

Short abstract
Review on techniques for inhibition of MPO activity and neutrophil response after injury to COPD and/or RA.
Keywords: bioluminescence, noninvasive imaging, neutrophil chemotaxis, reactive oxygen species, reactive nitrogen species
Abstract
Myeloperoxidase aids in clearance of microbes by generation of peroxidase‐mediated oxidants that kill leukocyte‐engulfed pathogens. In this review, we will examine 1) strategies for in vitro evaluation of myeloperoxidase function and its inhibition, 2) ways to monitor generation of certain oxidant species during inflammation, and 3) how these methods can be used to approximate the total polymorphonuclear neutrophil chemotaxis following insult. Several optical imaging probes are designed to target reactive oxygen and nitrogen species during polymorphonuclear neutrophil inflammatory burst following injury. Here, we review the following 1) the broad effect of myeloperoxidase on normal physiology, 2) the difference between myeloperoxidase and other peroxidases, 3) the current optical probes available for use as surrogates for direct measures of myeloperoxidase‐derived oxidants, and 4) the range of preclinical options for imaging myeloperoxidase accumulation at sites of inflammation in mice. We also stress the advantages and drawbacks of each of these methods, the pharmacokinetic considerations that may limit probe use to strictly cell cultures for some reactive oxygen and nitrogen species, rather than in vivo utility as indicators of myeloperoxidase function. Taken together, our review should shed light on the fundamental rational behind these techniques for measuring myeloperoxidase activity and polymorphonuclear neutrophil response after injury toward developing safe myeloperoxidase inhibitors as potential therapy for chronic obstructive pulmonary disease and rheumatoid arthritis.
Abbreviations
- 5‐HT
5‐hydroxytryptamide
- ADHP
10‐acetyl‐3,7‐dihydroxyphenoxazine
- APF
2‐[6‐(4′‐amino)phenoxy‐3H‐xanthen‐3‐on‐9‐yl]benzoic acid
- BODIPY
boron‐dipyrromethene
- CPH
1‐hydroxy‐3‐carboxy‐2,2,5,‐tetramethyl‐pyrrolidine hydrochloride
- DEPMO
5‐(diethoxyphosphoryl)‐5‐methyl‐1‐pyrroline N‐oxide
- DHR123
dihydrorhodamine 123
- DMPO
5,5‐dimethyl‐1‐pyrroline N‐oxide
- EC
endothelial cell
- EPO
eosinophil peroxidase
- EPR
electron paramagnetic resonance
- Fe
iron
- Gd
gadolinium
- H2DCFDA
2′,7′‐dichlorodihydrofluorescein diacetate
- H2O2
hydrogen peroxide
- HC
heavy chain of myeloperoxidase
- HOBr
hypobromous acid
- HOCl
hypochlorous acid
- HPF
2‐[6‐(4′‐hydroxy)phenoxy‐3H‐xanthen‐3‐on‐9‐yl]benzoic acid
- LPO
lactoperoxidase
- MPO
myeloperoxidase
- NO
nitric oxide radical
- O2
superoxide radical, −
- OCl
hypochlorite

hydroxyl radical
- ONOO−
peroxynitrite
- PDB
Protein Data Bank
- PMN
polymorphonuclear neutrophil
- RA
rheumatoid arthritis
- ROS
reactive oxygen species
- RNS
reactive nitrogen species
PHYSIOLOGIC IMPORTANCE OF MPO FUNCTION
MPO, a heme‐containing peroxidase in myeloid cells (PMNs and monocytes), has a central role in the development of the nascent inflammatory response and the perpetuation of chronic inflammation in diseases such as RA. From the initial trigger (i.e., damage or infection), patrolling monocytes [1], mast cells [2], and dendritic cells [3] respond to the acute injury almost immediately by increasing vascular permeability through histamine release and simultaneous establishment of chemokine and chemoattractant gradients required to arrest circulating leukocytes (see Fig. 1 ). A hallmark PMN burst occurs when responding leukocytes crawl to a slow roll through interaction of leukocyte‐derived L‐selectin and endothelial E‐selectin and P‐selectin [4]. In the mouse, PMN and other leukocytes essentially stop as the burden of the drag exerted on the cell overcomes blood flow pressures through, in part, binding of LFA‐1 and Mac1 (CD11b) with newly exposed ICAM on the activated EC surface (ECICAM‐1). This progress occurs near the damage and just before diapedesis [5]. Once at the inflammatory site, PMN, and later PMN‐signaled monocytes, begin the task of clearing foreign material (i.e., microbes) or cell debris [4]. To combat pathogens in the area, PMNs ensnare microbes through exocytosis of chromosomal material called neutrophil extracellular traps enabling phagocytosis of ensnared pathogens by neighboring PMNs [6, 7]. At the same time, there is probably an increased extracellular concentration of MPO in the local microenvironment as a direct result of MPO contributed by apoptotic PMNs that have sought to ensnare the microbes or debris in their chromosomal neutrophil extracellular traps. Because MPO makes up 30% of the total cellular PMN protein content, the resulting HOCl wave should effectively sanitize the area [8]. PMNs also experience an influx of K+ [9] and an increased pH [10], which awakens dormant proteases [11]. Given this, the engulfed pathogens ultimately face harsh pH conditions in the vacuoles [12], degradation of cell wall components by PMN proteases (i.e., cathepsin G and elastase), and oxidation of DNA and proteins by the powerful HOCl oxidant generated by the prolific MPO‐H2O2 system [13].
Figure 1.

Biology of PMNs in response to injury and inflammation. Localized overexpression of E‐selectin and P‐selectin on activated endothelial cells slow the PMN roll upon the endothelium via leukocyte‐derived L‐selectin. Responding PMNs transmigrate through the endothelial cells after LFA‐1 hooks intercellular adhesion molecule 1 (ICAM‐1) and arrive at the site of damage just before diapedesis caused by the increased vascular permeability from histamine released from the mast cells. PMNs undergo phagocytosis of the invasive microbes once they arrive at the infection site. In addition, MPO is secreted from the patrolled PMNs to produce the potent antimicrobial reagent HOCl in response to infection.
In humans, MPO has become a biomarker for heightened inflammation because of its role in the production of oxidized DNA and protein adducts [14]. For example, ROS and RNS production during oxidative stress has been linked to heightened MPO levels in chronic obstructive pulmonary disease [15], RA [16], atherosclerosis [17, 18, 19, 20–21], and even some cancers [22, 23]. Therefore, MPO is considered a druggable target with small‐molecule inhibitors already identified and more expected for limiting ROS/RNS production. In preclinical study, several MPO inhibitors have limited inflammation in diseases, including thioxanthine‐2 in chronic obstructive pulmonary disease studies [15], PF‐1355 in vasculitis and glomerulonephritis [24], and INV315 in atherosclerosis [25]. In this review, we will mainly discuss the recent, emerging tools for in vivo assessment of MPO as a target with the goal of monitoring disease progression, tracking PMN migration, and detecting inflammatory sites for preclinical or clinical applications.
WHAT SETS MPO APART FROM OTHER PEROXIDASES?
It is generally accepted that there are 2 distinct peroxidase superfamilies. One superfamily comprises all plant, fungal, and bacterial peroxidases, and the other comprises the mammalian peroxidases [26]. One primary difference between mammalian peroxidases (e.g., MPO, LPO, EPO, and thyroid peroxidase) and nonmammalian peroxidases (e.g., horseradish peroxidase, bacterial catalase‐peroxidase, lignin peroxidase, and ascorbate peroxidase) is the presence of specific linkages between the respective heme prosthetic group and the main peroxidase protein scaffold [26]. There are generally 2 ester bond linkages, but in the case of MPO, the presence of an additional unique sulfonium linkage distorts the heme from its normal planar state and allows MPO to produce HOCl.
