in the case of Nb infection, we found that transfer of NbM serum to NvM pups did not lead to a significant reduction in worm burdens when compared to recipients of NvM serum (Fig. 2B). In addition, WT NvM pups were nursed on IgMi mice that have a functional B cell receptor but do not secrete antibody (20). Here, NvM pups nursed by IgMi NbM and WT NbM mothers showed equivalent reductions in worm burdens that were independent of Nb-specific IgG1 antibody titers in either the pups or mother (Fig. 2C). Together, these data show that transfer of maternally derived antibody is not essential to the transfer of protection against Nb to offspring.
Fig. 2. Protection in NbM pups is maternal B cell dependent but not antibody dependent.
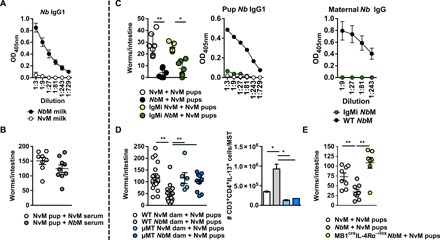
(A) Nb-specific IgG1 levels in breast milk taken from the stomach of pups nursed by NvM or NbM. (B) Intestinal worm burdens at day 5 PI of NvM offspring infected at 14 days old with 250 × Nb L3 following intravenous administration of 150 μl of heat-inactivated serum from either NvM or NbM. (C) Intestinal worm burdens and Nb-specific serum IgG1 levels at day 5 PI of NvM offspring infected at 14 days old with 250 × Nb L3 after being fostered on either WT and IgMi NbM mice or WT and IgMi maternal Nb-specific serum IgG1 levels. (D) Three-day-old NvM pups were fostered on WT NvM/NbM or μMT NvM/NbM dams. Pups were infected when 14 days old with 250 × Nb L3 and euthanized at day 5 PI. Offspring intestinal worm burden was analyzed, and MST IL-13+CD3+CD4+ T cells were quantified at day 5 PI. (E) Intestinal worm burdens at day 5 PI of NvM offspring infected at 14 days old with 250 × Nb L3 after being fostered on either WT or B cell–IL-4Rα–deficient (MB1creIL-4Rα−/lox) dams. All data are representative of a minimum of two experimental repeats. *P < 0.05 and **P < 0.01.
While antibody is not directly important in immunity to Nb, B effector cell cytokine responses and antigen presentation have been shown to contribute to CD4 T cell–driven recall immunity to Nb in an IL-4Rα–dependent manner (10). We nursed WT NvM pups on WT and μMT NvM and NbM dams and found that no protection was transferred to pups nursed by μMT NbM dams (Fig. 2D). Associated with this reduced protection were significantly reduced CD4+IL-13+ populations in offspring nursed by μMT NbM dams when compared to protected pups nursed by WT NbM (Fig. 2D). This effect was also dependent on maternal B cells being type 2 competent. Pups nursed on NbM dams deficient in IL-4Rα specifically on B cells (MB1creIL-4Rα−/lox) had significantly higher worm burdens than pups nursed by WT NbM mothers (Fig. 2E). These data show that a maternal B cell response reflective of B effector cell–dependent activation of CD4 T cells contributes to NbM dam–derived protection in nursed pups.
Transfer of protection from NbM is long term and associated with a systemic distribution of maternal TH2 CD4 T cells in the offspring
The systemic profile of raised TH2 CD4 T cell–associated immunity in NbM nursed pups when compared to NvM nursed pups is a striking effect on pup immunity. As milk contains significant populations of immune cells, we hypothesized that these cells may be important contributors to this enhanced anti-Nb immunity in NbM-nursed offspring. This could be affected by transferred Nb-experienced maternal CD4 T cells systemically influencing offspring protection derived from NbM dams. Moreover, such a systemic effect on immune readouts of protection associated with CD4 T cells may also suggest that the effect could result in transfer of immune memory that could persist into adulthood in NbM foster pups. Direct input from passively transferred maternal antibody would be minimal in this adult setting, as transferred antibody eventually degrades after transfer (21). To identify such an effect, naïve Thy1.2 offspring were nursed on either NvM or NbM syngeneic Thy1.1 dams and infected with Nb at 6 to 8 weeks of age (Fig. 3A).
Fig. 3. Offspring acquire, via nursing, persistent protection from Nb related to acquisition of a population of maternally derived CD4 T cells with a heightened TH2 responsiveness following Nb infection.
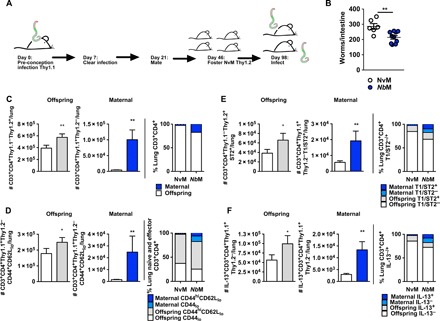
(A) Three-day-old NvM Thy1.2+ pups were fostered on Thy1.1+ WT NvM/NbM dams. Eight-week-old NvM Thy1.2+ mice that had been fostered on either NvM or NbM Thy1.1+ dams were infected with 500 × Nb L3 and euthanized at day 5 PI. (B) Intestinal worm burdens in offspring at day 5 PI in 8-week-old mice born to either NvM or NbM were established. (C to F) Numbers and proportions of offspring endogenous (Thy1.2+Thy1.1−) and maternally derived (Thy1.1+Thy1.2−) lung (C) Thy1+CD3+CD4+, (D) CD44+CD62Llo, (E) T1/ST2+, and (F) IL-13+ expressing T cell populations. All data are representative of a minimum of two experimental repeats. *P < 0.05 and **P < 0.01.
