

Abstract
Background
Breast cancer is one of the most common endocrine cancers among females worldwide. Distant metastasis of breast cancer is causing an increasing number of breast cancer-related deaths. However, the potential mechanisms of metastasis and candidate biomarkers remain to be further explored.
Results
The gene expression profiles of GSE102484 were downloaded from the Gene Expression Omnibus (GEO) database. Weighted gene co-expression network analysis (WGCNA) was used to screen for the most potent gene modules associated with the metastatic risk of breast cancer, and a total of 12 modules were identified based on the analysis. In the most significant module (R2 = 0.68), 21 network hub genes (MM > 0.90) were retained for further analyses. Next, protein-protein interaction (PPI) networks were used to further explore the biomarkers with the most interactions in gene modules. According to the PPI networks, five hub genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) were identified as key genes associated with breast cancer progression. Furthermore, the prognostic value and differential expression of these genes were validated based on data from The Cancer Genome Atlas (TCGA) and Kaplan-Meier (KM) Plotter. Receiver operating characteristic (ROC) curve analysis revealed that the mRNA expression levels of these five hub genes showed excellent diagnostic value for breast cancer and adjacent tissues. Moreover, these five hub genes were significantly associated with worse distant metastasis-free survival (DMFS) in the patient cohort based on KM Plotter.
Conclusion
Five hub genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) associated with the risk of distant metastasis were extracted for further research, which might be used as biomarkers to predict distant metastasis of breast cancer.
Keywords: Breast cancer, WGCNA, Bioinformatic analysis, Prognosis, Metastasis
Background
Breast cancer is one of the most common tumours threatening women’s health worldwide, and its incidence has shown a rising trend in the past decades. According to the prediction provided by the American Cancer Society (ACS), there will be more than 271,000 new incidences in the United States (US) in 2019 [1]. Current therapeutic approaches for breast cancer mainly focus on comprehensive treatment, including surgery, chemotherapy, radiotherapy, hormone therapy, and targeted therapy [2, 3]. Breast cancer in situ is usually not fatal to patients; however, advanced breast cancer with lymph nodes and/or distant metastasis tends to cause life-threatening outcomes for patients [4]. Although the aetiology and signatures of breast cancer have been preliminarily explored, there is no defined means of predicting the metastasis and recurrence of breast cancer in clinical practice [5], and further exploration of the potential mechanisms of the metastasis and biomarkers to monitor recurrence of breast cancer is urgently needed.
With the continuous development of biological research technologies, especially sequencing technologies and bioinformatic algorithms, massive amounts of genomic information are accumulating exponentially [6, 7]. Along with the successful implementation of numerous large-scale sequencing projects led by governments, biomedical research has been entering the era of “big data” [8–10]. Over the past few years, bioinformatics researchers have developed a series of analysis strategies and data mining algorithms for increasing the amount of transcriptomic data [11–13].
Weighted gene co-expression network analysis (WGCNA) is a systematic biological strategy for evaluating gene association patterns among different samples [11]. It can be applied to reveal highly correlative gene sets and explore potential biomarker genes or therapeutic targets according to the internal connectivity of gene clusters and the associations between gene clusters and phenotypes. By constructing a gene co-expression network and identifying related gene clusters, the correlation between gene modules and phenotypes can be calculated based on phenotypic information, and the most relevant gene modules can be found. Numerous potential biomarkers have been identified to date based on WGCNA for sequencing data [14–16]. For example, Tang et al. identified five genes as prognostic biomarkers for breast cancer, and Qiu et al. revealed several genes associated with the development of breast cancer for further basic and clinical research [14, 17]. However, there is currently no defined means of predicting the metastasis risk of breast cancer in clinical practice. Thus, we aim to further explore potential metastatic predictors via WGCNA.
GSE102484, a microarray containing 683 breast cancer samples, was submitted by Cheng et al. from Taiwan in 2017 [18]. Cheng et al. validated the prognostic value of an 18-gene classifier that predicted locoregional recurrence and metastasis risk in patients after mastectomy based on the sequencing data and clinical information of GSE102484 [18, 19]. Given the complete follow-up information and the defined risk scoring system of recurrence and metastasis for this microarray data, we re-evaluated these data in this study and identified five candidate biomarkers associated with the risk of metastasis through WGCNA and protein-protein interaction (PPI) networks. We further validated the differential expression and prognostic value of these five genes with another open database. Overall, we identified five genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) associated with distant metastasis, indicating these genes as potential biomarkers for assessing the risk of breast cancer recurrence and distant metastasis.
Materials and methods
Acquisition of microarray data
The workflow of our research is illustrated in Fig. 1. The array profiles of GSE102484 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE102484) contributed by Cheng et al. were downloaded from the Gene Expression Omnibus (GEO) database. GSE102484 was an array profile based on the GPL570 platform (Affymetrix Human Genome U133 Plus 2.0 Array) containing 683 breast cancer samples from Taiwan. The robust multi-array average (RMA) algorithm in the affy package within Bioconductor (http://www.bioconductor.org) in R was employed to preprocess the array profiles. After background correction, quantile normalization and probe summarization, the data set with 21,653 genes was further processed. The top 25% most variant genes (MVGs) according to analysis of variance (5413 genes) were selected for further WGCNA.
Fig. 1.
