

Abstract
Biological invasions are increasing globally in number and extent despite efforts to restrict their spread. Knowledge of incursion pathways is necessary to prevent new invasions and to design effective biosecurity protocols at source and recipient locations. This study uses genome‐wide single nucleotide polymorphisms (SNPs) to determine the origin of 115 incursive Aedes aegypti(yellow fever mosquito) detected at international ports in Australia and New Zealand. We also genotyped mosquitoes at three point mutations in the voltage‐sensitive sodium channel (Vssc) gene: V1016G, F1534C and S989P. These mutations confer knockdown resistance to synthetic pyrethroid insecticides, widely used for controlling invertebrate pests. We first delineated reference populations using Ae. aegypti sampled from 15 locations in Asia, South America, Australia and the Pacific Islands. Incursives were assigned to these populations using discriminant analysis of principal components (DAPC) and an assignment test with a support vector machine predictive model. Bali, Indonesia, was the most common origin of Ae. aegypti detected in Australia, while Ae. aegypti detected in New Zealand originated from Pacific Islands such as Fiji. Most incursives had the same allelic genotype across the three Vsscgene point mutations, which confers strong resistance to synthetic pyrethroids, the only insecticide class used in current, widely implemented aircraft disinsection protocols endorsed by the World Health Organization (WHO). Additionally, all internationally assigned Ae. aegypti had Vssc point mutations linked to pyrethroid resistance that are not found in Australian populations. These findings demonstrate that protocols for preventing introductions of invertebrates must consider insecticide resistance, and highlight the usefulness of genomic data sets for managing global biosecurity objectives.
Keywords: Aedes aegypti, assignment tests, biological invasions, biosecurity, discriminant analysis of principal components, genome‐wide SNPs, insecticide resistance, invasion pathways
1. INTRODUCTION
Invasive alien species are a major threat to global biodiversity, agriculture and human health and confer a great economic burden upon developed and developing countries alike (IUCN, 2000). Invasive species have been shown to disrupt soil processes (Purnima, Raghubanshi, & Singh, 2008) and ecological community structure (Hejda, Pyšek, & Jarošík, 2009) and are linked to reductions in the abundance and richness of native species (Blackburn, Cassey, Duncan, Evans, & Gaston, 2004). They are responsible for the global spread of human diseases such as malaria, dengue, Zika and chikungunya that together cause over 700,000 deaths annually and create heavy burdens on health systems worldwide (Lounibos, 2002). The spread of invasive species is closely tied to human economic activity, with many species transported as “stowaways” on human trade and transport vessels (Levine & D'Antonio, 2003; Westphal, Browne, MacKinnon, & Noble, 2008). Following the expansion of trade and transport activity in recent decades, global distributions of invasive species have increased markedly (Hulme, 2009), with current predictions still arguably underestimating their true extent (McGeoch et al., 2010).
Preventing the establishment of new alien species requires knowledge of incursion pathways (IUCN, 2000). Although border interceptions can be effective at stopping incursions (Bacon, Bacher, & Aebi, 2012; Caley, Ingram, & De Barro, 2015), resource limitations restrict the number of inspections on arriving goods and conveyances or the number of border sections that can be monitored. These strategies must therefore seek to maximize detections by incorporating knowledge of incursion pathways, such as more frequent inspection of goods and conveyances from likely source locations (Robinson, Burgman, & Cannon, 2011), or regular inspection of known entry points of highly destructive species (Mehta, Haight, Homans, Polasky, & Venette, 2007; Myers, Simberloff, Kuris, & Carey, 2000). Incursion pathways can also be monitored to estimate parameters indicating the likelihood of future invasions, such as propagule pressure and opportunities for selection (Wilson, Dormontt, Prentis, Lowe, & Richardson, 2009).
Genomic information can be a useful means for identifying source populations of incursions. Invading individuals of unknown source intercepted at borders (hereafter defined as “incursives”) can be genotyped using genetic markers and compared with reference populations from the species' known distribution. Analyses can then be performed to assign incursives to reference populations, in order to identify the likely source population or geographic region of origin. Genome‐wide single nucleotide polymorphisms (SNPs) in particular can provide clear geographical delineation of populations (Rašić, Filipović, Weeks, & Hoffmann, 2014), which allows for confident assignment of intercepted individuals to their population or region of origin. This method can in theory enable the assignment of all incursive individuals, and thus improves upon previous methods either that rely on the interception of incursives at the border as they arrive from a known location (Mehta et al., 2007), or that use lower‐resolution genetic markers that frequently cannot resolve individual origin to a fine geographic scale (Puckett & Eggert, 2016). Genome‐wide SNPs could also be used to identify the source of incursions postborder and improve the chance of eradication by using pathway management to prevent future incursions.
Aedes aegypti (L.) (yellow fever mosquito) is a highly invasive pest and a primary global vector of arboviruses such as dengue (Smith, 1956), Zika (Bogoch et al., 2016) and chikungunya (Weaver & Lecuit, 2015). Its pantropical distribution has grown rapidly over the last century (Kraemer et al., 2015), placing a greater part of the human population at risk of disease. In Australia, Ae. aegypti is currently only endemic to Queensland, but once had a broader distribution that extended into New South Wales, South Australia and Western Australia (Russell et al., 2009). Under current climate change scenarios, there is speculation that the species will increase its distribution in Australia again (Kearney, Porter, Williams, Ritchie, & Hoffmann, 2009; Russell et al., 2009). Furthermore, detections of Ae. aegypti at Australia's international airports increased markedly over the period from 2014 to 2016 (Figure 1), with 39 detections at Perth International Airport alone between October 2015 and April 2016. New Zealand does not currently support a local Ae. aegypti population, but historically has received incursives from places such as Fiji (Laird, 1951).
Figure 1.
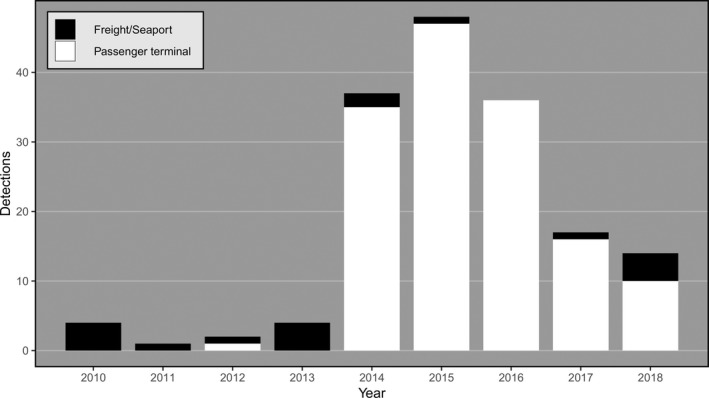
Number of Aedes aegypti detections per year at Australian international terminals. Detections at passenger airline terminals are shown in white, while those at seaports or freight terminals are in black. Each detection represents either a single adult retrieved from a trap or one or more larvae or pupae retrieved from an ovitrap
When dispersing by flight alone, Ae. aegypti is a weak disperser that prefers to remain close to human dwellings and sites of activity (Harrington et al., 2005). Its current distribution has been facilitated by several centuries of human transport, including terrestrial (Guagliardo et al., 2014), marine (Brown et al., 2014) and aerial (Sukehiro et al., 2013) conveyance. Studies of Ae. aegypti using genome‐wide SNPs have found genetic structure at fine scales of <5 km (Schmidt, Filipović, Hoffmann, & Rašić, 2018) and strong population differentiation at broader scales (Rašić et al., 2015, 2014). Similar SNP data sets to that used in this study have proved useful for investigating the source of newly established Ae. aegypti populations (Brown et al., 2014; Gloria‐Soria et al., 2018). The high level of genetic structuring at local and regional scales in this species suggests that genome‐wide SNP analyses are likely to be informative for determining the origin of incursive mosquitoes, provided reference mosquitoes are available in sufficient numbers from a range of potential source populations.