Intestinal worm burdens at day 5 PI were significantly reduced in offspring nursed by NbM dams compared to those nursed by NvM dams (Fig. 3B). This long-term maintenance of maternally acquired immunity is strongly associated with elevated numbers and proportions of both endogenous and maternally derived TH2-competent CD4 T cells in both secondary and peripheral immune sites associated with adaptive immune protection against Nb infection (10, 17) in NbM offspring (Fig. 3, C to F, and fig. S1).
Significantly higher numbers of maternally derived Thy1.1 CD4 T cells were detected in both the lungs (Fig. 3C) and MST (fig. S1) of NbM dam–nursed offspring than in pups nursed by NvM dams. Significantly more maternally derived cells were activated (CD4+CD44+CD62Llo) in NbM than NvM offspring (Fig. 3D). The NbM-derived populations of Thy1.1 CD4 T cells had higher levels of TH2 potential. Greater numbers of NbM dam–derived lung cells expressed the IL-33 receptor T1/ST2 (IL-33 signaling through this receptor can initiate type 2 immunity) when compared to offspring nursed by NvM dams (Fig. 3E). Moreover, higher numbers of NbM dam–derived CD4+ T cells were also IL-13+ (Fig. 3F). This raised type 2 immune potential was also apparent in offspring’s endogenous CD4 T cells in the lung (Fig. 4, C to F) but not the MST (fig. S1). Therefore, a maternal pre-conception Nb infection enabled transfer via nursing to, and systemic integration by, offspring of populations of Nb protective CD4+ T cells. In addition, the presence of these cells is also associated with raised endogenous type 2 protective immunity in the lungs of NbM offspring against Nb. However, pups nursed on mothers exposed to a single intestinally restricted helminth Hp for 17 days before pregnancy did not demonstrate raised populations of maternally derived CD4+ T cells in the mesenteric lymph nodes or lung (fig. S2). This suggests that differences in the host-parasite relationship may influence the ability of the mother to transfer a protective CD4 T cell response.
Fig. 4. Offspring acquire, via nursing, persistent protection from Nb related to raised TH2 CD4 T cell responses following Nb infection in an allogeneic setting.
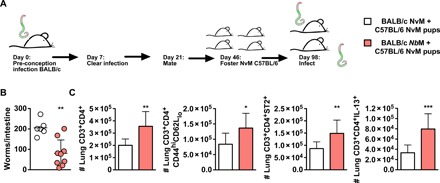
(A) Three-day-old NvM C57BL/6 pups were fostered on BALB/c NvM or WT NbM dams before being infected at 8 weeks old with 500 × Nb L3 and euthanized at day 5 PI. (B) Intestinal worm burdens at day 5 PI in 8-week-old mice born to either NvM or NbM were established. (C) Numbers of lung CD3+CD4+, ST2+, IL-13+, and CD44+CD62Llo expressing T cell populations were quantified. All data are representative of a minimum of two experimental repeats. *P < 0.05, **P < 0.01, and ***P < 0.001.
Therefore, NbM offspring acquired long-term protective maternal immunity that associated with a systemic dissemination of maternally derived TH2-competent CD4 T cells, which significantly enhanced both the overall and endogenous TH2 CD4 T cell compartment of the offspring.
Long-term protection is maintained in an allogeneic setting
To test whether our findings related to transfer of maternal CD4 T cells were relevant in an outbred setting representative of typical maternal-offspring relationships, we nursed naïve C57BL/6 pups on either NvM or NbM BALB/c dams (Fig. 4A) and found that 6- to 8-week-old NbM offspring had significantly reduced Nb burdens in the intestine when compared to NvM offspring (Fig. 4B). This is associated with increased numbers of maternally derived CD4 T cells (CD3+CD4+H2Dd+) in the lung (fig. S3), albeit in reduced numbers when compared to those seen in a major histocompatibility complex (MHC)–matched setting (Fig. 3). In addition, as seen in an MHC-matched setting (Fig. 3), offspring protection in an outbred, MHC-mismatched setting is associated with a raised type 2 CD4 T cell response. Here, NbM offspring displayed significantly increased levels of CD4 T cell activation, expression of ST2, and secretion of IL-13 (Fig. 4C). This effect was also found when NvM and NbM outbred BALB/c × C57BL/6 dams were bred and BALB/c pups were subsequently fostered by them (fig. S4).
