

Abstract
Dopamine is central to reinforcement processing and exerts this function in species ranging from humans to fruit flies. It can do so via two different types of receptors (i.e., D1 or D2) that mediate either augmentation or abatement of cellular cAMP levels. Whereas D1 receptors are known to contribute to Drosophila aversive odor learning per se, we here show that D2 receptors are specific for support of a consolidated form of odor memory known as anesthesia-resistant memory. By means of genetic mosaicism, we localize this function to Kenyon cells, the mushroom body intrinsic neurons, as well as GABAergic APL neurons and local interneurons of the antennal lobes, suggesting that consolidated anesthesia-resistant memory requires widespread dopaminergic modulation within the olfactory circuit. Additionally, dopaminergic neurons themselves require D2R, suggesting a critical role in dopamine release via its recognized autoreceptor function. Considering the dual role of dopamine in balancing memory acquisition (proactive function of dopamine) and its “forgetting” (retroactive function of dopamine), our analysis suggests D2R as central player of either process.
SIGNIFICANCE STATEMENT Dopamine provides different information; while it mediates reinforcement during the learning act (proactive function), it balances memory performance between two antithetic processes thereafter (retroactive function) (i.e., forgetting and augmentation). Such bidirectional design can also be found at level of dopamine receptors, where augmenting D1 and abating D2 receptors are engaged to balance cellular cAMP levels. Here, we report that consolidated anesthesia-resistant memory (ARM), but not other concomitant memory phases, are sensitive to bidirectional dopaminergic signals. By means of genetic mosaicism, we identified widespread dopaminergic modulation within the olfactory circuit that suggests nonredundant and reiterating functions of D2R in support of ARM. Our results oppose ARM to its concomitant memory phases that localize to mushroom bodies and propose a decentralized organization of consolidated ARM.
Keywords: ARM, cAMP, D2 receptor, dopamine, Drosophila, odor learning
Introduction
The dopaminergic system mediates reinforcement processing and exhibits similar functions in learning and memory formation from fruit flies to humans (Schultz, 2004; Berridge and Kringelbach, 2015; Nutt et al., 2015; Schultz, 2015; West et al., 2015). During the last decades, Drosophila has evolved as a model to unravel the anatomical design of neuronal circuits dedicated to reinforcement processing, and particular clusters of dopaminergic neurons (DANs) within the fly brain have been identified that contribute to aversive odor learning (Schwaerzel et al., 2003; Aso et al., 2010, 2012). DANs from the fly's PPL1, PPL2ab, and PAM clusters are of particular importance, and behavioral reinforcement can be substituted by experimental activation of DANs, resulting in formation of associative odor memories (Schroll et al., 2006; Claridge-Chang et al., 2009; Aso et al., 2010, 2012; Yamagata et al., 2015). These DANs project to the mushroom bodies (MBs) where they synapse onto Kenyon cells (KCs), the intrinsic neurons of the MBs, as well as onto MB output neurons that provide feedforward and feedback connections between KCs and DANs of distinct MB zones (Séjourné et al., 2011; Aso et al., 2014). These distinct zones appear to be critical for function of the KC-DANs-MB output neurons network because learning-induced plasticity occurs similarly in zonal patterns (Boto et al., 2014).
The KC-DANs-MB output neurons network exhibits complex signaling, and DANs mediate different information at different time points: Although DANs provide US-related impulses during the acquisition phase (Schwaerzel et al., 2003; Schroll et al., 2006), they impede on performance in a bidirectional way thereafter, either by abolishing performance via increased DAN activity, or by augmenting it through reduced DAN signaling (Berry et al., 2012, 2015). It thus appears that memory formation is not tuned to achieve maximal performance but rather actively balanced between two antithetic processes (i.e., “forgetting” and “augmentation”) by adjusted DAN activity.
Such bidirectional design also applies to dopamine receptors (DARs) that provide the central interface for DANs to communicate with appropriate downstream neurons. DARs converge onto intracellular cAMP levels in an antithetic fashion; whereas D1-type receptors augment cAMP levels, D2-type receptors can abate it (Doya, 2008; Nakano et al., 2010; Nutt et al., 2015). Apparently, the DAR interface is, as a matter of principle, designed to balance cellular cAMP levels. However, so far, only D1 receptors have been reported to contribute to Drosophila odor learning (Kim et al., 2007; Blum et al., 2009; Qin et al., 2012; Boto et al., 2014). Here, we addressed the contribution of inhibitory D2 receptors and show that D2 is specific for supporting a particular form of consolidated memory known as anesthesia-resistant memory (ARM). By means of genetic mosaicism, we localized this function to Kenyon cells (KCs), the MB intrinsic neurons, as well as GABAergic APL neurons and local interneurons of the antennal lobes, suggesting that consolidated ARM requires widespread dopaminergic modulation within the olfactory circuit.
Materials and Methods
Fly care.
Flies were raised at 24°C and 60% relative humidity with a 14:10 h light-dark cycle on cornmeal-based food following the Würzburg recipe (Guo et al., 1996). Genetic crosses were performed according to standard procedures. All experiments were performed with 3- to 5-d-old male F1 progeny of homozygous parental lines. All lines used in this study were outcrossed to Canton-S.
Behavioral experiments.
Flies were transferred to fresh-food vials for up to 48 h before the test. Behavioral experiments were performed in dim red light at 80% relative humidity with 3-octanol (1/100 dilution in mineral oil presented in a cup of 14 mm diameter) and 4-methyl-cyclohexanol (1/150 dilution in mineral oil presented in a cup of 14 mm diameter) used as olfactory stimuli. The US was represented by a train of 12 DC pulses of 120 V administered in fast succession with 5 s intervals as originally introduced by Tully and Quinn (1985).
For measuring performance of short-term memory (STM), flies were tested immediately after the end of the training session (i.e., exactly 3 min after onset of training). Performance of mid-term memory (MTM) and ARM was determined at 3 h after training, and flies were transferred to neutral containers without food for the resting period. For separation of consolidated ARM and labile ASM, two groups of flies were separately trained, and one group was cooled in an ice-bath (0°C) for 120 s at 2.5 h after training and tested for odor memory after 30 min of recovery (cold+ group). This treatment erases the labile anesthesia-sensitive memory (ASM) component. Thus, performance of the cold+ group is solely due to ARM. ASM was calculated by subtracting the performance index of the cold+ group (which performed due to ARM) from the performance index of the cold− group that performed due to ASM and ARM (Scheunemann et al., 2012). Calculation of behavioral indices was done as originally introduced by Tully and Quinn (1985). In case of 5 min memory, we applied 90 s ice-bath cooling immediately after the end of training followed by 60 s recovery before testing. Five minute controls received 90 s of ice-bath cooling followed by 60 s recovery immediately before training.
Generating D2 deletion strains.
We used the Drosophila FRT-derived deletion system (Parks et al., 2004) to generate a loss-of-function allele for the Drosophila D2-like receptor (CG33517). FRT-dependent remobilization of the trans-heterozygous P-element combinations f02905/f06521 and f02905/f02891 generated deletions D2RΔ1 spanning 32,386 bp and D2RΔ1 spanning 46,005 bp, respectively (see Fig. 1A). Deletions were verified by PCR and subsequent sequencing.
Figure 1.

