

Abstract
Wolbachia are maternally inherited bacteria that infect arthropod species worldwide and are deployed in vector control to curb arboviral spread using cytoplasmic incompatibility (CI). CI kills embryos when an infected male mates with an uninfected female, but the lethality is rescued if the female and her embryos are likewise infected. Two phage WO genes, cifAwMel and cifBwMel from the wMel Wolbachia deployed in vector control, transgenically recapitulate variably penetrant CI, and one of the same genes, cifAwMel, rescues wild type CI. The proposed Two-by-One genetic model predicts that CI and rescue can be recapitulated by transgenic expression alone and that dual cifAwMel and cifBwMel expression can recapitulate strong CI. Here, we use hatch rate and gene expression analyses in transgenic Drosophila melanogaster to demonstrate that CI and rescue can be synthetically recapitulated in full, and strong, transgenic CI comparable to wild type CI is achievable. These data explicitly validate the Two-by-One model in wMel-infected D. melanogaster, establish a robust system for transgenic studies of CI in a model system, and represent the first case of completely engineering male and female animal reproduction to depend upon bacteriophage gene products.
Author summary
Releases of Wolbachia-infected mosquitos are underway worldwide because Wolbachia block replication of Zika and Dengue viruses and spread themselves maternally through arthropod populations via cytoplasmic incompatibility (CI). The CI drive system depends on a Wolbachia-induced sperm modification that results in embryonic lethality when an infected male mates with an uninfected female, but this lethality is rescued when the female and her embryos are likewise infected. We recently reported that the phage WO genes, cifA and cifB, cause the sperm modification and cifA rescues the embryonic lethality caused by the wMel Wolbachia strain deployed in vector control. These reports motivated proposal of the Two-by-One model of CI whereby two genes cause lethality and one gene rescues it. Here we provide unequivocal support for the model in the Wolbachia strain used in vector control via synthetic methods that recapitulate CI and rescue in the absence of a Wolbachia infections. Our results reveal the set of phage WO genes responsible for this powerful genetic drive system, act as a proof-of-concept that these genes alone can induce gene drive like crossing patterns, and establish methodologies and hypotheses for future studies of CI in Drosophila. We discuss the implications of the Two-by-One model towards functional mechanisms of CI, the emergence of incompatibility between Wolbachia strains, vector control applications, and CI gene nomenclature.
Introduction
Wolbachia are the most widespread endosymbiotic bacteria on the planet and are estimated to infect half of all arthropod species [1,2] and half of the Onchocercidae family of filarial nematodes [3]. They specialize in infecting the cells of reproductive tissues, are primarily inherited maternally from ova to offspring, and often act in arthropods as reproductive parasites that enhance their maternal transmission by distorting host sex ratios and reproduction [4,5]. The most common type of reproductive parasitism is cytoplasmic incompatibility (CI), which manifests as a sperm modification in infected males that causes embryonic lethality or haploidization in matings with uninfected females upon fertilization [6–8]. This embryonic lethality is rescued if the female is infected with the same Wolbachia strain. As such, CI selfishly drives CI-inducing Wolbachia into host populations [9–13], and the incompatibilities between host populations cause reproductive isolation between recently diverged or incipient species [14–18].
In the last decade, Wolbachia and CI have garnered significant interest for their utility in combatting vector borne diseases worldwide. Two strategies are currently deployed: population suppression and population replacement. The population suppression strategy markedly crashes vector population sizes through the release of only infected males that induce CI upon mating with wild uninfected females [19–22]. In contrast, the population replacement strategy converts uninfected to infected populations through the release of both infected males and females that aid the spread Wolbachia via CI and rescue [23,24]. Replacing a vector competent, uninfected population with infected individuals can notably reduce the spread of arthropod borne diseases such as Zika and dengue [25,26] because Wolbachia appear to inhibit various stages of viral replication within arthropods based on diverse manipulations of the host cellular environment [27–33]. The combination of Wolbachia’s abilities to suppress arthropod populations, drive into host populations, and block the spread of viral pathogens have established Wolbachia in the vanguard of vector control efforts to curb arboviral transmission [22–25,34–36].
An unbiased, multi-omic analysis of CI-inducing and CI-incapable Wolbachia strains revealed two adjacent genes, cifA and cifB, in the eukaryotic association module of prophage WO [37] that strictly associate with CI induction [38]. Fragments of the CifA protein were found in the fertilized spermathecae of wPip infected Culex pipiens mosquitoes [39], and these genes are frequently missing or degraded in diverse CI-incapable strains [40,41]. Dual transgenic expression of cifA and cifB from either of the CI-inducing strains wMel or wPip in uninfected male flies causes a decrease in embryonic hatching corresponding to an increase in CI-associated cytological abnormalities including chromatin bridging and regional mitotic failures [38,42]. Single transgenic expression of either cifAwMel or cifBwMel in an uninfected male was insufficient to recapitulate CI, but single transgenic expression of either gene in an infected male enhances wMel-induced CI in a dose-dependent manner [38]. Importantly, dual transgenic CI induced by cifAwMel and cifBwMel expressing males was rescued when they were mated with wMel-infected females [38]. Moreover, transgenic expression of cifAwMel alone in uninfected females rescues embryonic lethality and nullifies cytological defects associated with wild type CI caused by a wMel infection [43].
As such, we recently proposed the Two-by-One genetic model of CI wherein dual expression of cifAwMel and cifBwMel causes CI when expressed in males and expression of cifAwMel rescues CI when expressed in females [43]. However, confirmation of the model’s central prediction requires the complete synthetic replication of CI-induced lethality and rescue in the absence of any Wolbachia infections since it remains possible that other Wolbachia or phage WO genes besides cifA and cifB contribute to wild type CI and rescue by wMel Wolbachia. Moreover, CI induced by dual cifAwMel and cifBwMel expression previously yielded variable offspring lethality with a median survival of 26.5% of embryos relative to survival of 0.0% of embryos from CI induced by a wild type infection under controlled conditions [38]. The inability to recapitulate strong wild type CI suggests other CI genes are required, other environmental factors need to be controlled, or the transgenic system requires optimization.
Here, we utilize transgenic expression, hatch rates, and gene expression assays in Drosophila melanogaster to test if an optimized expression system can generate strong transgenic CI and whether bacteriophage genes cifAwMel and cifBwMel can fully control fly reproduction by inducing and rescuing CI in the complete absence of Wolbachia (Fig 1). We further assess if both cifwMel genes are required for CI induction in the optimized system and whether cifAwMel in females can rescue transgenic CI. Results provide strong evidence for the Two-by-One model in wMel-infected D. melanogaster, offer context for conceptualizing CI mechanisms and the evolution of bidirectional incompatibilities between different Wolbachia strains, raise points for CI gene nomenclature, and motivate further research in developing these genes into a tool that combats vector borne diseases. To the best of our knowledge, they also represent the first case of completely engineering animal sexual reproduction to depend upon bacteriophage gene products.
Fig 1. Two-by-One model of CI is governed by cifA and cifB genes in the eukaryotic association module of prophage WO in Wolbachia.
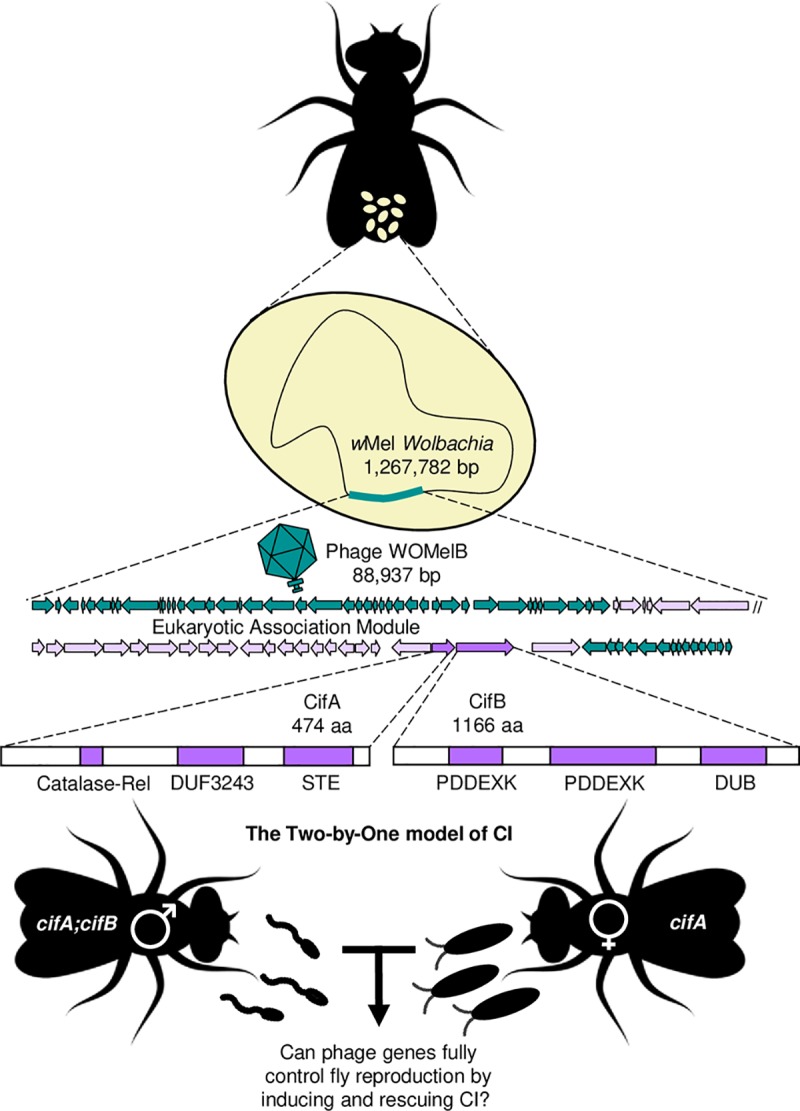
The Two-by-One model of CI predicts that D. melanogaster males and females can be engineered to recapitulate both CI and rescue phenotypes in the absence of Wolbachia, thus depending completely on phage genes for successful reproduction. Schematics are not to scale. Insect, sperm, and embryo art were obtained and modified using vecteezy.com. Phage gene schematics modified from [38]. CifA and CifB protein annotation from [40]. Purple indicates eukaryotic association module genes as indicated by [37].
