

Abstract
DpbCasX, also called Cas12e, is an RNA-guided DNA endonuclease isolated from Deltaproteobacteria. In this paper I characterized the CasX-compatible genome editing sites in the reference genomes of yeast (Saccharomyces cerevisiae), flatworms (Caenorhabditis elegans), flies (Drosophila melanogaster), zebrafish (Danio rerio), mouse (Mus musculus), rats (Rattus norvegicus), and humans (Homo sapiens). Across those genomes there were > 27,000 CasX sites per megabase on average. More than 90% of genes in each genome had at least one unique site overlapping an exon, with median unique sites per gene of 6–45. I also annotated sites in the GRCm38 reference and 15 additional mouse strain genomes. The presence of specific guide sequences varied amongst the strains, with CAST/EiJ and PWK/PhJ showing the greatest divergence from the reference strain. The high density of CasX sites and number of exon overlapping sites suggests that CasX has the potential to be used as a common genome editor.
Electronic supplementary material
The online version of this article (10.1186/s12864-019-5924-6) contains supplementary material, which is available to authorized users.
Keywords: Genome editing, CasX, Cas12e, Saccharomyces cerevisiae, Caenorhabditis elegans, Drosophila melanogaster, Danio rerio, Mus musculus, Rattus norvegicus, Homo sapiens
Background
Genome editing is a powerful molecular tool that allows for permanent genomic alteration. Currently the most widely used system is the Streptococcus pyogenes clustered regularly interspaced short palindromic repeats (CRISPR) / Cas9 endonuclease [1, 2]. Cas9 is an RNA-guided DNA endonuclease that has strong helicase activity, allowing for the opening and scanning of genomic DNA for a matching sequence. The target DNA is determined by an RNA component called a guide RNA. If a complementary DNA sequence is found along with an adjacent motif, called a protospacer adjacent motif (PAM), wildtype Cas9 introduces a double-strand break. For Cas9, the guide sequence is 20 basepairs and the PAM site is NGG, which means that a target locus requires the sequence N20-NGG. These properties mean that Cas9 can be used to knockout a target genomic locus. Double-strand breaks that are repaired by non-homologous end joining (NHEJ) result in a deletion.
Similar techniques can be used to knock-in specific genetic variants. If a DNA oligonucleotide containing the genetic variant of interest is supplied along with the Cas9 and guide RNA, the double-strand breaks are sometimes repaired by non-allelic homologous recombination (NAHR) using the oligo as a template, thereby introducing the variant of interest permanently into the target locus [3]. This method in particular has been touted as a potential mechanism to cure Mendelian-like genetic disorders by repairing the causative allele. A major concern for human gene-editing, however, is the possibility of introducing new genetic lesions through off-target activity of the editing enzyme. This has led to a general consensus that germline correction of a disease-causing variant in humans via Cas9 knock-in is not advisable at this time [4], and triggered intense research into ways to reduce off-target editing for clinical use.
One mechanism of reducing off-target effects would be to use an endonuclease with a longer PAM sequence. One recently identified RNA-guided genome editing endonuclease from Deltaproteobacteria is CasX (DpbCasX), tentatively designated as Cas12e [5, 6]. This endonuclease has a 4 basepair PAM site (TTCN) with a 20 basepair guide sequence. It introduces a staggered double-strand break by cutting downstream of the match on the strand complementary to the guide RNA and within the match on the opposite strand. The longer PAM site and smaller overall peptide size compared to Cas9 make CasX an attractive area of future genome editing development.
In this paper I present a catalog of CasX-compatible genome editing sites in 7 model organism genomes and in multiple mouse strains. The annotations are freely available from FigShare. Each site is annotated for chromosome, start and end position, PAM site, guide RNA target, uniqueness among editing sites, and any overlaps with the exons of known genes.
Construction and content
Identification of CasX sites
I coordinated part of the analysis using GNU Make (v4.1) on server running Ubuntu Linux (v16.04). The Makefile downloaded the gene annotations (GTF format) and genome sequences (FASTA format) for each of the specified genomes from release 95 of Ensembl [7]. The primary assembly FASTA file was used when it was available, and if none was found it fell back on using the top level file. I used the soft masked version (simple repeats as lower case characters rather than Ns) of the genome in either case. I calculated the GC content of each genome using a Python script with the pyfaidx package (v0.5.5.2) [8]. I then identified the potential CasX editing sites in each genome using Motif Scraper (v1.0.2) with the motif TTCNNNNNNNNNNNNNNNNNNNNN and multiple cores in file buffered mode [9]. The standard Motif Scraper options buffer all sites in memory for sorting later based on the order of contigs in the input FASTA file. For large genomes this uses a considerable amount of RAM. In file buffered mode the hits are printed by contig and strand into temporary files that are concatenated at the end of the analysis to form the full output, substantially reducing the overall memory burden.
