

Abstract
Purpose of Review
This review summarizes the role of hypoxia and hypoxia-inducible factors (HIFs) in the regulation of stem cell biology, specifically focusing on maintenance, differentiation, and stress responses in the context of several stem cell systems. Stem cells for different lineages/tissues reside in distinct niches, and are exposed to diverse oxygen concentrations. Recent studies have revealed the importance of the hypoxia signaling pathway for stem cell functions.
Recent Findings
Hypoxia and HIFs contribute to maintenance of embryonic stem cells, generation of induced pluripotent stem cells, functionality of hematopoietic stem cells, and survival of leukemia stem cells. Harvest and collection of mouse bone marrow and human cord blood cells in ambient air results in fewer hematopoietic stem cells recovered due to the phenomenon of Extra PHysiologic Oxygen Shock/Stress (EPHOSS).
Summary
Oxygen is an important factor in the stem cell microenvironment. Hypoxia signaling and HIFs play important roles in modeling cellular metabolism in both stem cells and niches to regulate stem cell biology, and represent an additional dimension that allows stem cells to maintain an undifferentiated status and multilineage differentiation potential.
Keywords: Hypoxia, HIF, Hematopoietic stem cells, Cancer stem cells, EPHOSS
Introduction
Interactions between stem cells and their niche microenvironment are critical for stem cell self-renewal and protection of the stem cell pool [1–4]. Oxygen is a vital molecule in the stem cell microenvironment, serving as a metabolic substrate and signaling mediator [5]. While organs and tissues experience a considerably lower range of oxygen tension compared with the ambient oxygen levels (normoxia, 21%), measurements in stem cell niche revealed even lower oxygen tensions (Table 1) [6–15]. Low oxygen levels (hypoxia) are appreciated to maintain a slow-cycling proliferation property, reduced oxidative stress, and undifferentiated status in several stem cell populations [16–18].
Table 1.
Low oxygen levels in diverse stem cell compartments
The effects of hypoxia are primarily mediated by hypoxiainducible factors (HIFs) [19, 20]. HIFs are DNA-binding transcriptional factors of the basic-helix-loop-helix-Per-Arnt-Sim (bHLH-PAS) superfamily proteins and bind to hypoxia regulated elements (5′-RCGTG-3′, HREs) in the promoter and enhancer of hypoxia-inducible genes [21–23]. HIFs are heterodimers composed of an alpha subunit (HIF-α) and a beta (HIF-β) subunit, also known as aryl-hydrocarbon receptor nuclear translocator (ARNT) [20]. To date, three alpha subunits (HIF1A, HIF2A, HIF3A) and two beta subunits (ARNT, ARNT2) have been identified [23–26]. All of the HIF-α subunits contain an N-terminal bHLH domain for DNA binding, a PAS domain for heterodimerization, and an oxygen-dependent degradation (ODD) domain for regulation of protein stability. HIF1A is ubiquitously expressed, while expression of HIF2A and HIF3A is restricted to specific cell types [27]. ARNT and ARNT2 share 57% amino acid sequence identity and both contain bHLH and PAS domains [28, 29].
The HIF-α subunit is regulated by an oxygen-dependent proteolytic degradation, whereas the HIF-β subunit is constitutively expressed. Under ambient oxygen levels, HIF-α is hydroxylated on proline residues within the ODD domain by prolyl hydroxylases (PHD), which is recognized by von Hippel-Lindau (VHL) E3 ubiquitin ligase. VHL targets HIF-α for ubiquitin conjugation and proteasomal degradation (Fig. 1) [30–32]. Under hypoxia, when PHD activity is suppressed, HIF-α is stabilized and translocated to the nucleus where it dimerizes with ARNT to promote transcription of various hypoxia-inducible genes (Fig. 1) [33, 34]. HIF-α/ ARNT heterodimer can recruit histone acetyltransferases p300 and CBP to activate transcription of target genes [35, 36]. HIF-α activity is also regulated by additional protein modifications, such as phosphorylation [37], acetylation [38], and sumoylation [39].
Fig. 1.
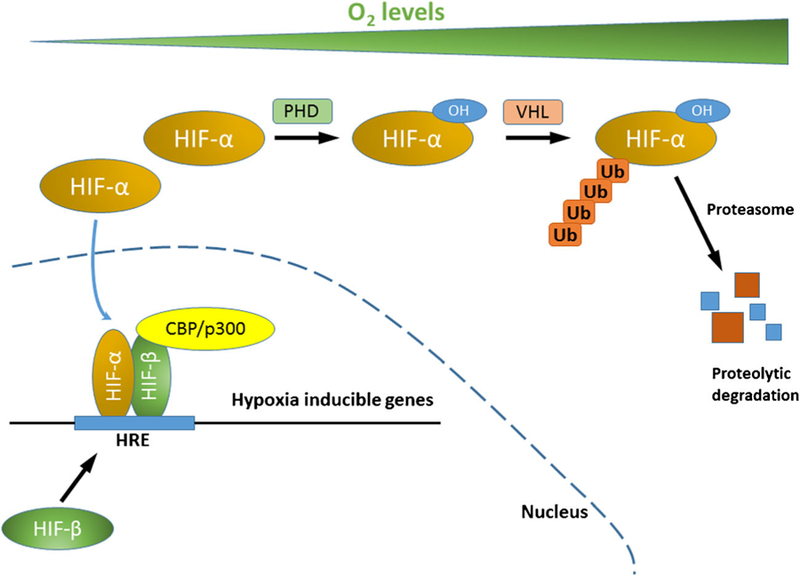
HIF mediated hypoxia signaling. Under normoxic oxygen tension, HIF-α is hydroxylated on proline residues by PHD, which is recognized by VHL E3 ubiquitin ligase. VHL targets HIF-α for polyubiquitin conjugation and proteasomal degradation. Under hypoxia, when PHD activity is suppressed, HIF-α is stabilized and translocated to the nucleus where it dimerizes with HIF-β. HIF-α/HIF-β heterodimer binds to HRE at the promoters of hypoxia-inducible genes, recruits histone acetyltransferases p300 and CBP to activate transcription of target genes. (HIF, hypoxia-inducible factor; PHD, prolyl hydroxylase; OH, hydroxyl group; VHL, von Hippel-Lindau; Ub, ubiquitin; HRE, hypoxia regulated elements; CBP, CREB binding protein)
While the roles of hypoxia signaling and HIFs in development have been reviewed by others, their effects on stem cells have received scant attention. In this review, we summarize recent advances on hypoxia signaling and HIFs with a focus on the regulation of the biology of several types of stem cells including pluripotent stem cells (PSCs), hematopoietic stem cells (HSCs), and cancer stem cells (CSCs).