Deletion of a Drosophila D2 receptor deteriorates aversive odor learning. Null D2R mutants were generated by targeted deletion of P-elements and tested for associative odor learning using the Tully–Quinn paradigm (Tully and Quinn, 1985). A, Deletions at the Drosophila D2R locus were generated by remobilizing FRT-containing P-elements: D2RΔ1 was generated by combining f02905 and f06521; D2RΔ2 by combining f02905 and f02891. BSC581 is a characterized deletion partially removing D2R coding sequence. B, Performance of STM was reduced in D2R mutants, whereas shock response and odor acuity were unaffected. C, The D2R alleles D2RΔ1, D2RΔ2, and BSC581 complement for each other for STM performance. D, Pan-neuronal knockdown of D2R by means of RNAi (D2RKO) (Draper et al., 2007) deteriorated STM performance. E, Pan-neuronal rescue of null D2R mutants by means of D2R+ cDNA restored STM performance. F, Conditional rescue at level of MBs, a recognized Drosophila learning center, restored STM under permissive conditions, whereas restricting expression of rescuing cDNA did not. Error bars indicate mean ± SEM of 8–10 biological repetitions (i.e., N = 8–10). Statistical differences at level of p ≤ 0.05 are denoted by different letters or asterisks.
Generating transgenic strains.
To construct the upstream activating sequence (UAS) expression vectors containing wild-type D2 cDNA, we obtained the cDNA clone RE06088 containing a D2 isoform (Hearn et al., 2002) from the Drosophila Genomic Resource Center. cDNA was amplified by PCR, subcloned into the pEntry vector (pENTR/DTOPO Cloning Kit, Invitrogen), and further cloned into the pUAST Drosophila transfection vector obtained from Drosophila Genomic Resource Center. Germ-line transformation was performed by BestGene.
Compendium of transgenic lines.
The following fly strains were used in this study: wild-type Canton-S; D2RΔ1 (current study), D2RΔ2 (current study), Df(1)BSC587 (Cook et al., 2012), dnc1 (Davis and Kiger, 1981), and rut1 (Livingstone et al., 1984). We used the following Gal4 lines: orco (Hummel et al., 2003), GH298 (Stocker et al., 1997), GH148 (Heimbeck et al., 2001), NP2426 (Das et al., 2008), NP1227 (Chou et al., 2010), NP2631 (Pitman et al., 2011), mb247 (Zars et al., 2000), 17d (Scheunemann et al., 2012), NP1131 (Akalal et al., 2006), c320 and c305a (Krashes et al., 2007), and TH (Schwaerzel et al., 2003). We used the following UAS lines: UAS-D2R+ (current study), UAS-D2R-RNAi (Draper et al., 2007), UAS-dnc-RNAi (Cheung et al., 1999; Scheunemann et al., 2012), and UAS-rut-RNAi (Pan et al., 2009). To modify spatial and temporal expression of particular Gal4 lines, we used tub-Gal80ts (McGuire et al., 2003) and Cha-Gal80 (Kitamoto, 2002).
Results
Engineering a null D2 receptor mutant by targeted P-element remobilization
To address the role of D2 receptors in odor learning, we targeted a small deletion to the Drosophila D2R locus by remobilizing FRT-containing P-elements (Parks et al., 2004). Thereby, we generated two deletions (D2RΔ1 and D2RΔ2, respectively) that partially covered the D2R locus (Fig. 1A). Putative null D2R mutants were homozygous viable and fertile but suffered from reduced performance of 3 min odor memory (one-way ANOVA: F(2,41) = 12.61, p < 0.001). Shock avoidance response (one-way ANOVA: F(2,39) = 0.03, p = 0.97) and olfactory responsiveness (one-way ANOVA: F(5,76) = 0.13, p = 0.98) were unaffected, thus excluding perceptual deficits as a potential cause of the behavioral defect (Fig. 1B). Because the mutations also removed CG17003, a gene of unknown function, we tested performance of D2R mutants in trans to BSC587, a putative D2 allele, because the deficiency covered D2R, but it did not affect CG17003 (Fig. 1C). D2Δ deletions in trans to BSC exhibited reduced performance indistinguishable from homozygote D2Δs, as did the trans combination of D2RΔ1 and D2RΔ2, whereas animals that carried appropriate deletions in heterozygous condition performed like wild-type animals (one-way ANOVA: F(7,102) = 10.80, p < 0.001). By means of this complementation analysis, we concluded that the memory phenotype of D2R mutants was recessive and that D2RΔ1, D2RΔ2, and BSC587 were allelic to each other.
Next, we used RNAi constructs that effectively target D2R transcripts to degradation (Draper et al., 2007). Thereby, we reduced memory performance to levels indistinguishable from that of D2R mutants when RNAi was expressed under control of elav-Gal4 (Fig. 1D). Genetic controls that carried either transgene in separation performed at wild-type levels (one-way ANOVA: F(5,96) = 6.66, p < 0.001). In a second test, we performed a genetic rescue experiment focusing on D2RΔ1 (Fig. 1E). Expressing wild-type D2R+ cDNA under control of the pan-neuronal elav Gal4 driver within an otherwise D2R mutant animal restored memory performance to wild-type levels, whereas genetic controls that carried either transgene alone performed at mutant levels (one-way ANOVA: F(4,55) = 10.46, p < 0.001). Finally, we aimed to distinguish between acute functions of D2R at the adult stage or, alternatively, during development by use of the TARGET system (McGuire et al., 2003). Toward that goal, we induced expression of the rescuing transgene within KCs of the MBs, a recognized Drosophila “learning center” (Heisenberg, 2003) by use of mb247-Gal4 (Fig. 1F). When we induced expression at the adult stage by shifting animals from 18°C to 30°C (permissive conditions), we obtained full rescue (Fig. 1F; one-way ANOVA: F(2,36) = 7.52, p < 0.01), whereas animals performed on mutant level under restrictive conditions (Fig. 1F; one-way ANOVA: F(2,38) = 10.52, p < 0.01). Together, these results established inhibitory D2R as critical regulator of aversive odor learning and identified MB KCs as a neuronal site of function.
D2 mutants lack consolidated ARM
Memory is composed of distinct memory phases that support behavioral performance during distinct time windows that separate at biochemical and neuronal levels (Margulies et al., 2005; Schwärzel and Müller, 2006). In case of Drosophila aversive olfactory learning, multiple memory phases are induced after single cycle training that separate by their resistance to amnestic cooling, and hence are referred to as ARM and ASM. Typically, this separation is applied to 3 h memory by use of 2 min cold treatment (Tempel et al., 1983; Scheunemann et al., 2012). When we separated 3 h total memory into its consolidated ARM and labile ASM components (Fig. 2A), we observed that ARM was abolished in D2R mutants (t test, t(25) = 4.49, p < 0.01), whereas ASM remained unaffected (t test, t(28) = 0.58, p = 0.57). How did total loss of ARM relate to a reduction of 3 min memory because D2R is a null mutant?
Figure 2.

D2R mutants lack consolidated ARM. Consolidated ARM and labile ASM are proportionate components of aversive odor memory and can be separately quantified by use of cold-amnestic treatment. A, Applying cold amnestic treatment to 3 h memory separated consolidated ARM and labile ASM from total memory. Whereas ARM was abolished in null D2R mutants, performance of ASM remained unaffected. B, Administering amnestic treatment directly after training revealed the existence of consolidated memory component that were disrupted in D2R mutants. Error bars indicate mean ± SEM of 8–10 biological repetitions (i.e., N = 8–10). **Statistical differences (p ≤ 0.05). ns, Not significant.