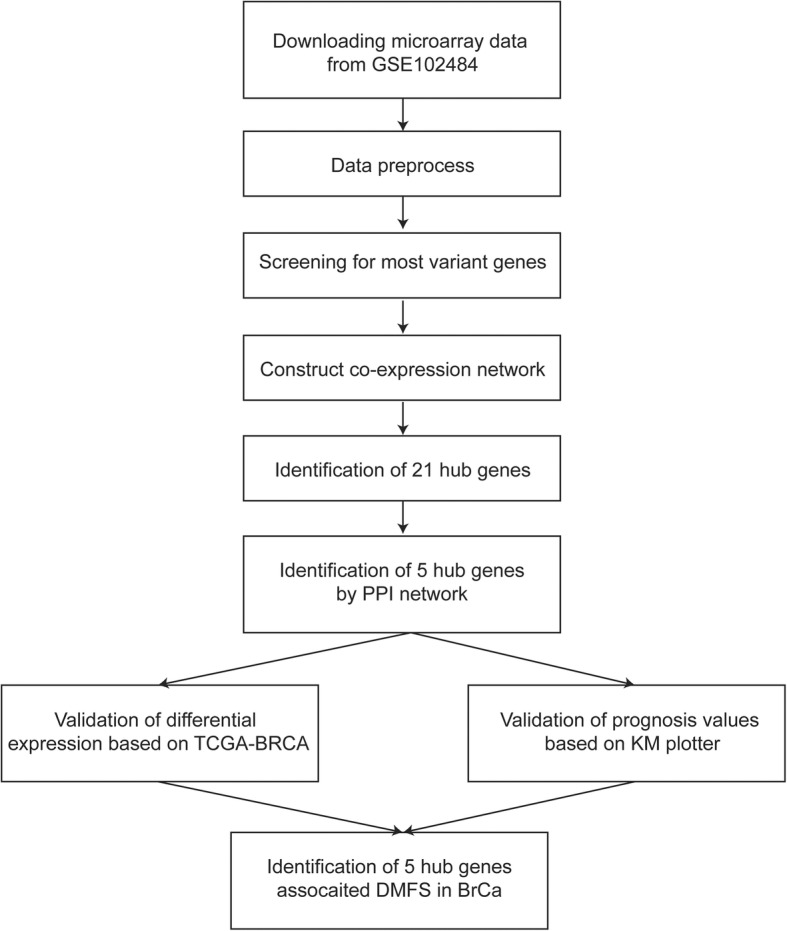
Flow chart of data preparation, processing, analysis, and validation. The gene expression profiles of GSE102484 were downloaded from the GEO database. WGCNA and PPI networks were further used to investigate potential biomarkers associated with the clinical stages and risk of distant metastasis. In addition, the prognostic value and differential expression of hub genes were validated based on data from TCGA-BRCA and KM Plotter
Co-expression network construction
After preprocessing the GSE102484 microarray data, the expression profile of these 5413 genes was employed to construct to a gene co-expression network using the WGCNA package in R [11]. The idea of a soft threshold is to continually elementize the elements in the Adjacency Matrix through a weight function. Because the choice of the soft threshold, β, is bound to affect the result of module identification and the relative network of the random average of each node, there is a scale-free network in which a few nodes exhibit a significantly higher degree than the general point, which is a more stable choice, so we need to choose a soft threshold, β, is our gene distribution in line with the scale-free network. To create a network with a nearly scale-free topology, we installed the soft threshold power of β = 4 (scale free R2 = 0.88). Adjacency matrices were calculated and transformed into the topological overlap matrix (TOM). The dynamic tree cut algorithm was applied to detect gene modules. Gene significance (GS) was defined as the correlation coefficient between gene expression and module traits. The module eigengene was calculated as a summary profile for each module. Module significance was defined as the correlation coefficient between a module’s eigengene and traits. Module membership (MM) was defined by the correlation coefficient of the module eigengene and gene expression profile. Genes with MM values above 0.90 were considered to be the modules’ representative genes with potential critical functions.
Identification of hub genes based on a PPI network
After WGCNA of the microarray data, we screened 21 hub genes in the yellow module, which were significantly associated with clinical stage, T stage, and risk of metastasis. We constructed the protein–protein interaction (PPI) network using the STRING database (https://string-db.org/) to load all the hub genes [20]. For all other parameters, the default settings were used. *.tsv format network files were loaded into the plug-in cytoHubba based on Cytoscape software version 3.5.1 (Institute for Systems Biology, Seattle, WA, USA). We defined the top 5 genes with the highest prediction scores calculated by the Stress algorithm as hub genes. We defined these genes with a minimum required interaction score greater than 0.90 as hub genes according to the PPI network, and the network diagram of all PPI hub genes was visualized with Cytoscape (version 3.5.1, Seattle, WA) [12].
Further authentication of key genes using other open data
To verify the significant values of five hub genes, the clinical files and RNA-seq data for breast cancer were downloaded from The Cancer Genome Atlas (TCGA, https://cancergenome.nih.gov/) database. The mRNA sequencing data were normalized using the edge R package in R software, and we used Student’s t-test and the receiver operating characteristic (ROC) curve to examine the differential mRNA levels of these five genes in breast cancer and adjacent tissues. The prognostic impacts of these five genes in breast cancer were analysed using the Kaplan-Meier Plotter (http://kmplot.com/analysis/), an online database containing gene expression profiles and survival data for breast cancer patients [21]. According to the mRNA expression of the genes, the cases in the database were ranked from high expression to low expression, which divided them into two groups; the top 50% were divided into the high expression group, and the bottom 50% belonged to the low expression group. All cohorts were compared with Kaplan-Meier survival plots. The hazard ratio (HR), 95% confidence interval (95%CI), and log rank P value were calculated and displayed online.