The increasing rate of incursive Ae. aegypti detections at Australian airports has occurred despite protocols that ensure all international flights into Australia undergo disinsection procedures (DAWR/MPI, 2016), wherein insecticides are used to kill incursives through residual and/or knockdown treatments before or after take‐off. However, dispersing pests from populations resistant to insecticides may be less affected by these measures. Although such resistance is not present in Australian Ae. aegypti(Endersby‐Harshman et al., 2017), it is common in Ae. aegypti populations globally (Endersby‐Harshman, Weeks, & Hoffmann, 2018; Ranson, Burhani, Lumjuan, & Black, 2010; Smith, Kasai, & Scott, 2016). Point mutations at the Vssc gene can confer target‐site knockdown resistance to synthetic pyrethroids in a broad range of insects (Scott et al., 2013; Scott, Yoshimizu, & Kasai, 2015; Seong et al., 2010) including Ae. aegypti(Donnelly et al., 2009). The only two chemicals registered for aircraft disinsection by the World Health Organization (WHO), permethrin and d‐phenothrin (IPCS, 2013; WHO, 2012), are synthetic pyrethroids.
In this study, we use genome‐wide SNPs to assign Ae. aegypti intercepted at international ports in Australia and New Zealand to their likely population, country or region of origin, by comparing incursives with reference mosquitoes collected from 15 locations around the world. We also genotype incursive and reference mosquitoes for three point mutations at the Vssc gene known to confer resistance to synthetic pyrethroids in Ae. aegypti(Wuliandari et al., 2015). Our results are informative to the control and monitoring of a broad range of invasive invertebrates and demonstrate how population genomic data can elucidate risks associated with species and genetic incursions.
2. MATERIALS AND METHODS
2.1. Mosquito collection
A total of 115 incursive Ae. aegypti were collected from passenger and freight terminals at airports and at seaports and Approved Arrangement facilities in Australia and New Zealand between December 27, 2012, and February 26, 2018. Mosquitoes were considered to be incursive if they fit either of two conditions: (a) they were collected at a terminal outside of Queensland, within which Ae. aegypti is endemic; or (b) they were collected within Queensland, but had alleles conferring resistance at the Vssc gene that are not normally found in Australian Ae. aegypti(Endersby‐Harshman et al., 2017). Overall, only a single incursive mosquito from Queensland was included (collected from Townsville seaport); all other incursives were collected outside the native range. The majority of incursives were collected at international airport passenger terminals (73.0%), generally around baggage areas airside. Supporting Information Appendix S1 lists data for each of the 115 incursives.
For the reference populations, we used 18 samples of Ae. aegypti collected throughout its range, with emphasis placed on regions with high frequency of passenger air traffic to Australia and New Zealand (Figure 2). This included samples from 15 locations: Bali (Indonesia), Yogyakarta (Indonesia), Kuala Lumpur (Malaysia), Bangkok (Thailand), Rio de Janeiro (Brazil), Cairns (Australia), Townsville (Australia), Nha Trang (Vietnam), Ho Chi Minh City (Vietnam), Fiji, Vanuatu, Kiribati, New Caledonia, Taiwan and Singapore. Bali, Kuala Lumpur and Rio de Janeiro were each sampled twice at different time points to determine the stability of genetic patterns. Where possible, we sampled close to airports with the highest frequency of passenger traffic into Australia. In the case of Rio de Janeiro, from which there are no direct flights into either Australia or New Zealand, the samples were intended to represent a general South American or Pan‐American reference population, with specific assignments within this continent prohibited without samples from other locations. Supporting Information Appendix S2 lists data for each of the 18 reference samples.
Figure 2.
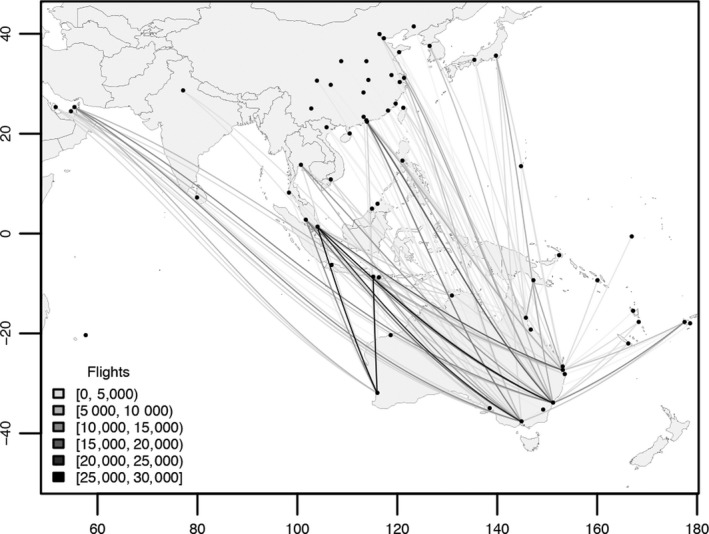
International passenger flights into Australia, 2014–2018. Total number of international passenger flights into Australian airports between January, 2014, and June, 2018, from regions in Asia, the South Pacific and the Middle East
DNA was extracted from mosquitoes using either Qiagen DNeasy Blood & Tissue Kits (Qiagen, Hilden, Germany) or Roche High Pure™ PCR Template Preparation Kits (Roche Molecular Systems, Inc., Pleasanton, CA, USA), each with an RNase A treatment step. Extracted DNA was used for SNP detection and genotyping at the Vssc gene.
2.2. SNP detection and filtering
We applied the double‐digest restriction‐site associated DNA sequencing (ddRADseq) protocol for Ae. aegyptideveloped by Rašić et al. (2014) to construct RAD libraries. The ddRADseq approach provides markers with excellent resolution at intraspecific scales (Peterson, Weber, Kay, Fisher, & Hoekstra, 2012), and is therefore suitable for both broad‐ and fine‐scale population assignment in this species, as well as for deciphering patterns of pairwise relatedness to help optimize analyses of genetic structure. Libraries were sequenced at either the Australian Genome Research Facility (AGRF, Melbourne, Australia) or the University of Melbourne (Pathology) on a HiSeq 2500 (Illumina, California, USA) in either High Output or Rapid Run modes.
We used the process_radtags program in Stacks v2.0 (Catchen, Hohenlohe, Bassham, Amores, & Cresko, 2013) to demultiplex sequence reads and trim the reads to 80 bp in length. Using a 15‐bp sliding window, low‐quality reads were discarded if the average phred score dropped below 20. Reads were aligned to the Ae. aegypti nuclear genome assembly AaegL4 (Dudchenko et al., 2017) with Bowtie v1.2.1.1 (Langmead, Trapnell, Pop, & Salzberg, 2009), allowing for a maximum of three mismatches (‐v 3 ‐‐tryhard ‐‐best). The Stacks pipeline was used in three stages: initial filtering of reference mosquitoes; establishment of reference populations from the 18 reference samples; and building a final catalog of incursive and reference mosquitoes for population assignment.