Generally, all peroxidases have an active‐site heme group that contains a central Fe atom that maintains its oxidation state (i.e., reactivity) through coordination with a distal His residue. MPO has a ferric heme, MPO‐Fe(III), which is oxidized to a short‐lived intermediate, termed Compound I (half life ∼100 ms [27]), by reacting with a 100–10,000‐fold lower relative concentration of H2O2, leading to generation of a ferryl porphyrin π cation radical, MPO‐FE(IV) = O·+π (Equation 1) [28, 29]. MPO can regenerate its ferric state in 2 consecutive 1‐electron steps with a classic peroxide electron donor (AH2) within a typical peroxidase catalytic cycle (Equations 2–3). The porphyrin radical is reduced to a ferryl heme, known as Compound II, in the first step (Equation 2). Compound II then is reduced back to ferric enzyme A by AH2 in the following step (Equation 3). At the same time, AH2 is oxidized to the free radical (AH) [30, 31].
 |
(1) |
 |
(2) |
| (3) |
| (4) |
In the chlorination cycle of MPO, Cl− ions are uniquely oxidized to HOCl by Compound I, and in the process, the MPO ferric state is regenerated (Equation 4).
Several MPO crystal structures have been solved that demonstrate complex formations between the MPO enzyme and a variety of ligands and inhibitors, which has provided a wealth of insight into MPO catalysis and inhibitor binding. Substrates gain access to the proximal surface of the cavern, wherein the heme prosthetic group acts as floor of the MPO active site. A narrower substrate‐binding chase leads from the enzyme and can be seen in the MPO isoform C structure (see PDB accession number 1LXP), MPO cocrystallized with cyanide and thiocyanate (1DNW), the complex of MPO with cyanide (1DNU), the thioxanthine‐inhibited MPO (3ZS1), the ceruloplasmin‐bound MPO (4EJX), and the MPO with aromatic hydroxamates (4C1M) [32, 33, 34, 35, 36, 37–38]. As mentioned, the protoporphyrin IX macrocycle of MPO has 3 covalent bonds with the parent protein as shown in Fig. 2 , and the planar distortion of the heme leads to the distant spectral signature of MPO [26]. These linkages correspond to the following: 1) an ester bond between the light chain of the myeloperoxidase Asp94 residue of the MPO light chain and the methyl side chain of pyrrole C, 2) another ester bond between glutamate on the heavy chain (HCGlu242) and the methyl side chain of pyrrole A, and 3) a vinyl sulfonium linkage between a heavy chain methionine (HCMet243) and pyrrole A [26].
Figure 2.

Surface representation of the active site of the MPO monomer. PDB numbers: 3F9P, the light chain is shown in pink, and the heavy chain is shown in blue. There are 3 linkages between the heme and protein through Asp94 on the light chain and Glu242 and Met243 on the heavy chain (highlighted as yellow). The catalytically essential residues Gln91, His95, and Arg239 (highlighted as red) compose the distal heme pocket; in the proximal site, the heme iron is coordinated through the His336 imidazole residue (highlighted as turquoise).
Classic biochemical studies [26, 39] have proven this vinyl sulfonium bond between MPO and the pyrrole A causes a distinct Soret (also known as an absorbance) band formation at 430 nm. Met243 variants of MPO have diminished peroxidase activity compared with native MPO, and site‐directed disruption of this linkage causes the Soret band at 430 nm to be lost in favor of a 413‐nm band, similar to that in absorbance spectra of LPO [26, 39]. Other Soret bands are responsible for the green coloration of the MPO protein, namely bands at 496, 570, 620, and 690 nm [40]. Our recent study found that some potent MPO inhibitors, like benzoic acid hydrazide and its analogs, cause disruption of these important ester linkages [41, 42]. In mammalian peroxidase, these linkages are thought to impart a level of resistance to the powerful oxidants that they produce, namely HOCl and HOBr, for MPO and LPO, respectively [43].
DETECTION OF MPO‐DERIVED ROS/RNS IN ISOLATED CELLS AND IN VITRO ASSAYS
To study MPO in any complex mixture, there must be an effective means to differentiate between MPO‐dependent ROS/RNS products and other reactive products. As such, a number of sensors have been developed to meet this need, such as OH, singlet oxygen, O2 −, ONOO−, and −OCl. A summary of these sensors is shown in Table 1 . This section will focus on introducing most of these sensors, attributing unique properties that drive their utility, and imparting their relative limitations.
Table 1.
A summary of the sensors for detection of MPO‐derived oxidants and other ROSs/RNSs
| Modality/ Agent | MW (Da) | Excitation λ (nm) | Emission λ (nm) | Target | Cell culture (in vitro) | Preclinical (in vivo) |
|---|---|---|---|---|---|---|
| EPR | ||||||
| DMPO | 113.2 | — | — |
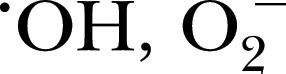
|
✓ | × |
| DEPMPO | 235.2 | — | — |
 , , 
|
✓ | × |
| CMH | 201.3 | — | — | ONOO−, 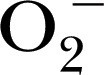
|
✓ | × |
| CPH | 223.7 | — | — | ONOO−, 
|
✓ | × |
| mito‐TEMPO‐H | 511.1 | — | — |
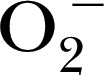
|
✓ | × |
| DBNBS | 367.0 | — | — |
 , ONOO−, , ONOO−, 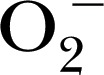 , MPO , MPO |
✓ | × |
| Fluorescence | ||||||
| APF | 423.4 | 490 | 515 |
 , ONOO−, and HOCl/Br , ONOO−, and HOCl/Br |
✓ | × |
| HPF | 424.4 | 490 | 515 |
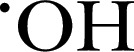 , ONOO− , ONOO−
|
✓ | × |
| DAF‐FM | 412.4 | 495 | 515 |
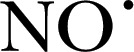
|
✓ | × |
| H2 DCFDA | 487.3 | 495 | 527 |
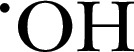 , ROO⋅, ONOO−, HOCl , ROO⋅, ONOO−, HOCl |
✓ | × |
| HKGreen‐3 | 613.1 | 520 | 535 | ONOO− | ✓ | × |
| mito‐SOX | 759.7 | 510 | 580 |
 , heme peroxidases , heme peroxidases |
✓ | × |
| DHR123 | 346.4 | 500 | 536 | ONOO−,  , HOCl , HOCl |
✓ | × |
| HKOCl‐1 | 512.2 | 520 | 541 | HOCl | ✓ | × |
| DHE | 315.4 | 518 | 605 |
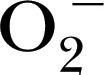 , heme peroxidases , heme peroxidases |
✓ | × |
| ADHP | 257.2 | 571 | 585 | Heme peroxidases | ✓ | ✓ |
| SNAPF | 640.0 | 614 | 676 | HOCl | ✓ | ✓ |
| Quenched nanoparticle | ||||||
| Oxazine nanoparticle | 423.3 (oxazine) | 620 | 672 | ONOO−, HOCl | ✓ | ✓ |
| LS601R‐PEG40 | 41,000 | 785 | 810 |
 , , 
|
✓ | ✓ |
| Lucigenin | 510.5 | — | 470 |
 and H2O2 and H2O2
|
✓ | ✓ |
| BLI | ||||||
| Luminol | 177.2 | — | 425 |
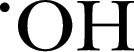 , ONOO−, , ONOO−, 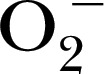 , HOCl and HOBr , HOCl and HOBr |
✓ | ✓ |
| L‐012 | 288.7 | — | 425 |
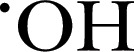 , ONOO−, HOCl and HOBr , ONOO−, HOCl and HOBr |
✓ | ✓ |
| MCLA | 291.7 | — | 465 |
 , MPO‐derived oxidants, ONOO− , MPO‐derived oxidants, ONOO−
|
✓ | ✓ |
| Pholasin | 34,600 | — | Blue light |
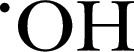 , ONOO−, , ONOO−, 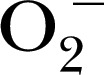 and MPO and MPO |
✓ | ✓ |
| CRET | ||||||
| Luminol‐QD800 | N/A | — | 800 | MPO‐derived HOCl | ✓ | ✓ |
| MCLA‐ BP‐AF594 | N/A | — | 620 |
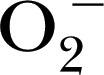 , and MPO‐derived oxidants , and MPO‐derived oxidants |
✓ | ✓ |
| FRET | ||||||
| HyPer sensor | 52,000 | 420/500 | 535 | H2O2 | ✓ | ✓ |
| Photo‐acoustic | ||||||
| NIR light‐absorbing RSPN1 | 553.7 (IR775S) | 680 | 838 | ONOO−, HOCl | ✓ | ✓ |
| MRI | ||||||
| Gd‐bis‐5‐HT‐DTPA/Gd‐ 5‐HT‐DOTA | 707.2, 788.7 | — | — | MPO | ✓ | ✓ |
Abbreviations: BLI, bioluminescence imaging; CMH, 1‐hydroxy‐3‐methoxycarbonyl‐2,2,5,5‐tetramethylpyrrolidine; CRET, chemiluminescence resonance energy transfer; DAF‐FM, 4‐amino‐5‐methylamino‐ 2′,7′‐difluorescein; DBNBS, 3,5‐dibromo‐4‐nitrosobenzenesulfonic acid; DHE, dihydroethidium; DTPA, diethylenetriaminepentaacetic acid; SPN, semiconducting polymer nanoparticle; mito‐TEMPO‐H, 1‐hydroxy‐4‐[2‐triphenylphosphonio)‐acetamido]‐2,2,6,6‐tetramethylpiperidine; N/A, not applicable.