Results
Optimizing transgenic CI
Dual transgenic expression of cifAwMel and cifBwMel was previously reported to induce highly variable and incomplete CI relative to CI caused by an age-controlled wMel infection [38], indicating either the presence of other genes necessary for strong CI, environmental factors uncontrolled in the study, or inefficiency of the transgenic system. Here, we test the latter hypothesis by dually expressing cifAwMel and cifBwMel in uninfected D. melanogaster males under two distinct GAL4 driver lines that express in reproductive tissues: nos-GAL4-tubulin and nos-GAL4:VP16 [44]. Both driver lines contain a nos promoter region, but differ in that nos-GAL4-tubulin produces a transcription factor with both the DNA binding and transcriptional activating region of the GAL4 protein, and nos-GAL4:VP16 produces a fusion protein of the GAL4 DNA binding domain and the virion protein 16 (VP16) activating region [45,46]. The GAL4:VP16 transcription factor is a particularly potent transcriptional activator because of its binding efficiency to transcription factors [47,48]. Additionally, the nos-GAL4-tubulin driver has a tubulin 3’ UTR, and nos-GAL4:VP16 has a nos 3’ UTR that may contribute to differences in localization within cells or between tissues [44–46]. As such, we predict that differences in the expression level or profile of these two driver lines will lead to differences in the penetrance of transgenic CI.
Since CI manifests as embryonic lethality, we measure hatching of D. melanogaster embryos into larvae to quantify the strength of CI. We confirm previous findings [38] that dual transgenic expression of cifAwMel and cifBwMel under nos-GAL4-tubulin in uninfected males yields low but variable embryonic hatching in crosses with uninfected females (Mdn = 26.3%, IQR = 10.4–38.1%) that can be rescued in crosses with wMel-infected females (Mdn = 97.5%; IQR = 94.2–100%) (Fig 2A). However, dual cifAwMel and cifBwMel expression under nos-GAL4:VP16 in uninfected males yields significantly reduced embryonic hatching relative to nos-GAL4-tubulin (p = 0.0002) with less variability (Mdn = 0%; IQR = 0.0–0.75%) and can be comparably rescued (Mdn = 98.65%; IQR = 95.93–100%; p > 0.99) (Fig 2A). Together, these results support that dual cifAwMel and cifBwMel expression under nos-GAL4:VP16 induces the strongest CI and that the transgenic system, not the absence of necessary CI factors, contributed to the prior inability to recapitulate strong wild type CI.
Fig 2. cifAwMel and cifBwMel induce strong CI when transgenically expressed in males under the nos-GAL4:VP16 driver.

(A) Two different driver lines, nos-GAL4-tubulin (purple; top) and nos-GAL4:VP16 (green; bottom) were tested for their ability to induce CI when transgenically expressed in uninfected, male Drosophila. Filled sex symbols represent infection with wMel Wolbachia, and gene names to the right of a symbol represent expression of those genes in the male line. Vertical bars represent medians. Letters to the right indicate significant differences with an α = 0.05 calculated by a Kruskal-Wallis analysis followed by Dunn’s multiple comparison test. (B,C) To test if nos-GAL4-tubulin and nos-GAL4:VP16 generate different levels of gene expression, (B) cifAwMel and (C) cifBwMel fold expression difference relative to the Drosophila housekeeping gene rp49 in male abdomens under the two drivers was measured using qPCR. Males tested for gene expression were the same used in the hatch rate experiment in A. Letters above indicate significant differences with an α = 0.05 calculated by a Mann-Whitney U test.
Next, we tested the hypothesis that differences in the penetrance of transgenic CI between the two drivers are due to differences in the strength of expression. To assess this, we used qPCR to measure the gene expression of cifAwMel and cifBwMel under the two drivers relative to a Drosophila housekeeping gene (rp49) in male abdomens (Fig 2B and 2C). Fold differences in RNA transcripts of cifAwMel relative to rp49 reveal nos-GAL4-tubulin (Mdn = 0.0098; IQR = 0.0082–0.122) drives significantly stronger and more variable cifAwMel expression relative to nos-GAL4:VP16 (Mdn = 0.0075; IQR = 0.0064–0.0090) (p = 0.016, MWU, Fig 2B). The same is true for cifBwMel expression where nos-GAL4-tubulin (Mdn = 0.022; IQR = 0.0165–0.0265) drives significantly stronger cifBwMel expression than nos-GAL4:VP16 (Mdn = 0.0168; IQR = 0.0135–0.0179) (p = 0.02, MWU, Fig 2C). Moreover, while cifAwMel and cifBwMel expression significantly correlate with each other under both nos-GAL4-tubulin (R2 = 0.85; p <0.0001) and nos-GAL4:VP16 (R2 = 0.75; p <0.0001; S1A Fig), neither cifAwMel (R2 = 0.02; p = 0.62; S1B Fig) nor cifBwMel (R2 = 0.04; p = 0.48; S1C Fig) expression levels under the nos-GAL4-tubulin driver correlate with the strength of CI measured via hatch rates. Notably, cifBwMel is consistently more highly expressed than cifAwMel within the same line (S1A Fig). We predict that expression differences are due to either differences in transgenic insertion sites or more rapid degradation of cifAwMel relative to cifBwMel. Taken together, these results suggest that an increase in CI penetrance in these crosses is not positively associated with higher transgene transcript abundance from different drivers.
Optimizing transgenic rescue
cifAwMel expression under the maternal triple driver (MTD) in uninfected females can rescue CI induced by a wild type infection [43]. MTD is comprised of three drivers in the same line: nos-GAL4-tubulin, nos-GAL4:VP16, and otu-GAL4:VP16 [44]. We previously reported that cifAwMel expression under the nos-GAL4-tubulin driver alone is rescue-incapable [43]. Here, we test if cifAwMel expression under either of the other components of the MTD driver independently recapitulate rescue of wMel CI. Hatch rate experiments indicate that CI is strong and expectedly not rescued when an infected male mates with a non-transgenic female whose genotype is otherwise nos-GAL4:VP16 (Mdn = 0.0%; IQR = 0.0–0.0%) or otu-GAL4:VP16 (Mdn = 0.0%; IQR = 0.0–0.0%) (Fig 3A). Transgenic expression of cifAwMel in uninfected females under either of the two drivers rescues CI induced by wMel. However, rescue is significantly weaker under cifAwMel expression with the otu-GAL4:VP16 driver (Mdn = 70.4%; IQR = 0.0–90.45%) as compared to the nos-GAL4:VP16 driver (Mdn = 94.2%; IQR = 83.3–97.1%; p = 0.0491) which produced strong transgenic rescue (Fig 3A). Gene expression analysis of cifAwMel relative to rp49 in the abdomens of uninfected females reveals that nos-GAL4:VP16 expresses cifAwMel significantly higher (Mdn = 1.08; p < 0.0001) than otu-GAL4:VP16 (Mdn = 0.03) (Fig 3B), suggesting that high expression in females may underpin the ability to rescue. Alternatively, nos-GAL4:VP16 and otu-GAL4:VP16 are known to express GAL4 at different times in oogenesis, with the former in all egg chambers and the latter in late stage egg chambers [44].
Fig 3. cifAwMel can induce strong rescue when expressed in uninfected females under the nos-GAL4:VP16 driver.