Initial filtering of reference samples aimed to remove closely related mosquitoes and those with high levels of missing data, and to normalize the number of mosquitoes in each reference population. We used the Stacks ref_map pipeline to build individual Stacks catalogs for each of the 18 reference samples, from which we called genotypes at RAD stacks at a 0.05 significance level. We generated VCF files for each catalog with the Stacks program populations. SNPs were required to be present in ≥75% of the mosquitoes and have a minor allele frequency ≥0.05 (‐r 0.75 ‐‐min_maf 0.05 ‐‐vcf). We used VCFtools (Danecek et al., 2011) to remove loci that deviated from Hardy–Weinberg equilibrium, and then thinned the data so that no two SNPs were within 250 kbp of one another. As the Ae. aegypti genome contains approximately 2.1 Mbp sequence per centimorgan (Brown, Severson, Smith, & Knudson, 2001), thinning the data in this way retains up to 8 SNPs per centimorgan, a density shown to sufficiently filter out linked loci (Cho & Dupuis, 2009). Reference mosquitoes with ≥30% of missing genotypes were omitted from all further analyses. We used SPAGeDi (Hardy & Vekemans, 2002) to calculate Loiselle's k (Loiselle, Sork, Nason, & Graham, 1995) among reference mosquitoes, identifying pairs with putative first‐degree relatedness (k ≥ 0.1875; Iacchei et al., 2013) and omitting related mosquitoes in order of missing data so that all remaining pairs had k < 0.1875. Finally, for reference samples with more than 18 mosquitoes, we omitted mosquitoes in order of missing data to reduce the number to 18.
To establish reference populations from the reference samples, we generated a new catalog containing all of the filtered reference mosquitoes. We retained SNPs that were present in ≥75% of the mosquitoes in each reference sample and that had minor allele frequencies ≥0.05. We used the R package assignPOP v1.1.4 (Chen et al., 2018) to perform Monte Carlo cross‐validation on reference samples (functions “assign.MC” and “accuracy.MC”), treating all samples as putative populations. In this process, a proportion of mosquitoes was resampled from each putative population, and this subset of mosquitoes were assigned to populations using the remaining mosquitoes from each putative population. We ran this cross‐validation multiple times, in turn resampling a proportion of 0.1, 0.2 and 0.3 of the mosquitoes from each reference sample, with 200 replicates run for each proportion.
Reference samples were treated as distinct reference populations if resampled mosquitoes were consistently assigned to their correct sample. In cases where resampled mosquitoes were assigned across multiple reference samples, we created composite reference populations from the multiple samples. If any composite reference populations contained more than 18 mosquitoes, we omitted mosquitoes in order of missing data proportion to reduce the number to 18.
A final catalog was built containing the 115 incursives and the 188 reference mosquitoes retained after filtering. This catalog was used to produce subsets of SNPs for the analyses delineated below. In each subset, we retained SNPs with minor allele frequency ≥0.05 that were present in ≥75% of the mosquitoes in each reference population and in ≥75% of incursives.
2.3. Geographical assignment of incursive Ae. aegypti
Population assignment comprised two separate processes: cluster detection with discriminant analysis of principal components (DAPC; Jombart, Devillard, & Balloux, 2010), performed in the R package adegenetv2.1.1 (Jombart, 2008); and a Monte Carlo assignment test using a support vector machine predictive model, performed in assignPOP. We used DAPC to partition reference mosquitoes and incursives into sets of clusters, and from these partitions infer population assignment. We used assignPOP to generate for each incursive a posterior probability of assignment to each of the reference populations, and from these probabilities infer confidence in assignment. Incursives showing inconsistent assignment for DAPC and assignPOP were not considered well‐assigned.
For population assignment with DAPC, we used the adegenet function “find.clusters” to establish partitions within 115 subsets of mosquitoes. Each subset included all 188 reference mosquitoes and a single incursive. We identified an optimal range for the number of clusters (K) to use for partitioning, using 1,000 principal components and 109 iterations for each K. The lower bound of this range was set as the value of K with the lowest Bayesian information criterion (BIC), and the upper bound was the value with the lowest Akaike information criterion (AIC), which provided a range of K from the conservative BIC to the less conservative AIC (Raftery, 1999). For each subset and for each value of Kwithin this range, the single incursive formed a cluster containing the incursive and one or more of the reference populations. This clustering indicated the assignment of the incursive for that value of K. Following this, we performed cluster detection among closely related reference populations, which allowed the use of more SNPs and thus more confident assignment among populations within a subregion. For incursives initially assigned to Asian populations (Bali, Bangkok, Kuala Lumpur, Singapore, Taiwan, Vietnam and Yogyakarta), we performed cluster detection using only the Asian reference populations and did likewise for incursives initially assigned to the Pacific Islands (Fiji, Kiribati, New Caledonia and Vanuatu).
For population assignment in assignPOP, we used the function “assign.X” to perform assignment tests on incursives. This analysis used the 188 reference mosquitoes to build a predictive model with which to assign the 115 incursives, using all principal components with eigenvalues >1, and a linear support vector machine classification function. This generated posterior probabilities of membership for each individual to each reference population. From each of these scores, we calculated “relative probability” of membership, which was defined as the probability of membership to the most likely population divided by the probability of membership to the second most likely population. We used these relative probabilities as an additional measure of confidence in assignment. For an incursive to be considered well‐assigned, it would need to have a relative probability >2; that is, the posterior probability of its assigned reference population would have to be at least twice that of the next most likely population.
2.4. Vssc genotyping
All incursives were genotyped for the presence of three target‐site mutations (V1016G, F1534C, S989P) in the Vssc gene known to provide resistance to synthetic pyrethroids (Du et al., 2013; Wuliandari et al., 2015). We also genotyped mosquitoes from the Bali and Fiji reference populations. The mutation site amino acid positions in this study are labelled according to the sequence of the most abundant splice variant of the house fly, Musca domestica, Vssc (GenBank accession nos. AAB47604 and AAB47605; Kasai et al., 2014). The V1016G mutation is widespread in South‐East Asia and is known to provide resistance to Type I and Type II synthetic pyrethroids when in the homozygous (GG) state (Wuliandari et al., 2015). The F1534C mutation is thought to confer resistance to Type I synthetic pyrethroids (Du et al., 2013) when homozygous. The S989P mutation alone has no effect, but in the presence of the V1016G mutation, it is thought to act synergistically, increasing resistance to Type II synthetic pyrethroids (but not Type I synthetic pyrethroids) (Hirata et al., 2014). We developed TaqMan® assays for all three target‐site mutations based on the high‐resolution melt genotyping assays in Wuliandari et al. (2015). Genotyping of each SNP was then undertaken on a LightCycler® 480 (Roche, Basel, Switzerland) real‐time PCR machine with three replicates in a 384‐well plate format.
3. RESULTS
3.1. Delineation of reference groupings
Monte Carlo cross‐validation using assignPOP showed perfect assignment of resampled mosquitoes among 8 of the 18 reference samples, indicating that these samples were genetically distinct and could be treated as separate reference populations. However, for the time‐separated pairs of reference samples from Bali, Kuala Lumpur and Rio de Janeiro, resampled mosquitoes were frequently assigned to the incorrect sample of the pair. This indicates that geographical genetic structure observed at these locations was stable over time. Additionally, resampled mosquitoes from the Cairns and Townsville reference samples showed 10% incorrect assignment with each other, which is not surprising given weak genetic structure across this area (Endersby et al., 2011), while 7% of resampled mosquitoes from the Ho Chi Minh City and Nha Trang reference samples were incorrectly assigned. Accordingly, each of these five pairs was combined into composite reference populations, leaving 13 reference populations for the assignment of incursives.
3.2. Geographical assignment of incursive Ae. aegypti
DAPC cluster detection using 41,834 SNPs gave a range for the number of clusters (K) of 3 ≤ K ≤ 13. At K = 3, populations were partitioned into a South American cluster, an Asian cluster, and a cluster containing Australia and the Pacific Islands. The Australian population split from the Pacific Islands at K = 4, following which new geographical partitions formed at each increase until K = 13 placed all reference populations into their own cluster. These same geographical partitions were established at K = 13 in each run, indicating that the inclusion of the single incursive was not biasing cluster detection.
