

Abstract
Mast cells are granulocytes derived from CD34+ pluripotent progenitor cells that demonstrate plasticity in their development, leaving the bone marrow and differentiating in the tissue where they ultimately reside. They are best known for their role in the allergic response, but also play a prominent immunoregulatory role in other processes, including immune tolerance, the innate immune response, angiogenesis, wound healing and tissue remodeling. Mast cells are found throughout the gastrointestinal tract; their metabolic products influence and regulate intestinal epithelial and endothelial function, gastrointestinal secretion, intestinal motility and absorption, and contribute to host defense. They also play an important role in the development of visceral hypersensitivity through bidirectional interaction with the enteric nervous system. Mast cells have been found to have an increasingly important role in the pathophysiology of a number of pediatric gastrointestinal diseases. This review summarizes the current understanding of the role that mast cells play in the development of pediatric gastrointestinal disorders, including eosinophilic esophagitis, functional dyspepsia, irritable bowel syndrome, celiac disease, inflammatory bowel disease, histologically negative appendicitis, Hirschsprung’s disease, intestinal neuronal dysplasia, and food protein-induced enterocolitis syndrome.
Keywords: Mast cell, eosinophilic esophagitis, inflammatory bowel disease, functional dyspepsia, irritable bowel syndrome, Hirschsprung’s disease, histologically negative appendicitis, food protein induced enterocolitis syndrome, intestinal neuronal dysplasia
Mast cell (MC) physiology
MCs are tissue-resident granulocytes that form an integral part of the allergic response and also play a role in immune tolerance, innate immunity, angiogenesis, and wound healing [1]. MCs are found throughout the body and are common at mucosal surfaces (skin, respiratory tract, and gastrointestinal [GI] tract) that interface with the external environment, often in proximity to blood vessels and nerves, where they form host barriers [2-4]. Their action against pathogens is important to innate immunity; they are important gatekeepers for fighting infectious organisms in portals of entry [5].
MCs do not represent a homogenous population of cells but exhibit significant functional and phenotypic heterogeneity depending on the microenvironment in which they mature [4-6]. This heterogeneity manifests as differences in morphology, biological functions, responsiveness to growth factors, and sensitivity to drugs and secretagogues [5]. Human MCs are classified by their content of serine proteases: tryptase only MC (MCT), chymase only (MCC), or both tryptase and chymase positive MC (MCTC) [5]. Human MCT are located predominately in mucosal tissues, such as the intestine and respiratory tract, whereas human MCTC are mainly found in connective tissues, such as the skin, but may also be found in the submucosa and muscularis propria of the intestinal tract [7].
MCs contain mediators active in both inflammatory and immunological reactions [8]. MCs are capable of reacting within minutes or hours to physical, biological, and chemical stimuli with either local or systemic effects [6]. These cells are activated by cross-linking of cell-surface IgE to antigens, as well as through non-IgE dependent stimulation (complement activation products, microbial components, medications, tissue trauma) [9,10]. Activation results in the release of a wide variety of bioactive mediators into adjacent tissues, where they exert physiological responses directly or indirectly via activation of other effector cells [11].
MCs release a number of mediators upon stimulation that differ in their potency and biological activities. The spectrum of mediators and cytokines produced and released by heterogeneous MCs differs depending on the MC subpopulation and the type of stimulation [6,11]. MCs isolated from human intestine express mRNA for a variety of pro-inflammatory cytokines (tumor necrosis factor [TNF]-α, interleukin [IL]-1β, IL-6, IL-8, IL-16, and IL-18) [12].
Mediators are categorized into 2 groups: preformed and newly synthesized. Preformed mediators are stored in granules and include biogenic amines (histamine), proteoglycans, neutral proteases, and cytokines—a combination of over 50 active factors that help facilitate arteriole vasodilation and vascular permeability [11,13,14] (Table 1). Newly synthesized mediators include arachidonic acid metabolites (prostaglandin D2, leukotriene B4, platelet-activating factor, leukotriene C4) and cytokines; these mediators need RNA transcription and are released within minutes to hours upon MC stimulation [1,11,14,15] (Table 2).
Table 1.
Preformed mast cell mediators [4]
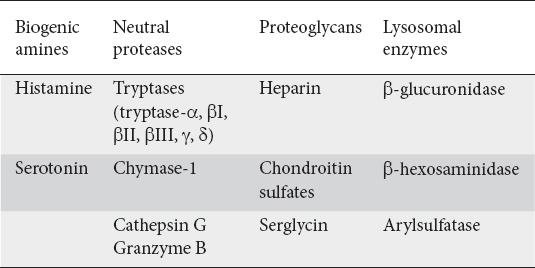
Table 2.

MCs are best known for their prompt secretion of granules (degranulation) that contain large stores of preformed mediators [4]. Preformed mediator release following MC stimulation can result in the release of large quantities of stored histamine within 15-90 sec [4]. This release enables both rapid anaphylactic reactions and allergic responses. It also initiates recruitment of leukocytes to sites of pathogen invasion, activation of innate immune processes and development of inflammatory responses [4]. Long-term responses associated with granule-derived mediators include wound healing and tissue remodeling processes through communication with other cells (fibroblast proliferation and extracellular matrix production) [4].
MCs and the GI tract
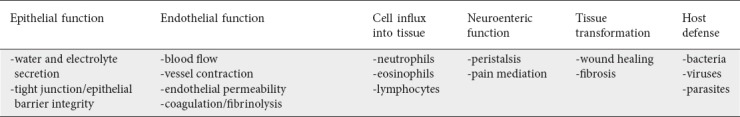
From birth and steadily through childhood there is an incremental rise in MC density that eventually reverses with aging [8]. The number of MCs varies throughout the body, depending on the location and immunologic function of a particular organ [16]. The highest number of progenitor cells was found in the GI tract [16,17]. Such a location may be optimal given the role that MCs play in the host’s defense against bacterial, viral, and parasitic infections. MCs are also thought to regulate a number of physiological functions in the intestine, including epithelial function (water and electrolyte secretion, tight junction/epithelial barrier integrity), endothelial function (blood flow, vessel contraction, endothelial permeability, coagulation/fibrinolysis), cell influx into tissue (neutrophils, eosinophils, lymphocytes), neuroenteric function (intestinal peristalsis, pain mediation), and tissue transformation (wound healing, fibrosis) [17] (Table 3).