Similar to its 3 h counterpart, early memory is composed of a consolidated and a nonconsolidated component that separate by resistance to amnestic cooling (Knapek et al., 2011; Bouzaiane et al., 2015). When we separate early (5 min) total memory into its consolidated and nonconsolidated forms (Fig. 2B), we revealed loss of early ARM in D2R mutants, t(18) = 4.44, p < 0.01. We thus conclude that consolidated ARM has a general requirement for inhibitory D2R. Given the well-established function of D2 receptors as inhibitor of the cAMP signaling pathway (Hearn et al., 2002; Bonci and Hopf, 2005; Pignatelli and Bonci, 2015), we next sought to investigate how Drosophila D2R interacts with learning mutants that affect cAMP signaling.
D2R interacts with a rut-independent cAMP signaling pathway
The cAMP-signaling pathway is central to Drosophila odor learning, and mutants affecting either the Type 1 adenylyl cyclase rutabaga (rut) or the cAMP-specific Type 4 phosphodiesterase dunce (dnc) suffer from reduced odor memory performance (Byers et al., 1981; Livingstone et al., 1984). However, rut and dnc mutants dissociate between ARM and ASM, showing that functionally distinct cAMP signals are engaged during odor learning that act within different neuronal circuits to support ARM or ASM, respectively (Scheunemann et al., 2012, 2013). Because D2R comprises a major route for dopamine to inhibit cAMP signaling (Hearn et al., 2002), we tested for genetic interactions between D2R and rut- or dnc-mutants, respectively. Toward that goal, we separated 3 h total memory into ARM and ASM by use of cold amnestic treatment (Fig. 3A) and established RNAi-mediated knockdown of rut or dnc transcripts to circumvent recombination because all three mutants were X-linked. Pan-neuronal expression of knockdown constructs targeting either rut- or dnc-transcripts (Cheung et al., 2006; Pan et al., 2009) affected appropriate memory components in a manner indistinguishable from the respective mutant (i.e., knockdown of dnc resulted in loss of consolidated ARM) (Fig. 3B; one-way ANOVA: F(9,105) = 15.53, p < 0.001), whereas rut abolished ASM (Fig. 3C; one-way ANOVA: F(9,105) = 10.54, p < 0.001). When knockdowns were executed in D2R null mutant background, we observed interaction between D2R and rut as indicated by an additional loss of ARM (one-way ANOVA: F(9, 1054) = 9.83, p < 0.001), whereas D2R and dnc-PDE4 did not interact as deduced by wild-type levels of ASM (Fig. 3D). Genetic controls that carried either transgene alone did not impair memory performance (Fig. 3E). Together, we conclude that D2R acts independent of the rut adenylyl cyclase but shares a common neuronal site with the former as revealed by our KC rescue experiment (Fig. 1E).
Figure 3.
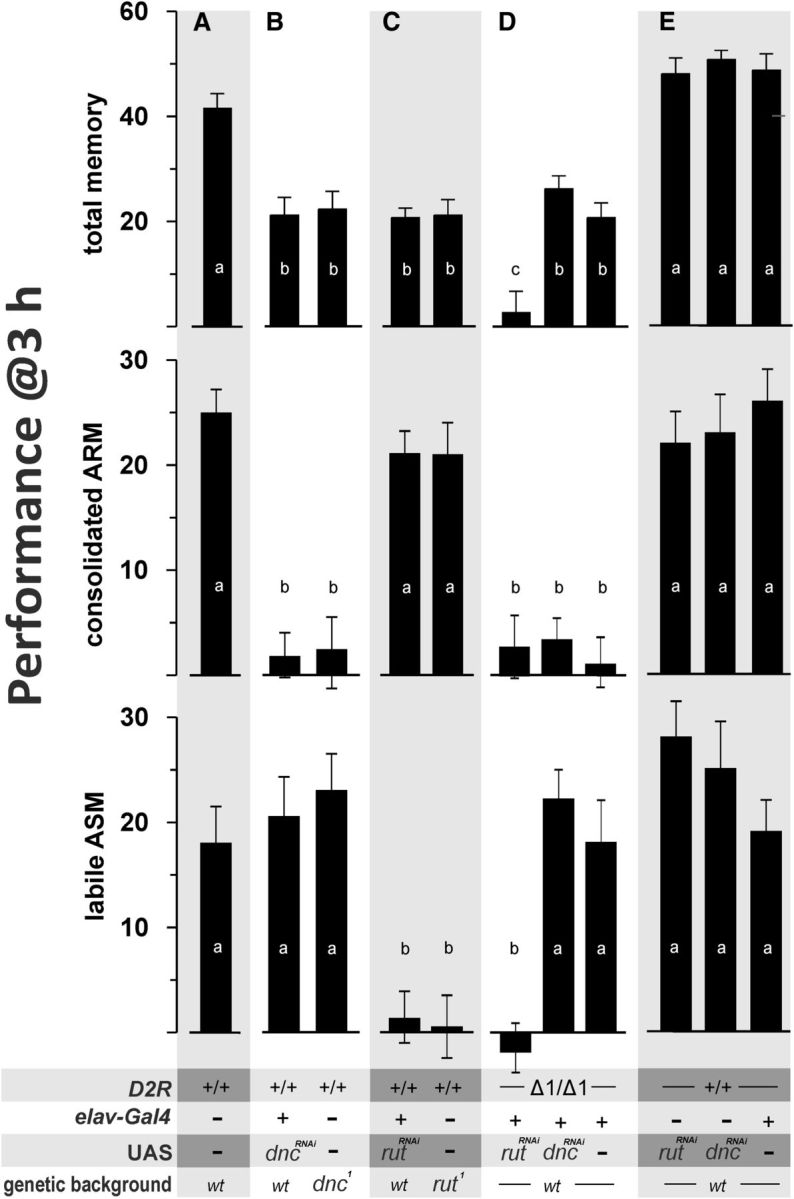
D2R acts in parallel to rut-dependent cAMP signals. A, Three hour total memory was separated into consolidated ARM and labile ASM to test for genetic interactions between D2R, dnc-PDE4, and rut-AC1. B, C, Targeted knockdown of either dnc-PDE4 or rut-AC1 dissociated between ARM and ASM in a manner indistinguishable from appropriate mutants. D, Knockdown of rut-AC1, but not dnc-PDE4, resulted in loss of all memory phases, suggesting parallel processing of appropriate cAMP signals. E, Genetic controls performed on wild-type levels. Error bars indicate mean ± SEM of 6–8 biological repetitions (i.e., N = 6–8). Statistical differences at level of p ≤ 0.05 are denoted by different letters.
RNAi-mediated knockdown identified a large circuit modified by D2R
A previous study has identified KCs as necessary neuronal site of D2R function in support of larval olfactory learning by means of RNAi knockdown (Qi and Lee, 2014). While this result is well in line with our KC rescue and suggestive for a common architecture of D2R in larval and adult fruit flies, the authors also reported requirement at level of larval DANs where D2R is hypothesized to act as autoreceptor in regulation of dopamine release. We analyzed the adult counterpart by targeting D2R knockdown to DANs. When assayed for 3 h memory, TH//D2R-RNAi animals showed reduced performance of total memory (Fig. 4A,B, top; t test: t(13) = 8.87, p < 0.001) due to loss of consolidated ARM (Fig. 4A,B, middle panel; t test: t(13) = 5.03, p < 0.001) with no change in labile ASM (Fig. 4A,B, bottom panel). These results suggest a common functional architecture of D2R in adult and larval fruit flies.