Site annotation
I performed the annotations on the Washington University Center for High Performance Computing cluster. The motif location output included the location of the hit in the genome (contig, start, end) and the associated sequences. I wanted to further analyze each output for uniqueness of the target site, PAM site usage, and overlap with the exons of known genes using R (v3.5.1) with the GenomicRanges, GenomicFeatures, here, knitr, multidplyr, and tidyverse packages [10]. I calculated the size of the reference genomes from their FASTA index files, determined the uniqueness of guide RNA sites amongst all editing sites, counted PAM usage, and annotated any overlaps with the exons of known genes. It is worth noting that uniqueness of a guide was determined only among editing sites. Presumably another site that matches the guide exactly, but does not have the PAM site would not be cleaved. A guide that overlaps an exon can likely be used to knockout the gene or knock-in coding variants at the exon. Identifying unique editing sites that overlap promoters would require additional analysis. For the mouse strains, I calculated the presence (1) / absence (0) of guide sequences in each genome. I used this binary table to calculate the Hamming distance [11] between all strains in Python using pandas and the scipy spatial packages.
Utility and discussion
CasX-compatible unique editing targets are common in model organism genomes
I was able to catalog and annotate potential CasX editing sites in 7 common model species: yeast (Saccharomyces cerevisiae), flatworms (Caenorhabditis elegans), fruit flies (Drosophila melanogaster), zebrafish (Danio rerio), mouse (Mus musculus), rats (Rattus norvegicus), and humans (Homo sapiens). I identified 263,193,973 total sites, of which 200,558,172 were unique cutters in their respective genomes (Table 1). Across the seven genomes there was an average of 1 site per 37 basepairs, and 1 unique site every 45 basepairs. The exon overlaps support the potential use of CasX to target genes of interest for editing. The median number of exon overlapping cutters per gene ranged from 7 to 52 across the genomes tested, with between 6 and 45 unique cutters per gene (Table 2). Importantly, at least 90% of annotated genes across all organisms had at least one unique CasX site overlapping at least one exon. There were more A/T PAM sites (TTCA, TTCT) compared to C/G (TTCC, TTCG) PAM sites in all the surveyed genomes (Fig. 1). The TTCG PAM site is particularly depleted in zebrafish, mouse, rat, and human genomes, perhaps due to the general depletion of CpG sites genome-wide (Additional file 1: Table S1).
Table 1.
CasX site genomic distribution in 7 model organisms
| Organism | Total sites | Unique sites | Unique (%) | Total sites / Mbp [median] | Unique sites / Mbp [median] |
|---|---|---|---|---|---|
| S. cerevisiae | 367,810 | 345,376 | 93.90 | 30,254.74 | 28,409.4 |
| C. elegans | 3,315,259 | 2,988,557 | 90.15 | 33,057.91 | 29,800.2 |
| D. melanogaster | 3,681,483 | 3,011,422 | 81.80 | 25,614.59 | 20,952.5 |
| D. rerio | 33,296,705 | 22,500,532 | 67.58 | 24,242.74 | 16,382.2 |
| M. musculus | 70,817,235 | 54,464,834 | 76.91 | 25,932.10 | 19,944.1 |
| R. norvegicus | 73,427,248 | 55,157,865 | 75.12 | 25,582.77 | 19,217.5 |
| H. sapiens | 78,288,233 | 62,089,586 | 79.31 | 25,256.30 | 20,030.5 |
Table 2.
CasX sites overlapping known gene exons
| Organisms | Genes | Cut (%) | Unique cut (%) | Sites / gene [median] | Unique sites / gene [median] |
|---|---|---|---|---|---|
| S. cerevisiae | 7036 | 99.97 | 96.93 | 31 | 29 |
| C. elegans | 46,778 | 96.46 | 94.73 | 7 | 6 |
| D. melanogaster | 17,737 | 99.86 | 97.30 | 38 | 37 |
| D. rerio | 32,520 | 99.94 | 95.43 | 52 | 45 |
| M. musculus | 54,838 | 99.58 | 93.14 | 36 | 26 |
| R. norvegicus | 32,883 | 99.40 | 90.44 | 33 | 25 |
| H. sapiens | 58,735 | 99.60 | 96.98 | 28 | 22 |
Fig. 1.