Nursing-derived protection corrects susceptibility to Nb in IL-4Rα−/− pups
We tested whether transfer of maternally derived cells via nursing to pups could support a type 2 immune environment in the lung capable of controlling the parasite (22–24) in susceptible IL-4Rα−/− pups to Nb. Transfer of WT immune cells into IL-4Rα−/− mice has been shown to reverse significant immunological pathology (i.e., graft-versus-host disease) (25). Moreover, transfer of maternal immune cells into offspring has been shown by others to correct profound immune phenotypes, e.g., in transforming growth factor–β (TGFβ−/−) offspring (26). We found that IL-4Rα−/− pups nursed on WT NbM showed significantly reduced worm burdens at day 5 PI when compared to IL-4Rα−/− pups nursed by WT NvM mothers (Fig. 5A). This protection in NbM IL-4Rα−/− pups correlated with systemically raised CD4 T cell–associated IL-13 responses when compared to NvM IL-4Rα−/− pups; specifically raised IL-13 in the lung and in CD4 T cells in the lung-draining lymph node along with increased secretion of IL-13 by anti-CD3 restimulated splenocytes (Fig. 5B).
Fig. 5. IL-4Rα−/− offspring nursed on NbM dams have enhanced lymphocyte-mediated immunity against Nb infection.
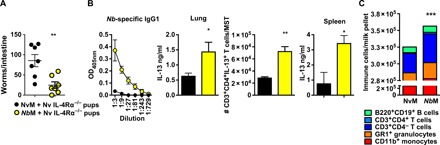
(A) Three-day-old IL-4Rα−/− pups were fostered on either WT NvM or NbM dams. Offspring were infected when 14 days old with 250 × Nb L3 and euthanized at day 5 PI, and intestinal worm burdens were quantified. (B) Nb-specific serum IgG1, anti-CD3 restimulated splenocyte IL-13, lung homogenate levels of IL-13, and CD3+CD4+IL-13+ T cells/MST were quantified in IL-4Rα−/− pups. (C) Numbers of immune cells in breast milk pellets from the stomachs of 10-day-old pups nursed by WT NvM or NbM dams. All data are representative of a minimum of two experimental repeats. *P < 0.05, **P < 0.01, and ***P < 0.001.
Such a transfer of protection to IL-4Rα−/− offspring would require transfer of a range of lymphoid and myeloid cells to achieve this effect. Therefore, protection related to transfer of immune cells may not be restricted to transfer of CD4 T cells. Analysis of immune cell populations in breast milk revealed increased cell numbers of both lymphoid and myeloid cells in breast milk from NbM when compared to NvM (Fig. 5C). This suggests that nursing by NbM could confer protective immunity to IL-4Rα−/− offspring through the transfer of a range of not only CD4 T cells but also populations of myeloid effector cells, which, together, could coordinate an adaptive-like immunity against Nb in the IL-4Rα−/− lung.
Offsprings’ nursing-derived immunity against Nb is dependent on maternally derived type 2 T cells, and these cells are enriched in the milk of NbM mothers
Together, our data show that transfer of immunity to offspring was nursing dependent but not dependent on passive antibody acquisition. Protection is instead associated with a systemic alteration of offspring immunity related to maternal TH2 CD4 T cells penetrating organs throughout the body. The subsequent enhanced type 2 immune footprint left on the offspring is also associated with transfer of protection in an allogeneic setting. In support of this transfer of protection coming via CD4 T cells in the milk, we identified significantly raised numbers of maternal CD3+CD4+ T cells in NbM dam milk when compared to NvM (Fig. 6A). Moreover, these NbM CD3+CD4+ T cells showed increased numbers of type 2 competent CD3+CD4+ST2+ T cells (Fig. 6A). To establish whether transfer of protection was dependent on maternal CD4 T cells being type 2 competent, we nursed naïve WT offspring by NbM dams with disrupted IL-4Rα expression specifically in their T cell compartment (ilckCreIL-4Rα−/lox mice). Offspring nursed on NbM ilckCreIL-4Rα−/lox failed to show enhanced control of Nb, related to an impaired systemic ability to launch a raised IL-13 response after restimulation (Fig. 6B). These data show that type 2 competent maternal CD4 T cells are required for transfer of immunity to offspring and that these protective CD4 T cells are enriched in NbM dam milk.
Fig. 6. Maternally derived type 2 CD4+T cells are enriched in NbM dam milk and are required for transfer of protection.
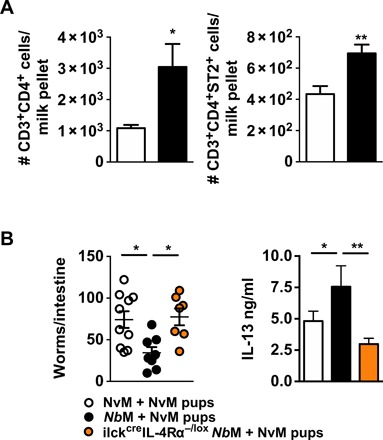
(A) Numbers of Thy1.1+CD4+ and Thy1.1+CD4+ST2+ T cells in milk pellet from the stomach of Thy1.2 pups at day 10 after birth. (B) Three-day-old NvM pups were fostered on WT NvM/NbM or ilckcreIL-4Rα−/lox dams. Pups were infected when 14 days old with 250 × Nb L3 and euthanized at day 5 PI. Offspring intestinal worm burden was analyzed, and IL-13 secretion by anti-CD3–restimulated splenocytes was quantified at day 5 PI. All data are representative of a minimum of two experimental repeats. *P < 0.05 and **P < 0.01.