Figure 4.

ARM requires ample distribution of D2R. Targeting knockdown of D2R to individual stages of the olfactory pathway defined necessary elements of D2R function. Performance of total memory (top panels), consolidated ARM (middle panels), and calculated ASM (bottom panels) was scored (A). Particular Gal4 lines were used for knockdown at individual stage of the circuitry, including dopaminergic neurons (B), olfactory receptor neurons (C), antennal lobe interneurons (D–F), projections neurons and/or APL neurons (G–I), and mushroom body KCs (K–O). Error bars indicate mean ± SEM of 8–10 biological repetitions (i.e., N = 8–10). **Statistical differences (p ≤ 0.05). ns, Not significant.
To test for further functions of D2R, we performed knockdown aided by Gal4 lines systematically addressing individual stage of the learning relevant circuitry. When D2R knockdown was applied to olfactory receptor neurons by use of OR83bGal4, we observed no effect on performance (Fig. 4C; t test: t(15) = 0.40, p = 0.70 for MTM and t(15) = 0.54, p = 0.59 for ARM). In contrast, memory was reduced and ARM abolished when RNAi was expressed under control of NP1227Gal4 (Fig. 4D; t test: t(14) = 5.18, p < 0.001 for MTM and t(15) = 6.32, p < 0.001 for ARM), NP2426Gal4 (Fig. 4E; t test: t(13) = 4.09, p < 0.01 for MTM and t(13) = 4.18, p < 0.01 for ARM), or GH298Gal4 (Fig. 4F; t test: t(15) = 7.51, p < 0.001 for MTM and t(15) = 6.46, p < 0.001 for ARM) (i.e., Gal4 lines that mark GABAergic antennal lobe interneurons) (Sachse et al., 2007; Tanaka et al., 2009; Chou et al., 2010). A similar effect was observed when knockdown was applied under control of GH146Gal4 (Fig. 4G; t test: t(15) = 6.40, p < 0.001 for MTM and t(15) = 4.37, p < 0.001 for ARM) that marked the majority of olfactory projection neurons as well as the GABAergic APL neurons (Pitman et al., 2011). To dissociate between requirements of either neuronal population, we used additional elements that either removed APL neurons from the GH146 pattern (GH146Gal4, ChaGal80, Fig. 4H; t test: t(14) = 0.61, p = 0.55 for MTM and t(14) = 0.53, p = 0.60 for ARM) or addressed APLs, but not olfactory projection neurons, by use of an alternative driver (i.e., NP2631Gal4) (Fig. 4I; t test: t(14) = 11.03, p < 0.001 for MTM and t(14) = 10.57, p < 0.001 for ARM). Our results clearly identified APL rather than projection neurons as site of D2R function. Finally, we addressed KCs by use of mb247Gal4 resulting in memory decline and ARM abolishment (Fig. 4K; t test: t(16) = 3.51, p < 0.01 for MTM and t(17) = 2.96, p < 0.01 for ARM). To further investigate on particular lobe systems, we used more restricted Gal4 lines and identified KCs of the αβ-lobes marked in 17dGal4 (Fig. 4L; t test: t(15) = 4.91, p < 0.001 for MTM and t(15) = 4.25, p < 0.001 for ARM) as well as γ-lobes marked in NP1131Gal4 (Fig. 4M; t test: t(18) = 4.78, p < 0.001 for MTM and t(18) = 4.49, p < 0.001 for ARM) but not prime lobe KCs marked in c320Gal4(Fig. 4N; t test, t(13) = 1.13, p = 0.28 for MTM and t(13) = 1.66, p = 0.12 for ARM) or c305aGal4 (Fig. 4O; t test, t(14) = 1.00, p = 0.34 for MTM and t(14) = 1.43, p = 0.17 for ARM). Together, this functional analysis revealed a broad distribution of D2R in support of ARM, including particular KC populations, GABAergic APL and antennal lobe interneurons, as well as dopaminergic neurons themselves. While this analysis has revealed necessary sites of D2R function, we next sought to identify those stages of the circuitry where D2R functions suffices ARM support.
Genetic rescue within the MBs suffices for ARM performance
Toward that goal, we expressed wild-type DR2+ cDNA within an otherwise D2R mutant brain and determined total memory (Fig. 5, top panels), consolidated ARM (Fig. 5, middle panels), or calculated labile ASM (Fig. 5, bottom panels) at 3 h after conditioning. First, we performed rescue within olfactory receptor neurons (OR83bGal4; Fig. 5B) but failed to rescue performance (t test, t(11) = 0.67, p = 0.54 for MTM and t(11) = 0.08, p = 0.93 for ARM). Similarly, local interneurons of the antennal lobes (GH298Gal4, Fig. 5C; t(10) = 0.13, p = 0.90 for MTM and t(11) = 1.24, p = 0.24 for ARM), APLs and/or olfactory projection neurons (GH146Gal4, Fig. 5D; t test, t(14) = 0.02, p = 0.94 for MTM and t(15) = 0.90, p = 0.37 for ARM), as well as dopaminergic neurons (THGal4, Fig. 5E; t test, t(12) = 0.07, p = 0.94 for MTM and t(12) = 0.51, p = 0.62 for ARM) failed to restore ARM. In contrast, expressing D2+ cDNA within KCs by use of mb247Gal4 fully restored ARM performance (Fig. 5F; t test, t(14) = 5.44, p < 0.001 for MTM and t(14) = 1.43, p < 0.001 for ARM), as did Gal4 lines that restricted expression to either αβ-lobes (17dGal4, Fig. 5G; t test, t(13) = 5.53, p < 0.0.001 for MTM and t(11) = 6.69, p < 0.001 for ARM) or the ϒ-lobes (NP1131Gal4, Fig. 5H; t test, t(14) = 5.52, p = 0.001 for MTM and t(11) = 4.45, p < 0.001 for ARM). Expression at level of prime lobe KCs did not rescue (c320Gal4, Fig. 5I; t test, t(12) = 1.34, p = 0.21 for MTM and t(12) = 1.61, p = 0.14 for ARM). Performance of labile ASM was unaffected in all genotypes.
Figure 5.

KCs comprise a sufficient site of D2R in support of ARM. Genetic rescue of D2R function was performed at individual stages of the olfactory pathway to define sufficient elements of D2R function. Performance of total memory (top panels), consolidated ARM (middle panels), and calculated ASM (bottom panels) was scored (A). Gal4 lines expressing the rescuing cDNA at different levels of the circuitry were used that mark dopaminergic neurons (B), olfactory receptor neurons (C), antennal lobe interneurons (D), projections neurons and/or APL neurons (E), the majority of MB KCs (F), KCs of the α/β-lobes (G), the γ-lobes (H), and the α/β-lobes (I). Error bars indicate mean ± SEM of 8–10 biological repetitions (i.e., N = 8–10). **Statistical differences (p ≤ 0.05). ns, Not significant.
Discussion
Here we performed functional mapping of a Drosophila D2R that is required for support of consolidated ARM. Thereby, our results extend the repertoire of dopamine receptors assigned to aversive odor learning for D2R, suggesting that, for ARM, excitatory and inhibitory dopamine inputs are equally important.