Table 3.
Physiologic role of mast cells in the intestine [17]
Most human tissues have mixed populations of MC types. In the human intestinal mucosa >80% of MCs are MCT and 20% MCTC, while the opposite applies to the intestinal submucosa [5]. MCs are present in all layers of the GI tract, but are located predominately in the lamina propria and the submucosal layer [11]. MCs comprise about 2-5% of mononuclear cells in the lamina propria and 1% of all cells found in the intestinal submucosa [16,18]. Typically, MCs do not show clustering (aggregate of 5 or more MCs) and are not present within the surface epithelium [19].
The number and distribution of MCs in the pediatric GI tract is not well defined. An average of 13 cells per high-power field (hpf) is seen in the duodenum and colon [16,18]. The largest number of MCs exists in the descending colon (17±7 MCs/hpf) and the lowest in the rectosigmoid (14±6 MCs/hpf) [19].
MCs are located in proximity to blood vessels and nerve fibers, which makes them an ideal candidate for modulating neural activity and nociception [20]. Approximately 70% of intestinal mucosal MCs are in direct contact with nerve fibers, while another 20% are within 2 μm. Given such proximity, MCs are thought to play a role in visceral sensitivity [16]. MCs can interact with the nervous system bidirectionally; MC mediators such as tryptase and histamine lead to the release of neuropeptides, such as substance P (SP) and calcitonin-gene related peptide, from the proximal nerve endings and SP can subsequently further activate MCs [20]. The motor neuron response, as a result of MC degranulation, can lead to hypersecretion in the GI tract, resulting in diarrhea, abdominal pain, and cramping. Other mediators released by MC degranulation can also sensitize nociceptors in the large intestine. In animal models, this mechanism was shown to reduce the threshold for pain with rectal distension—responses that can both be prevented and controlled with MC stabilizing drugs like cromolyn [16].
MCs are also responsive to both acute and chronic stress [16,17]. Stress can cause increased release of MC mediators (histamine and tryptase) in the proximal intestine in healthy controls and to a larger extent in those with an MC-associated disease (such as food allergy) [17]. There is additional evidence that stress affects intestinal barrier function. This pathway appears to be mediated by the release of corticotropin-releasing hormone, with subsequent activation of mucosal MCs [21].
While beneficial in fighting off pathogens and initiating protective measures, MCs in the GI tract are also implicated in pathologic phenomena under certain circumstances. MCs may transition from a protective, homeostatic role to one that is detrimental and proinflammatory, leading to the development of disease [13]. MC activation can lead to inflammation, which may affect motility, mucosal permeability, intestinal barrier function, and visceral sensitivity [16,17]. By increasing mucosal permeability, MCs can contribute to an ongoing inflammatory process, independently of whether the process is triggered by a harmful microbe or allergen [17]. Hence, MCs have been implicated in the underlying pathophysiology of a number of GI disorders that affect children.
MCs and diagnostic evaluation
The documentation of MC mediator release, at baseline as well as during symptomatic periods, is suggestive of the presence of MC involvement in the pathophysiology of clinical conditions. Pediatric data, however, are limited and the presence of these biomarkers in children must be interpreted with caution. Tryptase is the preferred biomarker for a diagnosis of MC-activation syndrome (MCAS) [22]. Tryptase can be used to reflect MC burden in the body. Consecutive measurements of serum tryptase were found to be helpful when assessing and predicting the clinical course of MCAS. The timing of sample collection and the severity of the clinical event should also be considered when interpreting tryptase results. For example, serum tryptase peaks 15-60 min after an anaphylactic event and declines with a half-life of about 2 h [23]. Other biomarkers, such as 24-h urinary histamine metabolites, 24-h urinary prostaglandin D2 or its metabolite 11β-prostaglandin-F2α, can be helpful, but meaningful elevation cutoffs have not been established [9,24].
Eosinophilic esophagitis (EoE)
EoE is a chronic, immune-mediated disease characterized by eosinophil-predominant inflammation isolated to the esophagus (peak count of ≥15 eosinophils per hpf) [25,26]. EoE is strongly linked to atopy and is often found in association with asthma, allergic rhinitis, eczema, food and aeroallergen hypersensitivity. A non-IgE-mediated mechanism, characterized by a Th2-predominant immune response, plays an important role in the development of disease [26]. Eosinophilic recruitment occurs via mediators like IL-5 and the eotaxin family of chemokines. Eotaxin-3 gene (encoded by CCL26) has been found to be abundantly expressed in the esophagus of EoE patients; expression correlates with disease activity [26,27]. Once activated, eosinophils then modulate various aspects of the immune response and can damage the surrounding tissue [26,27].
Recent studies suggest that MCs may be involved in the development and pathogenesis of EoE; several possible mechanisms have been proposed [7]. MCs produce cytokines (IL-4, IL-5, IL-13, granulocyte-macrophage colony-stimulating factor, eotaxin-1) that activate eosinophils. MCs also promote eosinophil recruitment by synthesizing and releasing eosinophil-selective chemokines and leukotrienes [7]. An increased number of MCs (20-fold increase found in the mucosal layer of the esophagus) has been noted in esophageal biopsies from patients with EoE [7,18]. MC density correlates with the severity of esophageal eosinophilia and the number of MCs decreases after treatment [7]. In addition, these MCs undergo substantial degranulation in EoE compared with healthy controls [18]. Genes associated with MC recruitment are upregulated in EoE and produce inflammatory mediators, implicated both in the migration of inflammatory cells as well as fibrosis generation [28].