(A) Two different driver lines, nos-GAL4:VP16 (green; top) and otu-GAL4:VP16 (pink; bottom), were tested for their ability to rescue wMel induced CI. Filled sex symbols represent infection with wMel Wolbachia, and gene names to the right of a symbol represent expression of those genes in the corresponding sex of that cross. Vertical bars represent medians. Letters to the right indicate significant differences with an α = 0.05 calculated by a Kruskal-Wallis analysis followed by Dunn’s multiple comparison test. (B) To test if nos-GAL4-tubulin and nos-GAL4:VP16 generate different levels of RNA expression, cifAwMel fold expression difference relative to the Drosophila housekeeping gene rp49 in male abdomens under the two drivers was measured using qPCR. Females tested for gene expression were the same used in the hatch rate experiment in A. Letters above indicate significant differences with an α = 0.05 calculated by a Mann-Whitney U test.
The Two-by-One model of CI
With the transgenic expression system optimized for both transgenic CI and rescue, we then tested the hypothesis that the Two-by-One model can be synthetically recapitulated by dual cifAwMel and cifBwMel expression in uninfected males to cause CI and single cifAwMel expression in uninfected females to rescue that transgenic CI. Indeed, dual cifAwMel and cifBwMel expression in uninfected males causes hatch rates comparable to wild type CI (Mdn = 0.0%; IQR = 0.0%-2.55; p > 0.99) (Fig 4). Transgenic CI cannot be rescued by single cifBwMel expression in uninfected females (Mdn = 1.25%; IQR = 0.0–3.35%). Transgenic CI can be rescued by single cifAwMel expression (Mdn = 98.6%; IQR = 97.35–100%; p = 0.41) or dual cifAwMel and cifBwMel expression (Mdn = 96.7%; IQR = 88.3–98.2%; p > 0.99) to levels comparable to rescue from a wild type infection (Mdn = 95.6%; IQR = 92.5–97.4%). In addition, cifAwMel rescues a wild type infection at comparable levels to wild type rescue (Mdn = 96.6%; IQR = 93.5–98.85%; p > 0.99). These data provide strong evidence for the Two-by-One model in wMel-infected D. melanogaster, namely that CI induced by transgenic dual cifAwMel and cifBwMel expression is sufficient to induce strong CI, and that cifAwMel alone is sufficient to rescue it.
Fig 4. CI and rescue can be synthetically recapitulated under transgenic expression in the absence of Wolbachia.

Single cifAwMel and dual cifAwMel and cifBwMel expression under nos-GAL4:VP16 in uninfected females (open circles) were tested for their ability to rescue transgenic CI under the same driver in uninfected males. Filled sex symbols represent infection with wMel Wolbachia, and gene names beside a symbol represent expression of those genes in the corresponding sex of that cross. Vertical bars represent medians. Letters to the right indicate significant differences with an α = 0.05 calculated by a Kruskal-Wallis analysis followed by Dunn’s multiple comparison test.
Next we reevaluated if single cifAwMel or cifBwMel expression under the more potent nos-GAL4:VP16 driver in uninfected males can recapitulate CI. Hatch rates indicate that dual cifAwMel and cifBwMel expression induces strong transgenic CI (Mdn = 0.0%; IQR = 0.0–1.15%) that can be rescued by a wild type infection (Mdn = 93.8%; IQR = 88.2–97.4%), whereas single expression of cifAwMel (Mdn = 96.1%; IQR = 97.78–98.55%; p < 0.0001) or cifBwMel (Mdn = 92.85%; IQR = 84.28–96.4%; p < 0.0001) failed once again to produce embryonic hatching comparable to expressing both genes together (Fig 5). In one replicate experiment, we note a statistically insignificant (p = 0.182) decrease in hatching under cifBwMel expression relative to wild type rescue cross (S1 Data file). Thus, both cifAwMel and cifBwMel are required for strong CI. Together, these and earlier results validate the Two-by-One model of CI in wMel whereby cifAwMel and cifBwMel expression are required and sufficient for strong CI, while cifAwMel expression is sufficient to rescue it.
Fig 5. Neither cifAwMel or cifBwMel alone can induce CI when expressed under nos-GAL4:VP16.

cifAwMel and cifBwMel were tested for their ability to induce CI individually under nos-GAL4:VP16 expression in uninfected males (open circles). Filled sex symbols represent infection with wMel Wolbachia and gene names to the right of a symbol represent expression of those genes in the corresponding sex of that cross. Vertical bars represent medians. Letters to the right indicate significant differences with an α = 0.05 calculated by a Kruskal-Wallis analysis followed by Dunn’s multiple comparison test.
Discussion
CI is the most common form of Wolbachia-induced reproductive parasitism and is currently at the forefront of vector control efforts to curb transmission of dengue, Zika, and other arthropod-borne human pathogens [22–25,34,35]. Two prophage WO genes from wMel Wolbachia cause CI (cifAwMel and cifBwMel) and one rescues wild type CI (cifAwMel) [38,43], supporting the proposal of a Two-by-One model for the genetic basis of CI [43]. However, dual transgenic expression of cifAwMel and cifBwMel recapitulates only weak and highly variable CI as compared to CI induced by a wild type infection [38]. In addition, the Two-by-One model predicts that both CI and rescue can be synthetically recapitulated by dual cifAwMel and cifBwMel expression in uninfected males and cifAwMel expression in uninfected females. Here we optimized the transgenic system for CI and rescue by these genes, further validated the necessity of expressing both cifAwMel and cifBwMel for CI, and synthetically recapitulated the Two-by-One model for CI with transgenics in the absence of Wolbachia.
CI induced by wMel Wolbachia can be highly variable and correlates with numerous factors including Wolbachia density [49], cifAwMel and cifBwMel expression levels [38], host age [50–52], mating rate [50], rearing density [53], development time [53], and host genetic factors [52,54–56]. Some of these factors, such as age, are known to also correlate with the level of cifwMel gene expression [38]. As such, we hypothesized that prior reports of weakened transgenic CI could be explained by low levels of transgenic cifAwMel and cifBwMel expression in male testes [38].
Indeed, strong CI with a median of 0% embryonic hatching was induced when both cifAwMel and cifBwMel were expressed under the nos-GAL4:VP16 driver. However, contrary to our expectations, nos-GAL4:VP16 generates significantly weaker cifAwMel and cifBwMel expression than the nos-GAL4-tubulin driver previously used to recapitulate weak CI [38]. Thus, the expression data conflict with previous reports in mammalian cells wherein the GAL4:VP16 fusion protein is a more potent transcriptional activator than GAL4 [48]. Other differences between the two driver constructs may explain phenotypic differences, including the presence of different 3’ UTRs that may contribute to differences in transcript localization [44]. While it remains possible, though unlikely, that other Wolbachia or phage WO genes may contribute to CI, the induction of near complete embryonic lethality confirms that cifAwMel and cifBwMel are sufficient to transgenically induce strong CI and do not require other Wolbachia or phage WO genes to do so. Moreover, comparative multi-omics demonstrated that cifA and cifB are the only two genes strictly associated with CI capability [38].
We previously recapitulated transgenic rescue of wMel-induced CI by expression of cifAwMel under the Maternal Triple Driver (MTD) [43], which is comprised of three independent drivers [44]. Expression of cifAwMel using one of the MTD drivers in flies was previously shown to be rescue-incapable [43]; the other drivers had not been evaluated. Here, we tested the hypothesis that expression of cifAwMel using either of the two remaining drivers is sufficient to rescue CI, and we found that cifAwMel expression under both driver lines recapitulates rescue, but at different strengths. Indeed, rescue is strongest when cifAwMel transgene expression is highest. These data are consistent with reports that cifAwMel is a highly expressed gene in transcriptomes of wMel-infected females [57] and the hypothesis that rescue capability is largely determined by the strength of cifAwMel expression in ovaries [43]. These results combined with those for transgenic expression of CI now establish a robust set of methods for future studies of transgene-induced CI and rescue in the D. melanogaster model.
The central prediction of the Two-by-One model is that transgenic CI can be synthetically rescued in the absence of Wolbachia through dual cifA and cifB expression in uninfected males and cifA expression in uninfected females. Here, we explicitly validate the model that two genes are required in males to cause CI, and one in females is required to rescue it using wMel cif gene variants. However, to confirm that the optimized expression system does not influence the ability of cifAwMel or cifBwMel alone to induce CI, we singly expressed them with the improved driver and found that embryonic hatching does not statistically differ from compatible crosses. Coupled with prior data in wMel [38,43], these results strongly support the Two-by-One genetic model whereby dual cifAwMel and cifBwMel expression is required in the testes to cause a sperm modification that can then be rescued by cifAwMel expression in the ovaries (Fig 6A).
Fig 6. The Two-by-One model of CI.