Statistical analysis
All statistical analyses were performed using SPSS 25.0 software and R 3.5.1 software. Most statistical comparisons among different groups were analysed with Student’s t-test and one-way analysis of variance (ANOVA). The ROC) curve was applied to examine the diagnostic value of differential hub gene mRNA expression for distinguishing tumour tissues from normal tissues. Kaplan-Meier survival plots were generated with survival curves compared by the log-rank test. For all analyses, differences were considered statistically significant if the P values were less than 0.05.
Results
Weighted co-expression network construction and key module identification
The R package for WGCNA was applied to construct a co-expression network, and 2370 MVGs with similar expression patterns were submitted to modules by cluster analysis. In our research, we selected the power of β = 4 (scale free R2 = 0.88) as the soft threshold to ensure a scale-free network (Fig. 2A-D). Then, we extracted twelve modules for further analysis (Fig. 2E). We next visualized the gene network with a heatmap and meta-modules (Fig. 3A, B). The yellow module, which was most significantly associated with clinical stage, T stage, and metastasis risk, was shown to be of notable value in the evaluation of breast cancer progression. Clinical stage (R = 0.21; P = 2e− 8), T stage (R = 0.20; P = 1e− 7) and metastasis risk (R = 0.68; P = 1e− 94) were all significantly correlated with the yellow module according to module-feature relationship analysis (Fig. 3C). Subsequently, we selected 21 genes in the yellow module with MM values above 0.90 for further analysis, which were considered representative genes exhibiting potential critical functions.
Fig. 2.

Determination of soft-thresholding power in WGCNA. (a) Analysis of the scale-free fitting indices for various soft-thresholding powers (β). (b) Mean connectivity analysis of various soft-thresholding powers. (c) Histogram of the connection distribution when β = 4. (d) Checking the scale-free topology when β = 4. According to Fig. 2C and D, k and p(k) are negatively correlated (correlation coefficient 0.85), indicating that a gene scale-free network can be resumed. (E) Clustering dendrograms of genes based on dissimilarity topological overlap and module colours. As a result, 12 co-expression modules were constructed and are shown in different colours. These modules are arranged from large to small according to the number of genes included
Fig. 3.

Identification of relevant modules of breast cancer clinical traits and construction of the PPI network. (a) Visualization of the gene network with a heatmap. The heatmap depicts the TOM among all genes in the analysis. A light colour represents low overlap, and progressively darker reds indicate higher overlap. Blocks of darker colours along the diagonal are the modules. The gene dendrogram and module assignment are also shown along the left and upper sides. (b) The eigengene dendrogram and heatmap identify groups of correlated eigengenes termed meta-modules. As a result, the dendrogram shows that yellow, pink and green-yellow modules are highly related to breast cancer clinical stage. (c) Heatmap of the correlation between module eigengenes and clinical traits of breast cancer. The yellow gene module was shown to exhibit the strongest correlation with the clinical stage (clinical stage, T stage, N stage, metastatic risk). (d) PPI network of genes in the yellow module for clinical stage. The nodes other than the pale blue nodes represent the hub genes with the highest prediction scores calculated with the stress algorithm
Identification of hub genes of modules and filtration via the PPI network
In this study, we extracted 21 hub genes with a high correlation with the yellow module (Fig. 3C). Subsequently, we constructed a PPI network using Cytoscape, and we screened these 21 PPI hub genes exhibiting intimate interactions with other genes (Fig. 3D). Based on the stress algorithm, we retained the top five genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) showing the closest connections with other genes for validation and subsequent investigation (Table 1).
Table 1.
Five hub genes identified by combined bioinformatic strategies in breast cancer
| Gene ID | Ensembl ID | Gene Description |
|---|---|---|
| TPX2 | ENSG00000088325.15 | Targeting protein for Xenopus kinesin-like protein 2 |
| KIF2C | ENSG00000142945.12 | Kinesin family member 2C |
| CDCA8 | ENSG00000134690.10 | Cell division cycle associated 8 |
| BUB1B | ENSG00000156970.12 | BUB1 mitotic checkpoint serine/threonine kinase B |
| CCNA2 | ENSG00000145386.9 | Cyclin A2 |
Validation of hub genes based on TCGA-BRCA data
The training dataset (TCGA-BRCA) was applied to validate the correlations between the five hub genes and clinical stages. We compared the expression of each candidate hub gene in breast cancer samples at different clinical stages and found that the five candidate hub genes were closely related to clinical stages (Fig. 4A-4E). In addition, Wang et al. verified a list of cancer/testis genes showing high specificity of testis-specific expression, which included these five hub genes [22]. We speculate that these five genes were upregulated in tumour tissues. We used the TCGA-BRCA dataset to validate our hypothesis, and the results showed that the mRNA level of each candidate hub gene was significantly upregulated in breast cancer tissues compared with paired adjacent breast tissues (Fig. 5A-5E). In addition, ROC analysis revealed that the mRNA levels of these five genes showed excellent diagnostic value for breast cancer and adjacent tissues (Fig. 6A-6E).