MCs may promote not only inflammation but also fibrosis in EoE, by secreting factors such as transforming growth factor-β, a proinflammatory cytokine that induces smooth muscle contractility contributing to esophageal dysfunction, and tryptase, which promotes proliferation and collagen secretion [27,29].
These findings suggest that MCs are actively involved in the development of EoE [7]. The effect of MC mediator production in EoE may enhance inflammation (through recruitment and activation of eosinophils), alter esophageal motility, and promote stricture formation [7].
Functional dyspepsia (FD)
FD, like other functional GI disorders, is best understood through a biopsychosocial model that incorporates the interaction between physiological factors (inflammation, visceral hypersensitivity, motility disturbances), psychological factors (depression, anxiety, somatization), and social factors (interaction with peers and family) [30]. There is evidence to implicate MCs as a key physiological factor in the pathogenesis of pediatric FD. Increased antral MC density is associated with slower gastric emptying and pre-prandial dysrhythmia in children with FD [30,31]. Moderate to extensive degranulation of MCs is seen in the antrum of pediatric FD patients [32]. Antral MC density also correlates with anxiety scores in children with FD [33]. A correlation has also been noted between gastric MC density and symptoms of postprandial fullness and early satiety commonly seen in FD [32].
The above data suggest that there is an interaction between antral inflammation, gastric electromechanical dysfunction, and development of symptoms in pediatric FD [32]. However, the increased small bowel permeability that has been demonstrated in subsets of patients with irritable bowel syndrome (IBS) is not seen in children with FD, even in the presence of elevated MC density [21]. A study by Yeom et al found that headache associated with dyspeptic symptoms in pediatric FD patients is associated with gastric mucosal MC density, suggesting that MCs may functionally link the brain and the gut in pediatric FD [34].
MCs appear to play a key role in the pathogenesis of FD, modulating both gastric motility and gastric visceral hypersensitivity. This may help us to understand why certain antihistamines, such as cyproheptadine, provide symptom relief in patients with FD. Future studies should include the use of mast-cell stabilizing medications in the treatment of FD.
Celiac disease
Celiac disease is characterized by immune-mediated mucosal damage that occurs as a result of both an adaptive and an innate response to the ingestion of dietary gliadin. A recent study conducted by Frossi et al suggests that MCs are central to the inflammatory process present in celiac disease [35]. Intestinal biopsy specimens of patients with celiac disease demonstrate a correlation between MC density and disease severity. Additionally, it was found that gliadin fragments bind to MCs, thereby activating these cells, inducing degranulation and providing a mechanism through which MCs have a direct role in both the onset and progression of disease [35].
While there is little in the current literature regarding the role of MCs in the pathogenesis of celiac disease, an increased awareness of how these cells promote and exacerbate inflammation may provide new treatment modalities in the future.
IBS
IBS is a functional disorder with a multifactorial pathogenesis, including visceral hypersensitivity, increased intestinal permeability, intestinal dysbiosis, food intolerance, psychological stress and brain-gut axis dysfunction [36]. It has been suggested that low grade immune activation and neuroimmune interactions within the colonic mucosa play an important role in the development of sensorimotor dysfunction and symptoms in IBS patients [37]. A number of studies have demonstrated an increased density of MCs in the duodenum, jejunum, terminal ileum and colon of adult IBS patients [37-40]. Additional studies have reported the proximity of MCs to enteric nerves in the colonic mucosa, indicating a key role in the development of visceral hypersensitivity that is central to the pathogenesis of IBS [36,39,41]. An increased number of degranulated MCs seen by electron microscopy in the colon of IBS patients has also been noted [36,39]. MC degranulation increases the excitability of vagal, splanchnic, and mesenteric afferents, contributing to nociceptive processes associated with visceral pain [36].
Additional studies have demonstrated a connection between a patient’s psychological state and an increased colonic mast-cell concentration. IBS patients with high fatigue and depression scores, as assessed by the Fatigue Impact Scale and Beck Depression Inventory, respectively, were specifically found to have a greater number of MCs in intestinal biopsies [42].
These findings have also been recognized in pediatric IBS patients. An increased number of MCs has been noted in the terminal ileum and colon of pediatric patients with IBS [37]. Furthermore, MC density is related to the intensity and frequency of abdominal pain in these patients [37]. Other studies have suggested increased MC activity in pediatric IBS patients. An increased amount of nerve growth factor (NGF), which may play a role in the development of visceral hypersensitivity and is produced by MCs, has been reported in the rectal mucosa of pediatric patients with diarrhea-predominant IBS [43].
These studies demonstrate a greater density and activity of MCs in the small and large intestine of patients with IBS compared with healthy controls. The role of MCs in the development of visceral hypersensitivity is important. IBS is a challenging disorder to treat; better understanding of how MCs are involved the pathogenesis of the disease may lead to more effective long-standing treatment options.
Inflammatory bowel disease (IBD)
IBD is chronic inflammatory disease attributed to a complex interaction between genetic and environmental factors leading to an aberrant immune response to intestinal microbiota [44]. Increased numbers of MCs in inflamed segments of bowel compared with the non-inflamed segments of bowel have been seen in patients with ulcerative colitis and Crohn’s disease [45]. Increased levels of MC mediators such as histamine, prostaglandin D2, and leukotrienes have also been noted in patients with IBD [46]. Studies demonstrate that activated MCs in vitro release TNF-α, shown to play a critical role in the pathogenesis of IBD. Approximately half of the lamina propria cells producing TNF-α were derived from MCs [47].