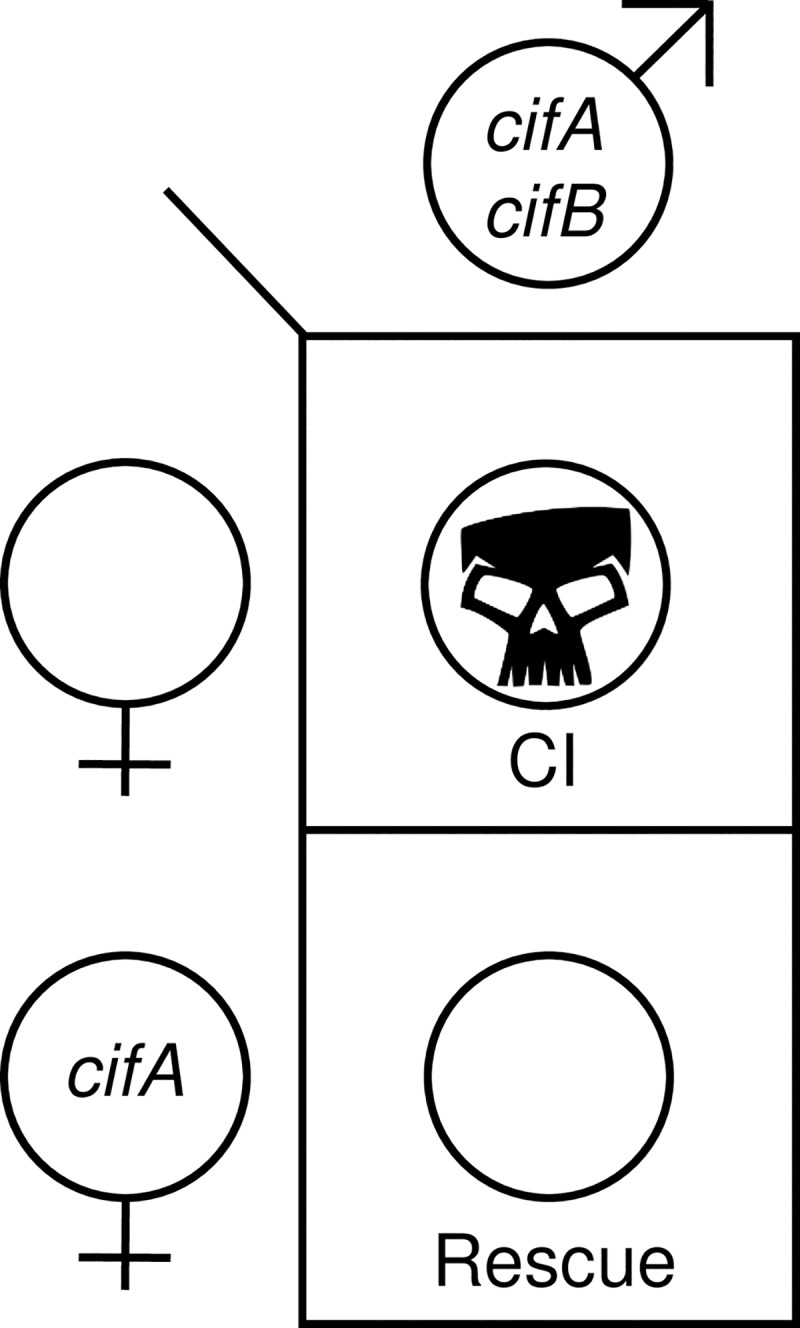
The Two-by-One genetic model of CI explains that cifA and cifB dual expression in uninfected males is necessary for embryonic lethality (CI; skull) when crossed to uninfected and non-expressing females. However, females expressing cifA can rescue CI in their offspring (rescue; open circle). Skull art is from vecteezy.com.
While the genetic basis of unidirectional CI appears resolved, it remains unclear how cifAwMel and cifBwMel functionally operate to generate these phenotypes. Numerous mechanistic models have been proposed over the last two decades [58–64]. We can broadly summarize these models into either host-modification (HM) [59] or toxin-antidote (TA) [58] models. HM models suggest that CI-inducing factors modify host products in such a way that would be lethal unless they are later reversed by rescue factors [59–64]. Conversely, TA models state that the CI-inducing factor is toxic to the developing embryo unless it is crucially bound to a cognate antidote provided by the female [42,58,59]. There are numerous lines of evidence in support of both sets of hypotheses and while the Two-by-One genetic model does not explicitly support or favor one set of models over the other, it can be used to generate hypotheses related to the mechanism of CI.
HM models [59] predict that CI factors directly interact with host products in the testes, modify them, and are displaced. These modifications travel with the sperm, in the absence of Wolbachia and Cif products, and would induce the canonical cytological embryonic defects including delayed paternal nuclear envelope breakdown, slowed Cdk1 activation, a failure of maternal histones to deposit onto the paternal genome, stalled or failed replication of the paternal DNA, a failure of paternal chromosomes to segregate, and later stage regional mitotic failures [7,38,60,61,64–67], or they are reversed by female-derived rescue factors. Leading HM models are the Mistiming [60,61] and Goalkeeper [63] models that leverage findings that male pronuclei are delayed in the first mitosis during embryonic development in CI crosses [61,65,67]. Since the first mitosis is initiated when the female pronucleus has developed, the delay of the male pronuclei leads to cytological defects [60]. It is thus proposed that rescue occurs through resynchronization of the first mitosis by comparably delaying the female pronucleus [60,61]. The Goalkeeper model expands the mistiming model to propose that the strength of the delay is what drives incompatibility between different Wolbachia strains [63]. There are numerous hypotheses to explain the role of the Cif products in these kinds of models. One such hypothesis would be that CifA is responsible for pronuclear delay, thus capable of delaying both the male and female pronuclei, but it requires CifB to properly interact with testis-associated targets. This hypothesis may predict that CifB acts to either protect CifA from ubiquitin tagging and degradation, localize it to a host target, or bind CifA to elicit a conformational change required for interacting with male-specific targets. Alternatively, CI-affected embryos express defective paternal histone deposition, protamine development, delayed nuclear breakdown, and delays in replication machinery [7,60,61,64–67]. Any of these factors could be explained by modifications occurring from HM-type interactions between Cif and host products.
TA models [58] contrast to HM models and require that the CI toxin transfers with or in the sperm and directly binds to a female-derived antidote in the embryo. If the antidote is absent, the CI toxin would induce cytological embryonic defects [7,38,60,61,64–67]. There is mixed evidence in support of this model. First, mass spectometry and SDS-PAGE analyses in Culex pipiens reveal that CifAwPip peptides are present in female spermatheca after mating, suggesting CifAwPip is transferred with or in the sperm [39]. CifBwPip was not detected in these analyses, curiously suggesting that the CifB toxin was not transferred [39]. These results are inconsistent with the TA model, but the lack of transferred CifB may occur because cifB gene expression is up to nine-fold lower than that of cifA [57], and the concentration may have been too low to be observed via these methods. Second, CifA and CifB bind in vitro [42]. However, it remains unclear if CifA-CifB binding enables rescue since this binding has no impact on known enzymatic activities of CifB [42]. While the Two-by-One model does not explicitly support or reject the TA model, it does further inform it. Most intriguing is to understand how CifA acts as a contributor to CI when expressed in testes and as a rescue factor when expressed in ovaries. One hypothesis is that CifA and CifB bind to form a toxin complex that is later directly inhibited by female derived CifA [43,59]. The difference in function between these two environments could be explained by post-translational modification and/or differential localization of CifA in testes and embryos [43,59]. Alternatively, CifB may be the primary toxin, but is incapable of inducing CI unless a CifA antidote is present in both the testes and the ovaries [58]. This hypothesis predicts that male-derived CifA rapidly degrades, leaving CifB with or in the sperm. On its own, CifB would induce lethal cytological embryonic defects [60–62,64] unless provided with a fresh supply of CifA from the embryo.
It has been suggested that divergence in CI and rescue factors causes the incipient evolution of reciprocal incompatibility, or bidirectional CI, between different Wolbachia strains [38,43,68,69]. Here, we review a non-exhaustive set of hypotheses that we previously proposed to explain the emergence of bidirectional CI and are consistent with the Two-by-One model [43]. First, the simplest explanation for CifA’s role in both CI and rescue is that it has similar functional effects in both testes/sperm and ovaries/embryos. Thus, instead of requiring a separate mutation for CI and another for rescue [69], bidirectional CI may emerge from a single CifA mutation that causes incompatibility against the ancestral strain while maintaining self-compatibility. Second, CifA in testes and ovaries may also have different functions, localizations, or posttranslational modifications that contribute to CI and rescue. If this occurs, or if CifB is also an incompatibility factor, the evolution of bidirectional CI may require two or more mutations, and the strain may pass through an intermediate phenotype wherein it becomes unidirectionally incompatible with the ancestral variant or loses the capability to induce either CI or rescue before becoming bidirectionally incompatible with the ancestral variant. In fact, some Wolbachia strains are incapable of inducing CI but capable of rescuing CI induced by other strains [70], and some can induce CI but cannot be rescued [71]. Furthermore, sequence variation in both cifA and cifB from Wolbachia strains in Drosophila [38] and in small regions among strains of wPip Wolbachia [68] have been correlated to incompatibility, suggesting that variation in both genes influence incompatibility.