Hypoxia and HIFs in Embryonic Stem Cells and Induced Pluripotent Stem Cells
Embryonic stem cells (ESCs) are capable of self-renewal and maintenance of pluripotency [40]. ESCs have been regarded as a potential replacement for tissues in regenerative medicine due to their pluripotency. Studies have demonstrated the advantage of generating ESC lines under hypoxic conditions (5% O2) over ambient air, in terms of increased colonies with alkaline phosphatase activity and enhanced cell proliferation [41, 42]. Culturing cells in hypoxia reduced spontaneous differentiation and enhanced formation of embryoid bodies, and seemed to maintain the pluripotency of the cells [43, 44]. A marker for early differentiation (SSEA-1) was less expressed when human ESCs were cultured under hypoxia conditions [43]. Culturing human ESCs at hypoxia was correlated with increased expression of NANOG, OCT4, and SOX2, main regulators of the undifferentiated status of embryonic cells [45]. In addition, HIF2A depletion hampered proliferation and protein expressions of OCT4, SOX2, and NANOG in human ESCs [45]. Enhanced maintenance of self-renewal and pluripotency of human ESCs was observed when cocultured with human fetal liver stromal cells expressing HIF1A, suggesting that the function of HIF1A in the microenvironment is important to maintain the undifferentiated state of human ESCs [46].
Hypoxia plays a crucial role in determining the fate of the stem cells [47]. However, there are conflicting reports on the effect of hypoxia on ESC differentiation. Reports showed that hypoxia maintained self-renewal and prevented differentiation of ESCs [43]. In contrast, others revealed that hypoxia promoted differentiation of human ESCs into cardiomyocytes [48] and chondrocytes [49]. Hypoxia also promoted mouse ESCs to differentiate to neurons [50], endothelial cells, and hematopoietic stem cells [51]. These apparently controversial reports on the effect of hypoxia on differentiation may be explained by the stage of stemness at which the hypoxia was introduced, and the duration and degree of hypoxia. The mechanism through which HIF induces differentiation to different lineages has been revealing. Hypoxia primed ESCs to commit to the vascular lineage by suppressing Oct4 expression via direct binding of HIF1A in the Oct4 promoter region [52]. Another group reported that Hif1a induced mouse ESC commitment to arterial endothelia cells through upregulation of the transcription factor Etv2 and Notch1 signaling [53•]. Moreover, overexpression of Hif2a primed mouse ESCs to commit to cardiomyocytes by upregulating β-catenin [54]. Knocking out HIF1A by the CRSIPR-Cas9 system blocked transition from naive to primed human ESCs [55].
The groundbreaking finding by Yamanaka and colleagues demonstrated that somatic cells could be reprogrammed into induced pluripotent stem cells (iPSCs) by four transcription factors (4F; Oct4, Sox2, Klf4 and c-Myc) [56]. The generation of iPSCs was inefficient under normoxic conditions at ambient air (21% O2), and the reprogramming efficiency was significantly higher under mildly hypoxic conditions (5% O2) [57]. This finding suggests that the condition of hypoxia enhances stem cell generation and maintenance, consistent with observations in ESCs. One distinct characteristic of ESC and iPSC is their reliance on glycolytic metabolism, regardless of oxygen availability [58, 59]. Cellular bioenergetics are also extensively remodeled upon generation of iPSCs, from mitochondrial oxidative to glycolytic metabolism [60, 61]. Cells with disrupted HIF1A function are refractory to reprogramming [61]. Also, a small molecule activation of HIF1A upregulated HIF1A targets (PKM2 and PDK3), resulting in increased glycolysis and enhanced reprogramming efficiency [61]. One group continued to dissect the HIF regulatory role in metabolic switch and reprogramming [62]. They proposed that HIF1A and HIF2A were sufficient to induce metabolic switching during reprogramming. They distinguished between HIF1A and HIF2A functions during reprogramming. Both were essential, but HIF2A applied its positive regulatory effect early in the process. Interestingly, prolonged stabilization of HIF2A impeded iPSC formation through TRAILinduced inhibition of caspase 3 [62]. Another report linked hypoxia, mitofusins (Mfn), and HIF1A with reprogramming [63]. Under hypoxic conditions, Mfn were downregulated and less expressed. In Mfn1/2 knockdown cells, HIF1A was stabilized, resulting in enhanced glycolytic metabolism and induced pluripotency [63].
Hypoxia Signaling and HIFs in Tissue-Specific Stem Cells, Using Hematopoietic Stem Cells as an Example
Each day, 100 billion new blood cells are produced by the human body. The continuous production of these cells depends on the presence of a rare population of HSCs [64–66]. HSCs are one of the best characterized adult stem cell types, and HSC transplantation is the only curative therapy for a variety of malignant and non-malignant blood diseases [67]. HSCs reside in a specialized bone marrow microenvironment, where they are adjacent to endothelial cells, osteoblasts, and stromal cells [68, 69]. Direct in vivo measurements revealed quite low oxygen tension in the BM of live mice with the lowest (1.3%) oxygen tension in deeper peri-sinusoidal regions [70••]. HSCs have a hypoxic profile including high levels of HIF1A protein expression [71], suggesting that hypoxia signaling plays an important role in HSC biology.
Genetic manipulations of mice have revealed the significance of HIFs in HSCs. Conditional deletion of Hif1a in the adult hematopoietic system resulted in loss of the quiescence state, and decreased repopulating capability of HSCs [72]. Hif1a null HSCs showed a metabolic shift from glycolysis to oxidative phosphorylation, with an increase in oxygen consumption rate and a decrease in anaerobic glycolysis [72, 73]. Consistently, the expression of pyruvate dehydrogenase kinases, which normally promotes glycolysis, was downregulated in Hif1a null HSCs [73]. Overexpression of pyruvate dehydrogenase kinases in Hif1a-null HSCs rescued the glycolysis phenotype and restored normal HSC activity. Conditional knockout of Meis1 in HSCs resulted in decreased expression of Hif1a and Hif2a, and demonstrated a similar metabolic shift pattern (from glycolysis to mitochondria metabolism), loss of HSC quiescence, and impaired bone marrow engraftment after transplantation [74, 75]. It has been reported that deletion of Hif2a within the hematopoietic system had no impact on HSC function [76]; however, knockdown of HIF2A in human CD34+ cells (enriched for HSCs and their progenitor cells) resulted in impaired reconstitution capacity [77], suggesting a potential distinct role of HIF2A between mouse and human HSCs. Of note, Hif2a-null recipient mice failed to sustain hematopoiesis when transplanted with wild-type donor cells suggesting the importance and contribution of Hif2a in the HSC microenvironment for normal hematopoiesis [78].