Despite this current understanding, the exact mechanism by which MC mediators participate in IBD is still unclear. One proposed mechanism suggests that MC dysfunction in regulating epithelial permeability may be important in the development of disease [13]. Tight junctions must be maintained to prevent the permeation of destructive pathogens and allergens into the mucosa, where they can further provoke MC-dependent inflammation. Disturbances in the epithelium can initiate or worsen inflammation in IBD [13]. This has been demonstrated in an animal model, where rats deficient in MCs (or MC mediator chymase) had lower intestinal permeability compared with wild-type mice, further supporting their role in epithelial membrane homeostasis in the intestinal tract [48].
Furthermore, patients with IBD commonly report IBS-like symptoms, such as abdominal pain and diarrhea, during remissive phases. Such findings are relevant in this setting, suggesting that as inflammation resolves, a group of mucosa and submucosal MCs remain active, contributing to GI symptoms even after the resolution of an IBD flare [13]. A greater number of MCs in close proximity to nerve endings was found in patients with ulcerative colitis, probably contributing to IBS-like symptoms during clinical remission of the underlying IBD [49].
While it has already been established that stress can precede IBD flares and induce changes in the epithelial barrier, studies are now supporting mast-cell involvement in stress-induced changes in IBD [50]. Santos et al conducted a study that analyzed MC-deficient and sufficient rats in times of stress. Both groups were subjected to 5 days of water deprivation, where rats were able to see but not drink water for several days. Results demonstrated that the colons of stressed, MC-sufficient rats had increased numbers of MCs, with evidence of MC activation for an entire 3-day period after the initial stressor [50].
MCs are also thought to play a role in the development of strictures and fistulae, 2 irreversible complications of chronic intestinal inflammation in IBD, related to poor wound healing [51]. MCs and their mediators play important roles in inflammation, proliferation, and remodeling of wound healing [51]. An ongoing presence of MCs for up to 20 days after an IBD exacerbation, even when other inflammatory cells were normalized, has been noted [52]. MCs also accumulate at the site of injury, releasing mediators and attracting other immune cells. They are capable of inducing nearby cells to increase their expression of biologically active factors important to the formation of strictures in Crohn’s disease [13].
IBD is due to an aberrant immune response to commensal flora; MCs appear to be involved in this process. Increased MC density has been noted in inflamed areas of intestine in patients with IBD. Activated MCs release TNF-α and other mediators involved in the propagation of intestinal inflammation, increased intestinal permeability, and the development of fistulae and strictures.
Appendicitis
The hallmark of acute appendicitis is neutrophilic predominance and infiltration in the mucosa and wall of the appendix. However, about 15-25% of appendices do not display neutrophilic infiltration and are therefore deemed histologically negative appendicitis [53]. In this scenario, MCs are thought to play a role. Yang et al studied 50 pediatric appendectomy cases and found that 30 of them were histologically negative. This study demonstrated the greatest MC density in the mucosa of both normal and histologically negative appendices. MC density, however, was elevated in all 4 layers of the intestinal tract in histologically negative appendices compared with normal controls, suggesting the importance of MCs in the pathogenesis of pediatric histologically negative appendicitis [53]. MC accumulation was therefore thought to provoke appendicitis-like pain in patients with histologically negative appendices via the release of cytokines, especially in close proximity to enteric nerve fibers [53]. These findings have also been reported in adults with histologically negative appendicitis [54,55].
In addition, MCs may also play a role in the pathophysiology of histologically evident appendicitis. Karakus et al examined the distribution of multiple cell types, including eosinophils and MCs, in children with normal and inflamed appendices. They found a statistically significant increase in the number of MCs in the appendicitis group compared with the control group, which correlated with the degree of inflammation. Such findings suggest that MCs and their metabolites are important in the development of appendicitis in children [56].
MCs appear to play a role in both histologically evident appendicitis as well as histologically negative appendicitis. Histologically negative appendicitis is mediated through MC activation of enteric nerve fibers, resulting in pain, while MC density is increased in histologically evident appendicitis, suggesting a role in the inflammatory process.
Hirschsprung’s disease (HD) and intestinal neuronal dysplasia (IND)
HD is a rare congenital disorder that is due to an absence of ganglia in the submucosal and intramuscular plexus of the GI tract [57-59]. Hypertrophic nerve trunks and increased numbers of adrenergic and cholinergic nerve (AchE-positive) fibers are seen in the aganglionic bowel segment [57,58,60]. Children below 6 months demonstrate hypertrophied nerve trunks and fibers only in the muscular mucosa and submucosa [59]. In children over 6 months there are abundant nerve fibers in all 3 layers of the mucosa [59]. Despite extensive research, the pathogenesis of the disease is not fully understood. There have been reports suggesting that MCs may play a role in the development of HD [58,60-62].
MCs have been observed in direct contact with nerve fibers, suggesting that they are essential for nerve growth and repair [58,61]. They synthesize, store, and release NGF, which supports the development and maintenance of nerve fibers [58,61]. Increased numbers of MCs have been reported in the aganglionic segment of the colon in patients with HD [58,60-62]. A transmural distribution was found in HD patients, with increases of MCs in all layers of the aganglionic segment [58,60,62]. MCs were also found to be in contact with hypertrophic nerve trunks in aganglionic segments in HD, suggesting that MCs may contribute to the development of hypertrophic nerve trunks through the release of NGF [58,60,61]. While the exact role of MCs in HD is not known, these findings suggest that MCs may be involved in the pathogenesis of the disease.
First described by Meier-Ruge in 1971, IND represents a range of neural-crest malformations [60]. Although IND is not clearly defined, suggested morphologic features include hyperplasia of enteric ganglia, giant and ectopic ganglia, and increased activity of acetylcholinergic nerve fibers [58,60]. Large numbers of MCs have been found in all layers of IND segments compared with non-IND segments and controls [58,60]. This suggests that MCs may be responsible for the development of giant ganglia. While depletion of mucin is seen in aganglionic segments in HD patients, this finding is not found in IND, indicating that innervation anomalies in IND do not affect colonic epithelial differentiation as in HD [60].