Fig. 4.

Validation of the differential expression of five hub genes in various clinical stages. (a) The correlation of TPX2 expression with clinical stage. (b) The correlation of KIF2C expression with clinical stage. (c) The correlation of CDCA8 expression with clinical stage. (d) The correlation of BUB1B expression with clinical stage. (e) The correlation of CCNA2 expression with clinical stage. ANOVA was used to assess the statistical significance of the differences
Fig. 5.

Expression of the five hub genes between normal and breast cancer tissues. The mRNA levels of (A) TPX2, (B) KIF2C, (C) CDCA8, (D) BUB1B, (E) CCNA2, were significantly upregulated in breast cancer tissues compared with adjacent breast tissues. Two-tailed Student’s t-tests were used to assess the statistical significance of differences
Fig. 6.

Diagnostic value of the five hub genes in identifying normal and breast cancer tissues. The ROC curve revealed that the mRNA levels of these five genes exhibited excellent diagnostic efficiency for breast cancer and adjacent tissues. (a) TPX2, (b) KIF2C, (c) CDCA8, (d) BUB1B, (E) CCNA2
Prognosis value of key genes for DMFS according to KM plotter
To validate the association between the expression levels of these five hub genes and metastasis risk in breast cancer, we used survival information from the KM Plotter database to perform survival analysis for the five hub genes. As shown in Fig. 7, high mRNA expression levels of TPX2 (HR = 1.87; 95%CI: 1.53–2.28; P < 0.001), KIF2C (HR = 1.87; 95%CI: 1.53–2.27; P < 0.001), CDCA8 (HR = 1.67; 95%CI: 1.37–2.04; P < 0.001), BUB1B (HR = 1.64; 95%CI: 1.35–1.99; P < 0.001), and CCNA2 (HR = 1.63; 95%CI: 1.34–1.98; P < 0.001) were significantly associated with worse distant metastasis-free survival (DMFS). Overall, these findings validated the prognostic value and the relationships between the five genes and the metastasis of breast cancer.
Fig. 7.

Prognostic value of the five hub genes in breast cancer patients based on KM Plotter. The patients were divided into a high-expression group and a low-expression group according to the median gene expression. (a) TPX2 (Affy ID: 210052_s_at). (b) KIF2C (Affy ID: 209408_at). (b) CDCA8 (Affy ID: 221520_s_at). (d) BUB1B (Affy ID: 203755_at). (es) CCNA2 (Affy ID: 213226_at)
Discussion
Breast cancer is one of the most common cancers that poses a threat to women’s life worldwide. Advanced breast cancer can spread to the whole body through blood vessels and lymphatics and directly cause cancer-related death. Although advanced therapies have achieved promising performance in controlling breast cancer with no metastasis, there are rarely available strategies to control advanced breast cancer, and even precise methods that can prevent the recurrence and metastasis of breast cancer are quite scarce [23, 24]. Therefore, it is urgent to explore potential mechanisms of metastasis and candidate biomarkers for monitoring the metastasis of breast cancer. Although Cheng et al. developed an 18-gene classifier for predicting local/regional recurrence after mastectomy and estimating distant metastasis risk after mastectomy, the complicated scoring system is difficult to widely replicate and apply clinically. Thus, in this study, we re-evaluated microarray data from GSE102484 through WGCNA and PPI analysis and finally identified five genes that exhibited the highest correlation with metastasis risk.
Co-expression analysis is an efficient strategy for gene/disease prediction analysis in large-scale datasets. In the current study, we employed WGCNA to construct a gene co-expression network, to measure the relationships between genes and modules and to investigate the relationships between modules and clinical traits. In the analysis of the top 25% most variant genes (5413 genes), the yellow module was found to show the closest correlation with the risk of metastasis as well as tumour stage, and 21 genes with high connectivity were screened from the module. We next constructed a PPI network and finally identified five (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) PPI hub genes that significantly interacted with other genes.
Targeting protein for Xenopus kinesin-like protein 2 (TPX2), which is considered to be a microtubule-associated protein, can participate in the activation of protein kinase activity, regulation of mitotic spindle tissue, apoptotic processes, mitosis, cell division and cell proliferation [25, 26]. Previous studies have shown that TPX2 is overexpressed in tumour tissues and promotes the tumorigenesis of multiple cancers [27–29]. In breast cancer cells, TPX2 promotes cancer cell invasion and migration via regulating MMP2 and MMP9 expression [30]. Additionally, silencing TPX2 can suppress cell proliferation and promote apoptosis by inhibiting PI3K/Akt signalling and activating P53 signalling [31].
Kinesin family member 2C (KIF2C) has been reported to be involved in mitosis and the cell cycle [32]. The encoded protein functions as a microtubule-dependent molecular motor that can polymerize microtubules at the plus end, thus promoting mitotic chromosome segregation. In addition, KIF2C is essential for inducing the deformation of microtubule structures and impairing cell motility [33]. In lung adenocarcinoma, KIF2C is highly expressed and is associated with the recurrence and stage of lung adenocarcinoma [34, 35]. In addition, the expression level of KIF2C mRNA has been identified as a potential independent biomarker for assessing glioma patient prognosis [36]. Accordingly, KIF2C has been reported to be a promising therapeutic target in invasive ductal carcinoma of breast cancer based on PPI network analysis [37].