Genetic knockouts on negative regulators of Hif resulted in enhanced hypoxia signaling and generated a significant phenotype in HSCs. Deletion of Phd2, a Hif prolyl hydroxylase, resulted in stabilization of both Hif1a and Hif2a [79]. Recipient mice transplanted with Phd2 deficient bone marrow cells showed increased engraftment and donor reconstitution which was reliant on Hif1a, indicating that Phd2 depletion and enhanced hypoxia signaling promote HSC proliferation after transplantation. Similarly, monoallelic loss of Vhl, an E3 ligase of Hif, enhanced cell cycle quiescence in HSCs and promoted engraftment after bone marrow transplantation [72]. Interestingly, biallelic loss of Vhl caused overstabilization of Hifs in HSCs and impaired transplantation capacity, possibly due to dysregulated cell cycle status and homing defects [72]. Furthermore, Vhl deletion in osteoblasts increased HSC and progenitor frequency, suggesting an important role of Vhl and Hifs in the HSC microenvironment [80]. In addition, conditional deletion of Cited2, another negative regulator of Hif1a, in the hematopoietic system resulted in loss of HSCs and bone marrow failure [81]. These results suggest that appropriate control of levels of Hifs in HSCs and their niche are essential for determining stem cell capacity.
HSC transplantation efficacy is a vital factor for successful donor engraftment during clinical therapy. Many efforts have been made to enhance human HSC transplantation efficacy via ex vivo expansion [82–86] or enhance homing [87•, 88, 89]. HIF1A has been reported to be involved in both of these efforts. Pharmacologic increase in HIF1A levels via prostaglandin E2 or dimethyloxalylglycine (DMOG) resulted in upregulation of surface CXCR4 expression, leading to enhanced HSC homing and engraftment [90]. In addition, HIF1A is essential for HSC mobilization induced by using G-CSF and AMD3100; pharmacologic stabilization of HIF1A by FG-4497 also enhanced mobilization of reconstituting HSCs [91]. NR-101, a novel c-MPL agonist, induced long-term accumulation of HIF1A protein and thus promoted ex vivo expansion of human cord blood HSCs, and enhanced their repopulating capacity [92].
Our laboratory recently reported that harvest, collection, and processing of mouse bone marrow and human cord blood HSCs under hypoxia (3% O2) resulted in 2–5-fold increases in the recovery of long-term repopulating HSCs compared with ambient air collection, through a phenomenon we named Bextra physiologic oxygen shock/stress^ (EPHOSS) [93••, 94]. EPHOSS is mediated by cyclophilin D and the mitochon-drial permeability transition pore (MPTP), with links to p53 and HIF1A. By modulating MPTP opening via cyclophilin D inhibition genetically or by using the small molecule inhibitor cyclosporine A, HSCs were protected from EPHOSS, resulting in increased collection of HSCs. This demonstrated that there are greater numbers of HSCs residing in bone marrow than previously reported, and suggesting that HSC transplantation could be improved if EPHOSS is suppressed by collecting and processing cells under hypoxia, or by using cyclosporine A or other modulators. Other means to suppress the phenomenon of EPHOSS for enhanced collection of HSCs are currently underway in our laboratory.
Hypoxia Signaling and HIFs in Cancer Stem Cells
Even though medical breakthroughs have advanced cancer care over the past decades, the high mortality rate due to disease recurrence remains one of the biggest challenges in the field. By definition, cancer stem cells (CSCs) are cells that have gained the abilities of self-renewal, as well as the capacity to reconstitute the heterogeneity of the tumor [95]. CSCs exist in several types of human cancers including blood malignancies [96] and solid tumors [97–99]. They are believed to be responsible for distant metastasis, tumor recurrence, and therapy failure [100]. The expression of either HIF1A or HIF2A is positively correlated with poor prognosis in most of cancer types [101]. Recent evidence suggested that HIF1A plays a critical role for metabolic switch in cancer cells by regulating PKM2 [102]. While the role of hypoxia in selecting for solid tumor CSCs has been well-established [103, 104], the role that hypoxia and hypoxia-inducible factors play in the development and maintenance of leukemia stem cells (LSCs) is still a subject under debate [105]. The striking similarity between LSCs and HSCs in self-renewal capacity and the finding that HSCs reside in hypoxic niches in the bone marrow spurred interest in whether hypoxia also plays a critical role in LSC biology. Unfortunately, this poses an inevitable obstacle in targeting LSCs, without harming healthy HSCs.
A recent study demonstrated that the HIF1A–Notch pathway was responsible for sustaining CSCs self-renewal in two models of lymphoma and acute myeloid leukemia (AML), suggesting that this pathway may be able to be targeted to eliminate CSCs or at least to diminish the leukemia-initiating property of AML LSCs [106]. Notably, in the murine lymphoma model used (with insertional mutation of the Epm2a gene), the overexpression of Hif1a which is critical for CSC survival is interestingly hypoxia-independent because the degradation pathway of Hif1a by VHL is concurrently downregulated. In line with this, the same group also showed that echinomycin, a Hif1a inhibitor, when administered alone can prevent relapse in the MllPTD/WT:Flt3ITD/WT murine model of AML. The authors pointed out that this model, by not having the immunological barriers of xenogeneic grafts and carrying a spontaneous mutation found in a subgroup of AML patients, faithfully represented the human disease [106]. In addition to studies in AML, the roles of HIFs in progression and recurrence of PML-RARα-driven acute promyelocytic leukemia (APL) have been evaluated in two different human APL cell lines—APL NB4 and U937-PR9 [107]. Their microarray analysis of leukemic APL samples showed a HIFregulated gene profile significantly distinguishable between normal human promyelocytes and leukemic promyelocytes. In xenograft experiments, shRNA-induced inhibition of HIF1A reduced cell migration, colony forming ability, and prolonged mouse survival [107]. The results were confirmed in acute HIF1A inhibition with an RNA antagonist EZN2968, which showed even higher efficiency than chronic silencing of HIF1A. Complementary to these studies, the role of HIF2A in both normal human HSCs and progenitors, and AML cells was characterized [77]. Interestingly, expression of HIF2A in primary AML samples was not significantly different from normal bone marrow and varied between patients mostly because of diverse genetic abnormalities. However, similar to the findings with normal bone marrow, HIF2Aknockdown AML cells showed defects in engraftment of leukemia including in a sample from a relapsed case; this defect resulted from endoplasmic reticulum stress-induced increase in apoptosis [77]. Thus, both HIF1A and HIF2A appear to be essential for protecting AML LSCs and maintenance of leukemia.
In contrast to the above findings, some skepticism has been posed on the therapeutic value of inhibition of HIFs in leukemia treatment [108]. Conditional knockout of Hif1a in three different well-characterized murine models of AML was used: AML1-ETO9a with no relation to Hif1a, and Meis1 and MLL with direct signals for Hif1a. In all three models, the status of Hif1a expression did not appear to be critical for either the initiation or progression of leukemia. In fact, loss of Hif1a did not have any impact on self-renewal of LSCs and even promoted their proliferation [108]. Adding to this study, others reported that HIF1A and HIF2A together acted as suppressors of AML development and had no significant impact on disease maintenance [109]. Loss of Hif2a unexpectedly enhanced LSC development and shortened AML latency in two murine models of AML (Mll-AF9 and Meis1/Hoxa9) [109].