Additionally, it remains possible that significant divergence in cifA, cifB, or both may be necessary to generate new phenotypes. Indeed, comparative genomic analyses reveal high levels of amino acid divergence in CifA and CifB that correlates with incompatibility between strains [38,40]. Moreover, some Wolbachia strains harbor numerous phage WO variants, each with their own, often divergent, cif genes, and the presence of multiple variants likewise correlates with incompatibility [38,40,68]. Thus, horizontal transfer of phage WO [37,72–76] can in theory rapidly introduce new compatibility relationships, and duplication of phage WO regions, or specifically cif genes, in the same Wolbachia genome may relax the selective pressure on the cif genes and enable their divergence. Determining which of the aforementioned models best explains the evolution of incompatibilities between Wolbachia strains will be assisted by additional sequencing studies to identify incompatible strains with closely related cif variants.
The genetic bases of numerous gene drives have been elucidated in plants [77], fungi [78–81], and nematodes [82,83]. Some gene drives have also been artificially replicated with transgenic constructs [84–86]. However, to our knowledge, the synthetic replication of the Two-by-One model of CI represents the first instance that a gene drive has been constructed by engineering eukaryotic reproduction to depend on phage proteins. Additionally, vector control programs using Wolbachia rely on their ability to suppress pathogens such as Zika and dengue viruses, reduce the size of vector populations, and spread Wolbachia into a host population via CI and rescue. However, there are limitations to these approaches. Most critically, not all pathogens are inhibited by Wolbachia infection and some are enhanced, such as West Nile Virus in Culex tarsalis infected with wAlbB Wolbachia [87]. Additionally, it requires substantial effort to establish a Wolbachia transinfection in a target non-native species [88] that could be obviated in genetically tractable vectors utilizing transgenic gene drives.
The complete synthetic replication of CI and rescue via the Two-by-One model represents a step towards transgenically using the cif genes in vector control efforts. The separation of CI mechanism from Wolbachia infection could theoretically expand CI’s utility to spread ‘payload’ genes that reduce the vectoral capacity of their hosts [89] into a vector population by, for instance, expressing the CI genes and the payload gene polycistronically under the same promoter in the vector’s nuclear or mitochondrial genomes. Moreover, these synthetic constructs have potential to increase the efficiency of Wolbachia-induced CI if they are transformed directly into Wolbachia genomes. For these efforts to be successful, considerable work is necessary to (i) generate a constitutively expressing cif gene drive that does not require GAL4 to operate, (ii) understand the spread dynamics of transgenic CI, (iii) characterize the impact of cif transgenic expression on insect fitness relative to wild vectors, (iv) generate and test effective payload genes in combination with cif drive, (v) explore and optimize the efficacy of cif drive in vector competent hosts such as mosquitoes, (vi) assess the impact of host factors on cif drive across age and development, (vii) compare the efficacy of a cif gene drive to other comparable technologies (CRISPR, homing drive, Medea, etc), in addition to numerous other lines of study. For example, while a substantial body of literature exists to describe the spread dynamics of CI [10,12,13,36,90,91], none yet describe how the Two-by-One model would translate into nuclear or mitochondrial spread dynamics in the absence of Wolbachia. As such, this study represents an early proof of concept that these genes alone are capable of biasing offspring survival in favor of flies expressing these genes under strictly controlled conditions, and should motivate additional study towards its application in vector control.
The generality of the Two-by-One model remains to be tested because it may be specific to certain strains of Wolbachia and/or phage haplotypes. For instance, transgenic expression of cifBwPip from C. pipiens in yeast yields temperature sensitive lethality that can be rescued by dual-expression of cifAwPip and cifBwPip [42]. Moreover, attempts to generate a cifBwPip transgenic line failed, possibly due to generalized toxicity from leaky expression [42]. Therefore, cifBwPip alone could in theory cause CI. However, this model has not been explicitly tested, it has not been explained how cifAwPip and cifBwPip dual-expression induces CI in transgenic Drosophila but prevents CI in yeast, and transgenic wPip CI has not been rescued in an insect. As such, it remains possible that cifBwPip lethality could be explained by artefactual toxicity of overexpression or toxic expression in a heterologous system. Thus, confirmation of an alternative model for CI in wPip is precluded by lack of evidence that cifBwPip alone can induce rescuable lethality in an insect. Since cifBwPip transgenic UAS constructs have not been generated due to toxicity from leaky expression, alternative PhiC31 landing sites or expression systems (i.e., the Q System) could prove valuable in addressing these questions.
Finally, these results further validate the importance of cifAwMel as an essential component of CI and underscore a community need to unify the nomenclature of the CI genes. When the CI genes were first reported, they were described as both CI factors (cif) and as CI deubiquitilases (cid), both of which are actively utilized in the literature. The cif nomenclature was proposed as a cautious naming strategy agnostic to the varied biochemical functions to be discovered, whereas the cid nomenclature was proposed based on the finding that the B protein is in part an in vitro deubiquitilase that, when ablated, inhibits CI-like induction [38,42]. A recent nomenclature proposal suggested that the cif gene family name be used as an umbrella label to describe all CI-associated factors whereas cidA and cidB would be used to describe the specific genes [58]. However, we do not agree with this nomenclature revision despite the appeal of combining the two nomenclatures. CifA protein is not a putative deubiquitilase [40], does not influence deubiquitilase activity of CifB [42], functions independently to rescue CI [43] and, as emphasized by the work in this study, is necessary for CI induction and rescue. The competing nomenclature presumes that it is appropriate to name the A protein cid because it could be expressed in an operon with the B protein. However, the evidence for the operon status of the genes is weak, and more work is needed to describe the regulatory control of these genes before they can be categorized as an operon [59]. Moreover, distant homologs that cluster into distinct phylogenetic groups are proposed to be named CI nucleases (cin) [42] yet the merger of these two groups into one name lacks phylogenetic rationality as the two lineages are as markedly divergent from each other as they are from cid [59]. In addition, none of these distant homologs have been functionally characterized as CI genes [38,40]. As such, it is more appropriate to call these genes “cif-like” to reflect their homology and unknown phenotypes. Thus, the holistic and conservative cif nomenclature with Types (e.g., I-IV) used to delineate phylogenetic clades is appropriately warranted in utilizing and unifying CI gene naming.
In conclusion, the results presented here support that both cifAwMel and cifBwMel phage genes are necessary and sufficient to induce strong CI. In addition, cifAwMel is the only gene necessary for rescue of either transgenic or wild type wMel CI. These results confirm the Two-by-One model of CI in wMel Wolbachia and phage WO with implications for the mechanism of CI and for the diversity of incompatibility between strains, and they provide additional context for understanding CI currently deployed in vector control efforts. The synthetic replication of CI in the absence of Wolbachia marks an early step in developing CI as a tool for genetic and mechanistic studies in D. melanogaster and for vector control efforts that may drive payload genes into vector competent populations.
Materials and methods
Fly rearing and strains
D. melanogaster stocks y1w* (BDSC 1495), nos-GAL4-tubulin (BDSC 4442), nos-GAL4:VP16 (BDSC 4937), otu-GAL4:VP16 (BDSC 58424), and UAS transgenic lines homozygous for cifA, cifB, and cifA;B [38] were maintained at 12:12 light:dark at 25o C and 70% relative humidity (RH) on 50 ml of a standard media. cifA insertion was performed with y1 M{vas-int.Dm}ZH-2A w*; P{CaryP}attP40 and cifB insertion was performed with y1 w67c23; P{CaryP}attP2, as previously described [38]. UAS transgenic lines and nos-GAL4:VP16 were uninfected whereas nos-GAL4-tubulin and otu-GAL4:VP16 lines were infected with wMel Wolbachia. Uninfected versions of infected lines were produced through tetracycline treatment as previously described [38]. WolbF and WolbR3 primers were regularly used to confirm infection status [38]. Stocks for virgin collections were stored at 18o C overnight to slow eclosion rate, and virgin flies were kept at room temperature.
Hatch rate assays
To test for CI, hatch rate assays were used as previously described [38,43]. Briefly, GAL4 adult females were aged 9–11 days post eclosion and mated with UAS males. Age controlled GAL4-UAS males and females were paired in 8 oz bottles affixed with a grape-juice agar plate smeared with yeast affixed to the opening with tape. 0–48 hour old males were used since CI strength rapidly declines with male age [50,52]. The flies and bottles were stored at 25o C for 24 h at which time the plates were replaced with freshly smeared plates and again stored for 24 h. Plates were then removed and the number of embryos on each plate were counted and stored at 25o C. After 30 h the remaining unhatched embryos were counted. The percent of embryos hatched into larvae was calculated by dividing the number of hatched embryos by the initial embryo count and multiplying by 100.