Dopamine receptors are functionally dissociated but converge at level of MBs
Several DARs are involved in Drosophila aversive odor learning: dDA1 (Kim et al., 2003), DAMB (Han et al., 1996), and D2R (current study). All three receptors exert their function within KCs as demonstrated for dDA1 (Qin et al., 2012) and D2R mutants by means of genetic rescue, whereas DAMB, another D1 type receptor, was localized by in situ hybridization (Han et al., 1996) and antibody staining (Crittenden et al., 1998). Mutant analysis revealed nonredundant functions: loss of dDA1 abolished all forms of aversive odor learning, including ARM (Qin et al., 2012), whereas DAMB mutants showed nearly normal learning yet impaired forgetting that results in augmented performance and revealed a retroactive dopamine function (Berry et al., 2012). Vis-à-vis these excitatory receptors is D2R, an inhibitory receptor capable of abating cellular cAMP levels (Hearn et al., 2002). However, D2R mutants do not categorize with either of the D1 phenotypes but exhibit discrete and specific loss of consolidated ARM. Thus, DAR functions dissociate at the behavioral level directly asking how this is implemented at a functional level.
Do dopamine receptors equilibrate KC cAMP levels in a concurrent fashion?
DARs are G-protein-coupled receptors and D1s activate Type 1 adenylyl cyclases (AC1s) via Gs/Golf, whereas D2s couple to Gαi/o subunits that bind to and inhibit AC1s, thereby preventing production of cAMP and activation of PKA. In this view, a primary action of dopamine is to elevate cellular cAMP levels through AC1s, and this activation is balanced by its concurrent D2-dependent inhibition (for review, see Neve et al., 2004; Bonci and Hopf, 2005). Drosophila DARs adhere to this mode of action (Han et al., 1996; Hearn et al., 2002), so that D2 has potential to operate on proactive and/or retroactive dopamine functions by counterbalancing dDA1 and/or DAMB actions, respectively.
ARM is sensitive to proactive (Qin et al., 2012) and retroactive dopamine actions (Plaçais et al., 2012), rendering loss of ARM performance possible via either process. Our data indicate that proactive and retroactive aspects are differentially affected by knockdown or rescue and localize to different stages of the learning-relevant circuitry: In particular, no sign of retroactive D2R actions was observed in case of genetic rescue. Rather, we observed full restoration of its proactive counterpart upon expression of cDNA at level of αβ- or γ-type KCs. This suggests that proactive D2R action is redundant at level of particular MB lobes.
On the contrary, knockdown of D2R at level of either αβ- or γ-lobes disrupted ARM performance, despite similar constellations of the MB network compared with corresponding rescue (for a comparison of circuit components affected upon rescue or knockdown approaches, see Table 1). We attribute this contrast to different D2R functionality of MB-adjacent circuits that remained wild-type like within the knockdown approach but changed to null D2R mutant background in case of rescue. It thus appears that retroactive dopamine actions are doubly dependent on D2R abundance (I) within the MB network in conjunction with (II) MB adjacent circuits.
Table 1.
Comparison of proactive and retroactive D2R functionsa
| Gal4 line | Marks | Retroactive function revealed by knockdown |
Proactive function revealed by rescue |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Behavior | MBs |
Adjacent circuit |
Behavior | MBs |
Adjacent circuit |
||||||||||
| αβ | γ | p | APL | DAN | LNs | αβ | γ | p | APL | DAN | LNs | ||||
| NP1227 | LN1 | − | F | F | F | F | F | NF | — | — | — | — | — | — | |
| GH298 | LN1 | − | F | F | F | F | F | NF | − | NF | NF | NF | NF | NF | F |
| NP2426 | LN2 | − | F | F | F | F | F | NF | — | — | — | — | — | — | |
| TH | DANs | − | F | F | F | F | NF | F | − | NF | NF | NF | NF | F | NF |
| GH146 | PNs and APL | − | F | F | F | NF | F | F | − | NF | NF | NF | F | NF | NF |
| NP2631 | APL | − | F | F | F | NF | F | F | — | — | — | — | — | — | |
| mb247 | αβ and γ KCs | − | NF | NF | F | F | F | F | + | F | F | NF | NF | NF | NF |
| 17d | αβ KCs | − | NF | F | F | F | F | F | + | F | NF | NF | NF | NF | NF |
| NP1131 | γ KCs | − | F | NF | F | F | F | F | + | NF | F | NF | NF | NF | NF |
| OR83b | ORNs | + | F | F | F | F | F | F | − | NF | NF | NF | NF | NF | NF |
| GH146; ChaG80 | PNs | + | F | F | F | F | F | F | — | — | — | — | — | — | |
| c320 | + | F | F | NF (no subset)b | F | F | F | − | NF | NF | F | NF | NF | NF | |
| c305a | + | F | F | NF (no subset)b | F | F | F | — | — | — | — | — | — | ||
aGal4 lines addressing particular stages of the learning relevant circuitry have been used to drive RNAi-mediated knockdown or genetic rescue of a Drosophila D2 receptor. Performance of ARM (i.e. results from Figures 4 and 5) is schematized according to wild-type (+) or mutant like (−) behavior. D2R function at level of MBs and adjacent circuits is coded as follows: F, Functional; NF, nonfunctional; —, not applicable.
bGal4 lines that did not impact on ARM performance with knockdown.
D2R mediates retroactive dopamine actions via a complex neuronal circuitry
In addition to KCs, D2R is required at level of GABAergic APLs and antennal lobe local interneurons, two sets of inhibitory neurons assigned to balancing network activity (Tanaka et al., 2009; Parnas et al., 2013). GABAergic local interneurons are site of DUNCE action, another negative regulator of cAMP signaling (Scheunemann et al., 2012) that impacts on ARM. However, whatever dunce impacts on memory acquisition or modulation is unknown, in contrast to a recognized function of APLs in retroactive modulation (Wu et al., 2013). APLs are connected to KCs in a reciprocal fashion and provide recurrent inhibitory feedback from the MB lobes to the sensory input region (Liu et al., 2007).
In the biochemical domain, vertebrate D2Rs excerpts numerous functions via interaction with the Gβγ subunit that impact on multiple signaling pathways, including Ca2+ (Yan et al., 1997; Zamponi and Snutch, 1998; Koga and Momiyama, 2000), MAPK (Faure et al., 1994; Ghahremani et al., 2000), phospholipases (Mitchell et al., 1998; Yan et al., 1999), and β-arrestin (Urs et al., 2015). Moreover, D2R sensitizes Type 2/4 adenylyl cyclases (AC2/4; i.e., recognized coincidence detectors of PKC and Gαs signals) via Gβγ, thereby providing an alternative route for D2R to cAMP-PKA signaling independent of concurrently regulated AC1s (Watts and Neve, 2005). Thus, a wealth of signaling pathways is available to D2R regardless of concurrent D1 action. However, it will require future experiments to elucidate which options are engaged in case of Drosophila ARM and how they map onto the learning relevant circuitry.
Does D2R regulate dopamine release via auto-regulation?