In addition to AML, the roles of hypoxia and HIFs in other types of LSCs have been investigated. Recent evidence suggested that HIF1A and hypoxia are crucial for the survival of chronic myeloid leukemia (CML) stem cells. HIF1A and its targets were overexpressed in BCRABL-expressing LSCs [110]. Using a BCR-ABL-driven CML murine model, it was observed that LSCs from Hif1a knockout mice had higher apoptotic rate and reduced colony formation, and were unable to propagate CML in secondary recipient. It was concluded that Hif1a is important for both development and maintenance of CML [110]. In agreement with these findings, another group also studied the role of hypoxia in CML but used primary human CML cells instead. They confirmed that hypoxia nurtured LSCs through upregulation of HIF1A even when BCR-ABL1 was effectively inhibited by imatinib [111]. They identified a profile of genes upregulated by hypoxia that are essential for cell survival in CML cells compared with healthy cord blood CD34+ cells [111]. Similar results were noted in bortezomib-resistant CML LSCs using both the K562 cell line and primary patient CML cells [112]. While research in the past 5 years has focused mostly on CML and AML, a study looked at T cell acute lymphoblastic leukemia (T-ALL) [113]. Intriguingly, HIF1A was found to induce expression of β-catenin, and the Wnt-β-catenin pathway was confined to the LSC subpopulation. Furthermore, deletion of HIF1A or β-catenin significantly reduced the LSC frequency but not the viability of bulk tumor cells.
Conclusions
HIFs are primary mediators of the metabolic switch in the hypoxic environment. Genetic manipulation of HIFs and other hypoxia signaling components by gene knockout in mice yielded different phenotypes in various stem cell compartments as reviewed above. This seems to support the idea that hypoxia signaling is critical in maintenance of stem cell function. Physiological oxygen environment or hypoxia greatly enhanced establishment of ESC lines, iPSC reprogramming efficiency, and recovery of HSCs. It will be interesting to see if they also share similar mechanisms involved in EPHOSS, as mediated by cyclophilin D and the MPTP [93••].
Hypoxia and HIFs contribute to the development and survival maintenance of LSCs in a number of blood malignancies. However, as in the case of AML, there have been conflicting results between studies, possibly due to different experimental designs and technical approaches, and further complicated by the genetic heterogeneity of human AML disease. Another very important point and particularly relevant for this topic to be considered is that the cells studied should be collected and processed in hypoxia to avoid the effects of EPHOSS [93••]. As demonstrated by our laboratory, collection of HSCs at ambient O2 levels induced ROS production and altered cell function, and likely gene expression patterns; hence, it would seem to be counterproductive for a true understanding of LSCs to study the physiology and pathology of LSCs without maintaining these cells in their native hypoxic status. In other types of leukemia, future studies are warranted as well because the concept of hypoxia in drugresistant LSCs is still emerging. Therefore, until a more thoroughly understanding of the role of HIFs in leukemia and LSCs is established, whether inhibition of HIFs promises therapeutic value will remain a question.
Acknowledgements
We thank other members in the Broxmeyer laboratory for helpful discussion and assistance. This work is supported by US Public Health Service Grants from the NIH to HEB: (R01 HL112669, R01 HL056416, U54 DK106846). AA is supported by NIH 5T32DK007519–32 to HEB, and TT is supported by NIH T32DK064466.
Footnotes
Conflict of Interest Xinxin Huang, Thao Trinh, and Arafat Aljoufi declare that they have no conflict of interest. Dr. Broxmeyer is a member of the Medical Scientific Advisory Board of CordUse, a cord blood banking company based in Orlando, Florida, but he reports no financial support.
Human and Animal Rights and Informed Consent This article does not contain any studies with human or animal subjects performed by any of the authors.
Papers of particular interests, published recently, have been highlighted as:
Of importance
Of major importance
References
- 1.Chen S, Lewallen M, Xie T. Adhesion in the stem cell niche: biological roles and regulation. Development 2013;140(2):255–65. 10.1242/dev.083139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mendelson A, Frenette PS. Hematopoietic stem cell niche maintenance during homeostasis and regeneration. Nat Med 2014;20(8):833–46. 10.1038/nm.3647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kfoury Y, Scadden DT. Mesenchymal cell contributions to the stem cell niche. Cell Stem Cell 2015;16(3):239–53. 10.1016/j.stem.2015.02.019. [DOI] [PubMed] [Google Scholar]
- 4.Fuentealba LC, Obernier K, Alvarez-Buylla A. Adult neural stem cells bridge their niche. Cell Stem Cell 2012;10(6):698–708. 10.1016/j.stem.2012.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jez M, Rozman P, Ivanovic Z, Bas T. Concise review: the role of oxygen in hematopoietic stem cell physiology. J Cell Physiol 2015;230(9):1999–2005. 10.1002/jcp.24953. [DOI] [PubMed] [Google Scholar]
- 6.Mohyeldin A, Garzon-Muvdi T, Quinones-Hinojosa A. Oxygen in stem cell biology: a critical component of the stem cell niche. Cell Stem Cell 2010;7(2):150–61. 10.1016/j.stem.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 7.Erecinska M, Silver IA. Tissue oxygen tension and brain sensitivity to hypoxia. Respir Physiol 2001;128(3):263–76. [DOI] [PubMed] [Google Scholar]
- 8.Dings J, Meixensberger J, Jager A, Roosen K. Clinical experience with 118 brain tissue oxygen partial pressure catheter probes. Neurosurgery 1998;43(5):1082–95. [DOI] [PubMed] [Google Scholar]
- 9.Fischer B, Bavister BD. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J Reprod Fertil 1993;99(2):673–9. [DOI] [PubMed] [Google Scholar]
- 10.Harrison JS, Rameshwar P, Chang V, Bandari P. Oxygen saturation in the bone marrow of healthy volunteers. Blood 2002;99(1): 394. [DOI] [PubMed] [Google Scholar]
- 11.Matsumoto A, Matsumoto S, Sowers AL, Koscielniak JW, Trigg NJ, Kuppusamy P, et al. Absolute oxygen tension (pO(2)) in murine fatty and muscle tissue as determined by EPR. Magn Reson Med 2005;54(6):1530–5. 10.1002/mrm.20714. [DOI] [PubMed] [Google Scholar]
- 12.Pasarica M, Sereda OR, Redman LM, Albarado DC, Hymel DT, Roan LE, et al. Reduced adipose tissue oxygenation in human obesity: evidence for rarefaction, macrophage chemotaxis, and inflammation without an angiogenic response. Diabetes 2009;58(3):718–25. 10.2337/db08-1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Eliasson P, Jonsson JI. The hematopoietic stem cell niche: low in oxygen but a nice place to be. J Cell Physiol 2010;222(1):17–22. 10.1002/jcp.21908. [DOI] [PubMed] [Google Scholar]