Of the 115 incursives, 111 demonstrated clustering for 3 ≤ K ≤ 12 that was consistent with their final cluster assignment at K = 13 (Appendix S1). The eight incursives collected at New Zealand terminals all showed consistent clustering with the Pacific Islands, while the incursives collected in Australia clustered with South‐East Asia, Australia and South America. Supporting Information Figure S1 shows a typical stepwise delineation of clusters for 3 ≤ K ≤ 13 for incursive IPT33.
Posterior probabilities of membership to the 13 reference populations calculated for incursives using assignPOPwere of range 0.16–0.82. Relative probabilities (see Section 22) ranged from 1.01 to 29.00. For incursives consistently assigned to either of the Asian or Pacific Islands clusters, we repeated assignment tests using only those reference populations, which allowed us to increase confidence in assignment by incorporating more SNPs: 55,443 and 146,142, respectively. Overall, 92 incursives (80%) had relative probabilities >2, while 23 (20%) had relative probabilities <2. Comparing assignment between DAPC and assignPOP, 112 of the 115 incursives (97.4%) had highest posterior probabilities for the same reference population they were assigned to at K = 13 (Supporting Information Appendix S1), indicating strong agreement between the two assignment methods.
Figure 3 shows the relationship between posterior probability, relative probability and proportion of missing data. The incursives with relative probabilities >2 (white and black circles) formed a group in which posterior probability of membership decreased as missing data increased. Five incursives with relative probabilities >2 were exceptions to this (black circles), grouping with the incursives with relative probabilities <2 (black squares). This latter group (black circles and squares) had low posterior probabilities and mostly low missing data, and these variables were not well correlated (linear regression: R 2 = 0.023, F 27 = 0.609, p = 0.442). Among the incursives of the first group (white circles), there was a strong negative correlation between posterior probability of membership and proportion of missing data (linear regression: R 2 = 0.795, F 86 = 329.6, p < 0.001), indicating that lower posterior probabilities in this group were likely due to missing data (e.g., from imperfect preservation of specimens). Conversely, 25 of the 28 incursives in the second group (black circles and squares) had posterior probabilities <0.5 despite having <0.13 missing data (Figure 3). As the low posterior and relative probabilities of these incursives could not be explained by high missing data, it seems likely that these incursives were from source populations not sampled in this study.
Figure 3.
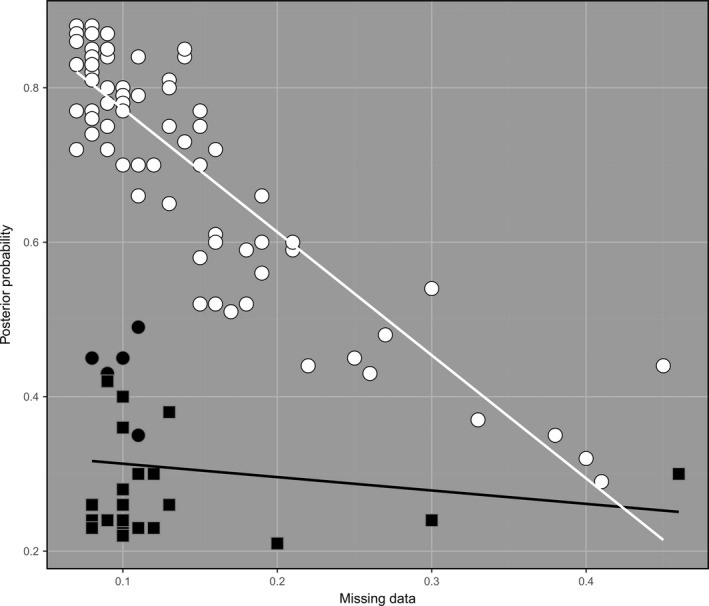
Posterior probabilities and proportion of missing data for the 115 incursive Aedes aegypti. White and black circles indicate incursives with relative probabilities >2, while black squares indicate incursives with relative probabilities <2. Incursives marked in white were considered well‐assigned, while those marked in black were considered poorly assigned. Among well‐assigned incursives, lower posterior probabilities correlated strongly with higher missing data (linear regression: R 2 = 0.795, F 86 = 329.6, p < 0.001). Among poorly assigned incursives, there was no similar relationship (linear regression: R 2 = 0.023, F 27 = 0.609, p = 0.442)
We considered the 87 incursives (75.7%) represented as white circles in Figure 3 as well‐assigned. These all showed consistent assignment for DAPC and assignPOP and consistent cluster assignment for 3 ≤ K ≤ 13, and had relative probabilities >2. Figure 4 shows a DAPC of the well‐assigned incursives plotted with the 13 reference populations. The well‐assigned incursives were assigned to the following source populations: 62 to Bali, 13 to Kuala Lumpur, 7 to Rio de Janeiro, 3 to Australia, 1 to Fiji and 1 to Bangkok (Supporting Information Appendix S1). The incursive assigned to Fiji was detected in New Zealand and was the only well‐assigned New Zealand incursive; the incursives assigned elsewhere were all detected in Australia. Supporting Information Figure S2 shows a DAPC of the seven Asian reference populations (Bali, Bangkok, Kuala Lumpur, Singapore, Taiwan, Vietnam, Yogyakarta) plotted with the 76 well‐assigned incursives putatively from Asia.
Figure 4.
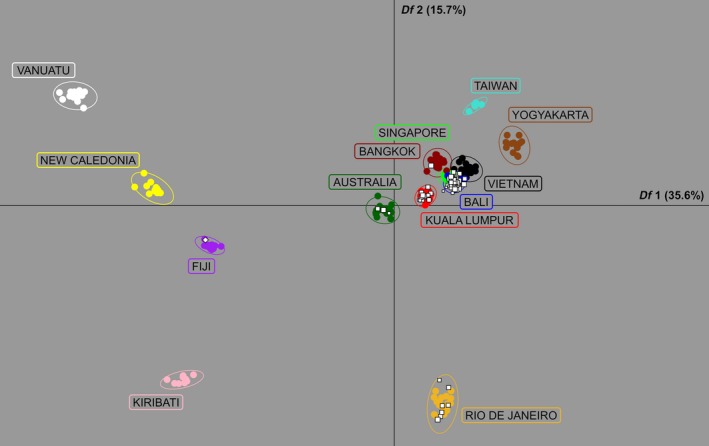
Discriminant analysis of principal components (DAPC). The plot shows the first and second discriminant functions of a DAPC of all 188 reference Aedes aegypti and all 87 well‐assigned incursive Ae. aegypti, using 41,834 SNPs and 100 principal components. White squares in normal orientation show incursives intercepted at Australian terminals, while white squares rotated 45° show incursives intercepted at New Zealand terminals. Squares are sized by a logarithmic function indicating relative probability of membership (see Section 22)
3.3. Vssc genotyping
TaqMan® (Life Technologies, CA, USA) assays for the three target‐site mutations showed that the most common genotype found in incursive mosquitoes across the three loci was homozygous resistant at V1016G, homozygous susceptible at F1534C, homozygous resistant at S989P (GG/TT/CC—the letters refer to the mutant or wild‐type bases at each of the loci, rather than the amino acid notation). This genotype putatively confers strong resistance to Type I and Type II synthetic pyrethroids (Wuliandari et al., 2015). Of the 72 incursives with the GG/TT/CC genotype, 62 were assigned to Bali, 5 were assigned to Kuala Lumpur, 1 was assigned to Bangkok, and 4 were uncertain but had a likely Asian origin. As well as occurring in 100% of putatively Balinese incursives, this genotype was also found in all reference mosquitoes from Bali.