DISCUSSION
This study reveals that a pre-conception maternal Nb infection transfers a strong and long-lasting protective immunity against Nb infection to her offspring and that this protection was acquired solely through offspring nursing. This maternal imprinting via nursing was profound as demonstrated by its ability to correct genetic immune susceptibility to infection. Moreover, this protection from infection persisted into adulthood and was maintained even in an allogeneic setting.
Protection was not dependent on passive antibody transfer. This is in agreement with our own and others’ findings that antibody has minimal input in immunity to Nb (10, 19). This is not the case for other helminths such as Hp, where antibody contributes significantly to host immunity against the parasite (27, 28). It also highlights that maternally derived immunological influence in offspring from helminth infection will differ depending on the species; unlike Nb, nursing-derived protection against Hp is dependent on antibody transfer (13). While our findings showed that antibody was not essential, we still found a requirement for maternal B cells in transfer of protection that related to offspring acquiring a type 2 CD4 T cell response. These findings show a maternally driven B effector–type response, which may be mediated via B cell–secreted cytokines and/or antigen presentation (10), driving the subsequent transfer of CD4 T cell protection.
Transfer of protection to NbM offspring was dependent on a systemic increase in offspring of CD4 T cell populations with raised levels of canonical anti-helminth TH2 competency and thereby an enhanced immunological ability to resolve an Nb infection (10). The biological potential of this effect to transfer protection was highlighted by our demonstration that nursing by WT NbM transferred protection against Nb to IL-4Rα−/− offspring, which have a genetically impaired ability to launch type 2 immune responses. This protection was coincident with IL-4Rα−/− offspring having a heightened systemic TH2 competency. Typically, IL-4Rα−/− mice display an abrogated ability to secrete TH2 cytokines such as IL-13 (29); however, here, we found that IL-4Rα−/− offspring nursed by NbM are able to secrete IL-13 at levels up to fourfold higher than IL-4Rα−/− offspring nursed by NvM. Moreover, we found that populations of maternally derived myeloid cells known to affect pulmonary anti-Nb immunity were also present in high numbers in the breast milk. Therefore, the required complement of type 2 immune cells for driving pulmonary immunity to Nb could be acquired via nursing.
Others have also shown that maternal cell transfer occurs and can dramatically influence early-life immunity in different contexts, e.g., by reversing the immune pathology resulting from lack of TGFβ (26). However, studies addressing immune cell transfer via nursing have not demonstrated whether this effect confers long-term influence on offspring ability to control infection. Our data show that pathogen-experienced maternal T cells can systemically train offspring immunity. We show that the influence and penetration of these cells in offspring are not restricted to the digestive tract (the site of cell uptake) (30); these cells can also be found in (for example) the lung and spleen. As this TH2 CD4 T cell–associated enhanced control of infection was maintained in an allogeneic setting, our observations are applicable in an immune setting reflective of normal MHC diversity in humans. In this allogeneic setting, antigen presentation would be impaired between maternally derived cells and foster pup cells. Moreover, transferred cells are likely to be exposed to high levels of cytotoxic killing by foster pup cells, although, as outlined below, the tolerogenic environment promoted during gestation and nursing may abrogate the cytotoxicity. Together, these data suggest that transfer of protection may be affected by secreted factors (such as cytokines) derived from maternally transferred cells as well as interactions dependent on direct contact between maternal and offspring cells.
Understanding the maternal-offspring interface in the context of type 2 immunity is also important. In early life, the immune system shows an inherent type 2 immune bias (31) and an ability to suppress TH1 immunity (32). This elevated prenatal type 2 immunity appears to contribute to fetal allograft survival by impairing potential cytotoxic events against it (33). This tolerogenic effect may also provide a permissive window that promotes an extended survival of maternal cells that is sufficient to allow immunological imprinting in the offspring to occur. In the neonate, the maintained TH2 bias can also contribute to the coordination of early-life pulmonary development (34), possibly at a cost of enhanced risk of neonatal allergic disease developing (35). Our findings add an important further dimension to this understanding of early-life TH2 bias: It may provide protection against helminth infection in early life.
Therefore, in this study, we demonstrate that maternal helminth infection before pregnancy can provide offspring with the ability throughout life to launch a heightened protective cellular immune response against the parasite. This protection is not dependent on antibody but is instead dependent on transfer of maternal type 2 competent cells. We show that maternal TH2-competent CD4 T cells were enriched in the milk of NbM dams, dispersed systemically throughout the offspring, and provided an environment conducive to initiating an enhanced and long-term ability in the offspring to launch a protective immune response against subsequent helminth infection. Therefore, transfer to offspring of maternally derived pathogen-conditioned lymphocytes via nursing represents a novel transgenerational mechanism by which offspring may acquire early-life immunity that persists into adulthood, but how this maternal cellular transfer of immunity manifests itself in clinically relevant human and veterinary settings is not yet defined and represents a potentially significant new paradigm in our understanding of maternal transfer of immunity to offspring.