Generally, the cellular redox homeostasis is maintained by a series of checks and balances between the production of ROS/RNS and the antioxidant system, wherein superoxide dismutase produces H2O2 from 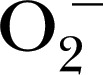 , and this is counteracted by the activity of catalase that degrades H2O2 into H2O. Other antioxidants act as general scavengers, such as NADPH, glutathione, thioredoxin, and peroxiredoxin [44], which further modulate the levels of certain oxidants. Up‐regulated NADPH oxidases increase the relative
, and this is counteracted by the activity of catalase that degrades H2O2 into H2O. Other antioxidants act as general scavengers, such as NADPH, glutathione, thioredoxin, and peroxiredoxin [44], which further modulate the levels of certain oxidants. Up‐regulated NADPH oxidases increase the relative  levels, contributing to certain pathologies, such as cardiac hypertrophy, fibrosis, ischemic stroke, and neurodegenerative diseases [45]. Central to this and important to the discussion here is that MPO functions to generate the potent oxidants HOCl from Cl− and H2O2, while generating other ROSs (e.g.,
levels, contributing to certain pathologies, such as cardiac hypertrophy, fibrosis, ischemic stroke, and neurodegenerative diseases [45]. Central to this and important to the discussion here is that MPO functions to generate the potent oxidants HOCl from Cl− and H2O2, while generating other ROSs (e.g., 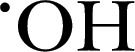 , singlet oxygen, and ONOO−) [46].
, singlet oxygen, and ONOO−) [46].
Given the relative importance of the redox state of the catalytic Fe in MPO, the use of EPR or spin resonance spectroscopy has become a sensitive method for in vitro assessment of the inhibitor‐ or substrate‐altered redox state [47]. Some probes for this application include DMPO for trapping 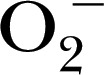 [48], DEPMPO for MPO and MPO‐derived oxidants [49], 1‐hydroxy‐3‐methoxycarbonyl‐2,2,5,5‐tetramethylpyrrolidine for the spin trapping of ONOO− and
[48], DEPMPO for MPO and MPO‐derived oxidants [49], 1‐hydroxy‐3‐methoxycarbonyl‐2,2,5,5‐tetramethylpyrrolidine for the spin trapping of ONOO− and 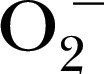 [48], CPH for stable trapping
[48], CPH for stable trapping 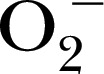 [47], 1‐hydroxy‐4‐[2‐triphenylphosphonio)‐acetamido]‐2,2,6,6‐tetramethylpiperidine for detection of mitochondrial
[47], 1‐hydroxy‐4‐[2‐triphenylphosphonio)‐acetamido]‐2,2,6,6‐tetramethylpiperidine for detection of mitochondrial 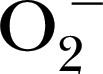 [50], and 3,5‐dibromo‐4‐nitrosobenzenesulfonic acid for trapping
[50], and 3,5‐dibromo‐4‐nitrosobenzenesulfonic acid for trapping  , ONOO−, and
, ONOO−, and 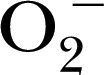 in biologic systems [51]. The distinct advantage of the CPH probe is that it generates a more‐stable
in biologic systems [51]. The distinct advantage of the CPH probe is that it generates a more‐stable 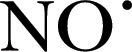 with a longer half‐life, which provides for a better detection window, whereas the use of nitrone spin traps, such as DMPO and DEPMPO, produce the more‐unstable intermediate
with a longer half‐life, which provides for a better detection window, whereas the use of nitrone spin traps, such as DMPO and DEPMPO, produce the more‐unstable intermediate 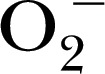 [47]. In addition, CPH probes are membrane permeable and can provide insights into the generation of ROS across both extracellular and intracellular membranes, allowing cytosol concentrations to be compared with those in the mitochondria [52].
[47]. In addition, CPH probes are membrane permeable and can provide insights into the generation of ROS across both extracellular and intracellular membranes, allowing cytosol concentrations to be compared with those in the mitochondria [52].
There are also cell‐permeable, fluorogenic ROS/RNS sensors, such as H2DCFDA and DHR123, which are oxidized by  , ONOO−, and −OCl [53]. H2DCFDA is converted into 2′,7′‐dichlorofluorescin by intracellular esterases within the cell [54]. Apart from the relative nonselectivity to ROS species and oxidants, both cytochrome c and heme peroxidases catalyze the oxidation of 2′,7′‐dichlorofluorescein and DHR123 [55].
, ONOO−, and −OCl [53]. H2DCFDA is converted into 2′,7′‐dichlorofluorescin by intracellular esterases within the cell [54]. Apart from the relative nonselectivity to ROS species and oxidants, both cytochrome c and heme peroxidases catalyze the oxidation of 2′,7′‐dichlorofluorescein and DHR123 [55]. 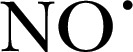 is able to oxidize another fluorescein‐based probe 4‐amino‐5‐methylamino‐ 2′,7′‐difluorescein to the fluorescent dye benzotriazole via oxidative cycloaddition reactions [56, 57]. The hydroethidine‐based probe mito‐SOX can detect
is able to oxidize another fluorescein‐based probe 4‐amino‐5‐methylamino‐ 2′,7′‐difluorescein to the fluorescent dye benzotriazole via oxidative cycloaddition reactions [56, 57]. The hydroethidine‐based probe mito‐SOX can detect 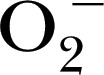 in the mitochondria, whereas dihydroethidium can detect
in the mitochondria, whereas dihydroethidium can detect  generally within cells [58]. These probes are not directly used for the detection of MPO‐derived products but can be used to confirm the oxidant generated or present in a complex mixture.
generally within cells [58]. These probes are not directly used for the detection of MPO‐derived products but can be used to confirm the oxidant generated or present in a complex mixture.