- 14.Kwan M, Niinikoski J, Hunt TK. In vivo measurements of oxygen tension in the cornea, aqueous humor, and anterior lens of the open eye. Investig Ophthalmol 1972;11(2):108–14. [PubMed] [Google Scholar]
- 15.Bath C, Yang S, Muttuvelu D, Fink T, Emmersen J, Vorum H, et al. Hypoxia is a key regulator of limbal epithelial stem cell growth and differentiation. Stem Cell Res 2013;10(3):349–60. 10.1016/j.scr.2013.01.004. [DOI] [PubMed] [Google Scholar]
- 16.Busuttil RA, Rubio M, Dolle ME, Campisi J, Vijg J. Oxygen accelerates the accumulation of mutations during the senescence and immortalization of murine cells in culture. Aging Cell 2003;2(6):287–94. [DOI] [PubMed] [Google Scholar]
- 17.Eliasson P, Rehn M, Hammar P, Larsson P, Sirenko O, Flippin LA, et al. Hypoxia mediates low cell-cycle activity and increases the proportion of long-term-reconstituting hematopoietic stem cells during in vitro culture. Exp Hematol 2010;38(4):301–10 e2. 10.1016/j.exphem.2010.01.005. [DOI] [PubMed] [Google Scholar]
- 18.Lekli I, Gurusamy N, Ray D, Tosaki A, Das DK. Redox regulation of stem cell mobilization. Can J Physiol Pharmacol 2009;87(12): 989–95. 10.1139/Y09-102. [DOI] [PubMed] [Google Scholar]
- 19.Majmundar AJ, Wong WJ, Simon MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol Cell 2010;40(2):294–309. 10.1016/j.molcel.2010.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kaelin WG Jr, Ratcliffe PJ. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Mol Cell 2008;30(4):393–402. 10.1016/j.molcel.2008.04.009. [DOI] [PubMed] [Google Scholar]
- 21.Kewley RJ, Whitelaw ML, Chapman-Smith A. The mammalian basic helix-loop-helix/PAS family of transcriptional regulators. Int J Biochem Cell Biol 2004;36(2):189–204. [DOI] [PubMed] [Google Scholar]
- 22.Wang GL, Semenza GL. Purification and characterization of hypoxia-inducible factor 1. J Biol Chem 1995;270(3):1230–7. [DOI] [PubMed] [Google Scholar]
- 23.Wang GL, Jiang BH, Rue EA, Semenza GL. Hypoxiainducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci U S A 1995;92(12):5510–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Makino Y, Kanopka A, Wilson WJ, Tanaka H, Poellinger L. Inhibitory PAS domain protein (IPAS) is a hypoxia-inducible splicing variant of the hypoxia-inducible factor-3alpha locus. J Biol Chem 2002;277(36):32405–8. 10.1074/jbc.C200328200. [DOI] [PubMed] [Google Scholar]
- 25.Maynard MA, Qi H, Chung J, Lee EH, Kondo Y, Hara S, et al. Multiple splice variants of the human HIF-3 alpha locus are targets of the von Hippel-Lindau E3 ubiquitin ligase complex. J Biol Chem 2003;278(13):11032–40. 10.1074/jbc.M208681200. [DOI] [PubMed] [Google Scholar]
- 26.Keith B, Adelman DM, Simon MC. Targeted mutation of the murine arylhydrocarbon receptor nuclear translocator 2 (Arnt2) gene reveals partial redundancy with Arnt. Proc Natl Acad Sci U S A 2001;98(12):6692–7. 10.1073/pnas.121494298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hu CJ, Wang LY, Chodosh LA, Keith B, Simon MC. Differential roles of hypoxia-inducible factor 1alpha (HIF-1alpha) and HIF2alpha in hypoxic gene regulation. Mol Cell Biol 2003;23(24): 9361–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sekine H, Mimura J, Yamamoto M, Fujii-Kuriyama Y. Unique and overlapping transcriptional roles of arylhydrocarbon receptor nuclear translocator (Arnt) and Arnt2 in xenobiotic and hypoxic responses. J Biol Chem 2006;281(49):37507–16. 10.1074/jbc.M606910200. [DOI] [PubMed] [Google Scholar]
- 29.Hirose K, Morita M, Ema M, Mimura J, Hamada H, Fujii H, et al. cDNA cloning and tissue-specific expression of a novel basic helix-loop-helix/PAS factor (Arnt2) with close sequence similarity to the aryl hydrocarbon receptor nuclear translocator (Arnt). Mol Cell Biol 1996;16(4):1706–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, et al. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O 2 sensing. Science 2001;292(5516):464–8. 10.1126/science.1059817. [DOI] [PubMed] [Google Scholar]
- 31.Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, et al. Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001;292(5516):468–72. 10.1126/science.1059796. [DOI] [PubMed] [Google Scholar]
- 32.Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O’Rourke J, Mole DR, et al. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 2001;107(1):43–54. [DOI] [PubMed] [Google Scholar]
- 33.Mole DR, Blancher C, Copley RR, Pollard PJ, Gleadle JM, Ragoussis J, et al. Genome-wide association of hypoxiainducible factor (HIF)-1alpha and HIF-2alpha DNA binding with expression profiling of hypoxia-inducible transcripts. J Biol Chem 2009;284(25):16767–75. 10.1074/jbc.M901790200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Schodel J, Oikonomopoulos S, Ragoussis J, Pugh CW, Ratcliffe PJ, Mole DR. High-resolution genome-wide mapping of HIFbinding sites by ChIP-seq. Blood 2011;117(23):e207–17. 10.1182/blood-2010-10-314427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Dames SA, Martinez-Yamout M, De Guzman RN, Dyson HJ, Wright PE. Structural basis for Hif-1 alpha /CBP recognition in the cellular hypoxic response. Proc Natl Acad Sci U S A 2002;99(8):5271–6. 10.1073/pnas.082121399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Freedman SJ, Sun ZY, Poy F, Kung AL, Livingston DM, Wagner G, et al. Structural basis for recruitment of CBP/p300 by hypoxiainducible factor-1 alpha. Proc Natl Acad Sci U S A 2002;99(8): 5367–72. 10.1073/pnas.082117899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Richard DE, Berra E, Gothie E, Roux D, Pouyssegur J. p42/p44 mitogen-activated protein kinases phosphorylate hypoxiainducible factor 1alpha (HIF-1alpha) and enhance the transcriptional activity of HIF-1. J Biol Chem 1999;274(46):32631–7. [DOI] [PubMed] [Google Scholar]
- 38.Jeong JW, Bae MK, Ahn MY, Kim SH, Sohn TK, Bae MH, et al. Regulation and destabilization of HIF-1alpha by ARD1-mediated acetylation. Cell 2002;111(5):709–20. [DOI] [PubMed] [Google Scholar]
- 39.Carbia-Nagashima A, Gerez J, Perez-Castro C, Paez-Pereda M, Silberstein S, Stalla GK, et al. RSUME, a small RWD-containing protein, enhances SUMO conjugation and stabilizes HIF-1alpha during hypoxia. Cell 2007;131(2):309–23. 10.1016/j.cell.2007.07.044. [DOI] [PubMed] [Google Scholar]
- 40.Cowan CA, Klimanskaya I, McMahon J, Atienza J, Witmyer J, Zucker JP, et al. Derivation of embryonic stem-cell lines from human blastocysts. N Engl J Med 2004;350(13):1353–6. 10.1056/NEJMsr040330. [DOI] [PubMed] [Google Scholar]
- 41.Wang F, Thirumangalathu S, Loeken MR. Establishment of new mouse embryonic stem cell lines is improved by physiological glucose and oxygen. Cloning Stem Cells 2006;8(2):108–16. 10.1089/clo.2006.8.108. [DOI] [PubMed] [Google Scholar]
- 42.Gibbons J, Hewitt E, Gardner DK. Effects of oxygen tension on the establishment and lactate dehydrogenase activity of murine embryonic stem cells. Cloning Stem Cells 2006;8(2):117–22. 10.1089/clo.2006.8.117. [DOI] [PubMed] [Google Scholar]