Cell division cycle-associated 8 (CDCA8) is a gene encoding a component of the chromosomal passenger complex, which functions as a crucial regulator of mitosis and cell division. This protein is required for chromatin-induced microtubule stabilization and spindle formation. Overexpression of CDCA8 in tumour tissues is observed in multiple cancers, including melanoma, bladder cancer, and breast cancer [38–40]. In addition, Dai et al. revealed that the activation of CDCA8 was upregulated by the transcription factor NF-Y in multiple cancer cells [41]. These results indicate that CDCA8 might be a novel target for reducing cell division and proliferation in multiple cancers.
BUB1 mitotic checkpoint serine/threonine kinase B (BUB1B) encodes a kinase participating in the spindle checkpoint; this protein plays a critical role in the inhibition of the anaphase-promoting complex/cyclosome (APC/C) and functions in delaying the onset of anaphase and ensuring proper chromosome segregation [42]. The oncogene role of BUB1B has been observed in cancers such as prostate cancer, glioblastoma and gastric cancer [43–45]. BUB1B is overexpressed in breast cancer, and the level of BUB1B mRNA is significantly correlated with intrachromosomal instability [46]. In addition, BUB1B is preferentially expressed in high-grade breast cancer, and its expression level exhibits significant associations with long-term survival [47].
Cyclin A2 (CCNA2), a protein encoded by the corresponding gene, belongs to the highly conserved cyclin family and functions as a regulator of CDK kinases. This protein binds and activates CDC2 or CDK2 kinases and. Thus. promotes the cell cycle G1/S and G2/M transitions [48]. In breast cancer, NEK5-dependent CCNA2 overexpression promotes the proliferation of tumour cells [49]. Moreover, CCNA2 had significant predictive value for the prognosis of ER+ breast cancer patients as well as tamoxifen resistance [50].
In this research, we further assessed the differential expression of five hub genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) in various clinical stages, and the results showed that their expression was significantly increased in advanced tumour stages. In addition, the expression levels of the five genes were significantly upregulated in tumour tissues compared with normal tissues. ROC analysis revealed that the mRNA levels of the five genes showed excellent diagnostic value for breast cancer and adjacent tissues. To further assess the association between the expression of these hub genes and the risk of breast cancer metastasis, we employed KM Plotter to evaluate the prognostic value of these genes in DMFS. The primary purpose of KM Plotter is to evaluate potential biomarkers in a meta-analysis-based manner, and many prognostic biomarkers have been identified based on this platform [51–53]. Five hub genes, including TPX2, KIF2C, CDCA8, BUB1B, and CCNA2, were validated to be notably associated with the shorter DMFS of breast cancer in the patient cohort based on KM Plotter.
Conclusion
Overall, our results provide valuable indications for biomarker research on breast cancer metastasis. We predicted key metastasis-associated genes based on WGCNA and PPI analysis and validated the primary findings using data from TCGA and KM Plotter. Finally, five hub genes (TPX2, KIF2C, CDCA8, BUB1B, and CCNA2) were identified for further research, which might be used as promising biomarkers to evaluate the distant metastasis of breast cancer.
Acknowledgements
We thank Dr. Qianghu Wang and Dr. Xue Lin (Nanjing Medical University) for giving us rational advice during the study.
Availability of data and material
All data are included in the article.
Abbreviations
- ANOVA
One-way analysis of variance
- BUB1B
BUB1 mitotic checkpoint serine/threonine kinase B
- CCNA2
Cyclin A2
- CDCA8
Cell division cycle associated 8
- DMFS
Distant-metastasis-free survival
- GEO
Gene Expression Omnibus
- GS
Gene significance
- KIF2C
Kinesin family member 2C
- KM Plotter
Kaplan-Meier Plotter.
- MM
Module membership
- MVGs
Most variant genes
- PPI
Protein-protein interaction
- ROC
Receiver operating characteristic
- TCGA
The Cancer Genome Atlas
- TOM
Topological overlap matrix
- TPX2
Targeting protein for Xenopus kinesin-like protein 2
- WGCNA
Weighted gene co-expression network analysis
Authors’ contributions
Y Zhang and Y Zhu conceived the study and participated in the study design, performance, coordination and manuscript writing. YC, JM, ZX, BX, and XJ carried out the assays and analysis. JM, Y Zhang and Y Zhu revised the manuscript. All authors reviewed and approved the final manuscript.
Funding
Not applicable.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Yun Cai, Jie Mei and Zhuang Xiao contributed equally to this work and should be considered co-first authors.
Contributor Information
Yun Cai, Email: yuncai@163.com.
Jie Mei, Email: meijie199621@163.com.
Zhuang Xiao, Email: xiaozhuang_325@163.com.
Bujie Xu, Email: bujiexu192@163.com.
Xiaozheng Jiang, Email: xiaoyubanana@163.com.
Yongjie Zhang, Phone: +86-25-86869485, Email: zhangyongjie@njmu.edu.cn.
Yichao Zhu, Phone: +86-25-86868318, Email: zhuyichao@njmu.edu.cn.