Other ROS, cell‐permeable, fluorogenic probes include APF and HPF, which undergo O‐dearylation oxidation to form highly fluorescence products in the presence of 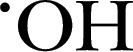 , ONOO−, and −OCl [57, 59]. HPF can selectively detect the presence of
, ONOO−, and −OCl [57, 59]. HPF can selectively detect the presence of 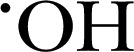 and ONOO−, whereas −OCl can also oxidize APF. MPO‐derived HOCl and EPO‐derived HOBr were specifically detected in PMNs and eosinophils by the combination of APF and HPF, respectively [59, 60]. Other oxidation‐mediated probes include HKGreen‐3, which detects ONOO− in vitro or in RAW 264.7 macrophages, generating fluorescent N‐methylrhodol by an N–dearylation reaction [61]. Similarly, a BODIPY‐based fluorescent probe, HKOCl‐1, detects HOCl from the formation of benzoquinone from p‐methoxyphenol [61, 62]. A red‐shifted fluorogenic sensor of peroxidase activity is ADHP, also known as Amplex Red, which has a strict H2O2 dependency, allowing H2O2 to be used as a trigger for inhibitor studies, whereby the enzyme and the compound can be allowed to reach equilibrium before initiation of the chemical reaction (
Fig. 3A
). Previously, we used stopped flow spectroscopy and the ADHP substrate to study MPO inhibition by benzoic acid hydrazide and its analogs [41, 42]. ADHP has also been used in tissue‐based assays for determining the presence of MPO activity, as a surrogate for assessing inflammation [63]. There is evidence that ADHP can undergo autoradical formation under prolonged exposure to excitation energy [58]. The BODIPY‐based fluorescent probe HKOCl‐1, specific for the detection of MPO‐derived HOCl, has never, to our knowledge, been used apart from the original reported study. Significant effort exists toward adapting these probes for use in intravital PMN recruitment and transmigration studies [64, 64–66].
and ONOO−, whereas −OCl can also oxidize APF. MPO‐derived HOCl and EPO‐derived HOBr were specifically detected in PMNs and eosinophils by the combination of APF and HPF, respectively [59, 60]. Other oxidation‐mediated probes include HKGreen‐3, which detects ONOO− in vitro or in RAW 264.7 macrophages, generating fluorescent N‐methylrhodol by an N–dearylation reaction [61]. Similarly, a BODIPY‐based fluorescent probe, HKOCl‐1, detects HOCl from the formation of benzoquinone from p‐methoxyphenol [61, 62]. A red‐shifted fluorogenic sensor of peroxidase activity is ADHP, also known as Amplex Red, which has a strict H2O2 dependency, allowing H2O2 to be used as a trigger for inhibitor studies, whereby the enzyme and the compound can be allowed to reach equilibrium before initiation of the chemical reaction (
Fig. 3A
). Previously, we used stopped flow spectroscopy and the ADHP substrate to study MPO inhibition by benzoic acid hydrazide and its analogs [41, 42]. ADHP has also been used in tissue‐based assays for determining the presence of MPO activity, as a surrogate for assessing inflammation [63]. There is evidence that ADHP can undergo autoradical formation under prolonged exposure to excitation energy [58]. The BODIPY‐based fluorescent probe HKOCl‐1, specific for the detection of MPO‐derived HOCl, has never, to our knowledge, been used apart from the original reported study. Significant effort exists toward adapting these probes for use in intravital PMN recruitment and transmigration studies [64, 64–66].
Figure 3.
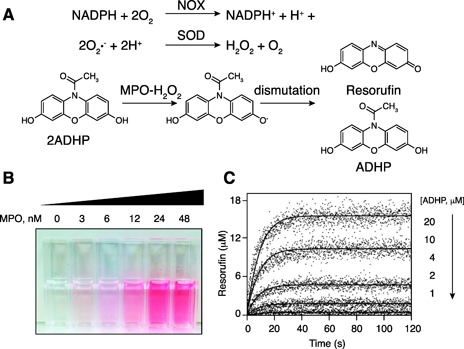
Fluorescence reaction for measuring MPO activity. (A) General mechanism of ADHP oxidation in a proposed 2‐step reaction, whereby the MPO–H2O2 complex generates 2 ADHP radicals that undergo a subsequent enzyme‐independent dismutation reaction to complete formation of 1 resorufin and 1 ADHP molecule. (B) Classic biochemical assays are possible using absorbance changes caused by the MPO–H2O2 system and the use of new fluorogenic probes, such as ADHP. Pictures are cuvettes containing increasing concentrations of the MPO–H2O2 system, as indicated, at a static ADHP level (40 μM). (C) Stopped‐flow progress curves of resorufin generation by MPO (23 nM) initiated by the addition of H2O2 (22 μM) for a series of given ADHP concentrations (adapted from Huang et al. [41]).
STRATEGIES FOR IMAGING MPO ACTIVITY IN PRECLINICAL MOUSE MODELS
Biodistribution studies indicate that there are static spatial and temporal differences in expression of mammalian peroxidases throughout host cells and tissues. Mobile immune cells are also an excellent source of stored peroxidases because these enzymes are sequestered in granules, as is the case for MPO and EPO [67]. In response to stimuli, PMNs flood injured tissue with MPO to catalyze the production of the disinfectant HOCl. Quantification of MPO levels can indicate overall inflammatory burden. Chronic inflammation leads to prolonged signal production and constant cellular recruitment, and, as such, both plasma and synovial fluids taken from patients with RA have increased MPO levels compared with healthy controls [16]. In addition, genetic production of MPO deficient‐mice attenuated RA severity in mouse models without altering circulating cytokine levels [68], indicating that MPO is a critical mediator of joint inflammation and damage in experimental RA. Cytokine levels are not altered in these mice, which may show the lack of MPO‐dependent feedback signaling, but that is conjecture at this stage.
Although use of the aforementioned ROS/RNS probes seems compelling, their hydrodynamic and pharmacokinetic properties limit their utility in animal studies. Their narrow imaging window is driven by their enhanced washout kinetics and reduced circulation half‐life. For example, solid tumors and regions of inflammation are known to be hyperpermeable to microparticles and nanoparticulates via enhanced permeability and retention properties [69]. Currently, there are already several in vivo imaging methods available, such as MRI of MPO‐induced oligomerization of chelated Gd containing serotonin analogs [70], fluorescence imaging of MPO‐induced release of oxazine probes from a nanoparticle scaffold [71], and MPO‐induced chemiluminescence using luminol (Sigma‐Aldrich, St. Louis, MO, USA) [72, 73]. Previously, bioluminescence imaging of MPO activity was successfully conducted in spontaneous, large, granular lymphocytic tumors in Gzmb:Tax mice with intraperitoneal administration of luminol [72]. On the surface, these results are not surprising because PMNs infiltrated these tumors. This “pseudo” in vivo Western blot is an interesting tool for studies of MPO activity but is limited to superficial sites because of photon scattering and the absorptive properties of blood and tissue [74]. This methodology is employed in Fig. 4 . In this example, intraperitoneal injection of luminol was used to detect MPO activity in inflammation sites associated with an acute cutaneous infection (Fig. 4B). This method has some limitations in that luminol has an extremely short half‐life (20 min) in mice [72] and is 90% cleared by the kidney and excreted through the urine [75]. Another limitation is that this imaging method can only be used as a paninflammation‐type sensor because it does not discriminate between sterile inflammation and active infection [76]. This process is highly H2O2‐dependent, and without an optimal H2O2 concentration available to MPO, the light production is severely affected (Fig. 4C).
Figure 4.
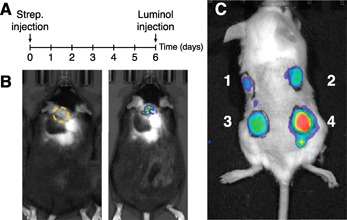
In vivo monitoring of MPO activity in mice. (A) Experimental design of the imaging study is shown. (B) A representative image of a mouse 6 d after s.c. injection of Streptococcus pyogenes (3 × 1010 CFU in phosphate‐buffered saline) in the upper back of the animal shown in white light (right image) and bioluminescence imaging (left image). A yellow dotted circle indicates the region of interest. (C) An example of the hydrogen peroxide (H2O2)‐dependence of MPO in vivo activity is shown. Albino C57BL/6 mice injected with 180 nM of MPO with either 400 μM of H2O2 (site 1), 2 mM of H2O2 (site 2), 4 mM of H2O2 (site 3), and 40 mM of H2O2 (site 4) were added to the matrigel (100 μl) before s.c. implant. (B–C) To visualize the MPO activity, an i.p. injection of luminol (Sigma‐Aldrich, 3 mg/kg) was given 10 min before luminescence imaging using the IVIS Lumina XRMS system (PerkinElmer, Waltham, MA, USA). The Institutional Animal Care and Use Committee of Auburn University approved all animal procedures.