Expression analyses
To assay transgenic RNA expression levels under the various gene drive systems, transgene expressing flies from hatch rates were immediately collected and frozen at -80°C for downstream application as previously described [43]. In brief, abdomens were dissected, RNA was extracted using the Direct-zol RNA MiniPrep Kit (Zymo), the DNA-free kit (Ambion, Life Technologies) was then used to remove DNA contamination, and cDNA was generated with SuperScript VILO (Invitrogen). Quantitative PCR was performed on a Bio-Rad CFX-96 Real-Time System in duplicate using iTaq Universal SYBR Green Supermix (Bio-Rad) using the cifA_opt and rp49 forward and reverse primers as previously described [43]. Samples with a standard deviation >0.3 between duplicates were excluded from analysis. Fold expression of cifA relative to rp49 was determined with 2−ΔΔCt. Each expression study was conducted once.
Statistical analyses
All statistical analyses were conducted in GraphPad Prism (Prism 8). Hatch rate statistical comparisons were made using Kruskal-Wallis followed by a Dunn’s multiple comparison test. A Mann-Whitney-U was used for statistical comparison of RNA fold expression. A linear regression was used to assess correlations between hatch rate and expression. All p-values are reported in S1 Table.
Supporting information
(XLSX)
(A) A linear regression of cifAwMel and cifBwMel expression reveals a positive correlation for both nos-GAL4-tubulin and nos-GAL4VP16. (B,C) A linear regression of (B) cifAwMel and (C) cifBwMel expression and embryonic hatching reveals no correlation for nos-GAL4-tubulin. Removal of data points corresponding to 0% embryonic hatching did not change the significance of the correlation. The nos-GAL4:VP16 driver was not included in analysis A or B since the majority of data points corresponded with 0% hatching. This analysis uses hatch rate samples from the experiment in Fig 2A and expression data from Fig 2B and Fig 2C.
(TIF)
(XLSX)
Acknowledgments
We thank Brittany Leigh, Jennie Shuman, and Mahip Kalra for comments on an earlier version of the manuscript. We also thank three anonymous reviewers and the editor for their excellent feedback, comments, questions, and critiques.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by National Institutes of Health (NIH) awards R01 AI132581 and R21 HD086833 to SRB, National Science Foundation award IOS 1456778 to SRB, the National Science Foundation Graduate Research Fellowship to JDS, and the Vanderbilt Microbiome Initiative. Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors(s) and do not necessarily reflect the views of the National Institutes of Health or the National Science Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Weinert LA, Araujo-Jnr EV, Ahmed MZ, Welch JJ. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc R Soc B. 2015;282: 20150249 10.1098/rspb.2015.0249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zug R, Hammerstein P. Still a Host of Hosts for Wolbachia: Analysis of Recent Data Suggests That 40% of Terrestrial Arthropod Species Are Infected. PLOS ONE. 2012;7: e38544 10.1371/journal.pone.0038544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ferri E, Bain O, Barbuto M, Martin C, Lo N, Uni S, et al. New Insights into the Evolution of Wolbachia Infections in Filarial Nematodes Inferred from a Large Range of Screened Species. PLOS ONE. 2011;6: e20843 10.1371/journal.pone.0020843 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.LePage D, Bordenstein SR. Wolbachia: Can we save lives with a great pandemic? Trends in Parasitology. 2013;29: 385–393. 10.1016/j.pt.2013.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Taylor MJ, Bordenstein SR, Slatko B. Microbe Profile: Wolbachia: a sex selector, a viral protector and a target to treat filarial nematodes. Microbiology. 2018;164: 1345–1347. 10.1099/mic.0.000724 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bordenstein SR, Uy JJ, Werren JH. Host genotype determines cytoplasmic incompatibility type in the haplodiploid genus Nasonia. Genetics. 2003;164: 223–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Serbus LR, Casper-Lindley C, Landmann F, Sullivan W. The Genetics and Cell Biology of Wolbachia-Host Interactions. Annual Review of Genetics. 2008;42: 683–707. 10.1146/annurev.genet.41.110306.130354 [DOI] [PubMed] [Google Scholar]
- 8.Yen JH, Barr AR. The etiological agent of cytoplasmic incompatibility in Culex pipiens. Journal of Invertebrate Pathology. 1973;22: 242–250. 10.1016/0022-2011(73)90141-9 [DOI] [PubMed] [Google Scholar]
- 9.Hancock PA, Sinkins SP, Godfray HCJ. Population Dynamic Models of the Spread of Wolbachia. The American Naturalist. 2011;177: 323–333. 10.1086/658121 [DOI] [PubMed] [Google Scholar]
- 10.Hoffmann A, Turelli M, Harshman L. Factors Affecting The Distribution of Cytoplasmic Incompatibility in Drosophila simulans. Genetics. 1990;126: 933–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Leftwich PT, Edgington MP, Harvey-Samuel T, Carabajal Paladino LZ, Norman VC, Alphey L. Recent advances in threshold-dependent gene drives for mosquitoes. Biochem Soc Trans. 2018;46: 1203–1212. 10.1042/BST20180076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Turelli M. Evolution of incompatibility-inducing microbes and their hosts. Evolution. 1994. October;48(5):1500–13. 10.1111/j.1558-5646.1994.tb02192.x [DOI] [PubMed] [Google Scholar]
- 13.Turelli M, Cooper BS, Richardson KM, Ginsberg PS, Peckenpaugh B, Antelope CX, et al. Rapid Global Spread of wRi-like Wolbachia across Multiple Drosophila. Current Biology. 2018;28: 963–971.e8. 10.1016/j.cub.2018.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bordenstein SR, O’Hara FP, Werren JH. Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature. 2001;409: 707–710. 10.1038/35055543 [DOI] [PubMed] [Google Scholar]
- 15.Brucker RM, Bordenstein SR. Speciation by symbiosis. Trends in Ecology & Evolution. 2012;27: 443–451. 10.1016/j.tree.2012.03.011 [DOI] [PubMed] [Google Scholar]
- 16.Jaenike J, Dyer KA, Cornish C, Minhas MS. Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biol. 2006. October;4(10):e325 10.1371/journal.pbio.0040325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Miller WJ, Ehrman L, Schneider D. Infectious speciation revisited: impact of symbiont-depletion on female fitness and mating behavior of Drosophila paulistorum. PLoS Pathog. 2010;6: e1001214 10.1371/journal.ppat.1001214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shropshire JD, Bordenstein SR. Speciation by Symbiosis: the Microbiome and Behavior. mBio. 2016;7: e01785–15. 10.1128/mBio.01785-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dobson SL, Fox Charles W., Jiggins Francis M. The effect of Wolbachia-induced cytoplasmic incompatibility on host population size in natural and manipulated systems. Proceedings of the Royal Society of London Series B: Biological Sciences. 2002;269: 437–445. 10.1098/rspb.2001.1876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lees RS, Gilles JR, Hendrichs J, Vreysen MJ, Bourtzis K. Back to the future: the sterile insect technique against mosquito disease vectors. Curr Opin Insect Sci. 2015;10: 156–162. 10.1016/j.cois.2015.05.011 [DOI] [PubMed] [Google Scholar]
- 21.Nikolouli K, Colinet H, Renault D, Enriquez T, Mouton L, Gibert P, et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. Journal of Pest Science. 2018;91: 489–503. 10.1007/s10340-017-0944-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.O’Connor L, Plichart C, Sang AC, Brelsfoard CL, Bossin HC, Dobson SL. Open release of male mosquitoes infected with a wolbachia biopesticide: field performance and infection containment. PLoS Negl Trop Dis. 2012;6: e1797 10.1371/journal.pntd.0001797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huang M, Luo J, Hu L, Zheng B, Yu J. Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations. J Theor Biol. 2018. March 7;440:1–11. 10.1016/j.jtbi.2017.12.012 [DOI] [PubMed] [Google Scholar]
- 24.O’Neill SL. The Use of Wolbachia by the World Mosquito Program to Interrupt Transmission of Aedes aegypti Transmitted Viruses. Adv Exp Med Biol. 2018;1062: 355–360. 10.1007/978-981-10-8727-1_24 [DOI] [PubMed] [Google Scholar]
- 25.Caragata EP, Dutra HLC, Moreira LA. Inhibition of Zika virus by Wolbachia in Aedes aegypti. Microb Cell. 2016;3: 293–295. 10.15698/mic2016.07.513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, Muzzi F, et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature. 2011;476: 454–457. 10.1038/nature10356 [DOI] [PubMed] [Google Scholar]