The pathogenesis of HD and IND has not been fully elucidated. The extensive presence of MCs in the intestinal tissue of both these disease entities suggests that these cells may contribute to the development of disease. MCs are involved in the development and growth of nerve fibers. In this context, aberrant function of MCs may lead to disease.
Food protein-induced proctocolitis (FPIES)
FPIES is a severe non-IgE-mediated food allergy that affects infants and young children [63]. Recent data suggest involvement of MCs in addition to both T cells and neutrophils. Markedly increased MCs (50 per hpf) have been found in the rectum during the acute phase of FPIES [64]. Elevated baseline serum tryptase levels have been reported in pediatric patients with active FPIES, suggesting that MC activation is part of the disease [65].
MCAS
MCAS comprise a heterogeneous group of conditions associated with recurrent signs and symptoms of MC mediator release, most notably histamine [1,23,66]. Symptoms depend on the affected organ and include skin (flushing, pruritus, urticaria, angioedema, dermatographism), GI (gastroesophageal reflux disorder, nausea, vomiting, diarrhea, and abdominal cramping), cardiovascular (hypotension, syncope, near syncope, tachycardia), respiratory (wheezing, throat swelling), naso-ocular (nasal congestion, conjunctival injection, nasal pruritus), anaphylaxis, and neurologic (headaches, poor concentration, memory issues, brain fog) manifestations [67,68].
MCAS can be classified into 3 distinct categories: primary (clonal), secondary (reactive), and idiopathic [1,23,66,67]. Primary MCAS are associated with KIT D816V or other KIT exon 17 mutations and are characterized by abnormal qualitative or quantitative production of MCs [9,69]. Primary MCAS includes mastocytosis and monoclonal MC activation syndrome; diagnosis is based on objective pathologic findings, as stated in the World Health Organization (WHO) diagnostic criteria [23,69]. The WHO classification divides mastocytosis into 7 categories, each with distinct clinicopathologic and prognostic features [9,69,70]. Cutaneous mastocytosis (CM) is the most common form in children, accounting for over 90% of pediatric mastocytosis cases [9,69-71]. Pediatric CM is limited to the skin, with no bone marrow or other organ involvement. Systemic mastocytosis (SM), where multiple organ systems are affected (with bone marrow involvement), is rare in children [69]. Virtually all patients with pediatric SM have skin involvement. Organomegaly in conjunction with evidence of MC mediator release is highly suggestive of SM in pediatric patients [72].
Secondary MCAS are more prevalent than primary and idiopathic MCAS and occur in IgE-dependent allergies, chronic inflammatory diseases, neoplastic disorders, physical urticarias, and chronic autoimmune urticaria [23,67]. MCs are produced normally and are generally present in normal numbers in tissues or can be increased in response to an underlying inflammatory process (reactive MC hyperplasia) [69].
Idiopathic MCAS has no identifiable cause; hypotheses concerning the underlying etiology include an intrinsic MC defect, an unidentified endogenous or environmental stimulus, or both, resulting in a hyperactive MC phenotype [23,73].
No consensus definition for idiopathic MCAS has been established, but proposed diagnostic criteria include: 1) episodic multisystem symptoms consistent with MC mediator release; 2) increased levels of MC mediators above baseline during symptomatic periods; 3) clinical improvement with H1 and H2-receptor antagonists, anti-leukotriene medications, or MC stabilizing agents; and 4) no evidence of a primary or secondary disorder of MC activation [24,67,69]. While this condition has predominately been described in adults, pediatric manifestations of idiopathic MCAS are not well recognized.
Pediatric MCAS includes primary, secondary, and idiopathic forms. Primary (clonal) MCAS is extremely rare in children and if present typically takes the form of CM, imited to the skin. Secondary MCAS is more prevalent in children and occurs secondarily to a variety of allergic and inflammatory conditions. Idiopathic MCAS is poorly understood and has not been clearly defined in the pediatric population.
Concluding remarks
MCs are active in both inflammatory and immunologic reactions, through the release of preformed or newly synthesized mediators in response to various stimuli. These cells have an important role in a multitude of physiologic processes. It is clear that MCs play an active role in the pathogenesis of a variety of pediatric GI disorders. A better understanding of how these cells contribute to pediatric GI disease will allow development of new therapeutic modalities in the future.