To overcome the depth limitation of the luminol‐based and normal fluorescent reflectance methods, hybrid techniques were developed that combines a near‐infrared–emitting quantum dot, tethered to a light‐production engine, by way of the MPO‐mediated oxidation of the luminol payload [73]. This probe, in an inflammatory environment, allows for bioluminescence‐resonance energy transfer to occur between the oxidized luminol and the probe. Another option available for in vivo probe sensors is to push to longer wavelengths to abrogate tissue interference, which was the case for the APF‐derived, near‐infrared dye sulfonaphthoaminophenyl fluorescein, used to assess HOCl production from human whole blood and advanced atherosclerotic plaques [77]. Previously, we were also able to modulate the poor pharmacokinetics of these small‐molecule ROS/RNS probes by attaching an activatable, near‐infrared oxazine probe to the surface of an iron‐oxide nanoparticle (increasing the half‐life to ∼9 h) [71]. In this case, the probe was oxidized and released from the particle only in the presence of HOCl or peroxynitrite [71]. Another report of similar conceptual design is the quenched nanoparticle LS601R‐PEG40, a nonfluorescent hydrocyanine probe conjugated with PEG40, which can be oxidized to fluorescent LS601 in the presence of various ROSs [78]. Shifts are also being seen in the in vivo methods used to collect this ROS/RNS information, namely, the advent of the photoacoustic or optoacoustic imaging modalities. One such probe used in this way was IR775S, a cyanine dye derivative sensitive to ONOO− and HOCl oxidation and used in a variety of inflammatory states [79].
Another option used in the preclinical setting is MRI, which allows for the capture of noninvasive, anatomic information, while gradient modulation can lead to differential relaxation profiles and probe visualization. An example of this is the MPO‐dependent polymerization of 5‐HT chelated to Gd, namely Gd‐bis‐5‐HT‐diethylenetriaminepentaacetic acid [70, 80]. As a polymer, the sensor has an enhanced magnetic resonance signal, which is prolonged up to 120 mins with the MPO‐Gd probe in matrigel experiments, mice brains, and the infarct zone in mice [81, 82, 83, 84–85]. This probe has been used extensively for in vivo inflammatory models that have been established in such a way as to prove the MPO‐dependent nature of the reporter using MPO‐deficient mice [81, 82–83].
WHAT DOES THE FUTURE HOLD FOR MONITORING MPO ACTIVITY AND PMN RECRUITMENT NONINVASIVELY?
New, emerging technology will allow for the development of better optical ROS/RNS sensors that take advantage of the spectrum of near‐infrared wavelengths to limit potential interference and scatter from blood and tissue during in vivo animal imaging studies. To complement this, there are ongoing attempts to develop the next generation of probes for in vivo imaging of MPO, which, hopefully, will have better pharmacodynamic properties for in vivo work to improve the detectability of MPO by increasing the signal‐to‐noise ratio. Besides new probe chemistry, improved imaging hardware has already begun to provide better resolution for researchers using optical imaging, especially with the advent of photoacoustic/optoacoustic techniques [86, 87–88]. Distinct from fluorescence, strong absorbing dyes are desired because the thermal expansion of the probe causes a resultant and proportional ultrasound signal [89, 90]. The benefit of these technologies over the standard reflectance‐fluorescence imaging is that they reduce the signal scattering as the light passes through tissue because sound waves are used instead of light for the later portion of the imaging strategy [91].
CONCLUDING REMARKS
Imaging MPO activity and assessing the imaging of this important peroxidase remains an active area of research because of its central role in inflammation. In designing new MPO inhibitors, the physiologic role of the enzyme, the elements that lead to HOCl generation, the gamut and specificity of secondary ROS/RNS reporter probes, and the lead compounds that ultimately cause toxicity in vivo must be understood. Often the best inhibitor for MPO is not the safest (e.g., sodium azide is a potent MPO inhibitor but toxic when applied to an animal). Taken together, a more‐holistic approach can be implemented, which would provide a streamlined approach for screening positive traits to accelerate new anti‐inflammatory drug designs.
AUTHORSHIP
J.H., A.M., J.R.P., and P.P. wrote this review. R.D.A., H.H., and F.S. edited the review and made critical comments throughout. J.H., A.M., and P.P. performed the experiments needed to demonstrate the luminol‐based detection of MPO activity.
DISCLOSURES
The authors declare no conflicts of interest.
ACKNOWLEDGMENTS
Supported, in part, by the U.S. National Institutes of Health through grants provided by the National Heart, Lung, and Blood Institute (Grant R01HL114477 to P.P.), the National Institute of Biomedical Engineering and Imaging (Grant R01EB EB016100 to R.D.A.), and Auburn University Research Initiative Cancer Graduate Fellowship (to J.H.).
REFERENCES
- 1. Auffray, C. , Fogg, D. , Garfa, M. , Elain, G. , Join‐Lambert, O. , Kayal, S. , Sarnacki, S. , Cumano, A. , Lauvau, G. , Geissmann, F. (2007) Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 317, 666–670. [DOI] [PubMed] [Google Scholar]
- 2. Theoharides, T. C. , Alysandratos, K. D. , Angelidou, A. , Delivanis, D. A. , Sismanopoulos, N. , Zhang, B. , Asadi, S. , Vasiadi, M. , Weng, Z. , Miniati, A. , Kalogeromitros, D. (2012) Mast cells and inflammation. Biochim. Biophys. Acta 1822, 21–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Palucka, K. , Banchereau, J. (2012) Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 12, 265–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Theilgaard‐Mönch, K. , Porse, B. T. , Borregaard, N. (2006) Systems biology of neutrophil differentiation and immune response. Curr. Opin. Immunol. 18, 54–60. [DOI] [PubMed] [Google Scholar]
- 5. Diacovo, T. G. , Roth, S. J. , Buccola, J. M. , Bainton, D. F. , Springer, T. A. (1996) Neutrophil rolling, arrest, and transmigration across activated, surface‐adherent platelets via sequential action of P‐selectin and the beta 2‐integrin CD11b/CD18. Blood 88, 146–157. [PubMed] [Google Scholar]
- 6. Brinkmann, V. , Reichard, U. , Goosmann, C. , Fauler, B. , Uhlemann, Y. , Weiss, D. S. , Weinrauch, Y. , Zychlinsky, A. (2004) Neutrophil extracellular traps kill bacteria. Science 303, 1532–1535. [DOI] [PubMed] [Google Scholar]
- 7. Branzk, N. , Lubojemska, A. , Hardison, S. E. , Wang, Q. , Gutierrez, M. G. , Brown, G. D. , Papayannopoulos, V. (2014) Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat. Immunol. 15, 1017–1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Parker, H. , Albrett, A. M. , Kettle, A. J. , Winterbourn, C. C. (2012) Myeloperoxidase associated with neutrophil extracellular traps is active and mediates bacterial killing in the presence of hydrogen peroxide. J. Leukoc. Biol. 91, 369–376. [DOI] [PubMed] [Google Scholar]