- 43.Ezashi T, Das P, Roberts RM. Low O2 tensions and the prevention of differentiation of hES cells. Proc Natl Acad Sci U S A 2005;102(13):4783–8. 10.1073/pnas.0501283102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gustafsson MV, Zheng X, Pereira T, Gradin K, Jin S, Lundkvist J, et al. Hypoxia requires notch signaling to maintain the undifferentiated cell state. Dev Cell 2005;9(5):617–28. 10.1016/j.devcel.2005.09.010. [DOI] [PubMed] [Google Scholar]
- 45.Forristal CE, Wright KL, Hanley NA, Oreffo RO, Houghton FD. Hypoxia inducible factors regulate pluripotency and proliferation in human embryonic stem cells cultured at reduced oxygen tensions. Reproduction 2010;139(1):85–97. 10.1530/REP-09-0300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ji L, Liu YX, Yang C, Yue W, Shi SS, Bai CX, et al. Self-renewal and pluripotency is maintained in human embryonic stem cells by co-culture with human fetal liver stromal cells expressing hypoxia inducible factor 1alpha. J Cell Physiol 2009;221(1):54–66. 10.1002/jcp.21826. [DOI] [PubMed] [Google Scholar]
- 47.Davy P, Allsopp R. Hypoxia: are stem cells in it for the long run? Cell Cycle 2011;10(2):206–11. 10.4161/cc.10.2.14535. [DOI] [PubMed] [Google Scholar]
- 48.Seewald MJ, Ellinghaus P, Kassner A, Stork I, Barg M, Niebrugge S, et al. Genomic profiling of developing cardiomyocytes from recombinant murine embryonic stem cells reveals regulation of transcription factor clusters. Physiol Genomics 2009;38(1):7–15. 10.1152/physiolgenomics.90287.2008. [DOI] [PubMed] [Google Scholar]
- 49.Koay EJ, Athanasiou KA. Hypoxic chondrogenic differentiation of human embryonic stem cells enhances cartilage protein synthesis and biomechanical functionality. Osteoarthr Cartil 2008;16(12):1450–6. 10.1016/j.joca.2008.04.007. [DOI] [PubMed] [Google Scholar]
- 50.Mondragon-Teran P, Tostoes R, Mason C, Lye GJ, Veraitch FS. Oxygen-controlled automated neural differentiation of mouse embryonic stem cells. Regen Med 2013;8(2):171–82. 10.2217/rme.13.12. [DOI] [PubMed] [Google Scholar]
- 51.Purpura KA, George SH, Dang SM, Choi K, Nagy A, Zandstra PW. Soluble Flt-1 regulates Flk-1 activation to control hematopoietic and endothelial development in an oxygen-responsive manner. Stem Cells 2008;26(11):2832–42. 10.1634/stemcells.2008-0237. [DOI] [PubMed] [Google Scholar]
- 52.Lee SW, Jeong HK, Lee JY, Yang J, Lee EJ, Kim SY, et al. Hypoxic priming of mESCs accelerates vascular-lineage differentiation through HIF1-mediated inverse regulation of Oct4 and VEGF. EMBO Mol Med 2012;4(9):924–38. 10.1002/emmm.201101107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.•.Tsang KM, Hyun JS, Cheng KT, Vargas M, Mehta D, UshioFukai M, et al. Embryonic stem cell differentiation to functional arterial endothelial cells through sequential activation of ETV2 and NOTCH1 signaling by HIF1alpha. Stem Cell Reports 2017;9(3):796–806. 10.1016/j.stemcr.2017.07.1. [DOI] [PMC free article] [PubMed] [Google Scholar]; This article provides evidence that HIF-1α induces mouse ESC commitment to arterial endothelia cells by activation of Etv2 and Notch1 signaling.
- 54.Sun X, Pang L, Shi M, Huang J, Wang Y. HIF2alpha induces cardiomyogenesis via Wnt/beta-catenin signaling in mouse embryonic stem cells. J Transl Med 2015;13:88 10.1186/s12967-015-0447-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sperber H, Mathieu J, Wang Y, Ferreccio A, Hesson J, Xu Z, et al. The metabolome regulates the epigenetic landscape during naiveto-primed human embryonic stem cell transition. Nat Cell Biol 2015;17(12):1523–35. 10.1038/ncb3264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006;126(4):663–76. 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 57.Yoshida Y, Takahashi K, Okita K, Ichisaka T, Yamanaka S. Hypoxia enhances the generation of induced pluripotent stem cells. Cell Stem Cell 2009;5(3):237–41. 10.1016/j.stem.2009.08.001. [DOI] [PubMed] [Google Scholar]
- 58.Folmes CD, Nelson TJ, Martinez-Fernandez A, Arrell DK, Lindor JZ, Dzeja PP, et al. Somatic oxidative bioenergetics transitions into pluripotency-dependent glycolysis to facilitate nuclear reprogramming. Cell Metab 2011;14(2):264–71. 10.1016/j.cmet.2011.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Panopoulos AD, Yanes O, Ruiz S, Kida YS, Diep D, Tautenhahn R, et al. The metabolome of induced pluripotent stem cells reveals metabolic changes occurring in somatic cell reprogramming. Cell Res 2012;22(1):168–77. 10.1038/cr.2011.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lee MR, Mantel C, Lee SA, Moon SH, Broxmeyer HE. MiR-31/ SDHA axis regulates reprogramming efficiency through mitochondrial metabolism. Stem Cell Reports 2016;7(1):1–10. 10.1016/j.stemcr.2016.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Prigione A, Rohwer N, Hoffmann S, Mlody B, Drews K, Bukowiecki R, et al. HIF1alpha modulates cell fate reprogramming through early glycolytic shift and upregulation of PDK1–3 and PKM2. Stem Cells 2014;32(2):364–76. 10.1002/stem.1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mathieu J, Zhou W, Xing Y, Sperber H, Ferreccio A, Agoston Z, et al. Hypoxia-inducible factors have distinct and stage-specific roles during reprogramming of human cells to pluripotency. Cell Stem Cell 2014;14(5):592–605. 10.1016/j.stem.2014.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Son MJ, Kwon Y, Son MY, Seol B, Choi HS, Ryu SW, et al. Mitofusins deficiency elicits mitochondrial metabolic reprogramming to pluripotency. Cell Death Differ 2015;22(12): 1957–69. 10.1038/cdd.2015.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nemeth MJ, Bodine DM. Regulation of hematopoiesis and the hematopoietic stem cell niche by Wnt signaling pathways. Cell Res 2007;17(9):746–58. 10.1038/cr.2007.69. [DOI] [PubMed] [Google Scholar]
- 65.Ng AP, Alexander WS. Haematopoietic stem cells: past, present and future. Cell Death Discov 2017;3:17002 10.1038/cddiscovery.2017.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Orkin SH, Zon LI. Hematopoiesis: an evolving paradigm for stem cell biology. Cell 2008;132(4):631–44. 10.1016/j.cell.2008.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Copelan EA. Hematopoietic stem-cell transplantation. N Engl J Med 2006;354(17):1813–26. 10.1056/NEJMra052638. [DOI] [PubMed] [Google Scholar]
- 68.Crane GM, Jeffery E, Morrison SJ. Adult haematopoietic stem cell niches. Nat Rev Immunol 2017;17(9):573–90. 10.1038/nri.2017.53. [DOI] [PubMed] [Google Scholar]
- 69.Morrison SJ, Scadden DT. The bone marrow niche for haematopoietic stem cells. Nature 2014;505(7483):327–34. 10.1038/nature12984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.••.Spencer JA, Ferraro F, Roussakis E, Klein A, Wu J, Runnels JM, et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014;508(7495):269–73. 10.1038/nature13034. [DOI] [PMC free article] [PubMed] [Google Scholar]; This article demonstrates a very low oxygen tension in the BM of live mice.