Biography
Rush University Children’s Hospital, Chicago, IL, USA
Footnotes
Conflict of Interest: None
References
- 1.Cookson H, Grattan C. An update on mast cell disorders. Clin Med (Lond) 2016;16:580–583. doi: 10.7861/clinmedicine.16-6-580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dawicki W, Marshall JS. New and emerging roles for mast cells in host defence. Curr Opin Immunol. 2007;19:31–38. doi: 10.1016/j.coi.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 3.Heib V, Becker M, Taube C, Stassen M. Advances in the understanding of mast cell function. Br J Haematol. 2008;142:683–694. doi: 10.1111/j.1365-2141.2008.07244.x. [DOI] [PubMed] [Google Scholar]
- 4.Moon TC, Befus AD, Kulka M. Mast cell mediators:their differential release and the secretory pathways involved. Front Immunol. 2014;5:569. doi: 10.3389/fimmu.2014.00569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vliagoftis H, Befus AD. Rapidly changing perspectives about mast cells at mucosal surfaces. Immunol Rev. 2005;206:190–203. doi: 10.1111/j.0105-2896.2005.00279.x. [DOI] [PubMed] [Google Scholar]
- 6.Mekori YA, Metcalfe DD. Mast cells in innate immunity. Immunol Rev. 2000;173:131–140. doi: 10.1034/j.1600-065x.2000.917305.x. [DOI] [PubMed] [Google Scholar]
- 7.Wershil BK. Exploring the role of mast cells in eosinophilic esophagitis. Immunol Allergy Clin North Am. 2009;29:189–195. doi: 10.1016/j.iac.2008.09.006. [DOI] [PubMed] [Google Scholar]
- 8.Dollberg L, Gurevitz M, Freier S. Gastrointestinal mast cells in health, and in coeliac disease and other conditions. Arch Dis Child. 1980;55:702–705. doi: 10.1136/adc.55.9.702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Akin C. Mast cell activation disorders. J Allergy Clin Immunol Pract. 2014;2:252–257.e1. doi: 10.1016/j.jaip.2014.03.007. [DOI] [PubMed] [Google Scholar]
- 10.Sattar HA. Fundamentals of pathology:medical course and step 1 review. Chicago, IL: Pathoma.com; 2017. [Google Scholar]
- 11.Yu LC, Perdue MH. Role of mast cells in intestinal mucosal function:studies in models of hypersensitivity and stress. Immunol Rev. 2001;179:61–73. doi: 10.1034/j.1600-065x.2001.790107.x. [DOI] [PubMed] [Google Scholar]
- 12.Lorentz A, Schwengberg S, Sellge G, Manns MP, Bischoff SC. Human intestinal mast cells are capable of producing different cytokine profiles:role of IgE receptor cross-linking and IL-4. J Immunol. 2000;164:43–48. doi: 10.4049/jimmunol.164.1.43. [DOI] [PubMed] [Google Scholar]
- 13.Hamilton MJ, Frei SM, Stevens RL. The multifaceted mast cell in inflammatory bowel disease. Inflamm Bowel Dis. 2014;20:2364–2378. doi: 10.1097/MIB.0000000000000142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bischoff SC. Physiological and pathophysiological functions of intestinal mast cells. Semin Immunopathol. 2009;31:185–205. doi: 10.1007/s00281-009-0165-4. [DOI] [PubMed] [Google Scholar]
- 15.Boyce JA. Mast cells:beyond IgE. J Allergy Clin Immunol. 2003;111:24–32. doi: 10.1067/mai.2003.60. [DOI] [PubMed] [Google Scholar]
- 16.Ramsay DB, Stephen S, Borum M, et al. Mast cells in gastrointestinal disease. Gastroenterol Hepatol (N Y) 2010;6:772–777. [PMC free article] [PubMed] [Google Scholar]
- 17.Bischoff SC, Krämer S. Human mast cells, bacteria, and intestinal immunity. Immunol Rev. 2007;217:329–337. doi: 10.1111/j.1600-065X.2007.00523.x. [DOI] [PubMed] [Google Scholar]
- 18.Abonia JP, Blanchard C, Butz BB, et al. Involvement of mast cells in eosinophilic esophagitis. J Allergy Clin Immunol. 2010;126:140–149. doi: 10.1016/j.jaci.2010.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Saad AG. Normal quantity and distribution of mast cells and eosinophils in the pediatric colon. Pediatr Dev Pathol. 2011;14:294–300. doi: 10.2350/10-07-0878-OA.1. [DOI] [PubMed] [Google Scholar]
- 20.Aich A, Afrin LB, Gupta K. Mast cell-mediated mechanisms of nociception. Int J Mol Sci. 2015;16:29069–29092. doi: 10.3390/ijms161226151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Neilan NA, Garg UC, Schurman JV, Friesen CA. Intestinal permeability in children/adolescents with functional dyspepsia. BMC Res Notes. 2014;7:275. doi: 10.1186/1756-0500-7-275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Frieri M, Quershi M. Pediatric mastocytosis:a review of the literature. Pediatr Allergy Immunol Pulmonol. 2013;26:175–180. doi: 10.1089/ped.2013.0275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Valent P. Mast cell activation syndromes:definition and classification. Allergy. 2013;68:417–424. doi: 10.1111/all.12126. [DOI] [PubMed] [Google Scholar]
- 24.Valent P, Akin C, Arock M, et al. Definitions, criteria and global classification of mast cell disorders with special reference to mast cell activation syndromes:a consensus proposal. Int Arch Allergy Immunol. 2012;157:215–225. doi: 10.1159/000328760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.O'Shea KM, Aceves SS, Dellon ES, et al. Pathophysiology of eosinophilic esophagitis. Gastroenterology. 2018;154:333–345. doi: 10.1053/j.gastro.2017.06.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wechsler JB, Bryce PJ. Allergic mechanisms in eosinophilic esophagitis. Gastroenterol Clin North Am. 2014;43:281–296. doi: 10.1016/j.gtc.2014.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rothenberg ME. Molecular, genetic, and cellular bases for treating eosinophilic esophagitis. Gastroenterology. 2015;148:1143–1157. doi: 10.1053/j.gastro.2015.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lomazi EA, Brandalise NA, Servidoni MFPC, Cardoso SR, Meirelles LR. Mast cells distinguish eosinophilic esophagitis in pediatric patients. Arq Gastroenterol. 2017;54:192–196. doi: 10.1590/S0004-2803.201700000-23. [DOI] [PubMed] [Google Scholar]