- 9. Reeves, E. P. , Lu, H. , Jacobs, H. L. , Messina, C. G. , Bolsover, S. , Gabella, G. , Potma, E. O. , Warley, A. , Roes, J. , Segal, A. W. (2002) Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature 416, 291–297. [DOI] [PubMed] [Google Scholar]
- 10. Segal, A. W. , Geisow, M. , Garcia, R. , Harper, A. , Miller, R. (1981) The respiratory burst of phagocytic cells is associated with a rise in vacuolar pH. Nature 290, 406–409. [DOI] [PubMed] [Google Scholar]
- 11. Segal, A. W. (2005) How neutrophils kill microbes. Annu. Rev. Immunol. 23, 197–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Levine, A. P. , Duchen, M. R. , de Villiers, S. , Rich, P. R. , Segal, A. W. (2015) Alkalinity of neutrophil phagocytic vacuoles is modulated by HVCN1 and has consequences for myeloperoxidase activity. PLoS One 10, e0125906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Nathan, C. (2006) Neutrophils and immunity: challenges and opportunities. Nat. Rev. Immunol. 6, 173–182. [DOI] [PubMed] [Google Scholar]
- 14. Klebanoff, S. J. (2005) Myeloperoxidase: friend and foe. J. Leukoc. Biol. 77, 598–625. [DOI] [PubMed] [Google Scholar]
- 15. Churg, A. , Marshall, C. V. , Sin, D. D. , Bolton, S. , Zhou, S. , Thain, K. , Cadogan, E. B. , Maltby, J. , Soars, M. G. , Mallinder, P. R. , Wright, J. L. (2012) Late intervention with a myeloperoxidase inhibitor stops progression of experimental chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 185, 34–43. [DOI] [PubMed] [Google Scholar]
- 16. Stamp, L. K. , Khalilova, I. , Tarr, J. M. , Senthilmohan, R. , Turner, R. , Haigh, R. C. , Winyard, P. G. , Kettle, A. J. (2012) Myeloperoxidase and oxidative stress in rheumatoid arthritis. Rheumatology (Oxford) 51, 1796–1803. [DOI] [PubMed] [Google Scholar]
- 17. Heinecke, J. W. (2003) Oxidative stress: new approaches to diagnosis and prognosis in atherosclerosis. Am. J. Cardiol. 91 (3A), 12A–16A. [DOI] [PubMed] [Google Scholar]
- 18. Nicholls, S. J. , Hazen, S. L. (2005) Myeloperoxidase and cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 25, 1102–1111. [DOI] [PubMed] [Google Scholar]
- 19. Nicholls, S. J. , Hazen, S. L. (2009) Myeloperoxidase, modified lipoproteins, and atherogenesis. J. Lipid Res. 50 (Suppl), S346–S351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Panizzi, P. , Swirski, F. K. , Figueiredo, J. L. , Waterman, P. , Sosnovik, D. E. , Aikawa, E. , Libby, P. , Pittet, M. , Weissleder, R. , Nahrendorf, M. (2010) Impaired infarct healing in atherosclerotic mice with Ly‐6Chi monocytosis. J. Am. Coll. Cardiol. 55, 1629–1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Warnatsch, A. , Ioannou, M. , Wang, Q. , Papayannopoulos, V. (2015) Inflammation: neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 349, 316–320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ambrosone, C. B. , Barlow, W. E. , Reynolds, W. , Livingston, R. B. , Yeh, I. T. , Choi, J. Y. , Davis, W. , Rae, J. M. , Tang, L. , Hutchins, L. R. , Ravdin, P. M. , Martino, S. , Osborne, C. K. , Lyss, A. P. , Hayes, D. F. , Albain, K. S. (2009) Myeloperoxidase genotypes and enhanced efficacy of chemotherapy for early‐stage breast cancer in SWOG‐8897. J. Clin. Oncol. 27, 4973–4979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Ramsey, M. R. , Sharpless, N. E. (2006) ROS as a tumour suppressor? Nat. Cell Biol. 8, 1213–1215. [DOI] [PubMed] [Google Scholar]
- 24. Zheng, W. , Warner, R. , Ruggeri, R. , Su, C. , Cortes, C. , Skoura, A. , Ward, J. , Ahn, K. , Kalgutkar, A. , Sun, D. , Maurer, T. S. , Bonin, P. D. , Okerberg, C. , Bobrowski, W. , Kawabe, T. , Zhang, Y. , Coskran, T. , Bell, S. , Kapoor, B. , Johnson, K. , Buckbinder, L. (2015) PF‐1355, a mechanism‐based myeloperoxidase inhibitor, prevents immune complex vasculitis and anti‐glomerular basement membrane glomerulonephritis. J. Pharmacol. Exp. Ther. 353, 288–298. [DOI] [PubMed] [Google Scholar]
- 25. Liu, C. , Desikan, R. , Ying, Z. , Gushchina, L. , Kampfrath, T. , Deiuliis, J. , Wang, A. , Xu, X. , Zhong, J. , Rao, X. , Sun, Q. , Maiseyeu, A. , Parthasarathy, S. , Rajagopalan, S. (2012) Effects of a novel pharmacologic inhibitor of myeloperoxidase in a mouse atherosclerosis model. PLoS One 7, e50767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Furtmüller, P. G. , Zederbauer, M. , Jantschko, W. , Helm, J. , Bogner, M. , Jakopitsch, C. , Obinger, C. (2006) Active site structure and catalytic mechanisms of human peroxidases. Arch. Biochem. Biophys. 445, 199–213. [DOI] [PubMed] [Google Scholar]
- 27. Harrison, J. E. , Araiso, T. , Palcic, M. M. , Dunford, H. B. (1980) Compound I of myeloperoxidase. Biochem. Biophys. Res. Commun. 94, 34–40. [DOI] [PubMed] [Google Scholar]
- 28. Kettle, A. J. , van Dalen, C. J. , Winterbourn, C. C. (1997) Peroxynitrite and myeloperoxidase leave the same footprint in protein nitration. Redox Rep. 3, 257–258. [DOI] [PubMed] [Google Scholar]
- 29. Everse, J. (1998) The structure of heme proteins Compounds I and II: some misconceptions. Free Radic. Biol. Med. 24, 1338–1346. [DOI] [PubMed] [Google Scholar]
- 30. Andrews, P. C. , Krinsky, N. I. (1982) A kinetic analysis of the interaction of human myeloperoxidase with hydrogen peroxide, chloride ions, and protons. J. Biol. Chem. 257, 13240–13245. [PubMed] [Google Scholar]
- 31. Andrews, P. C. , Krinsky, N. I. (1982) Quantitative determination of myeloperoxidase using tetramethylbenzidine as substrate. Anal. Biochem. 127, 346–350. [DOI] [PubMed] [Google Scholar]
- 32. Fiedler, T. J. , Davey, C. A. , Fenna, R. E. (2000) X‐ray crystal structure and characterization of halide‐binding sites of human myeloperoxidase at 1.8 A resolution. J. Biol. Chem. 275, 11964–11971. [DOI] [PubMed] [Google Scholar]
- 33. Tidén, A. K. , Sjögren, T. , Svensson, M. , Bernlind, A. , Senthilmohan, R. , Auchère, F. , Norman, H. , Markgren, P. O. , Gustavsson, S. , Schmidt, S. , Lundquist, S. , Forbes, L. V. , Magon, N. J. , Paton, L. N. , Jameson, G. N. , Eriksson, H. , Kettle, A. J. (2011) 2‐Thioxanthines are mechanism‐based inactivators of myeloperoxidase that block oxidative stress during inflammation. J. Biol. Chem. 286, 37578–37589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Samygina, V. R. , Sokolov, A. V. , Bourenkov, G. , Petoukhov, M. V. , Pulina, M. O. , Zakharova, E. T. , Vasilyev, V. B. , Bartunik, H. , Svergun, D. I. (2013) Ceruloplasmin: macromolecular assemblies with iron‐containing acute phase proteins. PLoS ONE 8, e67145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Carpena, X. , Vidossich, P. , Schroettner, K. , Calisto, B. M. , Banerjee, S. , Stampler, J. , Soudi, M. , Furtmüller, P. G. , Rovira, C. , Fita, I. , Obinger, C. (2009) Essential role of proximal histidine‐asparagine interaction in mammalian peroxidases. J. Biol. Chem. 284, 25929–25937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Blair‐Johnson, M. , Fiedler, T. , Fenna, R. (2001) Human myeloperoxidase: structure of a cyanide complex and its interaction with bromide and thiocyanate substrates at 1.9 A resolution. Biochemistry 40, 13990–13997. [DOI] [PubMed] [Google Scholar]