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69:7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 2.Dean A. Primary breast cancer: risk factors, diagnosis and management. Nurs Stand. 2008;22:47–55. doi: 10.7748/ns2008.06.22.40.47.c6572. [DOI] [PubMed] [Google Scholar]
- 3.Peart O. Breast intervention and breast cancer treatment options. Radiol Technol. 2015;86:535M–558M. [PubMed] [Google Scholar]
- 4.Shen T, Gao C, Zhang K, Siegal GP, Wei S. Prognostic outcomes in advanced breast cancer: the metastasis-free interval is important. Hum Pathol. 2017;70:70–76. doi: 10.1016/j.humpath.2017.10.002. [DOI] [PubMed] [Google Scholar]
- 5.Li SC, Stucky A, Chen X, Kabeer MH, Loudon WG, Plant AS, Torno L, Nangia CS, Cai J, Zhang G, Zhong JF. Single-cell transcriptomes reveal the mechanism for a breast cancer prognostic gene panel. Oncotarget. 2018;9:33290–33301. doi: 10.18632/oncotarget.26044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Orton RJ, Gu Q, Hughes J, Maabar M, Modha S, Vattipally SB, Wilkie GS, Davison AJ. Bioinformatics tools for analysing viral genomic data. Rev Sci Tech. 2016;35:271–285. doi: 10.20506/rst.35.1.2432. [DOI] [PubMed] [Google Scholar]
- 7.Chen J, Coppola G. Bioinformatics and genomic databases. Handb Clin Neurol. 2018;147:75–92. doi: 10.1016/B978-0-444-63233-3.00007-5. [DOI] [PubMed] [Google Scholar]
- 8.Heng HH. Cancer genome sequencing: the challenges ahead. Bioessays. 2007;29:783–794. doi: 10.1002/bies.20610. [DOI] [PubMed] [Google Scholar]
- 9.Hanauer DA, Rhodes DR, Sinha-Kumar C, Chinnaiyan AM. Bioinformatics approaches in the study of cancer. Curr Mol Med. 2007;7:133–141. doi: 10.2174/156652407779940431. [DOI] [PubMed] [Google Scholar]
- 10.Li SC, Tachiki LM, Kabeer MH, Dethlefs BA, Anthony MJ, Loudon WG. Cancer genomic research at the crossroads: realizing the changing genetic landscape as intratumoral spatial and temporal heterogeneity becomes a confounding factor. Cancer Cell Int. 2014;14:115. doi: 10.1186/s12935-014-0115-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics. 2008;9:559. doi: 10.1186/1471-2105-9-559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chin CH, Chen SH, Wu HH, Ho CW, Ko MT, Lin CY. cytoHubba: identifying hub objects and sub-networks from complex interactome. BMC Syst Biol. 2014;8:S11. doi: 10.1186/1752-0509-8-S4-S11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Martin-Antoniano I, Alonso L, Madrid M, Lopez de Maturana E, Malats N. DoriTool: a bioinformatics integrative tool for post-association functional annotation. Public Health Genomics. 2017;20:126–135. doi: 10.1159/000477561. [DOI] [PubMed] [Google Scholar]
- 14.Tang J, Kong D, Cui Q, Wang K, Zhang D, Gong Y, Wu G. Prognostic genes of breast Cancer identified by gene co-expression network analysis. Front Oncol. 2018;8:374. doi: 10.3389/fonc.2018.00374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu M, Qiu YL, Jin T, Zhou Y, Mao ZY, Zhang YJ. Meta-analysis of microarray datasets identify several chromosome segregation-related cancer/testis genes potentially contributing to anaplastic thyroid carcinoma. PeerJ. 2018;6:e5822. doi: 10.7717/peerj.5822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guo X, Xiao H, Guo S, Dong L, Chen J. Identification of breast cancer mechanism based on weighted gene coexpression network analysis. Cancer Gene Ther. 2017;24:333–341. doi: 10.1038/cgt.2017.23. [DOI] [PubMed] [Google Scholar]
- 17.Qiu J, Du Z, Wang Y, Zhou Y, Zhang Y, Xie Y, Lv Q. Weighted gene co-expression network analysis reveals modules and hub genes associated with the development of breast cancer. Medicine (Baltimore) 2019;98:e14345. doi: 10.1097/MD.0000000000014345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cheng SH, Huang TT, Cheng YH, Tan TBK, Horng CF, Wang YA, Brian NS, Shih LS, Yu BL. Validation of the 18-gene classifier as a prognostic biomarker of distant metastasis in breast cancer. PLoS One. 2017;12:e0184372. doi: 10.1371/journal.pone.0184372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cheng SH, Horng CF, West M, Huang E, Pittman J, Tsou MH, Dressman H, Chen CM, Tsai SY, Jian JJ, Liu MC, Nevins JR, Huang AT. Genomic prediction of locoregional recurrence after mastectomy in breast cancer. J Clin Oncol. 2006;24:4594–4602. doi: 10.1200/JCO.2005.02.5676. [DOI] [PubMed] [Google Scholar]