- 27.Bhattacharya T, Newton ILG, Hardy RW. Wolbachia elevates host methyltransferase expression to block an RNA virus early during infection. PLOS Pathogens. 2017;13: e1006427 10.1371/journal.ppat.1006427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brennan LJ, Haukedal JA, Earle JC, Keddie B, Harris HL. Disruption of redox homeostasis leads to oxidative DNA damage in spermatocytes of Wolbachia-infected Drosophila simulans. Insect Mol Biol. 2012;21: 510–520. 10.1111/j.1365-2583.2012.01155.x [DOI] [PubMed] [Google Scholar]
- 29.Caragata EP, Rancès E, Hedges LM, Gofton AW, Johnson KN, O’Neill SL, et al. Dietary cholesterol modulates pathogen blocking by Wolbachia. PLoS Pathog. 2013;9: e1003459 10.1371/journal.ppat.1003459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Geoghegan V, Stainton K, Rainey SM, Ant TH, Dowle AA, Larson T, et al. Perturbed cholesterol and vesicular trafficking associated with dengue blocking in Wolbachia-infected Aedes aegypti cells. Nat Commun. 2017;8: 526 10.1038/s41467-017-00610-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lindsey A, Bhattacharya T, Newton I, Hardy R. Conflict in the Intracellular Lives of Endosymbionts and Viruses: A Mechanistic Look at Wolbachia-Mediated Pathogen-blocking. Viruses. 2018;10: 141 10.3390/v10040141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Molloy JC, Sommer U, Viant MR, Sinkins SP. Wolbachia Modulates Lipid Metabolism in Aedes albopictus Mosquito Cells. Appl Environ Microbiol. 2016;82: 3109–3120. 10.1128/AEM.00275-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schultz MJ, Isern S, Michael SF, Corley RB, Connor JH, Frydman HM. Variable Inhibition of Zika Virus Replication by Different Wolbachia Strains in Mosquito Cell Cultures. J Virol. 2017;91 10.1128/JVI.00339-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hughes GL, Koga R, Xue P, Fukatsu T, Rasgon JL. Wolbachia Infections Are Virulent and Inhibit the Human Malaria Parasite Plasmodium Falciparum in Anopheles Gambiae. PLOS Pathogens. 2011;7: e1002043 10.1371/journal.ppat.1002043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Schmidt TL, Barton NH, Rašić G, Turley AP, Montgomery BL, Iturbe-Ormaetxe I, et al. Local introduction and heterogeneous spatial spread of dengue-suppressing Wolbachia through an urban population of Aedes aegypti. PLOS Biology. 2017;15: e2001894 10.1371/journal.pbio.2001894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Turelli M, Barton NH. Deploying dengue-suppressing Wolbachia: Robust models predict slow but effective spatial spread in Aedes aegypti. Theor Popul Biol. 2017;115: 45–60. 10.1016/j.tpb.2017.03.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bordenstein SR, Bordenstein SR. Eukaryotic association module in phage WO genomes from Wolbachia. Nature Communications. 2016;7: 13155 10.1038/ncomms13155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.LePage DP, Metcalf JA, Bordenstein SR, On J, Perlmutter JI, Shropshire JD, et al. Prophage WO genes recapitulate and enhance Wolbachia-induced cytoplasmic incompatibility. Nature. 2017. March 9;543(7644):243–7. 10.1038/nature21391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Beckmann JF, Fallon AM. Detection of the Wolbachia protein WPIP0282 in mosquito spermathecae: implications for cytoplasmic incompatibility. Insect Biochem Mol Biol. 2013;43: 867–878. 10.1016/j.ibmb.2013.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lindsey A, Rice DW, Bordenstein SR, Brooks AW, Bordenstein SR, Newton ILG. Evolutionary Genetics of Cytoplasmic Incompatibility Genes cifA and cifB in Prophage WO of Wolbachia. Genome Biol Evol. 2018. February 1;10(2):434–51. 10.1093/gbe/evy012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sutton ER, Harris SR, Parkhill J, Sinkins SP. Comparative genome analysis of Wolbachia strain wAu. BMC Genomics. 2014. October 24;15:928 10.1186/1471-2164-15-928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Beckmann JF, Ronau JA, Hochstrasser M. A Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility. Nature Microbiology. 2017;2: 17007 10.1038/nmicrobiol.2017.7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Shropshire JD, On J, Layton EM, Zhou H, Bordenstein SR. One prophage WO gene rescues cytoplasmic incompatibility in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2018. May 8;115(19):4987–91. 10.1073/pnas.1800650115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ni J-Q, Zhou R, Czech B, Liu L-P, Holderbaum L, Yang-Zhou D, et al. A genome-scale shRNA resource for transgenic RNAi in Drosophila. Nature Methods. 2011;8: 405–407. 10.1038/nmeth.1592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Doren MV, Williamson AL, Lehmann R. Regulation of zygotic gene expression in Drosophila primordial germ cells. Current Biology. 1998;8: 243–246. 10.1016/S0960-9822(98)70091-0 [DOI] [PubMed] [Google Scholar]
- 46.Tracey WD, Ning X, Klingler M, Kramer SG, Gergen JP. Quantitative analysis of gene function in the Drosophila embryo. Genetics. 2000;154: 273–284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.He Z, Brinton BT, Greenblatt J, Hassell JA, Ingles CJ. The transactivator proteins VP16 and GAL4 bind replication factor A. Cell. 1993;73: 1223–1232. [DOI] [PubMed] [Google Scholar]
- 48.Sadowski I, Ma J, Triezenberg S, Ptashne M. GAL4-VP16 is an unusually potent transcriptional activator. Nature. 1988;335: 563–564. 10.1038/335563a0 [DOI] [PubMed] [Google Scholar]
- 49.Bourtzis K, Nirgianaki A, Markakis G, Savakis C. Wolbachia infection and cytoplasmic incompatibility in Drosophila species. Genetics. 1996;144: 1063–1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Awrahman ZA, Champion de Crespigny F, Wedell N. The impact of Wolbachia, male age and mating history on cytoplasmic incompatibility and sperm transfer in Drosophila simulans. J Evol Biol. 2014;27: 1–10. 10.1111/jeb.12270 [DOI] [PubMed] [Google Scholar]
- 51.Reynolds KT, Thomson LJ, Hoffmann AA. The effects of host age, host nuclear background and temperature on phenotypic effects of the virulent Wolbachia strain popcorn in Drosophila melanogaster. Genetics. 2003. July;164(3):1027–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Reynolds KT, Hoffmann AA. Male age, host effects and the weak expression or non-expression of cytoplasmic incompatibility in Drosophila strains infected by maternally transmitted Wolbachia. Genet Res. 2002;80: 79–87. [DOI] [PubMed] [Google Scholar]
- 53.Yamada R, Floate KD, Riegler M, O’Neill SL. Male Development Time Influences the Strength of Wolbachia-Induced Cytoplasmic Incompatibility Expression in Drosophila melanogaster. Genetics. 2007;177: 801–808. 10.1534/genetics.106.068486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Poinsot D, Bourtzis K, Markakis G, Savakis C, Mercot H. Wolbachia transfer from Drosophila melanogaster into D. simulans: Host effect and cytoplasmic incompatibility relationships. Genetics. 1998. September;150(1):227–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jaenike J. Spontaneous emergence of a new wolbachia phenotype. Evolution. 2007;61: 2244–2252. 10.1111/j.1558-5646.2007.00180.x [DOI] [PubMed] [Google Scholar]
- 56.Cooper BS, Ginsberg PS, Turelli M, Matute DR. Wolbachia in the Drosophila yakuba Complex: Pervasive Frequency Variation and Weak Cytoplasmic Incompatibility, but No Apparent Effect on Reproductive Isolation. Genetics. 2017. January;205(1):333–51. 10.1534/genetics.116.196238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gutzwiller F, Carmo CR, Miller DE, Rice DW, Newton ILG, Hawley RS, et al. Dynamics of Wolbachia pipientis Gene Expression Across the Drosophila melanogaster Life Cycle. G3 (Bethesda). 2015;5: 2843–2856. 10.1534/g3.115.021931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Beckmann JF, Bonneau M, Chen H, Hochstrasser M, Poinsot D, Merçot H, et al. The Toxin–Antidote Model of Cytoplasmic Incompatibility: Genetics and Evolutionary Implications. Trends in Genetics. 2019; 10.1016/j.tig.2018.12.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Shropshire JD, Leigh B, Bordenstein SR, Duplouy A, Riegler M, Brownlie JC, et al. Models and Nomenclature for Cytoplasmic Incompatibility: Caution over Premature Conclusions–A Response to Beckmann et al. Trends in Genetics. 2019;0 10.1016/j.tig.2019.03.004 [DOI] [PubMed] [Google Scholar]