- 37. Fenna, R. , Zeng, J. , Davey, C. (1995) Structure of the green heme in myeloperoxidase. Arch. Biochem. Biophys. 316, 653–656. [DOI] [PubMed] [Google Scholar]
- 38. Forbes, L. V. , Sjögren, T. , Auchère, F. , Jenkins, D. W. , Thong, B. , Laughton, D. , Hemsley, P. , Pairaudeau, G. , Turner, R. , Eriksson, H. , Unitt, J. F. , Kettle, A. J. (2013) Potent reversible inhibition of myeloperoxidase by aromatic hydroxamates. J. Biol. Chem. 288, 36636–36647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Kooter, I. M. , Moguilevsky, N. , Bollen, A. , Sijtsema, N. M. , Otto, C. , Wever, R. (1997) Site‐directed mutagenesis of Met243, a residue of myeloperoxidase involved in binding of the prosthetic group. J. Biol. Inorg. Chem. 2, 191–197. [Google Scholar]
- 40. Wever, R. , Plat, H. (1981) Spectral properties of myeloperoxidase and its ligand complexes. Biochim. Biophys. Acta 661, 235–239. [DOI] [PubMed] [Google Scholar]
- 41. Huang, J. , Smith, F. , Panizzi, P. (2014) Ordered cleavage of myeloperoxidase ester bonds releases active site heme leading to inactivation of myeloperoxidase by benzoic acid hydrazide analogs. Arch. Biochem. Biophys. 548, 74–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Huang, J. , Smith, F. , Panizzi, J. R. , Goodwin, D. C. , Panizzi, P. (2015) Inactivation of myeloperoxidase by benzoic acid hydrazide. Arch. Biochem. Biophys. 570, 14–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Huang, L. , Wojciechowski, G. , Ortiz de Montellano, P. R. (2006) Role of heme‐protein covalent bonds in mammalian peroxidases: protection of the heme by a single engineered heme‐protein link in horseradish peroxidase. J. Biol. Chem. 281, 18983–18988. [DOI] [PubMed] [Google Scholar]
- 44. Nordberg, J. , Arnér, E. S. (2001) Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 31, 1287–1312. [DOI] [PubMed] [Google Scholar]
- 45. Bedard, K. , Krause, K. H. (2007) The NOX family of ROS‐generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87, 245–313. [DOI] [PubMed] [Google Scholar]
- 46. Hampton, M. B. , Kettle, A. J. , Winterbourn, C. C. (1998) Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood 92, 3007–3017. [PubMed] [Google Scholar]
- 47. Cai, H. , Dikalov, S. , Griendling, K. K. , Harrison, D. G. (2007) Detection of reactive oxygen species and nitric oxide in vascular cells and tissues: comparison of sensitivity and specificity. Methods Mol. Med. 139, 293–311. [DOI] [PubMed] [Google Scholar]
- 48. Dikalov, S. , Skatchkov, M. , Bassenge, E. (1997) Spin trapping of superoxide radicals and peroxynitrite by 1‐hydroxy‐3‐carboxy‐pyrrolidine and 1‐hydroxy‐2,2,6,6‐tetramethyl‐4‐oxo‐piperidine and the stability of corresponding nitroxyl radicals towards biological reductants. Biochem. Biophys. Res. Commun. 231, 701–704. [DOI] [PubMed] [Google Scholar]
- 49. Britigan, B. E. , Hamill, D. R. (1989) The interaction of 5,5‐dimethyl‐1‐pyrroline‐N‐oxide with human myeloperoxidase and its potential impact on spin trapping of neutrophil‐derived free radicals. Arch. Biochem. Biophys. 275, 72–81. [DOI] [PubMed] [Google Scholar]
- 50. Dikalov, S. I. , Kirilyuk, I. A. , Voinov, M. , Grigor'ev, I. A. (2011) EPR detection of cellular and mitochondrial superoxide using cyclic hydroxylamines. Free Radic. Res. 45, 417–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ichimori, K. , Arroyo, C. M. , Nakazawa, H. (1996) Electron spin resonance for spin trapping of 3,5‐dibromo‐4‐nitrosobenzene sulfonate. Methods Enzymol. 268, 203–211. [DOI] [PubMed] [Google Scholar]
- 52. Dikalov, S. I. , Harrison, D. G. (2014) Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal 20, 372–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Hempel, S. L. , Buettner, G. R. , O'Malley, Y. Q. , Wessels, D. A. , Flaherty, D. M. (1999) Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: comparison with 2',7'‐dichlorodihydrofluorescein diacetate, 5(and 6)‐carboxy‐2',7'‐dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 27, 146–159. [DOI] [PubMed] [Google Scholar]
- 54. Bass, D. A. , Parce, J. W. , Dechatelet, L. R. , Szejda, P. , Seeds, M. C. , Thomas, M. (1983) Flow cytometric studies of oxidative product formation by neutrophils: a graded response to membrane stimulation. J. Immunol. 130, 1910–1917. [PubMed] [Google Scholar]
- 55. Wang, X. , Fang, H. , Huang, Z. , Shang, W. , Hou, T. , Cheng, A. , Cheng, H. (2013) Imaging ROS signaling in cells and animals. J. Mol. Med. (Berl.) 91, 917–927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Kojima, H. , Nakatsubo, N. , Kikuchi, K. , Kawahara, S. , Kirino, Y. , Nagoshi, H. , Hirata, Y. , Nagano, T. (1998) Detection and imaging of nitric oxide with novel fluorescent indicators: diaminofluoresceins. Anal. Chem. 70, 2446–2453. [DOI] [PubMed] [Google Scholar]
- 57. Chan, J. , Dodani, S. C. , Chang, C. J. (2012) Reaction‐based small‐molecule fluorescent probes for chemoselective bioimaging. Nat. Chem. 4, 973–984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Robinson, K. M. , Janes, M. S. , Pehar, M. , Monette, J. S. , Ross, M. F. , Hagen, T. M. , Murphy, M. P. , Beckman, J. S. (2006) Selective fluorescent imaging of superoxide in vivo using ethidium‐based probes. Proc. Natl. Acad. Sci. U. S. A. 103, 15038–15043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Setsukinai, K. , Urano, Y. , Kakinuma, K. , Majima, H. J. , Nagano, T. (2003) Development of novel fluorescence probes that can reliably detect reactive oxygen species and distinguish specific species. J. Biol. Chem. 278, 3170–3175. [DOI] [PubMed] [Google Scholar]
- 60. Flemmig, J. , Zschaler, J. , Remmler, J. , Arnhold, J. (2012) The fluorescein‐derived dye aminophenyl fluorescein is a suitable tool to detect hypobromous acid (HOBr)‐producing activity in eosinophils. J. Biol. Chem. 287, 27913–27923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Peng, T. , Yang, D. (2010) HKGreen‐3: a rhodol‐based fluorescent probe for peroxynitrite. Org. Lett. 12, 4932–4935. [DOI] [PubMed] [Google Scholar]
- 62. Sun, Z. N. , Liu, F. Q. , Chen, Y. , Tam, P. K. , Yang, D. (2008) A highly specific BODIPY‐based fluorescent probe for the detection of hypochlorous acid. Org. Lett. 10, 2171–2174. [DOI] [PubMed] [Google Scholar]
- 63. Pulli, B. , Ali, M. , Forghani, R. , Schob, S. , Hsieh, K. L. , Wojtkiewicz, G. , Linnoila, J. J. , Chen, J. W. (2013) Measuring myeloperoxidase activity in biological samples. PLoS One 8, e67976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Mempel, T. R. , Scimone, M. L. , Mora, J. R. , von Andrian, U. H. (2004) In vivo imaging of leukocyte trafficking in blood vessels and tissues. Curr. Opin. Immunol. 16, 406–417. [DOI] [PubMed] [Google Scholar]
- 65. Sumen, C. , Mempel, T. R. , Mazo, I. B. , von Andrian, U. H. (2004) Intravital microscopy: visualizing immunity in context. Immunity 21, 315–329. [DOI] [PubMed] [Google Scholar]