- 20.Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos A, Tsafou KP, Kuhn M, Bork P, Jensen LJ, von Mering C. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015;43:D447–D452. doi: 10.1093/nar/gku1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lanczky A, Nagy A, Bottai G, Munkacsy G, Szabo A, Santarpia L, Gyorffy B. miRpower: a web-tool to validate survival-associated miRNAs utilizing expression data from 2178 breast cancer patients. Breast Cancer Res Treat. 2016;160:439–446. doi: 10.1007/s10549-016-4013-7. [DOI] [PubMed] [Google Scholar]
- 22.Wang C, Gu Y, Zhang K, Xie K, Zhu M, Dai N, Jiang Y, Guo X, Liu M, Dai J, Wu L, Jin G, Ma H, Jiang T, Yin R, Xia Y, Liu L, Wang S, Shen B, Huo R, Wang Q, Xu L, Yang L, Huang X, Shen H, Sha J, Hu Z. Systematic identification of genes with a cancer-testis expression pattern in 19 cancer types. Nat Commun. 2016;7:10499. doi: 10.1038/ncomms10499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Weigelt B, Peterse JL, van ‘t Veer LJ. Breast cancer metastasis: markers and models. Nat Rev Cancer. 2005;5:591–602. doi: 10.1038/nrc1670. [DOI] [PubMed] [Google Scholar]
- 24.McGuire A, Brown JA, Kerin MJ. Metastatic breast cancer: the potential of miRNA for diagnosis and treatment monitoring. Cancer Metastasis Rev. 2015;34:145–155. doi: 10.1007/s10555-015-9551-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wieczorek M, Bechstedt S, Chaaban S, Brouhard GJ. Microtubule-associated proteins control the kinetics of microtubule nucleation. Nat Cell Biol. 2015;17:907–916. doi: 10.1038/ncb3188. [DOI] [PubMed] [Google Scholar]
- 26.Janecek M, Rossmann M, Sharma P, Emery A, Huggins DJ, Stockwell SR, Stokes JE, Tan YS, Almeida EG, Hardwick B, Narvaez AJ, Hyvonen M, Spring DR, McKenzie GJ, Venkitaraman AR. Allosteric modulation of AURKA kinase activity by a small-molecule inhibitor of its protein-protein interaction with TPX2. Sci Rep. 2016;6:28528. doi: 10.1038/srep28528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tomii C, Inokuchi M, Takagi Y, Ishikawa T, Otsuki S, Uetake H, Kojima K, Kawano T. TPX2 expression is associated with poor survival in gastric cancer. World J Surg Oncol. 2017;15:14. doi: 10.1186/s12957-016-1095-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Glaser ZA, Love HD, Guo S, Gellert L, Chang SS, Herrell SD, Barocas DA, Penson DF, Cookson MS, Clark PE. TPX2 as a prognostic indicator and potential therapeutic target in clear cell renal cell carcinoma. Urol Oncol. 2017;35:286–293. doi: 10.1016/j.urolonc.2016.12.012. [DOI] [PubMed] [Google Scholar]
- 29.Pan HW, Su HH, Hsu CW, Huang GJ, Wu TT. Targeted TPX2 increases chromosome missegregation and suppresses tumor cell growth in human prostate cancer. Onco Targets Ther. 2017;10:3531–3543. doi: 10.2147/OTT.S136491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yang Y, Li DP, Shen N, Yu XC, Li JB, Song Q, Zhang JH. TPX2 promotes migration and invasion of human breast cancer cells. Asian Pac J Trop Med. 2015;8:1064–1070. doi: 10.1016/j.apjtm.2015.11.007. [DOI] [PubMed] [Google Scholar]
- 31.Chen M, Zhang H, Zhang G, Zhong A, Ma Q, Kai J, Tong Y, Xie S, Wang Y, Zheng H, Guo L, Lu R. Targeting TPX2 suppresses proliferation and promotes apoptosis via repression of the PI3k/AKT/P21 signaling pathway and activation of p53 pathway in breast cancer. Biochem Biophys Res Commun. 2018;507:74–82. doi: 10.1016/j.bbrc.2018.10.164. [DOI] [PubMed] [Google Scholar]
- 32.Manning AL, Ganem NJ, Bakhoum SF, Wagenbach M, Wordeman L, Compton DA. The kinesin-13 proteins Kif2a, Kif2b, and Kif2c/MCAK have distinct roles during mitosis in human cells. Mol Biol Cell. 2007;18:2970–2979. doi: 10.1091/mbc.e07-02-0110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jung YD, Cho JH, Park S, Kang M, Park SJ, Choi DH, Jeong M, Park KC, Yeom YI, Lee DC. Lactate activates the E2F pathway to promote cell motility by up-regulating microtubule modulating genes. Cancers (Basel). 2019:11. [DOI] [PMC free article] [PubMed]
- 34.Bai Y, Xiong L, Zhu M, Yang Z, Tang H, Zhao J. Co-expression network analysis identified KIF2C in association with progression and prognosis in lung adenocarcinoma. Cancer Biomark. 2019. [DOI] [PubMed]
- 35.Bai Y, Xiong L, Zhu M, Yang Z, Zhao J, Tang H. Co-expression network analysis identified KIF2C in association with progression and prognosis in lung adenocarcinoma. Cancer Biomark. 2019;24:371–382. doi: 10.3233/CBM-181512. [DOI] [PubMed] [Google Scholar]
- 36.Bie L, Zhao G, Wang YP, Zhang B. Kinesin family member 2C (KIF2C/MCAK) is a novel marker for prognosis in human gliomas. Clin Neurol Neurosurg. 2012;114:356–360. doi: 10.1016/j.clineuro.2011.11.005. [DOI] [PubMed] [Google Scholar]