- 29.Cianferoni A, Spergel JM, Muir A. Recent advances in the pathological understanding of eosinophilic esophagitis. Expert Rev Gastroenterol Hepatol. 2015;9:1501–1510. doi: 10.1586/17474124.2015.1094372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Friesen CA, Schurman JV, Colombo JM, Abdel-Rahman SM. Eosinophils and mast cells as therapeutic targets in pediatric functional dyspepsia. World J Gastrointest Pharmacol Ther. 2013;4:86–96. doi: 10.4292/wjgpt.v4.i4.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schäppi MG, Borrelli O, Knafelz D, et al. Mast cell-nerve interactions in children with functional dyspepsia. J Pediatr Gastroenterol Nutr. 2008;47:472–480. doi: 10.1097/MPG.0b013e318186008e. [DOI] [PubMed] [Google Scholar]
- 32.Friesen CA, Lin Z, Singh M, et al. Antral inflammatory cells, gastric emptying, and electrogastrography in pediatric functional dyspepsia. Dig Dis Sci. 2008;53:2634–2640. doi: 10.1007/s10620-008-0207-0. [DOI] [PubMed] [Google Scholar]
- 33.Friesen CA, Schurman JV, Qadeer A, Andre L, Welchert E, Cocjin J. Relationship between mucosal eosinophils and anxiety in pediatric dyspepsia. Gastroenterology. 2005;129:A–158. [Google Scholar]
- 34.Yeom JS, Choi MB, Seo JH, et al. Relationship between headache and mucosal mast cells in pediatric Helicobacter pylori-negative functional dyspepsia. Cephalalgia. 2013;33:323–329. doi: 10.1177/0333102412472070. [DOI] [PubMed] [Google Scholar]
- 35.Frossi B, Tripodo C, Guarnotta C, et al. Mast cells are associated with the onset and progression of celiac disease. J Allergy Clin Immunol. 2017;139:1266–1274. doi: 10.1016/j.jaci.2016.08.011. [DOI] [PubMed] [Google Scholar]
- 36.Zhang L, Song J, Hou X. Mast cells and irritable bowel syndrome:from the bench to the bedside. J Neurogastroenterol Motil. 2016;22:181–192. doi: 10.5056/jnm15137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Di Nardo G, Barbara G, Cucchiara S, et al. Neuroimmune interactions at different intestinal sites are related to abdominal pain symptoms in children with IBS. Neurogastroenterol Motil. 2014;26:196–204. doi: 10.1111/nmo.12250. [DOI] [PubMed] [Google Scholar]
- 38.Jakate S, Demeo M, John R, Tobin M, Keshavarzian A. Mastocytic enterocolitis:increased mucosal mast cells in chronic intractable diarrhea. Arch Pathol Lab Med. 2006;130:362–367. doi: 10.5858/2006-130-362-MEIMMC. [DOI] [PubMed] [Google Scholar]
- 39.Lee KN, Lee OY. The role of mast cells in irritable bowel syndrome. Gastroenterol Res Pract. 2016;2016:2031480. doi: 10.1155/2016/2031480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sohn W, Lee OY, Lee SP, et al. Mast cell number, substance P and vasoactive intestinal peptide in irritable bowel syndrome with diarrhea. Scand J Gastroenterol. 2014;49:43–51. doi: 10.3109/00365521.2013.857712. [DOI] [PubMed] [Google Scholar]
- 41.Cheng P, Yao J, Wang C, Zhang L, Kong W. Molecular and cellular mechanisms of tight junction dysfunction in the irritable bowel syndrome. Mol Med Rep. 2015;12:3257–3264. doi: 10.3892/mmr.2015.3808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Piche T, Saint-Paul MC, Dainese R, et al. Mast cells and cellularity of the colonic mucosa correlated with fatigue and depression in irritable bowel syndrome. Gut. 2008;57:468–473. doi: 10.1136/gut.2007.127068. [DOI] [PubMed] [Google Scholar]
- 43.Willot S, Gauthier C, Patey N, Faure C. Nerve growth factor content is increased in the rectal mucosa of children with diarrhea-predominant irritable bowel syndrome. Neurogastroenterol Motil. 2012;24:734–739.e347. doi: 10.1111/j.1365-2982.2012.01933.x. [DOI] [PubMed] [Google Scholar]
- 44.Podolsky DK. Inflammatory bowel disease. N Engl J Med. 2002;347:417–429. doi: 10.1056/NEJMra020831. [DOI] [PubMed] [Google Scholar]
- 45.Nishida Y, Murase K, Isomoto H, et al. Different distribution of mast cells and macrophages in colonic mucosa of patients with collagenous colitis and inflammatory bowel disease. Hepatogastroenterology. 2002;49:678–682. [PubMed] [Google Scholar]
- 46.Fox CC, Lazenby AJ, Moore WC, Yardley JH, Bayless TM, Lichtenstein LM. Enhancement of human intestinal mast cell mediator release in active ulcerative colitis. Gastroenterology. 1990;99:119–124. doi: 10.1016/0016-5085(90)91238-2. [DOI] [PubMed] [Google Scholar]
- 47.Bischoff SC, Lorentz A, Schwengberg S, Weier G, Raab R, Manns MP. Mast cells are an important cellular source of tumour necrosis factor alpha in human intestinal tissue. Gut. 1999;44:643–652. doi: 10.1136/gut.44.5.643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Groschwitz KR, Ahrens R, Osterfeld H, et al. Mast cells regulate homeostatic intestinal epithelial migration and barrier function by a chymase/Mcpt4-dependent mechanism. Proc Natl Acad Sci U S A. 2009;106:22381–22386. doi: 10.1073/pnas.0906372106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van Hoboken EA, Thijssen AY, Verhaaren R, et al. Symptoms in patients with ulcerative colitis in remission are associated with visceral hypersensitivity and mast cell activity. Scand J Gastroenterol. 2011;46:981–987. doi: 10.3109/00365521.2011.579156. [DOI] [PubMed] [Google Scholar]