Another common genotype among incursives was homozygous susceptible at V1016G, homozygous resistant at F1534C, homozygous susceptible at S989P (TT/GG/TT), which putatively confers strong resistance to Type I synthetic pyrethroids (Du et al., 2013). This genotype was found in 18 incursive Ae. aegyptiand 9 of the 11 Fijian reference mosquitoes. Of these incursives, seven were assigned to Rio de Janeiro, and one each to Bangkok and Fiji; the remainder were uncertain. Overall, 95 incursives (82.6%) had genotypes putatively conferring strong resistance to Type I synthetic pyrethroids.
Among all incursives, only three had the genotype that lacks all three resistance‐associated point mutations (TT/TT/TT). These incursives were all well‐assigned to the Australian reference population, indicating that they were likely transported via domestic means rather than an international pathway. This genotype was also found in one of the Fijian reference mosquitoes.
4. DISCUSSION
Assignment tests using genome‐wide SNPs were able to identify the global source populations from which pyrethroid‐resistant Ae. aegypti disperse into Australia and New Zealand. We observed evidence for different dispersal pathways into Australia and into New Zealand, as all the incursives intercepted in Australia were assigned to South‐East Asia, South America and Australia, while all the incursives intercepted in New Zealand were assigned to the Pacific Islands. Among well‐assigned incursives, most (71.2%) were assigned to Bali. All incursives assigned to Bali were collected airside at international passenger terminals in Australia, with the majority detected in Perth (56.3%), one of the most active routes between Australia and Denpasar International Airport in Bali (Figure 2). All Ae. aegyptisuspected (incursives) and known (reference mosquitoes) to be from Bali shared a Vsscgenotype (1016/1534/989; GG/TT/CC) linked to strong resistance to Type I and Type II synthetic pyrethroids (Wuliandari et al., 2015). Most of the suspected and known Fijian Ae. aegypti shared a genotype (TT/GG/TT) linked to strong resistance to Type I synthetic pyrethroids (Du et al., 2013). If the resistance observed in laboratory studies is indicative of field resistance, then the WHO aircraft disinsection protocols that rely on the use of Type I synthetic pyrethroids (IPCS, 2013; WHO, 2012) are unlikely to stop the dispersal of Ae. aegyptifrom regions where resistance to these insecticides is common. The protocols used in this study may be implementable for Ae. aegyptiand other invasive species internationally, which may help indicate the extent to which pyrethroid‐based protocols alone are insufficient for achieving global biosecurity objectives.
Our results strongly suggest that resistance to synthetic pyrethroids used during aircraft disinsection is likely to be contributing to the detections of Ae. aegypti at the Australian and New Zealand borders. There are currently only two chemicals registered for aircraft disinsection by the WHO, permethrin and d‐phenothrin (IPCS, 2013; WHO, 2012); both are Type I synthetic pyrethroids (permethrin is first generation; d‐phenothrin is second generation). Resistance to these pyrethroids varies substantially throughout populations of Ae. aegpyti worldwide (Smith et al., 2016), though Australian populations remain wholly susceptible (Endersby‐Harshman et al., 2017). Of the 115 incursive mosquitoes analysed in this study, only three incursives had the wholly susceptible genotype (TT/TT/TT). These were collected in Brisbane, Australia, and were assigned to the Australian population, and thus are likely to have been transported on a domestic transport route where disinsection was not conducted as required on all international flights to Australia (DAWR/MPI, 2016). Within most of the non‐Australian Ae. aegypti reference populations in this study, there is considerable diversity in resistance genotypes (Endersby‐Harshman et al., 2018; unpublished data), with populations made up of homozygotes with theoretically strong resistance, susceptible wild‐type homozygotes and heterozygotes of varying resistance status. Our finding that all incursive mosquitoes from international sources had alleles associated with resistance to Type I pyrethroids, and that 82.6% were likely to have displayed strong levels of resistance, suggests that aircraft disinsection may be successfully eliminating pyrethroid‐susceptible mosquitoes, while pyrethroid resistance may be permitting others to survive and reach the Australian and New Zealand borders.
International incursions of pyrethroid‐resistant Ae. aegypti into Australia present three main management challenges. They could introduce alleles conferring pyrethroid resistance into the current, pyrethroid‐susceptible Australian population and thereby reduce the effectiveness of current control programmes; they could bring disease into nonendemic regions (Siala et al., 2015); and they could lead to the establishment of new Australian Ae. aegypti populations outside the current native range in Queensland. Considering the current high frequency at which resistant Ae. aegypti are being detected at the Australian border, it is critical that future research focuses on determining whether the target‐site mutations provide “operational resistance” to disinsection and whether this differs for the various methods used for disinsection (e.g., preflight and top‐of‐descent vs. pre‐embarkation/residual treatment; DAWR/MPI, 2016). If operational resistance is shown to disinsection, then management of Ae. aegypti may need to focus more on suppressing populations at source locations. Interestingly, there has been a large reduction in the number of Ae. aegypti incursives detected at Australian airports over the last 2 years (Figure 1), which coincides with discussions between the Australian and Indonesian Governments as the results of this study were becoming available and may indicate a change in density of local mosquito populations in Bali through either local control efforts or other local dynamics. Nevertheless, due to the likely high incidence of pyrethroid resistance in these populations, it may be more effective to use noninsecticidal methods to suppress them, such as by releasing Ae. aegypti males that are sterile or that carry an introduced infection of endosymbiotic bacteria (McGraw & O'Neill, 2013). Local suppression around Denpasar International Airport may also help reduce the dengue burden in this area, which has one of the highest incidences of dengue in Bali (Purnama & Baskoro, 2013). Noninsecticidal suppression may also be useful for eliminating small populations of resistant mosquitoes in newly invaded regions.
This study has applied protocols for the assignment of incursive individuals to their likely population of origin. Following their application to Ae. aegypti, our protocols could be applied to other invasive species that are also frequently detected at borders. One candidate would be the related dengue vector Ae. albopictus, which has invaded and continues to invade tropical and temperate regions globally (Benedict, Levine, Hawley, & Lounibos, 2007; Kraemer et al., 2015), including the Torres Strait Islands in northern Australia (Ritchie et al., 2006), from which it threatens invasion of the mainland. This approach may be limited by the weak genetic structure between Ae. albopictuspopulations compared with Ae. aegypti(Goubert, Minard, Vieira, & Boulesteix, 2016), even when investigated with high‐density markers (Schmidt et al., 2017). However, genome‐level population genetic studies on Ae. albopictus are currently scarce, and a large database of reference samples may be necessary to identify distinct populations and produce confident assignments as in this study.
For invasive species with population differentiation equivalent to or greater than that of Ae. aegypti, the methods used in this study are likely to provide similar sensitivity for determining the origin of incursions. Our methodology could also be applied for postborder detections, where newly invasive species are detected and there is a need to determine their likely source. For instance, Diuraphis noxia (Russian wheat aphid) was first detected in Australia in 2016, and successful control of this species now depends, in part, on identifying past and present sources of incursions (Yazdani et al., 2018). Although a recent genome assembly of D. noxia suggests that this species has low genetic variability (Burger & Botha, 2017), genome‐wide SNPs may yet provide sufficient power to determine broad patterns of gene flow. Likewise, similar methodologies may help untangle the complexity of marine invasion narratives, in which many processes remain cryptic when analysed without the appropriate molecular tools (Ojaveer et al., 2014).