- 37.Li C, Luo L, Wei S, Wang X. Identification of the potential crucial genes in invasive ductal carcinoma using bioinformatics analysis. Oncotarget. 2018;9:6800–6813. doi: 10.18632/oncotarget.23239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Phan NN, Wang CY, Li KL, Chen CF, Chiao CC, Yu HG, Huang PL, Lin YC. Distinct expression of CDCA3, CDCA5, and CDCA8 leads to shorter relapse free survival in breast cancer patient. Oncotarget. 2018;9:6977–6992. doi: 10.18632/oncotarget.24059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bi Y, Chen S, Jiang J, Yao J, Wang G, Zhou Q, Li S. CDCA8 expression and its clinical relevance in patients with bladder cancer. Medicine (Baltimore) 2018;97:e11899. doi: 10.1097/MD.0000000000011899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ci C, Tang B, Lyu D, Liu W, Qiang D, Ji X, Qiu X, Chen L, Ding W. Overexpression of CDCA8 promotes the malignant progression of cutaneous melanoma and leads to poor prognosis. Int J Mol Med. 2019;43:404–412. doi: 10.3892/ijmm.2018.3985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dai C, Miao CX, Xu XM, Liu LJ, Gu YF, Zhou D, Chen LS, Lin G, Lu GX. Transcriptional activation of human CDCA8 gene regulated by transcription factor NF-Y in embryonic stem cells and cancer cells. J Biol Chem. 2015;290:22423–22434. doi: 10.1074/jbc.M115.642710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Vleugel M, Hoek TA, Tromer E, Sliedrecht T, Groenewold V, Omerzu M, Kops GJ. Dissecting the roles of human BUB1 in the spindle assembly checkpoint. J Cell Sci. 2015;128:2975–2982. doi: 10.1242/jcs.169821. [DOI] [PubMed] [Google Scholar]
- 43.Ma Q, Liu Y, Shang L, Yu J, Qu Q. The FOXM1/BUB1B signaling pathway is essential for the tumorigenicity and radioresistance of glioblastoma. Oncol Rep. 2017;38:3367–3375. doi: 10.3892/or.2017.6032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fu X, Chen G, Cai ZD, Wang C, Liu ZZ, Lin ZY, Wu YD, Liang YX, Han ZD, Liu JC, Zhong WD. Overexpression of BUB1B contributes to progression of prostate cancer and predicts poor outcome in patients with prostate cancer. Onco Targets Ther. 2016;9:2211–2220. doi: 10.2147/OTT.S100633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hudler P, Britovsek NK, Grazio SF, Komel R. Association between polymorphisms in segregation genes BUB1B and TTK and gastric cancer risk. Radiol Oncol. 2016;50:297–307. doi: 10.1515/raon-2015-0047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Scintu M, Vitale R, Prencipe M, Gallo AP, Bonghi L, Valori VM, Maiello E, Rinaldi M, Signori E, Rabitti C, Carella M, Dallapiccola B, Altomare V, Fazio VM, Parrella P. Genomic instability and increased expression of BUB1B and MAD2L1 genes in ductal breast carcinoma. Cancer Lett. 2007;254:298–307. doi: 10.1016/j.canlet.2007.03.021. [DOI] [PubMed] [Google Scholar]
- 47.Mukherjee A, Joseph C, Craze M, Chrysanthou E, Ellis IO. The role of BUB and CDC proteins in low-grade breast cancers. Lancet. 2015;385(Suppl 1):S72. doi: 10.1016/S0140-6736(15)60387-7. [DOI] [PubMed] [Google Scholar]
- 48.Zhang QH, Yuen WS, Adhikari D, Flegg JA, FitzHarris G, Conti M, Sicinski P, Nabti I, Marangos P, Carroll J. Cyclin A2 modulates kinetochore-microtubule attachment in meiosis II. J Cell Biol. 2017;216:3133–3143. doi: 10.1083/jcb.201607111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pei J, Zhang J, Yang X, Wu Z, Sun C, Wang Z, Wang B. NEK5 promotes breast cancer cell proliferation through up-regulation of cyclin A2. Mol Carcinog. 2019. [DOI] [PubMed]
- 50.Gao T, Han Y, Yu L, Ao S, Li Z, Ji J. CCNA2 is a prognostic biomarker for ER+ breast cancer and tamoxifen resistance. PLoS One. 2014;9:e91771. doi: 10.1371/journal.pone.0091771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mei J, Yan T, Huang Y, Xia T, Chang F, Shen S, Hao L, Chen Y, Wang Z, Jiang X, Xu B, Zhu Y. A DAAM1 3′-UTR SNP mutation regulates breast cancer metastasis through affecting miR-208a-5p-DAAM1-RhoA axis. Cancer Cell Int. 2019;19:55. doi: 10.1186/s12935-019-0747-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Manicum Theasha, Ni Fubiao, Ye Yiming, Fan Xuhui, Chen Bi-Cheng. Prognostic values of E2F mRNA expression in human gastric cancer. Bioscience Reports. 2018;38(6):BSR20181264. doi: 10.1042/BSR20181264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gayed DT, Wodeyar J, Wang ZX, Wei X, Yao YY, Chen XX, Du Z, Chen JC. Prognostic values of inhibitory kappaB kinases mRNA expression in human gastric cancer. Biosci Rep. 2019;39. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data are included in the article.