MATERIALS AND METHODS
Animal work
Mice used
The following BALB/c background mouse strains were used in the study: BALB/c (CD90.2), IL-4Rα−/− (described as IL4ratm1Fbb/IL4ratm1Fbb), μMT (B cell deficient), Thy1.1 (CD90.1), MB1creIL-4Rα−/lox [B cell–specific disruption of IL-4Rα expression (36)], and ilckcreIL-4Rα−/lox [CD4 T cell–specific disruption of IL-4Rα expression (37)]. C57BL/6 background mouse strains used were C57BL/6 and IgMi (20). Mice were bred and housed in specific pathogen–free conditions at the Animal Unit of the University of Cape Town, South Africa. All experimental mice were used between 1 and 15 weeks of age with appropriate littermate controls of the same generation. Mice were euthanized by lethal halothane inhalation.
Ethics statement
All studies were carried out in accordance with protocol 012/054 or 014/021 approved by the Faculty of Health Sciences Animal Ethics Committee from the University of Cape Town.
Mating and litter swaps
Female mice aged between 7 and 8 weeks were mated with male mice at one male per cage of two females over 2 weeks, after which the male was removed. Females gave birth approximately 21 days after fertilization, and the birth of pups was monitored daily to ensure age matching. A maximum of two mothers and eight pups per mother were housed per cage. In the long-term experiments, pups were weaned at 3 weeks of age, separated from their mothers, and housed by gender.
Infection with Nb
Adult mice (older than 6 weeks) were injected subcutaneously with 500 × Nb L3 larvae. Pups were infected with a half-dose of 250 × Nb L3 larvae. To enumerate adult worms, mice were euthanized at various times PI, intestines were opened longitudinally and incubated in 10 ml of saline for 3 hours at 37°C, and parasites were counted under a dissecting microscope.
The mice used (except IL-4Rα−/−) for infection experiments would naturally clear Nb at approximately 9 days PI, but to ensure that all worms were expelled, mothers were treated with ivermectin (10 mg/ml) in their drinking water from days 7 to 14 after primary infection to clear the pathogen before mating.
Flow cytometry
Single-cell suspensions were prepared, and 1 × 106 cells were incubated in phosphate-buffered saline + 0.5% bovine serum albumin, 1% normal rat serum, and appropriate antibody cocktails. Cell populations were acquired and determined on a BD FACS Fortessa (Becton Dickinson). Cell populations were identified by the following antibody staining strategies: CD4 T cells: CD3+CD4+. CD4 T cells populations were additionally stratified into activated effector (CD44+CD62Llo), T1/ST2+ and Thy1.1+Thy1.2−, or Thy1.2+Thy1.1− T cell populations.
Intracellular cytokine staining was also carried out on T cells to determine IL-13 expression. Cells were resuspended in complete medium [Iscove's modified Dulbecco’s medium (IMDM; GIBCO/Invitrogen, Carlsbad, CA) + 10% fetal calf serum (FCS), penicillin/streptomycin] at 2.5 × 107/ml and stimulated with phorbol 12-myristate 13-acetate/ionomycin (10 μg/ml) and GolgiStop (as per the manufacturer’s protocol; BD Pharmingen) at 37°C for 4 hours. After restimulation, cells were surface-stained for CD3 and CD4 and then fixed and permeabilized with Cytofix/Cytoperm Plus (as per the manufacturer’s instructions; BD Pharmingen). Intracellular staining was performed by staining cells with IL-13 or appropriate control. All analyses were performed with FlowJo software.
Antibody ELISA
Relative antigen-specific serum antibody levels were determined by enzyme-linked immunosorbent assay (ELISA). Briefly, plates were coated with Nb somatic antigen (10 μg/ml). Blood serum was added and incubated overnight before the commercially available alkaline phosphatase–conjugated secondary antibody for IgG1 (BD Pharmingen) was added. 4-p-Nitrophenol-phosphate was used to develop the reaction. Absorbance was read at λ = 405 nm against a reference measurement of λ = 490 nm using the SoftMax Pro software on a VersaMax microplate reader. Relative antigen-specific antibody levels were plotted as dilution graphs. Nb somatic antigen was generated by isolating the soluble fraction from homogenized Nb L3 larvae.
Milk preparation
A modified method of the one described by Cabinian et al. (30) was used to prepare milk for analysis of cell populations. The milk bolus was extracted from the stomachs of 10-day-old pups and conserved in complete medium (IMDM + 10% FCS, penicillin/streptomycin, Hepes). The bolus was passed through a 100-μm cell strainer to generate a single-cell suspension, and the strainer was washed with 0.17 M tris (pH 7.6) and 0.83% NH4Cl buffer. The cell suspension was centrifuged at 400g for 15 min. Pellets containing cells were washed a further three times in complete medium before further analysis.