5. CONCLUSIONS
Current WHO protocols for preventing the unintentional dispersal of invasive invertebrates rely upon Type I synthetic pyrethroids for disinsection of aircraft. This study demonstrates that these measures are likely to have been insufficient for preventing the dispersal of resistant strains of the major disease vector, Ae. aegypti, to the Australian and New Zealand borders through international air traffic. Most Ae. aegypti collected in Australia came from Bali, while all those collected in New Zealand came from Pacific Islands like Fiji. A large majority of incursives (82.6%) had point mutations at the Vssc gene that putatively confer strong resistance to Type I or Type I and Type II synthetic pyrethroids. These mutations were found at high frequency in the greater Balinese and Fijian Ae. aegypti populations. Our findings indicate that insecticide‐based strategies are likely to be limited in their capacity to stop biological invasions when insecticide resistance is present, and alternative control strategies should be considered, particularly for important disease vectors such as Ae. aegypti.
CONFLICT OF INTEREST
None declared.
Supporting information
ACKNOWLEDGEMENTS
We thank Ashley Callahan, Jason Axford, Tim Hurst, Stephen Doggett, Elizabeth Valerie, Craig Williams and Joe Davis for the collection of reference samples used in this study. We also thank Ashley Callahan for conducting some of the DNA extraction and library preparation. We thank Gordana Rašić for input in conceptualization and study design. We thank staff from the Department of Agriculture and Water Resources (DAWR), Australian Government, for the collection of all incursive mosquitoes and DAWR for providing funding for this work. AAH was supported by Program and Fellowship grants from the National Health and Medical Research Council (NHMRC), no. 1037003. TLS and AAH were also supported by the Wellcome Trust UK, no. 108508. These grants provided the necessary support to establish the reference database used in this study.
Schmidt TL, van Rooyen AR, Chung J, et al. Tracking genetic invasions: Genome‐wide single nucleotide polymorphisms reveal the source of pyrethroid‐resistant Aedes aegypti (yellow fever mosquito) incursions at international ports. Evol Appl. 2019;12:1136–1146. 10.1111/eva.12787
Contributor Information
Ary A. Hoffmann, Email: ary@unimelb.edu.au
Andrew R. Weeks, Email: aweeks@unimelb.edu.au
DATA ACCESSIBILITY
Aligned.bam files for 188 reference and 115 incursive Ae. aegypti have been archived at NCBI Genbank, accessible with accession number PRJNA522930.
REFERENCES
- Bacon, S. J. , Bacher, S. , & Aebi, A. (2012). Gaps in border controls are related to quarantine alien insect invasions in Europe. PLoS ONE, 7(10), e47689 10.1371/journal.pone.0047689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benedict, M. Q. , Levine, R. S. , Hawley, W. A. , & Lounibos, L. P. (2007). Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus . Vector‐Borne and Zoonotic Diseases, 7(1), 76–85. 10.1089/vbz.2006.0562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackburn, T. M. , Cassey, P. , Duncan, R. P. , Evans, K. L. , & Gaston, K. J. (2004). Avian extinction and mammalian introductions on oceanic islands. Science, 305(5692), 1955–1958. 10.1126/science.1101617 [DOI] [PubMed] [Google Scholar]
- Bogoch, I. I. , Brady, O. J. , Kraemer, M. U. G. , German, M. , Creatore, M. I. , Kulkarni, M. A. , … Khan, K. (2016). Anticipating the international spread of Zika virus from Brazil. Lancet (London, England), 387(10016), 335–336. 10.1016/S0140-6736(16)00080-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, J. E. , Evans, B. R. , Zheng, W. , Obas, V. , Barrera‐Martinez, L. , Egizi, A. , … Powell, J. R. (2014). Human impacts have shaped historical and recent evolution in Aedes aegypti, the dengue and yellow fever mosquito. Evolution, 68(2), 514–525. 10.1111/evo.12281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, S. E. , Severson, D. W. , Smith, L. A. , & Knudson, D. L. (2001). Integration of the Aedes aegypti Mosquito Genetic Linkage and Physical Maps. Genetics, 157(3), 1299–1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burger, N. F. V. , & Botha, A.‐M. (2017). Genome of Russian wheat aphid an economically important cereal aphid. Standards in Genomic Sciences, 12(1), 90 10.1186/s40793-017-0307-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caley, P. , Ingram, R. , & De Barro, P. (2015). Entry of exotic insects into Australia: Does border interception count match incursion risk? Biological Invasions, 17(4), 1087–1094. 10.1007/s10530-014-0777-z [DOI] [Google Scholar]
- Catchen, J. , Hohenlohe, P. A. , Bassham, S. , Amores, A. , & Cresko, W. A. (2013). Stacks: An analysis tool set for population genomics. Molecular Ecology, 22(11), 3124–3140. 10.1111/mec.12354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, K.‐Y. , Marschall, E. A. , Sovic, M. G. , Fries, A. C. , Gibbs, H. L. , & Ludsin, S. A. (2018). assignPOP: An R package for population assignment using genetic, non‐genetic, or integrated data in a machine‐learning framework. Methods in Ecology and Evolution, 9(2), 439–446. 10.1111/2041-210X.12897 [DOI] [Google Scholar]
- Cho, K. , & Dupuis, J. (2009). Handling linkage disequilibrium in qualitative trait linkage analysis using dense SNPs: A two‐step strategy. BMC Genetics, 10(1), 44 10.1186/1471-2156-10-44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danecek, P. , Auton, A. , Abecasis, G. , Albers, C. a. , Banks, E. , DePristo, M. a. , … Durbin, R. (2011). The variant call format and VCFtools. Bioinformatics, 27(15), 2156–2158. 10.1093/bioinformatics/btr330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DAWR/MPI (2016). Schedule of aircraft disinsection procedures for flights into Australia and New Zealand. Version 4.0. Department of Agriculture and Water Resources, Canberra, and Ministry for Primary Industries, Wellington.