- 71.Nombela-Arrieta C, Pivarnik G, Winkel B, Canty KJ, Harley B, Mahoney JE, et al. Quantitative imaging of haematopoietic stem and progenitor cell localization and hypoxic status in the bone marrow microenvironment. Nat Cell Biol 2013;15(5):533–43. 10.1038/ncb2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Takubo K, Goda N, Yamada W, Iriuchishima H, Ikeda E, Kubota Y, et al. Regulation of the HIF-1alpha level is essential for hematopoietic stem cells. Cell Stem Cell 2010;7(3):391–402. 10.1016/j.stem.2010.06.020. [DOI] [PubMed] [Google Scholar]
- 73.Takubo K, Nagamatsu G, Kobayashi CI, Nakamura-Ishizu A, Kobayashi H, Ikeda E, et al. Regulation of glycolysis by Pdk functions as a metabolic checkpoint for cell cycle quiescence in hematopoietic stem cells. Cell Stem Cell 2013;12(1):49–61. 10.1016/j.stem.2012.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Unnisa Z, Clark JP, Roychoudhury J, Thomas E, Tessarollo L, Copeland NG, et al. Meis1 preserves hematopoietic stem cells in mice by limiting oxidative stress. Blood 2012;120(25):4973–81. 10.1182/blood-2012-06-435800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kocabas F, Zheng J, Thet S, Copeland NG, Jenkins NA, DeBerardinis RJ, et al. Meis1 regulates the metabolic phenotype and oxidant defense of hematopoietic stem cells. Blood 2012;120(25):4963–72. 10.1182/blood-2012-05-432260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Guitart AV, Subramani C, Armesilla-Diaz A, Smith G, Sepulveda C, Gezer D, et al. Hif-2alpha is not essential for cell-autonomous hematopoietic stem cell maintenance. Blood 2013;122(10):1741–5. 10.1182/blood-2013-02-484923. [DOI] [PubMed] [Google Scholar]
- 77.Rouault-Pierre K, Lopez-Onieva L, Foster K, Anjos-Afonso F, Lamrissi-Garcia I, Serrano-Sanchez M, et al. HIF-2alpha protects human hematopoietic stem/progenitors and acute myeloid leukemic cells from apoptosis induced by endoplasmic reticulum stress. Cell Stem Cell 2013;13(5):549–63. 10.1016/j.stem.2013.08.011. [DOI] [PubMed] [Google Scholar]
- 78.Scortegagna M, Morris MA, Oktay Y, Bennett M, Garcia JA. The HIF family member EPAS1/HIF-2alpha is required for normal hematopoiesis in mice. Blood 2003;102(5):1634–40. 10.1182/blood-2003-02-0448. [DOI] [PubMed] [Google Scholar]
- 79.Singh RP, Franke K, Kalucka J, Mamlouk S, Muschter A, Gembarska A, et al. HIF prolyl hydroxylase 2 (PHD2) is a critical regulator of hematopoietic stem cell maintenance during steadystate and stress. Blood 2013;121(26):5158–66. 10.1182/blood-2012-12-471185. [DOI] [PubMed] [Google Scholar]
- 80.Rankin EB, Wu C, Khatri R, Wilson TL, Andersen R, Araldi E, et al. The HIF signaling pathway in osteoblasts directly modulates erythropoiesis through the production of EPO. Cell 2012;149(1): 63–74. 10.1016/j.cell.2012.01.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kranc KR, Schepers H, Rodrigues NP, Bamforth S, Villadsen E, Ferry H, et al. Cited2 is an essential regulator of adult hematopoietic stem cells. Cell Stem Cell 2009;5(6):659–65. 10.1016/j.stem.2009.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Boitano AE, Wang J, Romeo R, Bouchez LC, Parker AE, Sutton SE, et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science 2010;329(5997):1345–8. 10.1126/science.1191536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Fares I, Chagraoui J, Gareau Y, Gingras S, Ruel R, Mayotte N, et al. Cord blood expansion. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science 2014;345(6203):1509–12. 10.1126/science.1256337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Chaurasia P, Gajzer DC, Schaniel C, D’Souza S, Hoffman R. Epigenetic reprogramming induces the expansion of cord blood stem cells. J Clin Invest 2014;124(6):2378–95. 10.1172/JCI70313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Huang X, Lee MR, Cooper S, Hangoc G, Hong KS, Chung HM, et al. Activation of OCT4 enhances ex vivo expansion of human cord blood hematopoietic stem and progenitor cells by regulating HOXB4 expression. Leukemia 2016;30(1):144–53. 10.1038/leu.2015.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Guo B, Huang X, Lee MR, Lee SA, Broxmeyer HE. Antagonism of PPAR-gamma signaling expands human hematopoietic stem and progenitor cells by enhancing glycolysis. Nat Med 2018. doi: 10.1038/nm.4477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.•.Guo B, Huang X, Cooper S, Broxmeyer HE. Glucocorticoid hormone-induced chromatin remodeling enhances human hematopoietic stem cell homing and engraftment. Nat Med 2017;23(4):424–8. 10.1038/nm.4298. [DOI] [PMC free article] [PubMed] [Google Scholar]; This article provides evidence that glucocorticoid treatment enhances human cord blood HSC homing and engraftment by epigentic regulation.