Cytokine ELISA
Cytokine ELISAs were performed as previously described (38) using coating and biotinylated detection antibodies from R&D Systems. Streptavidin-conjugated horseradish peroxidase was used for detection with a commercially available substrate solution. Spleen cells were plated at 1 × 106 cells per well in 48-well plates precoated with anti-CD3 (20 μg/ml) and restimulated for 72 hours before the supernatant was analyzed for cytokines. Homogenates of lung sections were prepared using a Polytron homogenizer, and the protein concentration of the soluble fraction of the lung samples was equalized before ELISA.
Statistics
The results are expressed as either individual mice/data points or group mean ± SD. P values and significances were determined using either the one-tailed Mann-Whitney t test or nonparametric one-way analysis of variance (ANOVA) (GraphPad Prism Software). Groups were judged to be significantly different if the P value was less than 0.05 (*P < 0.05, **P < 0.01, and ***P < 0.001).
Supplementary Material
Acknowledgments
Funding: The work presented here was supported by grants from the Carnegie Corporation of New York (to M.G.D.), Horizon 2020 (to W.G.C.H.), National Research Foundation (78912 to W.G.C.H. and 81578 to M.G.D.), Deutsche Forschungsgemeinschaft (LA 2746/2 to W.G.C.H.), and BBSRC (BB/R015856/1 to W.G.C.H. and BB/M025292/1 to K.-M.T.). Author contributions: Experiments were carried out by W.G.C.H., M.G.D., A.C., M.R., D.M., B.G.D., D.S., C.M., and K.S. Data analysis was performed by W.G.C.H., M.G.D., A.C., M.R., D.M., B.G.D., D.S., C.M., D.N., A.F.C., and K.S. Reagents and materials were provided by V.Q., B.R., F.B., A.W., and K.-M.T. Study design was carried out by W.G.C.H., B.G.D., A.C., B.R., and H.J. Manuscript was prepared by W.G.C.H., B.G.D., A.C., F.B., B.R., H.J., M.G.D., A.F.C., and D.M. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/5/eaav3058/DC1
Fig. S1. Offspring acquire, via nursing, a population of maternally derived CD4 T cells with a heightened TH2 responsiveness following Nb infection.
Fig. S2. Offspring nursed on Hp-infected mothers do not demonstrate maternal cell transfer.
Fig. S3. Allogeneic NbM offspring have raised numbers of maternally derived lung CD4+ T cells following Nb infection.
Fig. S4. Offspring acquire, via nursing, persistent protection from Nb related to raised TH2 CD4+ T cell responses following Nb infection in an allogeneic setting.
REFERENCES AND NOTES
- 1.Neuzil K. M., Mellen B. G., Wright P. F., Mitchel E. F. Jr., Griffin M. R., The effect of influenza on hospitalizations, outpatient visits, and courses of antibiotics in children. N. Engl. J. Med. 342, 225–231 (2000). [DOI] [PubMed] [Google Scholar]
- 2.E. A. F. Simoes, E. A. F. Simoes, T. Cherian, J. Chow, S. A. Shahid-Salles, R. Laxminarayan, T. J. John, Acute respiratory infections in children, in Disease Control Priorities in Developing Countries, D. T. Jamison, J. G. Breman, A. R. Measham, G. Alleyne, M. Claeson, D. B. Evans, P. Jha, A. Mills, P. Musgrove, Eds. (Oxford Univ. Press, 2006). [Google Scholar]
- 3.Jones G., Steketee R. W., Black R. E., Bhutta Z. A., Morris S. S.; Bellagio Child Survival Study Group , How many child deaths can we prevent this year? Lancet 362, 65–71 (2003). [DOI] [PubMed] [Google Scholar]
- 4.Wolf J. H., Low breastfeeding rates and public health in the United States. Am. J. Public Health 93, 2000–2010 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Heath P. T., Culley F. J., Jones C. E., Kampmann B., le Doare K., Nunes M. C., Sadarangani M., Chaudhry Z., Baker C. J., Openshaw P. J. M., Group B streptococcus and respiratory syncytial virus immunisation during pregnancy: A landscape analysis. Lancet Infect. Dis. 17, e223–e234 (2017). [DOI] [PubMed] [Google Scholar]
- 6.Marchant A., Sadarangani M., Garand M., Dauby N., Verhasselt V., Pereira L., Bjornson G., Jones C. E., Halperin S. A., Edwards K. M., Heath P., Openshaw P. J., Scheifele D. W., Kollmann T. R., Maternal immunisation: Collaborating with mother nature. Lancet Infect. Dis. 17, e197–e208 (2017). [DOI] [PubMed] [Google Scholar]
- 7.Verhasselt V., Is infant immunization by breastfeeding possible? Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 1–6 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ghosh M. K., Nguyen V., Muller H. K., Walker A. M., Maternal milk T cells drive development of transgenerational Th1 immunity in offspring thymus. J. Immunol. 197, 2290–2296 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dewals B. G., Layland L. E., Prazeres da Costa C., Horsnell W. G., Maternal helminth infections and the shaping of offspring immunity. Parasite Immunol. 41, e12599 (2019). [DOI] [PubMed] [Google Scholar]