- Donnelly, M. J. , Corbel, V. , Weetman, D. , Wilding, C. S. , Williamson, M. S. , & Black, W. C. (2009). Does kdr genotype predict insecticide‐resistance phenotype in mosquitoes? Trends in Parasitology, 25(5), 213–219. 10.1016/J.PT.2009.02.007 [DOI] [PubMed] [Google Scholar]
- Du, Y. , Nomura, Y. , Satar, G. , Hu, Z. , Nauen, R. , He, S. y. , … Dong, K. (2013). Molecular evidence for dual pyrethroid‐receptor sites on a mosquito sodium channel. Proceedings of the National Academy of Sciences of the United States of America, 110(29), 11785–11790. 10.1073/pnas.1305118110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudchenko, O. , Batra, S. S. , Omer, A. D. , Nyquist, S. K. , Hoeger, M. , Durand, N. C. , Aiden, E. L. (2017). De novo assembly of the Aedes aegypti genome using Hi‐C yields chromosome‐length scaffolds. Science (New York, NY), 356(6333), 92–95. 10.1126/science.aal3327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edman, J. D. , Kittayapong, P. , Coleman, R. C. , Clark, G. G. , Sithiprasasna, R. , Kitthawee, S. , … Harrington, L. C. (2005). Dispersal of the dengue vector Aedes aegypti within and between rural communities. American Journal of Tropical Medicine and Hygiene, 72(2), 209–220. 10.4269/ajtmh.2005.72.209 [DOI] [PubMed] [Google Scholar]
- Endersby, N. M. , Hoffmann, A. A. , White, V. L. , Ritchie, S. A. , Johnson, P. H. , & Weeks, A. R. (2011). Changes in the genetic structure of Aedes aegypti (Diptera: Culicidae) populations in Queensland, Australia, across two seasons: Implications for potential mosquito releases. Journal of Medical Entomology, 48(5), 999–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endersby‐Harshman, N. M. , Weeks, A. R. , & Hoffmann, A. A. (2018). The detection and significance of emerging insecticide resistance in mosquitoes. Microbiology Australia, 39, 80–83. 10.1071/MA18022 [DOI] [Google Scholar]
- Endersby‐Harshman, N. M. , Wuliandari, J. R. , Harshman, L. G. , Frohn, V. , Johnson, B. J. , Ritchie, S. A. , & Hoffmann, A. A. (2017). Pyrethroid susceptibility has been maintained in the dengue vector, Aedes aegypti (Diptera: Culicidae), in Queensland, Australia. Journal of Medical Entomology, 54(6), 1649–1658. 10.1093/jme/tjx145 [DOI] [PubMed] [Google Scholar]
- Gloria‐Soria, A. , Lima, A. , Lovin, D. D. , Cunningham, J. M. , Severson, D. W. , & Powell, J. R. (2018). Origin of a High‐Latitude Population of Aedes aegypti in Washington, DC. American Journal of Tropical Medicine and Hygiene, 98(2), 445–452. 10.4269/ajtmh.17-0676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goubert, C. , Minard, G. , Vieira, C. , & Boulesteix, M. (2016). Population genetics of the Asian tiger mosquito Aedes albopictus, an invasive vector of human diseases. Heredity, 117(3), 125–134. 10.1038/hdy.2016.35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guagliardo, S. A. , Barboza, J. L. , Morrison, A. C. , Astete, H. , Vazquez‐Prokopec, G. , & Kitron, U. (2014). Patterns of geographic expansion of Aedes aegypti in the Peruvian Amazon. PLoS Neglected Tropical Diseases, 8(8), e3033 10.1371/journal.pntd.0003033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardy, O. J. , & Vekemans, X. (2002). SPAGeDi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Molecular Ecology Notes, 2(4), 618–620. 10.1046/j.1471-8286.2002.00305.x [DOI] [Google Scholar]
- Hejda, M. , Pyšek, P. , & Jarošík, V. (2009). Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 97(3), 393–403. 10.1111/j.1365-2745.2009.01480.x [DOI] [Google Scholar]
- Hirata, K. , Komagata, O. , Itokawa, K. , Yamamoto, A. , Tomita, T. , & Kasai, S. (2014). A single crossing‐over event in voltage‐sensitive Na+ channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Neglected Tropical Diseases, 8(8), e3085 10.1371/journal.pntd.0003085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hulme, P. E. (2009). Trade, transport and trouble: Managing invasive species pathways in an era of globalization. Journal of Applied Ecology, 46(1), 10–18. 10.1111/j.1365-2664.2008.01600.x [DOI] [Google Scholar]
- Iacchei, M. , Ben‐Horin, T. , Selkoe, K. A. , Bird, C. E. , García‐Rodríguez, F. J. , & Toonen, R. J. (2013). Combined analyses of kinship and F ST suggest potential drivers of chaotic genetic patchiness in high gene‐flow populations. Molecular Ecology, 22(13), 3476–3494. 10.1111/mec.12341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- IPCS (2013). Aircraft disinsection insecticides (Environmental health criteria 243). Geneva, UK: European Commission and the Policy Research Programme. [Google Scholar]
- IUCN (2000). Guidelines for the prevention of biodiversity loss caused by alien invasive species. Gland, Switzerland: International Union for the Conservation of Nature. [Google Scholar]
- Jombart, T. (2008). adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics, 24(11), 1403–1405. 10.1093/bioinformatics/btn129 [DOI] [PubMed] [Google Scholar]
- Jombart, T. , Devillard, S. , & Balloux, F. (2010). Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genetics, 11(1), 94–108. 10.1186/1471-2156-11-94 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasai, S. , Komagata, O. , Itokawa, K. , Shono, T. , Ng, L. C. , Kobayashi, M. , & Tomita, T. (2014). Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: target site insensitivity, penetration, and metabolism. PLoS Neglected Tropical Diseases, 8(6), e2948 10.1371/journal.pntd.0002948 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearney, M. , Porter, W. P. , Williams, C. , Ritchie, S. , & Hoffmann, A. A. (2009). Integrating biophysical models and evolutionary theory to predict climatic impacts on species' ranges: The dengue mosquito Aedes aegypti in Australia. Functional Ecology, 23(3), 528–538. 10.1111/j.1365-2435.2008.01538.x [DOI] [Google Scholar]
- Kraemer, M. U. G. , Sinka, M. E. , Duda, K. A. , Mylne, A. Q. N. , Shearer, F. M. , Barker, C. M. , Hay, S. I. (2015). The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus . Elife, 4, e08347 10.7554/eLife.08347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird, M. (1951). Insects collected from Aircraft arriving in New Zealand from Abroad. Zoology Publications, Victoria University Collection, (11). Retrieved from https://www.cabdirect.org/cabdirect/abstract/19542901358
- Langmead, B. , Trapnell, C. , Pop, M. , & Salzberg, S. L. (2009). Ultrafast and memory‐efficient alignment of short DNA sequences to the human genome. Genome Biology, 10(3), R25 10.1186/gb-2009-10-3-r25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levine, J. M. , & D'Antonio, C. M. (2003). Forecasting biological invasions with increasing international trade. Conservation Biology, 17(1), 322–326. 10.1046/j.1523-1739.2003.02038.x [DOI] [Google Scholar]
- Loiselle, B. A. , Sork, V. L. , Nason, J. , & Graham, C. (1995). Spatial genetic structure of a tropical understory shrub, Psychotria officinalis (Rubiaceae). American Journal of Botany, 82(11), 1420–1425. 10.1002/j.1537-2197.1995.tb12679.x [DOI] [Google Scholar]
- Lounibos, L. P. (2002). Invasions by insect vectors of human disease. Annual Review of Entomology, 47(1), 233–266. 10.1146/annurev.ento.47.091201.145206 [DOI] [PubMed] [Google Scholar]
- McGeoch, M. A. , Butchart, S. H. M. , Spear, D. , Marais, E. , Kleynhans, E. J. , Symes, A. , … Hoffmann, M. (2010). Global indicators of biological invasion: Species numbers, biodiversity impact and policy responses. Diversity and Distributions, 16(1), 95–108. 10.1111/j.1472-4642.2009.00633.x [DOI] [Google Scholar]
- McGraw, E. A. , & O'Neill, S. L. (2013). Beyond insecticides: New thinking on an ancient problem. Nature Reviews Microbiology, 11(3), 181–193. 10.1038/nrmicro2968 [DOI] [PubMed] [Google Scholar]
- Mehta, S. V. , Haight, R. G. , Homans, F. R. , Polasky, S. , & Venette, R. C. (2007). Optimal detection and control strategies for invasive species management. Ecological Economics, 61(2–3), 237–245. 10.1016/J.ECOLECON.2006.10.024 [DOI] [Google Scholar]