D2R exerts additional support of ARM at level of DANs as revealed by RNAi knockdown. In vertebrate midbrain neurons, D2R acts as autoreceptor of dopamine release by decreasing the excitability of DANs and by curtailing dopamine abundance via control of tyrosine hydroxylase, the rate-limiting enzyme of dopamine synthesis, as well as via plasma membrane dopamine transporters (Beaulieu and Gainetdinov, 2011; Ford, 2014). Lack of a putative D2 autoreceptor in Drosophila DARs could lead to increased dopamine release that in turn has potential to unbalance proactive and/or retroactive processes. Actually, Drosophila DANs are both upstream of KCs during acquisition (Schroll et al., 2006; Aso et al., 2012), as well as downstream of KCs in case they impact on retroactive modulation (Plaçais et al., 2012), so that both scenarios appear equally plausible. Conditional block of DANs in combination with appropriate DAR manipulation may be applied within future experiments to fully understand the reiterating and nonredundant contributions of dopamine and its receptors in associative learning and memory consolidation.
Footnotes
This work was supported by Forschungsgemeinschaft Grant SI841/5-1 to M.S. and Deutsche Forschungsgemeinschaft research unit FOR 1363. Fly stocks were obtained from the Bloomington Stock Center and from Vienna Drosophila RNAi Centre (Vienna, Austria). We thank André Fiala for helpful comments on the manuscript.
The authors declare no competing financial interests.
References
- Akalal DB, Wilson CF, Zong L, Tanaka NK, Ito K, Davis RL. Roles for Drosophila mushroom body neurons in olfactory learning and memory. Learn Mem. 2006;13:659–668. doi: 10.1101/lm.221206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aso Y, Siwanowicz I, Bräcker L, Ito K, Kitamoto T, Tanimoto H. Specific dopaminergic neurons for the formation of labile aversive memory. Curr Biol. 2010;20:1445–1451. doi: 10.1016/j.cub.2010.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aso Y, Herb A, Ogueta M, Siwanowicz I, Templier T, Friedrich AB, Ito K, Scholz H, Tanimoto H. Three dopamine pathways induce aversive odor memories with different stability. PLoS Genet. 2012;8:e1002768. doi: 10.1371/journal.pgen.1002768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aso Y, Hattori D, Yu Y, Johnston RM, Iyer NA, Ngo TT, Dionne H, Abbott LF, Axel R, Tanimoto H, Rubin GM. The neuronal architecture of the mushroom body provides a logic for associative learning. Elife. 2014;3:e04577. doi: 10.7554/eLife.04577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaulieu JM, Gainetdinov RR. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev. 2011;63:182–217. doi: 10.1124/pr.110.002642. [DOI] [PubMed] [Google Scholar]
- Berridge KC, Kringelbach ML. Pleasure systems in the brain. Neuron. 2015;86:646–664. doi: 10.1016/j.neuron.2015.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry JA, Cervantes-Sandoval I, Nicholas EP, Davis RL. Dopamine is required for learning and forgetting in Drosophila. Neuron. 2012;74:530–542. doi: 10.1016/j.neuron.2012.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry JA, Cervantes-Sandoval I, Chakraborty M, Davis RL. Sleep facilitates memory by blocking dopamine neuron-mediated forgetting. Cell. 2015;161:1656–1667. doi: 10.1016/j.cell.2015.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blum AL, Li W, Cressy M, Dubnau J. Short- and long-term memory in Drosophila require cAMP signaling in distinct neuron types. Curr Biol. 2009;19:1341–1350. doi: 10.1016/j.cub.2009.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonci A, Hopf FW. The dopamine D2 receptor: new surprises from an old friend. Neuron. 2005;47:335–338. doi: 10.1016/j.neuron.2005.07.015. [DOI] [PubMed] [Google Scholar]
- Boto T, Louis T, Jindachomthong K, Jalink K, Tomchik SM. Dopaminergic modulation of cAMP drives nonlinear plasticity across the Drosophila mushroom body lobes. Curr Biol. 2014;24:822–831. doi: 10.1016/j.cub.2014.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouzaiane E, Trannoy S, Scheunemann L, Plaçais PY, Preat T. Two independent mushroom body output circuits retrieve the six discrete components of Drosophila aversive memory. Cell Rep. 2015;11:1280–1292. doi: 10.1016/j.celrep.2015.04.044. [DOI] [PubMed] [Google Scholar]
- Byers D, Davis RL, Kiger JA., Jr Defect in cyclic AMP phosphodiesterase due to the dunce mutation of learning in Drosophila melanogaster. Nature. 1981;289:79–81. doi: 10.1038/289079a0. [DOI] [PubMed] [Google Scholar]
- Cheung US, Shayan AJ, Boulianne GL, Atwood HL. Drosophila larval neuromuscular junction's responses to reduction of cAMP in the nervous system. J Neurobiol. 1999;40:1–13. doi: 10.1002/(SICI)1097-4695(199907)40:1<1::AID-NEU1>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- Cheung U, Atwood HL, Zucker RS. Presynaptic effectors contributing to cAMP-induced synaptic potentiation in Drosophila. J Neurobiol. 2006;66:273–280. doi: 10.1002/neu.20218. [DOI] [PubMed] [Google Scholar]
- Chou YH, Spletter ML, Yaksi E, Leong JC, Wilson RI, Luo L. Diversity and wiring variability of olfactory local interneurons in the Drosophila antennal lobe. Nat Neurosci. 2010;13:439–449. doi: 10.1038/nn.2489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claridge-Chang A, Roorda RD, Vrontou E, Sjulson L, Li H, Hirsh J, Miesenböck G. Writing memories with light-addressable reinforcement circuitry. Cell. 2009;139:405–415. doi: 10.1016/j.cell.2009.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook RK, Christensen SJ, Deal JA, Coburn RA, Deal ME, Gresens JM, Kaufman TC, Cook KR. The generation of chromosomal deletions to provide extensive coverage and subdivision of the Drosophila melanogaster genome. Genome Biol. 2012;13:R21. doi: 10.1186/gb-2012-13-3-r21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crittenden JR, Skoulakis EM, Han KA, Kalderon D, Davis RL. Tripartite mushroom body architecture revealed by antigenic markers. Learn Mem. 1998;5:38–51. [PMC free article] [PubMed] [Google Scholar]
- Das A, Sen S, Lichtneckert R, Okada R, Ito K, Rodrigues V, Reichert H. Drosophila olfactory local interneurons and projection neurons derive from a common neuroblast lineage specified by the empty spiracles gene. Neural Dev. 2008;3:33. doi: 10.1186/1749-8104-3-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis RL, Kiger JA., Jr Dunce mutants of Drosophila melanogaster: mutants defective in the cyclic AMP phosphodiesterase enzyme system. J Cell Biol. 1981;90:101–107. doi: 10.1083/jcb.90.1.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doya K. Modulators of decision making. Nat Neurosci. 2008;11:410–416. doi: 10.1038/nn2077. [DOI] [PubMed] [Google Scholar]