- 10.Horsnell W. G., Darby M. G., Hoving J. C., Nieuwenhuizen N., McSorley H. J., Ndlovu H., Bobat S., Kimberg M., Kirstein F., Cutler A. J., DeWals B., Cunningham A. F., Brombacher F., IL-4Rα-associated antigen processing by B cells promotes immunity in Nippostrongylus brasiliensis infection. PLOS Pathog. 9, e1003662 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gause W. C., Maizels R. M., Macrobiota—Helminths as active participants and partners of the microbiota in host intestinal homeostasis. Curr. Opin. Microbiol. 32, 14–18 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Maizels R. M., Parasitic helminth infections and the control of human allergic and autoimmune disorders. Clin. Microbiol. Infect. 22, 481–486 (2016). [DOI] [PubMed] [Google Scholar]
- 13.Harris N. L., Spoerri I., Schopfer J. F., Nembrini C., Merky P., Massacand J., Urban J. F. Jr., Lamarre A., Burki K., Odermatt B., Zinkernagel R. M., Macpherson A. J., Mechanisms of neonatal mucosal antibody protection. J. Immunol. 177, 6256–6262 (2006). [DOI] [PubMed] [Google Scholar]
- 14.Straubinger K., Paul S., Prazeres da Costa O., Ritter M., Buch T., Busch D. H., Layland L. E., Prazeres da Costa C. U., Maternal immune response to helminth infection during pregnancy determines offspring susceptibility to allergic airway inflammation. J. Allergy Clin. Immunol. 134, 1271–1279.e10 (2014). [DOI] [PubMed] [Google Scholar]
- 15.Guadalupe I., Mitre E., Benitez S., Chico M. E., Nutman T. B., Cooper P. J., Evidence for in utero sensitization to Ascaris lumbricoides in newborns of mothers with ascariasis. J. Infect. Dis. 199, 1846–1850 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mpairwe H., Ndibazza J., Webb E. L., Nampijja M., Muhangi L., Apule B., Lule S., Akurut H., Kizito D., Kakande M., Jones F. M., Fitzsimmons C. M., Muwanga M., Rodrigues L. C., Dunne D. W., Elliott A. M., Maternal hookworm modifies risk factors for childhood eczema: Results from a birth cohort in Uganda. Pediatr. Allergy Immunol. 25, 481–488 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Thawer S. G., Horsnell W. G. C., Darby M., Hoving J. C., Dewals B., Cutler A. J., Lang D., Brombacher F., Lung-resident CD4+ T cells are sufficient for IL-4Rα-dependent recall immunity to Nippostrongylus brasiliensis infection. Mucosal Immunol. 7, 239–248 (2014). [DOI] [PubMed] [Google Scholar]
- 18.Hanson L. A., Korotkova M., Lundin S., Håversen L., Silfverdal S. A., Mattsby-Baltzer I., Strandvik B., Telemo E., The transfer of immunity from mother to child. Ann. N. Y. Acad. Sci. 987, 199–206 (2003). [DOI] [PubMed] [Google Scholar]
- 19.Liu Q., Kreider T., Bowdridge S., Liu Z., Song Y., Gaydo A. G., Urban J. F. Jr., Gause W. C., B cells have distinct roles in host protection against different nematode parasites. J. Immunol. 184, 5213–5223 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Waisman A., Kraus M., Seagal J., Ghosh S., Melamed D., Song J., Sasaki Y., Classen S., Lutz C., Brombacher F., Nitschke L., Rajewsky K., IgG1 B cell receptor signaling is inhibited by CD22 and promotes the development of B cells whose survival is less dependent on Igα/β. J. Exp. Med. 204, 747–758 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vieira P., Rajewsky K., The half-lives of serum immunoglobulins in adult mice. Eur. J. Immunol. 18, 313–316 (1988). [DOI] [PubMed] [Google Scholar]
- 22.Chen F., Wu W., Millman A., Craft J. F., Chen E., Patel N., Boucher J. L., Urban J. F. Jr., Kim C. C., Gause W. C., Neutrophils prime a long-lived effector macrophage phenotype that mediates accelerated helminth expulsion. Nat. Immunol. 15, 938–946 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Harvie M., Camberis M., Tang S. C., Delahunt B., Paul W., Le Gros G., The lung is an important site for priming CD4 T-cell-mediated protective immunity against gastrointestinal helminth parasites. Infect. Immun. 78, 3753–3762 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bouchery T., Kyle R., Camberis M., Shepherd A., Filbey K., Smith A., Harvie M., Painter G., Johnston K., Ferguson P., Jain R., Roediger B., Delahunt B., Weninger W., Forbes-Blom E., Le Gros G., ILC2s and T cells cooperate to ensure maintenance of M2 macrophages for lung immunity against hookworms. Nat. Commun. 6, 6970 (2015). [DOI] [PubMed] [Google Scholar]