- 50.Santos J, Yang PC, Söderholm JD, Benjamin M, Perdue MH. Role of mast cells in chronic stress induced colonic epithelial barrier dysfunction in the rat. Gut. 2001;48:630–636. doi: 10.1136/gut.48.5.630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Noli C, Miolo A. The mast cell in wound healing. Vet Dermatol. 2001;12:303–313. doi: 10.1046/j.0959-4493.2001.00272.x. [DOI] [PubMed] [Google Scholar]
- 52.Iba Y, Shibata A, Kato M, Masukawa T. Possible involvement of mast cells in collagen remodeling in the late phase of cutaneous wound healing in mice. Int Immunopharmacol. 2004;4:1873–1880. doi: 10.1016/j.intimp.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 53.Yang Z, Esebua M, Layfield L. The role of mast cells in histologically “normal”appendices following emergency appendectomy in pediatric patients. Ann Diagn Pathol. 2016;24:1–3. doi: 10.1016/j.anndiagpath.2016.06.003. [DOI] [PubMed] [Google Scholar]
- 54.Amber S, Mathai AM, Naik R, Pai MR, Kumar S, Prasad K. Neuronal hypertrophy and mast cells in histologically negative, clinically diagnosed acute appendicitis:a quantitative immunophenotypical analysis. Indian J Gastroenterol. 2010;29:69–73. doi: 10.1007/s12664-010-0016-1. [DOI] [PubMed] [Google Scholar]
- 55.Singh UR, Malhotra A, Bhatia A. Eosinophils, mast cells, nerves and ganglion cells in appendicitis. Indian J Surg. 2008;70:231–234. doi: 10.1007/s12262-008-0066-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Karakuş E, Azili MN, Karabulut B, et al. Is there a role of interstitial cells of cajal and mast cells and eosinophils in appendicitis in children? Turk J Med Sci. 2015;45:800–803. doi: 10.3906/sag-1408-21. [DOI] [PubMed] [Google Scholar]
- 57.Kapur RP. Practical pathology and genetics of Hirschsprung's disease. Semin Pediatr Surg. 2009;18:212–223. doi: 10.1053/j.sempedsurg.2009.07.003. [DOI] [PubMed] [Google Scholar]
- 58.Kobayashi H, Yamataka A, Fujimoto T, Lane GJ, Miyano T. Mast cells and gut nerve development:implications for Hirschsprung's disease and intestinal neuronal dysplasia. J Pediatr Surg. 1999;34:543–548. doi: 10.1016/s0022-3468(99)90069-6. [DOI] [PubMed] [Google Scholar]
- 59.Szylberg L, Marszałek A. Diagnosis of Hirschsprung's disease with particular emphasis on histopathology. A systematic review of current literature. Prz Gastroenterol. 2014;9:264–269. doi: 10.5114/pg.2014.46160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Demirbilek S, Ozardali HI, Aydm G. Mast-cells distribution and colonic mucin composition in Hirschsprung's disease and intestinal neuronal dysplasia. Pediatr Surg Int. 2001;17:136–139. doi: 10.1007/s003830000467. [DOI] [PubMed] [Google Scholar]
- 61.Hermanowicz A, Debek W, Dzienis-Koronkiewicz E, Chyczewski L. Topography and morphometry of intestinal mast cells in children with Hirschsprung's disease. Folia Histochem Cytobiol. 2008;46:65–68. doi: 10.2478/v10042-008-0008-5. [DOI] [PubMed] [Google Scholar]
- 62.Yadav AK, Mishra K, Mohta A, Agarwal S. Hirschsprung's disease:is there a relationship between mast cells and nerve fibers? World J Gastroenterol. 2009;15:1493–1498. doi: 10.3748/wjg.15.1493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Michelet M, Schluckebier D, Petit LM, Caubet JC. Food protein-induced enterocolitis syndrome - a review of the literature with focus on clinical management. J Asthma Allergy. 2017;10:197–207. doi: 10.2147/JAA.S100379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ishige T, Yagi H, Tatsuki M, et al. Endoscopic findings in the acute phase of food protein-induced enterocolitis syndrome. Pediatr Allergy Immunol. 2015;26:90–91. doi: 10.1111/pai.12327. [DOI] [PubMed] [Google Scholar]
- 65.Caubet JC, Bencharitiwong R, Ross A, Sampson HA, Berin MC, Nowak-Węgrzyn A. Humoral and cellular responses to casein in patients with food protein-induced enterocolitis to cow's milk. J Allergy Clin Immunol. 2017;139:572–583. doi: 10.1016/j.jaci.2016.02.047. [DOI] [PubMed] [Google Scholar]
- 66.Lee MJ, Akin C. Mast cell activation syndromes. Ann Allergy Asthma Immunol. 2013;111:5–8. doi: 10.1016/j.anai.2013.02.008. [DOI] [PubMed] [Google Scholar]
- 67.Akin C, Valent P, Metcalfe DD. Mast cell activation syndrome:Proposed diagnostic criteria. J Allergy Clin Immunol. 2010;126:1099–1104.e4. doi: 10.1016/j.jaci.2010.08.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Picard M, Giavina-Bianchi P, Mezzano V, Castells M. Expanding spectrum of mast cell activation disorders:monoclonal and idiopathic mast cell activation syndromes. Clin Ther. 2013;35:548–562. doi: 10.1016/j.clinthera.2013.04.001. [DOI] [PubMed] [Google Scholar]
- 69.Akin C. Mast cell activation syndromes. J Allergy Clin Immunol. 2017;140:349–355. doi: 10.1016/j.jaci.2017.06.007. [DOI] [PubMed] [Google Scholar]
- 70.Akin C, Valent P. Diagnostic criteria and classification of mastocytosis in 2014. Immunol Allergy Clin North Am. 2014;34:207–218. doi: 10.1016/j.iac.2014.02.003. [DOI] [PubMed] [Google Scholar]
- 71.Méni C, Bruneau J, Georgin-Lavialle S, et al. Paediatric mastocytosis:a systematic review of 1747 cases. Br J Dermatol. 2015;172:642–651. doi: 10.1111/bjd.13567. [DOI] [PubMed] [Google Scholar]
- 72.Carter MC, Clayton ST, Komarow HD, et al. Assessment of clinical findings, tryptase levels, and bone marrow histopathology in the management of pediatric mastocytosis. J Allergy Clin Immunol. 2015;136:1673–1679. doi: 10.1016/j.jaci.2015.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Frieri M. Mast cell activation syndrome. Clin Rev Allergy Immunol. 2018;54:353–365. doi: 10.1007/s12016-015-8487-6. [DOI] [PubMed] [Google Scholar]