- Myers, J. H. , Simberloff, D. , Kuris, A. M. , & Carey, J. R. (2000). Eradication revisited: Dealing with exotic species. Trends in Ecology & Evolution, 15(8), 316–320. 10.1016/S0169-5347(00)01914-5 [DOI] [PubMed] [Google Scholar]
- Ojaveer, H. , Galil, B. S. , Minchin, D. , Olenin, S. , Amorim, A. , Canning‐Clode, J. , … Zenetos, A. (2014). Ten recommendations for advancing the assessment and management of non‐indigenous species in marine ecosystems. Marine Policy, 44, 160–165. 10.1016/J.MARPOL.2013.08.019 [DOI] [Google Scholar]
- Peterson, B. K. , Weber, J. N. , Kay, E. H. , Fisher, H. S. , & Hoekstra, H. E. (2012). Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non‐model species. PLoS ONE, 7(5), e37135 10.1371/journal.pone.0037135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puckett, E. E. , & Eggert, L. S. (2016). Comparison of SNP and microsatellite genotyping panels for spatial assignment of individuals to natal range: A case study using the American black bear (Ursus americanus). Biological Conservation, 193, 86–93. 10.1016/J.BIOCON.2015.11.020 [DOI] [Google Scholar]
- Purnama, S. , & Baskoro, T. (2013). Maya Index and density of Aedes aegypti larvae of the dengue infection. Makara Journal of Health Research, 11(3), 57–64. [Google Scholar]
- Purnima, R. , Raghubanshi, A. S. , & Singh, J. S. (2008). Impact of invasive alien plant species on soil processes: A review. Proceedings of the National Academy of Sciences India. Section B, Biological Sciences, 78(4), 288–298. [Google Scholar]
- Raftery, A. E. (1999). Bayes factors and BIC: Comment on “A critique of the Bayesian information criterion for model selection”. Sociological Methods & Research, 27(3), 411–427. 10.1177/0049124199027003005 [DOI] [Google Scholar]
- Ranson, H. , Burhani, J. , Lumjuan, N. , & Black, W. C. I. (2010). Insecticide resistance in dengue vectors. Retrieved from http://archive.lstmliverpool.ac.uk/999/?utm_source=rss&utm_medium=rss&utm_campaign=insecticide-resistance-in-dengue-vectors
- Rašić, G. , Endersby‐Harshman, N. , Tantowijoyo, W. , Goundar, A. , White, V. , Yang, Q. , … Arguni, E. (2015). Aedes aegypti has spatially structured and seasonally stable populations in Yogyakarta, Indonesia. Parasites & Vectors, 8(1), 610 10.1186/s13071-015-1230-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rašić, G. , Filipović, I. , Weeks, A. R. , & Hoffmann, A. A. (2014). Genome‐wide SNPs lead to strong signals of geographic structure and relatedness patterns in the major arbovirus vector, Aedes aegypti . BMC Genomics, 15(1), 275 10.1186/1471-2164-15-275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie, S. A. , Moore, P. , Carruthers, M. , Williams, C. , Montgomery, B. , Foley, P. , … Russell, R. C. (2006). Discovery of a widespread infestation of Aedes albopictus in the Torres Strait, Australia. Journal of the American Mosquito Control Association, 22(3), 358–365. 10.2987/8756-971X(2006)22[358:DOAWIO]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- Robinson, A. , Burgman, M. A. , & Cannon, R. (2011). Allocating surveillance resources to reduce ecological invasions: Maximizing detections and information about the threat. Ecological Applications, 21(4), 1410–1417. 10.1890/10-0195.1 [DOI] [PubMed] [Google Scholar]
- Russell, R. C. , Currie, B. J. , Lindsay, M. D. , Mackenzie, J. S. , Ritchie, S. A. , & Whelan, P. I. (2009). Dengue and climate change in Australia: Predictions for the future should incorporate knowledge from the past. Medical Journal of Australia, 190(5), 265–268. [DOI] [PubMed] [Google Scholar]
- Schmidt, T. L. , Filipović, I. , Hoffmann, A. A. , & Rašić, G. (2018). Fine‐scale landscape genomics helps explain the slow spatial spread of Wolbachia through the Aedes aegypti population in Cairns, Australia. Heredity, 120(5), 386–395. 10.1038/s41437-017-0039-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt, T. L. , Rašić, G. , Zhang, D. , Zheng, X. , Xi, Z. , & Hoffmann, A. A. (2017). Genome‐wide SNPs reveal the drivers of gene flow in an urban population of the Asian Tiger Mosquito, Aedes albopictus . PLOS Neglected Tropical Diseases, 11(10), e0006009 10.1371/journal.pntd.0006009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott, J. G. , Leichter, C. A. , Rinkevihc, F. D. , Harris, S. A. , Su, C. , Aberegg, L. C. , … Zurek, L. (2013). Insecticide resistance in house flies from the United States: Resistance levels and frequency of pyrethroid resistance alleles. Pesticide Biochemistry and Physiology, 107(3), 377–384. 10.1016/J.PESTBP.2013.10.006 [DOI] [PubMed] [Google Scholar]
- Scott, J. G. , Yoshimizu, M. H. , & Kasai, S. (2015). Pyrethroid resistance in Culex pipiens mosquitoes. Pesticide Biochemistry and Physiology, 120, 68–76. 10.1016/J.PESTBP.2014.12.018 [DOI] [PubMed] [Google Scholar]
- Seong, K. M. , Lee, D.‐Y. , Yoon, K. S. , Kwon, D. H. , Kim, H. C. , Klein, T. A. , … Lee, S. H. (2010). Establishment of quantitative sequencing and filter contact vial bioassay for monitoring pyrethroid resistance in the common bed bug, Cimex lectularius . Journal of Medical Entomology, 47(4), 592–599. 10.1093/jmedent/47.4.592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siala, E. , Gamara, D. , Kallel, K. , Daaboub, J. , Zouiten, F. , Houzé, S. , … Aoun, K. (2015). Airport malaria: Report of four cases in Tunisia. Malaria Journal, 14(1), 42 10.1186/s12936-015-0566-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, C. E. G. (1956). The history of dengue in tropical Asia and its probable relationship to the mosquito Aedes aegypti . Journal of Tropical Medicine and Hygiene, 59(10), 243–251. [PubMed] [Google Scholar]
- Smith, L. B. , Kasai, S. , & Scott, J. G. (2016). Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pesticide Biochemistry and Physiology, 133, 1136–12. 10.1016/J.PESTBP.2016.03.005 [DOI] [PubMed] [Google Scholar]
- Sukehiro, N. , Kida, N. , Umezawa, M. , Murakami, T. , Arai, N. , Jinnai, T. , … Tsuda, Y. (2013). First report on invasion of yellow fever mosquito, Aedes aegypti, at Narita International Airport, Japan in August 2012. Japanese Journal of Infectious Diseases, 66(3), 189–194. 10.7883/yoken.66.189 [DOI] [PubMed] [Google Scholar]
- Weaver, S. C. , & Lecuit, M. (2015). Chikungunya virus and the global spread of a mosquito‐borne disease. New England Journal of Medicine, 372(13), 1231–1239. 10.1056/NEJMra1406035 [DOI] [PubMed] [Google Scholar]
- Westphal, M. I. , Browne, M. , MacKinnon, K. , & Noble, I. (2008). The link between international trade and the global distribution of invasive alien species. Biological Invasions, 10(4), 391–398. 10.1007/s10530-007-9138-5 [DOI] [Google Scholar]
- WHO (2012). Guidelines for testing the efficacy of insecticide products used in aircraft. Geneva, Switzerland: WHO. [Google Scholar]
- Wilson, J. R. U. , Dormontt, E. E. , Prentis, P. J. , Lowe, A. J. , & Richardson, D. M. (2009). Something in the way you move: Dispersal pathways affect invasion success. Trends in Ecology & Evolution, 24(3), 136–144. 10.1016/J.TREE.2008.10.007 [DOI] [PubMed] [Google Scholar]
- Wuliandari, J. , Lee, S. , White, V. , Tantowijoyo, W. , Hoffmann, A. A. , & Endersby‐Harshman, N. M. (2015). Association between three mutations, F1565C, V1023G and S996P, in the voltage‐sensitive sodium channel gene and knockdown resistance in Aedes aegypti from Yogyakarta, Indonesia. Insects, 6(3), 658–685. 10.3390/insects6030658 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yazdani, M. , Baker, G. , DeGraaf, H. , Henry, K. , Hill, K. , Kimber, B. , … Nash, M. A. (2018). First detection of Russian wheat aphid Diuraphis noxia Kurdjumov (Hemiptera: Aphididae) in Australia: A major threat to cereal production. Austral Entomology, 57(4), 410–417. 10.1111/aen.12292 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Aligned.bam files for 188 reference and 115 incursive Ae. aegypti have been archived at NCBI Genbank, accessible with accession number PRJNA522930.