- 25.Urso K., Alvarez D., Cremasco V., Tsang K., Grauel A., Lafyatis R., von Andrian U. H., Ermann J., Aliprantis A. O., IL4RA on lymphatic endothelial cells promotes T cell egress during sclerodermatous graft versus host disease. JCI Insight 1, e88057 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Letterio J. J., Geiser A. G., Kulkarni A. B., Roche N. S., Sporn M. B., Roberts A. B., Maternal rescue of transforming growth factor-beta 1 null mice. Science 264, 1936–1938 (1994). [DOI] [PubMed] [Google Scholar]
- 27.Esser-von Bieren J., Mosconi I., Guiet R., Piersgilli A., Volpe B., Chen F., Gause W. C., Seitz A., Verbeek J. S., Harris N. L., Antibodies trap tissue migrating helminth larvae and prevent tissue damage by driving IL-4Rα-independent alternative differentiation of macrophages. PLOS Pathog. 9, e1003771 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.McCoy K. D., Stoel M., Stettler R., Merky P., Fink K., Senn B. M., Schaer C., Massacand J., Odermatt B., Oettgen H. C., Zinkernagel R. M., Bos N. A., Hengartner H., Macpherson A. J., Harris N. L., Polyclonal and specific antibodies mediate protective immunity against enteric helminth infection. Cell Host Microbe 4, 362–373 (2008). [DOI] [PubMed] [Google Scholar]
- 29.Barner M., Mohrs M., Brombacher F., Kopf M., Differences between IL-4R alpha-deficient and IL-4-deficient mice reveal a role for IL-13 in the regulation of Th2 responses. Curr. Biol. 8, 669–672 (1998). [DOI] [PubMed] [Google Scholar]
- 30.Cabinian A., Sinsimer D., Tang M., Zumba O., Mehta H., Toma A., Sant’Angelo D., Laouar Y., Laouar A., Transfer of maternal immune cells by breastfeeding: Maternal cytotoxic T lymphocytes present in breast milk localize in the Peyer’s patches of the nursed infant. PLOS ONE 11, e0156762 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sornasse T., Larenas P. V., Davis K. A., de Vries J. E., Yssel H., Differentiation and stability of T helper 1 and 2 cells derived from naive human neonatal CD4+ T cells, analyzed at the single-cell level. J. Exp. Med. 184, 473–483 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li L., Lee H. H., Bell J. J., Gregg R. K., Ellis J. S., Gessner A., Zaghouani H., IL-4 utilizes an alternative receptor to drive apoptosis of Th1 cells and skews neonatal immunity toward Th2. Immunity 20, 429–440 (2004). [DOI] [PubMed] [Google Scholar]
- 33.Sefrioui H., Donahue J., Gilpin E. A., Srivastava A. S., Carrier E., Tolerance and immunity following in utero transplantation of allogeneic fetal liver cells: The cytokine shift. Cell Transplant. 12, 75–82 (2003). [DOI] [PubMed] [Google Scholar]
- 34.Jones C. V., Williams T. M., Walker K. A., Dickinson H., Sakkal S., Rumballe B. A., Little M. H., Jenkin G., Ricardo S. D., M2 macrophage polarisation is associated with alveolar formation during postnatal lung development. Respir. Res. 14, 41 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.de Kleer I. M., Kool M., de Bruijn M. J. W., Willart M., van Moorleghem J., Schuijs M. J., Plantinga M., Beyaert R., Hams E., Fallon P. G., Hammad H., Hendriks R. W., Lambrecht B. N., Perinatal activation of the interleukin-33 pathway promotes type 2 immunity in the developing lung. Immunity 45, 1285–1298 (2016). [DOI] [PubMed] [Google Scholar]
- 36.Hoving J. C., Kirstein F., Nieuwenhuizen N. E., Fick L. C. E., Hobeika E., Reth M., Brombacher F., B cells that produce immunoglobulin E mediate colitis in BALB/c mice. Gastroenterology 142, 96–108 (2012). [DOI] [PubMed] [Google Scholar]
- 37.Radwanska M., Cutler A. J., Hoving J. C., Magez S., Holscher C., Bohms A., Arendse B., Kirsch R., Hunig T., Alexander J., Kaye P., Brombacher F., Deletion of IL-4Rα on CD4 T cells renders BALB/c mice resistant to Leishmania major infection. PLOS Pathog. 3, e68 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Darby M., Schnoeller C., Vira A., Culley F., Bobat S., Logan E., Kirstein F., Wess J., Cunningham A. F., Brombacher F., Selkirk M. E., Horsnell W. G. C., The M3 muscarinic receptor is required for optimal adaptive immunity to helminth and bacterial infection. PLOS Pathog. 11, e1004636 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/5/eaav3058/DC1
Fig. S1. Offspring acquire, via nursing, a population of maternally derived CD4 T cells with a heightened TH2 responsiveness following Nb infection.
Fig. S2. Offspring nursed on Hp-infected mothers do not demonstrate maternal cell transfer.
Fig. S3. Allogeneic NbM offspring have raised numbers of maternally derived lung CD4+ T cells following Nb infection.
Fig. S4. Offspring acquire, via nursing, persistent protection from Nb related to raised TH2 CD4+ T cell responses following Nb infection in an allogeneic setting.