- 66. Megens, R. T. , Kemmerich, K. , Pyta, J. , Weber, C. , Soehnlein, O. (2011) Intravital imaging of phagocyte recruitment. Thromb. Haemost. 105, 802–810. [DOI] [PubMed] [Google Scholar]
- 67. Doyle, A. D. , Jacobsen, E. A. , Ochkur, S. I. , McGarry, M. P. , Shim, K. G. , Nguyen, D. T. , Protheroe, C. , Colbert, D. , Kloeber, J. , Neely, J. , Shim, K. P. , Dyer, K. D. , Rosenberg, H. F. , Lee, J. J. , Lee, N. A. (2013) Expression of the secondary granule proteins major basic protein 1 (MBP‐1) and eosinophil peroxidase (EPX) is required for eosinophilopoiesis in mice. Blood 122, 781–790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Odobasic, D. , Yang, Y. , Muljadi, R. C. , O'Sullivan, K. M. , Kao, W. , Smith, M. , Morand, E. F. , Holdsworth, S. R. (2014) Endogenous myeloperoxidase is a mediator of joint inflammation and damage in experimental arthritis. Arthritis Rheumatol. 66, 907–917. [DOI] [PubMed] [Google Scholar]
- 69. Maeda, H. (2001) The enhanced permeability and retention (EPR) effect in tumor vasculature: the key role of tumor‐selective macromolecular drug targeting. Adv. Enzyme Regul. 41, 189–207. [DOI] [PubMed] [Google Scholar]
- 70. Chen, J. W. , Querol Sans, M. , Bogdanov, Jr., A. , Weissleder, R. (2006) Imaging of myeloperoxidase in mice by using novel amplifiable paramagnetic substrates. Radiology 240, 473–481. [DOI] [PubMed] [Google Scholar]
- 71. Panizzi, P. , Nahrendorf, M. , Wildgruber, M. , Waterman, P. , Figueiredo, J. L. , Aikawa, E. , McCarthy, J. , Weissleder, R. , Hilderbrand, S. A. (2009) Oxazine conjugated nanoparticle detects in vivo hypochlorous acid and peroxynitrite generation. J. Am. Chem. Soc. 131, 15739–15744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Gross, S. , Gammon, S. T. , Moss, B. L. , Rauch, D. , Harding, J. , Heinecke, J. W. , Ratner, L. , Piwnica‐Worms, D. (2009) Bioluminescence imaging of myeloperoxidase activity in vivo. Nat. Med. 15, 455–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Zhang, N. , Francis, K. P. , Prakash, A. , Ansaldi, D. (2013) Enhanced detection of myeloperoxidase activity in deep tissues through luminescent excitation of near‐infrared nanoparticles. Nat. Med. 19, 500–505. [DOI] [PubMed] [Google Scholar]
- 74. Stamatas, G. N. , Southall, M. , Kollias, N. (2006) In vivo monitoring of cutaneous edema using spectral imaging in the visible and near infrared. J. Invest. Dermatol. 126, 1753–1760. [DOI] [PubMed] [Google Scholar]
- 75. Sanders, J. M. , Chen, L. J. , Burka, L. T. , Matthews, H. B. (2000) Metabolism and disposition of luminol in the rat. Xenobiotica 30, 263–272. [DOI] [PubMed] [Google Scholar]
- 76. Eggleston, H. , Panizzi, P. (2014) Molecular imaging of bacterial infections in vivo: the discrimination between infection and inflammation. Informatics 1, 72–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Shepherd, J. , Hilderbrand, S. A. , Waterman, P. , Heinecke, J. W. , Weissleder, R. , Libby, P. (2007) A fluorescent probe for the detection of myeloperoxidase activity in atherosclerosis‐associated macrophages. Chem. Biol. 14, 1221–1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Magalotti, S. , Gustafson, T. P. , Cao, Q. , Abendschein, D. R. , Pierce, R. A. , Berezin, M. Y. , Akers, W. J. (2013) Evaluation of inflammatory response to acute ischemia using near‐infrared fluorescent reactive oxygen sensors. Mol. Imaging Biol. 15, 423–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Pu, K. , Shuhendler, A. J. , Jokerst, J. V. , Mei, J. , Gambhir, S. S. , Bao, Z. , Rao, J. (2014) Semiconducting polymer nanoparticles as photoacoustic molecular imaging probes in living mice. Nat. Nanotechnol. 9, 233–239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Chen, J. W. , Pham, W. , Weissleder, R. , Bogdanov, A., Jr. (2004) Human myeloperoxidase: a potential target for molecular MR imaging in atherosclerosis. Magn. Reson. Med. 52, 1021–1028. [DOI] [PubMed] [Google Scholar]
- 81. Rodríguez, E. , Nilges, M. , Weissleder, R. , Chen, J. W. (2010) Activatable magnetic resonance imaging agents for myeloperoxidase sensing: mechanism of activation, stability, and toxicity. J. Am. Chem. Soc. 132, 168–177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Pulli, B. , Bure, L. , Wojtkiewicz, G. R. , Iwamoto, Y. , Ali, M. , Li, D. , Schob, S. , Hsieh, K. L. , Jacobs, A. H. , Chen, J. W. (2015) Multiple sclerosis: myeloperoxidase immunoradiology improves detection of acute and chronic disease in experimental model. Radiology 275, 480–489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Nahrendorf, M. , Sosnovik, D. , Chen, J. W. , Panizzi, P. , Figueiredo, J. L. , Aikawa, E. , Libby, P. , Swirski, F. K. , Weissleder, R. (2008) Activatable magnetic resonance imaging agent reports myeloperoxidase activity in healing infarcts and noninvasively detects the antiinflammatory effects of atorvastatin on ischemia‐reperfusion injury. Circulation 117, 1153–1160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Breckwoldt, M. O. , Chen, J. W. , Stangenberg, L. , Aikawa, E. , Rodriguez, E. , Qiu, S. , Moskowitz, M. A. , Weissleder, R. (2008) Tracking the inflammatory response in stroke in vivo by sensing the enzyme myeloperoxidase. Proc. Natl. Acad. Sci. U. S. A. 105, 18584–18589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Chen, J. W. , Breckwoldt, M. O. , Aikawa, E. , Chiang, G. , Weissleder, R. (2008) Myeloperoxidase‐targeted imaging of active inflammatory lesions in murine experimental autoimmune encephalomyelitis. Brain 131, 1123–1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Yao, J. , Kaberniuk, A. A. , Li, L. , Shcherbakova, D. M. , Zhang, R. , Wang, L. , Li, G. , Verkhusha, V. V. , Wang, L. V. (2016) Multiscale photoacoustic tomography using reversibly switchable bacterial phytochrome as a near‐infrared photochromic probe. (Published online ahead of print November 9, 2015) Nat. Methods 13, 67–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Jathoul, A. P. , Laufer, J. , Ogunlade, O. , Treeby, B. , Cox, B. , Zhang, E. , Johnson, P. , Pizzey, A. R. , Philip, B. , Marafioti, T. , Lythgoe, M. F. , Pedley, R. B. , Pule, M. A. , Beard, P. (2015) Deep in vivo photoacoustic imaging of mammalian tissues using a tyrosinase‐based genetic reporter. Nat. Photonics 9, 239–246. [Google Scholar]
- 88. Manohar, S. , van Apeldoorn, A. , Steenbergen, W. (2015) Photoacoustic imaging: cells make themselves heard. Nat. Photonics 9, 216–218. [Google Scholar]
- 89. Razansky, D. , Distel, M. , Vinegoni, C. , Ma, R. , Perrimon, N. , Köster, R. W. , Ntziachristos, V. (2009) Multispectral opto‐acoustic tomography of deep‐seated fluorescent proteins in vivo. Nat. Photonics 3, 412–417. [Google Scholar]
- 90. Ntziachristos, V. (2010) Going deeper than microscopy: the optical imaging frontier in biology. Nat. Methods 7, 603–614. [DOI] [PubMed] [Google Scholar]
- 91. Ntziachristos, V. , Razansky, D. (2010) Molecular imaging by means of multispectral optoacoustic tomography (MSOT). Chem. Rev. 110, 2783–2794. [DOI] [PubMed] [Google Scholar]