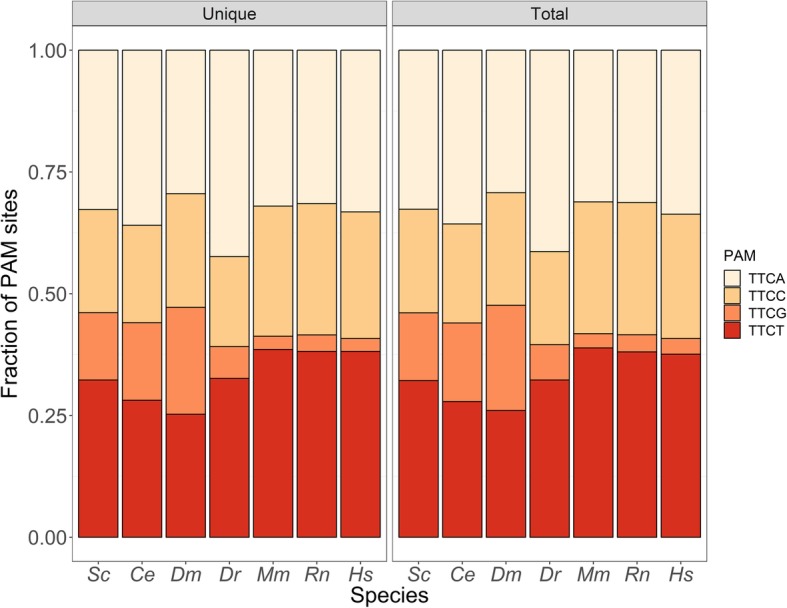
CasX PAM site usage. Shown in this figure are the 7 species on the x-axis (abbreviated as the first letter of the genus and species), and a stacked bar chart of fractional PAM site usage on the y-axis. The plot is divided into two subplots with the distribution of only unique cutters and of all sites. The A and T PAM sites are generally the most used. The TTCC and TTCG sites are used much less often in zebrafish, mouse, rat, and humans. The TTCG site, which contains a CpG dinucleotide, is seldom observed in those four species in particular
Mouse strains vary in guide RNA site availability in their reference genomes
I also annotated the CasX sites in the main Ensembl mouse reference (GRCm38; Mus musculus) and in multiple strains: 129S1/SvImJ, A/J, AKR/J, BALB/cJ, C3H/HeJ, C57BL/6NJ, CAST/EiJ, CBA/J, DBA/2 J, FVB/NJ, LP/J, NOD/ShiLtJ, NZO/HlLtJ, PWK/PhJ, and WSB/EiJ. The GRCm38 reference is built primarily from the C57BL/6 J strain. Genome-editing is especially important in the generation of mouse models, drastically reduced the time and effort required to generate knockouts and knock-ins. However, databases of editing sites for mice are built mostly from the GRCm38 reference, and therefore most applicable to C57BL/6 J.
I used a binary table of presence / absence of guide RNA sequences across all 16 genomes to calculate the Hamming distance between all pair-wise strain comparisons (Fig. 2). One important finding is that there is different site availability in the strains. The GRCm38 reference and C57BL/6NJ had few differences (distance 0.017), as expected. The site availability was also similar between 129S1/SvImJ and LP/J (distance 0.038). Two strains stood out as most different from the C57BL/6 J reference, CAST/EiJ (distance 0.297) and PWK/PhJ (distance 0.291), and from each other (distance 0.289). These strains in particular might benefit from strain-specific guide RNA development.
Fig. 2.

Hamming distance for editing targets amongst Ensembl mouse strains. Shown is the Hamming distance between sites for the GRCm38 reference (denoted Mus musculus), and genomes of 15 strains available via Ensembl. The matrix was ordered by calculating the hierarchical clustering of the distances with complete linkage. The strains CAST/EiJ and PWK/PhJ were the furthest from the GRCm38 reference among the tested strains, though all had some degree of difference. Those two strains in particular might require site annotations for their specific genomes rather than site selection from the general mouse reference
Conclusions
Identifying new RNA-guided endonucleases to use as genome editors is an area of intense research. There are currently many modifications of Cas9 that can help to decrease the number of off-target cuts (such as using dual Cas9 nickases), but it is still worth it to explore other editors with more favorable characteristics for clinical use. CasX appears to use a mechanism distinct from both Cas9 and Cas12a, suggesting it may have different benefits and limitations [6]. CasX guide sites are relatively common in all the tested genomes, and most genes have at least one CasX site overlapping an exon. This supports the potential utility of CasX in genome editing. The expanded PAM site also may reduce the number of off-target near matches in candidate genomes. Amongst the mouse strains, there were some substantial differences in site availability. In some strains, particularly CAST/EiJ and PWK/PhJ, there are many differences in site availability between them and the GRCm38 reference. It is important to note that the resolution of these differences is directly dependent on the quality of genome assembly. Any strains with poor assembly may have dropout of sites that is technical rather than biological. Regardless, this catalog of CasX editing sites will be an important resource in the future testing of this new class of RNA-guided genome editor.