- 88.Hoggatt J, Singh P, Sampath J, Pelus LM. Prostaglandin E2 enhances hematopoietic stem cell homing, survival, and proliferation. Blood 2009;113(22):5444–55. 10.1182/blood-2009-01-201335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Capitano ML, Hangoc G, Cooper S, Broxmeyer HE. Mild heat treatment primes human CD34(+) cord blood cells for migration toward SDF-1alpha and enhances engraftment in an NSG mouse model. Stem Cells 2015;33(6):1975–84. 10.1002/stem.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Speth JM, Hoggatt J, Singh P, Pelus LM. Pharmacologic increase in HIF1alpha enhances hematopoietic stem and progenitor homing and engraftment. Blood 2014;123(2):203–7. 10.1182/blood-2013-07-516336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Forristal CE, Nowlan B, Jacobsen RN, Barbier V, Walkinshaw G, Walkley CR, et al. HIF-1alpha is required for hematopoietic stem cell mobilization and 4-prolyl hydroxylase inhibitors enhance mobilization by stabilizing HIF-1alpha. Leukemia 2015;29(6):1366–78. 10.1038/leu.2015.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nishino T, Miyaji K, Ishiwata N, Arai K, Yui M, Asai Y, et al. Ex vivo expansion of human hematopoietic stem cells by a smallmolecule agonist of c-MPL. Exp Hematol 2009;37(11):1364–77 e4. 10.1016/j.exphem.2009.09.001. [DOI] [PubMed] [Google Scholar]
- 93.••.Mantel CR, O’Leary HA, Chitteti BR, Huang X, Cooper S, Hangoc G, et al. Enhancing hematopoietic stem cell transplantation efficacy by mitigating oxygen shock. Cell 2015;161(7): 1553–65. 10.1016/j.cell.2015.04.054. [DOI] [PMC free article] [PubMed] [Google Scholar]; This article demonstrates that hypoxia collection and harvest increases HSC numbers by mitigating EPHOSS mediated by a p53CypD-MPTP axis.
- 94.Broxmeyer HE, O’Leary HA, Huang X, Mantel C. The importance of hypoxia and extra physiologic oxygen shock/stress for collection and processing of stem and progenitor cells to understand true physiology/pathology of these cells ex vivo. Curr Opin Hematol 2015;22(4):273–8. 10.1097/MOH.0000000000000144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kreso A, Dick JE. Evolution of the cancer stem cell model. Cell Stem Cell 2014;14(3):275–91. 10.1016/j.stem.2014.02.006. [DOI] [PubMed] [Google Scholar]
- 96.Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, CaceresCortes J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994;367(6464): 645–8. 10.1038/367645a0. [DOI] [PubMed] [Google Scholar]
- 97.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A 2003;100(7):3983–8. 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006;444(7120):756–60. 10.1038/nature05236. [DOI] [PubMed] [Google Scholar]
- 99.Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, et al. Identification of pancreatic cancer stem cells. Cancer Res 2007;67(3):1030–7. 10.1158/0008-5472.CAN-06-2030. [DOI] [PubMed] [Google Scholar]
- 100.Dawood S, Austin L, Cristofanilli M. Cancer stem cells: implications for cancer therapy. Oncology (Williston Park) 2014;28(12): 1101–7. 10 [PubMed] [Google Scholar]
- 101.Keith B, Johnson RS, Simon MC. HIF1alpha and HIF2alpha: sibling rivalry in hypoxic tumour growth and progression. Nat Rev Cancer 2011;12(1):9–22. 10.1038/nrc3183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Luo W, Hu H, Chang R, Zhong J, Knabel M, O’Meally R, et al. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell 2011;145(5):732–44. 10.1016/j.cell.2011.03.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Keith B, Simon MC. Hypoxia-inducible factors, stem cells, and cancer. Cell 2007;129(3):465–72. 10.1016/j.cell.2007.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Qing G, Simon MC. Hypoxia inducible factor-2alpha: a critical mediator of aggressive tumor phenotypes. Curr Opin Genet Dev 2009;19(1):60–6. 10.1016/j.gde.2008.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Vyas P Targeting HIF function: the debate continues. Blood 2014;124(24):3510–1. 10.1182/blood-2014-10-605055. [DOI] [PubMed] [Google Scholar]
- 106.Wang Y, Liu Y, Tang F, Bernot KM, Schore R, Marcucci G, et al. Echinomycin protects mice against relapsed acute myeloid leukemia without adverse effect on hematopoietic stem cells. Blood 2014;124(7):1127–35. 10.1182/blood-2013-12-544221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Coltella N, Percio S, Valsecchi R, Cuttano R, Guarnerio J, Ponzoni M, et al. HIF factors cooperate with PML-RARalpha to promote acute promyelocytic leukemia progression and relapse. EMBO Mol Med 2014;6(5):640–50. 10.1002/emmm.201303065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Velasco-Hernandez T, Hyrenius-Wittsten A, Rehn M, Bryder D, Cammenga J. HIF-1alpha can act as a tumor suppressor gene in murine acute myeloid leukemia. Blood 2014;124(24):3597–607. 10.1182/blood-2014-04-567065. [DOI] [PubMed] [Google Scholar]
- 109.Vukovic M, Guitart AV, Sepulveda C, Villacreces A, O’Duibhir E, Panagopoulou TI, et al. Hif-1alpha and Hif-2alpha synergize to suppress AML development but are dispensable for disease maintenance. J Exp Med 2015;212(13):2223–34. 10.1084/jem.20150452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Zhang H, Li H, Xi HS, Li S. HIF1alpha is required for survival maintenance of chronic myeloid leukemia stem cells. Blood 2012;119(11):2595–607. 10.1182/blood-2011-10-387381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Ng KP, Manjeri A, Lee KL, Huang W, Tan SY, Chuah CT, et al. Physiologic hypoxia promotes maintenance of CML stem cells despite effective BCR-ABL1 inhibition. Blood 2014;123(21): 3316–26. 10.1182/blood-2013-07-511907. [DOI] [PubMed] [Google Scholar]
- 112.Tanturli M, Giuntoli S, Barbetti V, Rovida E, Dello SP. Hypoxia selects bortezomib-resistant stem cells of chronic myeloid leukemia. PLoS One 2011;6(2):e17008 10.1371/journal.pone.0017008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Giambra V, Jenkins CE, Lam SH, Hoofd C, Belmonte M, Wang X, et al. Leukemia stem cells in T-ALL require active Hif1alpha and Wnt signaling. Blood 2015;125(25):3917–27. 10.1182/blood-2014-10-609370. [DOI] [PMC free article] [PubMed] [Google Scholar]