- Draper I, Kurshan PT, McBride E, Jackson FR, Kopin AS. Locomotor activity is regulated by D2-like receptors in Drosophila: an anatomic and functional analysis. Dev Neurobiol. 2007;67:378–393. doi: 10.1002/dneu.20355. [DOI] [PubMed] [Google Scholar]
- Faure M, Voyno-Yasenetskaya TA, Bourne HR. cAMP and beta gamma subunits of heterotrimeric G proteins stimulate the mitogen-activated protein kinase pathway in COS-7 cells. J Biol Chem. 1994;269:7851–7854. [PubMed] [Google Scholar]
- Ford CP. The role of D2-autoreceptors in regulating dopamine neuron activity and transmission. Neuroscience. 2014;282C:13–22. doi: 10.1016/j.neuroscience.2014.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghahremani MH, Forget C, Albert PR. Distinct roles for Galpha(i)2 and Gbetagamma in signaling to DNA synthesis and Galpha(i)3 in cellular transformation by dopamine D2S receptor activation in BALB/c 3T3 cells. Mol Cell Biol. 2000;20:1497–1506. doi: 10.1128/MCB.20.5.1497-1506.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo A, Li L, Xia SZ, Feng CH, Wolf R, Heisenberg M. Conditioned visual flight orientation in Drosophila: dependence on age, practice, and diet. Learn Mem. 1996;3:49–59. doi: 10.1101/lm.3.1.49. [DOI] [PubMed] [Google Scholar]
- Han KA, Millar NS, Grotewiel MS, Davis RL. DAMB, a novel dopamine receptor expressed specifically in Drosophila mushroom bodies. Neuron. 1996;16:1127–1135. doi: 10.1016/S0896-6273(00)80139-7. [DOI] [PubMed] [Google Scholar]
- Hearn MG, Ren Y, McBride EW, Reveillaud I, Beinborn M, Kopin AS. A Drosophila dopamine 2-like receptor: molecular characterization and identification of multiple alternatively spliced variants. Proc Natl Acad Sci U S A. 2002;99:14554–14559. doi: 10.1073/pnas.202498299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heimbeck G, Bugnon V, Gendre N, Keller A, Stocker RF. A central neural circuit for experience-independent olfactory and courtship behavior in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2001;98:15336–15341. doi: 10.1073/pnas.011314898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heisenberg M. Mushroom body memoir: from maps to models. Nat Rev Neurosci. 2003;4:266–275. doi: 10.1038/nrn1074. [DOI] [PubMed] [Google Scholar]
- Hummel T, Vasconcelos ML, Clemens JC, Fishilevich Y, Vosshall LB, Zipursky SL. Axonal targeting of olfactory receptor neurons in Drosophila is controlled by Dscam. Neuron. 2003;37:221–231. doi: 10.1016/S0896-6273(02)01183-2. [DOI] [PubMed] [Google Scholar]
- Kim YC, Lee HG, Seong CS, Han KA. Expression of a D1 dopamine receptor dDA1/DmDOP1 in the central nervous system of Drosophila melanogaster. Gene Expr Patterns. 2003;3:237–245. doi: 10.1016/S1567-133X(02)00098-4. [DOI] [PubMed] [Google Scholar]
- Kim YC, Lee HG, Han KA. D1 dopamine receptor dDA1 is required in the mushroom body neurons for aversive and appetitive learning in Drosophila. J Neurosci. 2007;27:7640–7647. doi: 10.1523/JNEUROSCI.1167-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitamoto T. Conditional disruption of synaptic transmission induces male-male courtship behavior in Drosophila. Proc Natl Acad Sci U S A. 2002;99:13232–13237. doi: 10.1073/pnas.202489099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knapek S, Sigrist S, Tanimoto H. Bruchpilot, a synaptic active zone protein for anesthesia-resistant memory. J Neurosci. 2011;31:3453–3458. doi: 10.1523/JNEUROSCI.2585-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koga E, Momiyama T. Presynaptic dopamine D2-like receptors inhibit excitatory transmission onto rat ventral tegmental dopaminergic neurones. J Physiol. 2000;523:163–173. doi: 10.1111/j.1469-7793.2000.t01-2-00163.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krashes MJ, Keene AC, Leung B, Armstrong JD, Waddell S. Sequential use of mushroom body neuron subsets during Drosophila odor memory processing. Neuron. 2007;53:103–115. doi: 10.1016/j.neuron.2006.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Krause WC, Davis RL. GABA(A) receptor RDL inhibits Drosophila olfactory associative learning. Neuron. 2007;56:1090–1102. doi: 10.1016/j.neuron.2007.10.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livingstone MS, Sziber PP, Quinn WG. Loss of calcium/calmodulin responsiveness in adenylate cyclase of rutabaga, a Drosophila learning mutant. Cell. 1984;37:205–215. doi: 10.1016/0092-8674(84)90316-7. [DOI] [PubMed] [Google Scholar]
- Margulies C, Tully T, Dubnau J. Deconstructing memory in Drosophila. Curr Biol. 2005;15:R700–R713. doi: 10.1016/j.cub.2005.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuire SE, Le PT, Osborn AJ, Matsumoto K, Davis RL. Spatiotemporal rescue of memory dysfunction in Drosophila. Science. 2003;302:1765–1768. doi: 10.1126/science.1089035. [DOI] [PubMed] [Google Scholar]
- Mitchell R, McCulloch D, Lutz E, Johnson M, MacKenzie C, Fennell M, Fink G, Zhou W, Sealfon SC. Rhodopsin-family receptors associate with small G proteins to activate phospholipase D. Nature. 1998;392:411–414. doi: 10.1038/32937. [DOI] [PubMed] [Google Scholar]
- Nakano T, Doi T, Yoshimoto J, Doya K. A kinetic model of dopamine- and calcium-dependent striatal synaptic plasticity. PLoS Comput Biol. 2010;6:e1000670. doi: 10.1371/journal.pcbi.1000670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neve KA, Seamans JK, Trantham-Davidson H. Dopamine receptor signaling. J Recept Signal Transduct Res. 2004;24:165–205. doi: 10.1081/RRS-200029981. [DOI] [PubMed] [Google Scholar]
- Nutt DJ, Lingford-Hughes A, Erritzoe D, Stokes PR. The dopamine theory of addiction: 40 years of highs and lows. Nat Rev Neurosci. 2015;16:305–312. doi: 10.1038/nrn3939. [DOI] [PubMed] [Google Scholar]
- Pan Y, Zhou Y, Guo C, Gong H, Gong Z, Liu L. Differential roles of the fan-shaped body and the ellipsoid body in Drosophila visual pattern memory. Learn Mem. 2009;16:289–295. doi: 10.1101/lm.1331809. [DOI] [PubMed] [Google Scholar]
- Parks AL, Cook KR, Belvin M, Dompe NA, Fawcett R, Huppert K, Tan LR, Winter CG, Bogart KP, Deal JE, Deal-Herr ME, Grant D, Marcinko M, Miyazaki WY, Robertson S, Shaw KJ, Tabios M, Vysotskaia V, Zhao L, Andrade RS, et al. Systematic generation of high-resolution deletion coverage of the Drosophila melanogaster genome. Nat Genet. 2004;36:288–292. doi: 10.1038/ng1312. [DOI] [PubMed] [Google Scholar]
- Parnas M, Lin AC, Huetteroth W, Miesenböck G. Odor discrimination in Drosophila: from neural population codes to behavior. Neuron. 2013;79:932–944. doi: 10.1016/j.neuron.2013.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pignatelli M, Bonci A. Role of dopamine neurons in reward and aversion: a synaptic plasticity perspective. Neuron. 2015;86:1145–1157. doi: 10.1016/j.neuron.2015.04.015. [DOI] [PubMed] [Google Scholar]