- 60.Tram U, Sullivan W. Role of delayed nuclear envelope breakdown and mitosis in Wolbachia-induced cytoplasmic incompatibility. Science. 2002. May 10;296(5570):1124–6. 10.1126/science.1070536 [DOI] [PubMed] [Google Scholar]
- 61.Ferree PM, Sullivan W. A genetic test of the role of the maternal pronucleus in Wolbachia-induced cytoplasmic incompatibility in Drosophila melanogaster. Genetics. 2006. June;173(2):839–47. 10.1534/genetics.105.053272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Poinsot D, Charlat S, Mercot H. On the mechanism of Wolbachia-induced cytoplasmic incompatibility: confronting the models with the facts. Bioessays. 2003. March;25(3):259–65. 10.1002/bies.10234 [DOI] [PubMed] [Google Scholar]
- 63.Bossan B, Koehncke A, Hammerstein P. A New Model and Method for Understanding Wolbachia-Induced Cytoplasmic Incompatibility. PLOS ONE. 2011. May 10;6(5):e19757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Landmann F, Orsi GA, Loppin B, Sullivan W. Wolbachia-Mediated Cytoplasmic Incompatibility Is Associated with Impaired Histone Deposition in the Male Pronucleus. PLOS Pathogens. 2009. Mar 20;5(3):e1000343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Callaini G, Dallai R, Riparbelli MG. Wolbachia-induced delay of paternal chromatin condensation does not prevent maternal chromosomes from entering anaphase in incompatible crosses of Drosophila simulans. Journal of Cell Science. 1997. January;110:271–80. [DOI] [PubMed] [Google Scholar]
- 66.Lassy CW, Karr TL. Cytological analysis of fertilization and early embryonic development in incompatible crosses of Drosophila simulans. Mechanisms of Development. 1996. June;57(1):47–58. [DOI] [PubMed] [Google Scholar]
- 67.Tram U, Fredrick K, Werren JH, Sullivan W. Paternal chromosome segregation during the first mitotic division determines Wolbachia-induced cytoplasmic incompatibility phenotype. Journal of Cell Science. 2006. September 1;119(17):3655–63. [DOI] [PubMed] [Google Scholar]
- 68.Bonneau M, Atyame C, Beji M, Justy F, Cohen-Gonsaud M, Sicard M, et al. Culex pipiens crossing type diversity is governed by an amplified and polymorphic operon of Wolbachia. Nature Communications. 2018;9 10.1038/s41467-017-01881-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Charlat S, Calmet C, Merçot H. On the mod resc Model and the Evolution of Wolbachia Compatibility Types. Genetics. 2001;159: 1415–1422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bourtzis K, Dobson SL, Braig HR, O’Neill SL. Rescuing Wolbachia have been overlooked. Nature. 1998;391: 852–853. 10.1038/36017 [DOI] [PubMed] [Google Scholar]
- 71.Zabalou S, Apostolaki A, Pattas S, Veneti Z, Paraskevopoulos C, Livadaras I, et al. Multiple rescue factors within a Wolbachia strain. Genetics. 2008;178: 2145–2160. 10.1534/genetics.107.086488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Chafee ME, Funk DJ, Harrison RG, Bordenstein SR. Lateral phage transfer in obligate intracellular bacteria (Wolbachia): verification from natural populations. Mol Biol Evol. 2010;27: 501–505. 10.1093/molbev/msp275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kent BN, Funkhouser LJ, Setia S, Bordenstein SR. Evolutionary genomics of a temperate bacteriophage in an obligate intracellular bacteria (Wolbachia). PLoS ONE. 2011;6: e24984 10.1371/journal.pone.0024984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang GH, Sun BF, Xiong TL, Wang YK, Murfin KE, Xiao JH, et al. Bacteriophage WO Can Mediate Horizontal Gene Transfer in Endosymbiotic Wolbachia Genomes. Front Microbiol. 2016;7: 1867 10.3389/fmicb.2016.01867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wang GH, Jia L-Y, Xiao J-H, Huang D-W. Discovery of a new Wolbachia supergroup in cave spider species and the lateral transfer of phage WO among distant hosts. Infect Genet Evol. 2016;41: 1–7. 10.1016/j.meegid.2016.03.015 [DOI] [PubMed] [Google Scholar]
- 76.Wang N, Jia S, Xu H, Liu Y, Huang D-W. Multiple Horizontal Transfers of Bacteriophage WO and Host Wolbachia in Fig Wasps in a Closed Community. Front Microbiol. 2016;7: 136 10.3389/fmicb.2016.00136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yang J, Zhao X, Cheng K, Du H, Ouyang Y, Chen J, et al. A killer-protector system regulates both hybrid sterility and segregation distortion in rice. Science. 2012;337: 1336–1340. 10.1126/science.1223702 [DOI] [PubMed] [Google Scholar]
- 78.Grognet P, Lalucque H, Malagnac F, Silar P. Genes that bias Mendelian segregation. PLoS Genet. 2014;10: e1004387 10.1371/journal.pgen.1004387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hammond TM, Rehard DG, Xiao H, Shiu PKT. Molecular dissection of Neurospora Spore killer meiotic drive elements. Proc Natl Acad Sci USA. 2012;109: 12093–12098. 10.1073/pnas.1203267109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hu W, Jiang Z-D, Suo F, Zheng J-X, He W-Z, Du L-L. A large gene family in fission yeast encodes spore killers that subvert Mendel’s law. Elife. 2017;6 10.7554/eLife.26057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Nuckolls NL, Bravo Núñez MA, Eickbush MT, Young JM, Lange JJ, Yu JS, et al. wtf genes are prolific dual poison-antidote meiotic drivers. Elife. 2017;6 10.7554/eLife.26033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ben-David E, Burga A, Kruglyak L. A maternal-effect selfish genetic element in Caenorhabditis elegans. Science. 2017;356: 1051–1055. 10.1126/science.aan0621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Seidel HS, Ailion M, Li J, van Oudenaarden A, Rockman MV, Kruglyak L. A novel sperm-delivered toxin causes late-stage embryo lethality and transmission ratio distortion in C. elegans. PLoS Biol. 2011;9: e1001115 10.1371/journal.pbio.1001115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Akbari OS, Chen C-H, Marshall JM, Huang H, Antoshechkin I, Hay BA. Novel synthetic Medea selfish genetic elements drive population replacement in Drosophila; a theoretical exploration of Medea-dependent population suppression. ACS Synth Biol. 2014;3: 915–928. 10.1021/sb300079h [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Akbari OS, Matzen KD, Marshall JM, Huang H, Ward CM, Hay BA. A synthetic gene drive system for local, reversible modification and suppression of insect populations. Curr Biol. 2013;23: 671–677. 10.1016/j.cub.2013.02.059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Chen C-H, Huang H, Ward CM, Su JT, Schaeffer LV, Guo M, et al. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science. 2007;316: 597–600. 10.1126/science.1138595 [DOI] [PubMed] [Google Scholar]
- 87.Dodson BL, Hughes GL, Paul O, Matacchiero AC, Kramer LD, Rasgon JL. Wolbachia Enhances West Nile Virus (WNV) Infection in the Mosquito Culex tarsalis. PLOS Neglected Tropical Diseases. 2014;8: e2965 10.1371/journal.pntd.0002965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hughes GL, Rasgon JL. Transinfection: a method to investigate Wolbachia–host interactions and control arthropod-borne disease. Insect Molecular Biology. 2014. April 1;23(2):141–51. 10.1111/imb.12066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Champer J, Buchman A, Akbari OS. Cheating evolution: engineering gene drives to manipulate the fate of wild populations. Nature Reviews Genetics. 2016;17: 146–159. 10.1038/nrg.2015.34 [DOI] [PubMed] [Google Scholar]
- 90.Jansen VAA, Turelli M, Godfray HCJ. Stochastic spread of Wolbachia. Proc Biol Sci. 2008. December 7;275(1652):2769–76. 10.1098/rspb.2008.0914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rasgon JL. Using Predictive Models to Optimize Wolbachia-Based Strategies for Vector-Borne Disease Control In: Aksoy S, editor. Transgenesis and the Management of Vector-Borne Disease. New York, NY: Springer New York; 2008. pp. 114–125. 10.1007/978-0-387-78225-6_10 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(A) A linear regression of cifAwMel and cifBwMel expression reveals a positive correlation for both nos-GAL4-tubulin and nos-GAL4VP16. (B,C) A linear regression of (B) cifAwMel and (C) cifBwMel expression and embryonic hatching reveals no correlation for nos-GAL4-tubulin. Removal of data points corresponding to 0% embryonic hatching did not change the significance of the correlation. The nos-GAL4:VP16 driver was not included in analysis A or B since the majority of data points corresponded with 0% hatching. This analysis uses hatch rate samples from the experiment in Fig 2A and expression data from Fig 2B and Fig 2C.
(TIF)
(XLSX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.