Additional file
Table S1. GC PAM sites depleted in reference genomes (DOCX 15 kb)
Acknowledgements
Not applicable.
Abbreviations
- CRISPR
clustered regularly interspaced short palindromic repeats
- DpbCasX or CasX
Deltaproteobacteria CasX
- GTF
Gene Transfer Format
- NAHR
non-allelic homologous recombination
- NHEJ
non-homologous end joining
- PAM
protospacer adjacent motif
Author’s contribution
EDOR designed the study, analyzed the data, and wrote the manuscript. The author read and approved the final manuscript.
Funding
E.D.O.R. was partially supported by NIH grant P30-AR073752. The funder had no role in the design, analysis, interpretation of data, or writing of the manuscript. Some of the computations in this paper were performed using the facilities of the Washington University Center for High Performance Computing, which were partially provided through NIH grant S10 OD018091.
Availability of data and materials
The code to generate this analysis is available on GitHub:
https://github.com/RobersonLab/2019CasXModelOrgCatalog
The cataloged CasX sites are available on FigShare collected as a project:
https://figshare.com/projects/2019_CasX_genome_editing_site_annotations/61103
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 2012;337(6096):816–821. doi: 10.1126/science.1225829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA, et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 2013;339(6121):819–823. doi: 10.1126/science.1231143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang H, Yang H, Shivalila CS, Dawlaty MM, Cheng AW, Zhang F, Jaenisch R. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell. 2013;153(4):910–918. doi: 10.1016/j.cell.2013.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ormond KE, Mortlock DP, Scholes DT, Bombard Y, Brody LC, Faucett WA, Garrison NA, Hercher L, Isasi R, Middleton A, et al. Human germline genome editing. Am J Hum Genet. 2017;101(2):167–176. doi: 10.1016/j.ajhg.2017.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Koonin EV, Makarova KS, Zhang F. Diversity, classification and evolution of CRISPR-Cas systems. Curr Opin Microbiol. 2017;37:67–78. doi: 10.1016/j.mib.2017.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Liu JJ, Orlova N, Oakes BL, Ma E, Spinner HB, Baney KLM, Chuck J, Tan D, Knott GJ, Harrington LB, et al. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nature. 2019;566(7743):218–223. doi: 10.1038/s41586-019-0908-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Flicek P, Amode MR, Barrell D, Beal K, Brent S, Chen Y, Clapham P, Coates G, Fairley S, Fitzgerald S, et al. Ensembl 2011. Nucleic Acids Res. 2011;39(Database issue):D800–D806. doi: 10.1093/nar/gkq1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shirley MD, Ma Z, Pedersen BS, Wheelan SJ. Efficient “pythonic” access to FASTA files using pyfaidx. PeerJ PrePrints. 2015;3:e970v971. [Google Scholar]
- 9.Roberson EDO. Motif scraper: a cross-platform, open-source tool for identifying degenerate nucleotide motif matches in FASTA files. Bioinformatics. 2018;34(22):3926–3928. doi: 10.1093/bioinformatics/bty437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lawrence M, Huber W, Pages H, Aboyoun P, Carlson M, Gentleman R, Morgan MT, Carey VJ. Software for computing and annotating genomic ranges. PLoS Comput Biol. 2013;9(8):e1003118. doi: 10.1371/journal.pcbi.1003118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hamming RW. Error detecting and error correcting codes. Bell Syst Tech J. 1950;29(2):147–160. doi: 10.1002/j.1538-7305.1950.tb00463.x. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. GC PAM sites depleted in reference genomes (DOCX 15 kb)
Data Availability Statement
The code to generate this analysis is available on GitHub:
https://github.com/RobersonLab/2019CasXModelOrgCatalog
The cataloged CasX sites are available on FigShare collected as a project:
https://figshare.com/projects/2019_CasX_genome_editing_site_annotations/61103