- Pitman JL, Huetteroth W, Burke CJ, Krashes MJ, Lai SL, Lee T, Waddell S. A pair of inhibitory neurons are required to sustain labile memory in the Drosophila mushroom body. Curr Biol. 2011;21:855–861. doi: 10.1016/j.cub.2011.03.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plaçais PY, Trannoy S, Isabel G, Aso Y, Siwanowicz I, Belliart-Guérin G, Vernier P, Birman S, Tanimoto H, Preat T. Slow oscillations in two pairs of dopaminergic neurons gate long-term memory formation in Drosophila. Nat Neurosci. 2012;15:592–599. doi: 10.1038/nn.3055. [DOI] [PubMed] [Google Scholar]
- Qi C, Lee D. Pre- and postsynaptic role of dopamine D2 receptor DD2R in Drosophila olfactory associative learning. Biology. 2014;3:831–845. doi: 10.3390/biology3040831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin H, Cressy M, Li W, Coravos JS, Izzi SA, Dubnau J. Gamma neurons mediate dopaminergic input during aversive olfactory memory formation in Drosophila. Curr Biol. 2012;22:608–614. doi: 10.1016/j.cub.2012.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sachse S, Rueckert E, Keller A, Okada R, Tanaka NK, Ito K, Vosshall LB. Activity-dependent plasticity in an olfactory circuit. Neuron. 2007;56:838–850. doi: 10.1016/j.neuron.2007.10.035. [DOI] [PubMed] [Google Scholar]
- Scheunemann L, Jost E, Richlitzki A, Day JP, Sebastian S, Thum AS, Efetova M, Davies SA, Schwärzel M. Consolidated and labile odor memory are separately encoded within the Drosophila brain. J Neurosci. 2012;32:17163–17171. doi: 10.1523/JNEUROSCI.3286-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheunemann L, Skroblin P, Hundsrucker C, Klussmann E, Efetova M, Schwärzel M. AKAPS act in a two-step mechanism of memory acquisition. J Neurosci. 2013;33:17422–17428. doi: 10.1523/JNEUROSCI.1303-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroll C, Riemensperger T, Bucher D, Ehmer J, Völler T, Erbguth K, Gerber B, Hendel T, Nagel G, Buchner E, Fiala A. Light-induced activation of distinct modulatory neurons triggers appetitive or aversive learning in Drosophila larvae. Curr Biol. 2006;16:1741–1747. doi: 10.1016/j.cub.2006.07.023. [DOI] [PubMed] [Google Scholar]
- Schultz W. Neural coding of basic reward terms of animal learning theory, game theory, microeconomics and behavioural ecology. Curr Opin Neurobiol. 2004;14:139–147. doi: 10.1016/j.conb.2004.03.017. [DOI] [PubMed] [Google Scholar]
- Schultz W. Neuronal reward and decision signals: from theories to data. Physiol Rev. 2015;95:853–951. doi: 10.1152/physrev.00023.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwaerzel M, Monastirioti M, Scholz H, Friggi-Grelin F, Birman S, Heisenberg M. Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in Drosophila. J Neurosci. 2003;23:10495–10502. doi: 10.1523/JNEUROSCI.23-33-10495.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwärzel M, Müller U. Dynamic memory networks: dissecting molecular mechanisms underlying associative memory in the temporal domain. Cell Mol Life Sci. 2006;63:989–998. doi: 10.1007/s00018-006-6024-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Séjourné J, Plaçais PY, Aso Y, Siwanowicz I, Trannoy S, Thoma V, Tedjakumala SR, Rubin GM, Tchénio P, Ito K, Isabel G, Tanimoto H, Preat T. Mushroom body efferent neurons responsible for aversive olfactory memory retrieval in Drosophila. Nat Neurosci. 2011;14:903–910. doi: 10.1038/nn.2846. [DOI] [PubMed] [Google Scholar]
- Stocker RF, Heimbeck G, Gendre N, de Belle JS. Neuroblast ablation in Drosophila P[GAL4] lines reveals origins of olfactory interneurons. J Neurobiol. 1997;32:443–456. doi: 10.1002/(SICI)1097-4695(199705)32:5<443::AID-NEU1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Tanaka NK, Ito K, Stopfer M. Odor-evoked neural oscillations in Drosophila are mediated by widely branching interneurons. J Neurosci. 2009;29:8595–8603. doi: 10.1523/JNEUROSCI.1455-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tempel BL, Bonini N, Dawson DR, Quinn WG. Reward learning in normal and mutant Drosophila. Proc Natl Acad Sci U S A. 1983;80:1482–1486. doi: 10.1073/pnas.80.5.1482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tully T, Quinn WG. Classical conditioning and retention in normal and mutant Drosophila melanogaster. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 1985;157:263–277. doi: 10.1007/BF01350033. [DOI] [PubMed] [Google Scholar]
- Urs NM, Bido S, Peterson SM, Daigle TL, Bass CE, Gainetdinov RR, Bezard E, Caron MG. Targeting beta-arrestin2 in the treatment of l-DOPA-induced dyskinesia in Parkinson's disease. Proc Natl Acad Sci U S A. 2015;112:E2517–E2526. doi: 10.1073/pnas.1502740112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watts VJ, Neve KA. Sensitization of adenylate cyclase by Galpha i/o-coupled receptors. Pharmacol Ther. 2005;106:405–421. doi: 10.1016/j.pharmthera.2004.12.005. [DOI] [PubMed] [Google Scholar]
- West RJ, Furmston R, Williams CA, Elliott CJ. Neurophysiology of Drosophila models of Parkinson's disease. Parkinsons Dis. 2015;2015:381281. doi: 10.1155/2015/381281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu CL, Shih MF, Lee PT, Chiang AS. An octopamine-mushroom body circuit modulates the formation of anesthesia-resistant memory in Drosophila. Curr Biol. 2013;23:2346–2354. doi: 10.1016/j.cub.2013.09.056. [DOI] [PubMed] [Google Scholar]
- Yamagata N, Ichinose T, Aso Y, Plaçais PY, Friedrich AB, Sima RJ, Preat T, Rubin GM, Tanimoto H. Distinct dopamine neurons mediate reward signals for short- and long-term memories. Proc Natl Acad Sci U S A. 2015;112:578–583. doi: 10.1073/pnas.1421930112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan Z, Song WJ, Surmeier J. D2 dopamine receptors reduce N-type Ca2+ currents in rat neostriatal cholinergic interneurons through a membrane-delimited, protein-kinase-C-insensitive pathway. J Neurophysiol. 1997;77:1003–1015. doi: 10.1152/jn.1997.77.2.1003. [DOI] [PubMed] [Google Scholar]
- Yan Z, Feng J, Fienberg AA, Greengard P. D(2) dopamine receptors induce mitogen-activated protein kinase and cAMP response element-binding protein phosphorylation in neurons. Proc Natl Acad Sci U S A. 1999;96:11607–11612. doi: 10.1073/pnas.96.20.11607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zamponi GW, Snutch TP. Decay of prepulse facilitation of N type calcium channels during G protein inhibition is consistent with binding of a single Gbeta subunit. Proc Natl Acad Sci U S A. 1998;95:4035–4039. doi: 10.1073/pnas.95.7.4035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zars T, Wolf R, Davis R, Heisenberg M. Tissue-specific expression of a type I adenylyl cyclase rescues the rutabaga mutant memory defect: in search of the engram. Learn Mem. 2000;7:18–31. doi: 10.1101/lm.7.1.18. [DOI] [PMC free article] [PubMed] [Google Scholar]