

Abstract
Angelman syndrome (AS) is caused by the loss of Ube3A, an ubiquitin ligase that commits specific proteins to proteasomal degradation. How this defect causes autism and other pathological phenotypes associated with AS is unknown. Long-term depression (LTD) of excitatory synaptic transmission mediated by type 5 metabotropic glutamate (mGlu5) receptors was enhanced in hippocampal slices of Ube3Am−/p+ mice, which model AS. No changes were found in NMDA-dependent LTD induced by low-frequency stimulation. mGlu5 receptor-dependent LTD in AS mice was sensitive to the protein synthesis inhibitor anisomycin, and relied on the same signaling pathways as in wild-type mice, e.g., the mitogen-activated protein kinase (MAPK) pathway, the phosphatidylinositol-3-kinase (PI3K)/mammalian target of rapamycine pathway, and protein tyrosine phosphatase. Neither the stimulation of MAPK and PI3K nor the increase in Arc (activity-regulated cytoskeleton-associated protein) levels in response to mGlu5 receptor activation were abnormal in hippocampal slices from AS mice compared with wild-type mice. mGlu5 receptor expression and mGlu1/5 receptor-mediated polyphosphoinositide hydrolysis were also unchanged in the hippocampus of AS mice. In contrast, AS mice showed a reduced expression of the short Homer protein isoform Homer 1a, and an increased coupling of mGlu5 receptors to Homer 1b/c proteins in the hippocampus. These findings support the link between Homer proteins and monogenic autism, and lay the groundwork for the use of mGlu5 receptor antagonists in AS.
Keywords: Angelman syndrome, hippocampus, Homer proteins, LTD, metabotropic glutamate receptors
Introduction
Long-term depression (LTD) of excitatory synaptic transmission mediated by type 5 metabotropic glutamate (mGlu5) receptors is amplified in the hippocampus of Fmr1 knock-out mice modeling fragile X syndrome (FXS; Huber et al., 2002; Bear et al., 2004), a genetic disorder associated with autism in ∼30–35% of affected children (Kelleher and Bear, 2008). Pathological behavioral phenotypes of Fmr1 knock-out mice are corrected by germline manipulations that reduce the expression of mGlu5 receptors (Dölen et al., 2007) or by treatments with negative allosteric modulators (NAMs) of mGlu5 receptors (Bhakar et al., 2012; Michalon et al., 2012), suggesting that exaggerated mGlu5 receptor activity contributes to the pathophysiology of FXS. Moving from these findings, clinical studies are underway to test the effectiveness of mGlu5 receptor NAMs in the treatment of FXS (for review, see Krueger and Bear, 2011; Hagerman et al., 2012).
Aberrant protein synthesis lies at the core of synaptic modifications associated with FXS (Feng et al., 1995), and mGlu5 receptor-dependent LTD in the hippocampus relies on dendritic protein synthesis (Huber et al., 2000; but see also Moult et al., 2008; Waung and Huber, 2009). Because an aberrant protein synthesis is a common motif of autism spectrum disorders (Kelleher and Bear, 2008), there is increasing interest in examining mGlu5 receptor activity in other models of monogenic autism. Auerbach et al. (2011) found that mice carrying heterozygous loss-of-function mutations of the tuberous sclerosis complex-2 (Tsc2) showed a reduced mGlu5 receptor-dependent LTD in the hippocampus. Some of the phenotypes of Tsc2+/− mice were corrected by cross-breeding with Fmr1 knock-out mice or by treatment with a positive allosteric modulator of mGlu5 receptors. Thus, deviations in either direction in mGlu5 receptor-mediated protein synthesis and synaptic plasticity can lead to shared pathological phenotypes (Auerbach et al., 2011).
Here, we examined mGlu5 receptor-dependent synaptic plasticity in a mouse model of Angelman syndrome (AS), a disorder characterized by developmental delay, epilepsy, hyperactivity, and autistic features (Steffenburg et al., 1996; Williams, 2005). AS is caused by mutations or deletions of the maternally inherited Ube3A gene, because the paternal allele of Ube3A is epigenetically silenced in neurons (Kishino et al., 1997). A mouse model of AS has been generated by knocking out 3 kb of the sequence orthologous to exon 2 of the human Ube3A gene (Jiang et al., 1998). Ube3A is an E3 ubiquitin ligase, which provides substrate specificity to the ubiquitin proteasome system (UPS). The UPS plays a critical role in the regulation of synaptic plasticity (Ehlers, 2003; Dong et al., 2008), and Ube3A knock-out mice display impaired hippocampal long-term potentiation (Jiang et al., 1998, Weeber et al., 2003) and visual cortex plasticity (Yashiro et al., 2009; Sato and Stryker, 2010). Here, we report that AS mice show a selective amplification of mGlu5 receptor-mediated LTD in the hippocampus and alterations in mGlu5 receptor coupling to Homer proteins.
Materials and Methods
Drugs.
(RS)-3,5-dihydroxyphenylglycine (DHPG), 2-methyl-6-(phenylethynyl)-pyridine (MPEP), (E)-2-methyl-6-stryrylpyridine(-)-2-oxa-4-aminocyclo[3.1.0]hexane-4,6-dicarboxylic acid (LY367385), U0126, d-2-amino-5-phosphonopentanoic acid (d-AP5), UBE1-41, and anysomicin were obtained from Tocris Cookson. Phenylarsine oxide (PAO) and rapamycin were obtained from Sigma-Aldrich.
Animals.
Heterozygous Ube3A mice were purchased from The Jackson Laboratory (Jackson code: 129-Ube3atm1Alb/J) and maintained in a C57BL/6 background. The genotyping was carried by PCR analysis using the following primers: 5′-GCTCAAGGTTGTATGCCTTGGTGCT-3′ (oIMR1965); 5′-AGTTCTCAAGGTAAGCTGAGCTTGC-3′ (oIMR1966); and 5′-TGCATCGCATTGTCTGAGTAGGTGTC-3′ (oIMR1967; The Jackson Laboratory).
Mice were kept under environmentally controlled conditions (ambient temperature, 22°C; humidity, 40%) on a 12 h light/dark cycle with food and water ad libitum. All experiments were performed on mice of either sex. Experiments were performed following the Guidelines for Animal Care and Use of the National Institutes of Health. All efforts were made to minimize animal suffering and to reduce the number of animals used.
Electrophysiology.
Hippocampal slices were prepared from 4-week-old to 5-week-old Ube3A maternal deficient mice (Ube3Am−/p+ “AS” mice) and their wild-type (Ube3Am+/p+) littermates, as previously described (Nisticò et al., 2013). Brains were rapidly dissected out and parasagittal slices (400 μm) were prepared and incubated in artificial CSF (ACSF) containing the following (in mm): 124 NaCl, 3.0 KCl, 1.0 MgCl2, 2.0 CaCl2, 1.25 NaH2PO4, 26 NaHCO3, 10 glucose, saturated with 95% O2, 5% CO2, pH 7.4. The CA3 region was not removed from the slices. Slices were allowed to recover for 2–4 h and then placed on a nylon mesh, completely submerged in a small chamber (0.8 ml), and superfused with oxygenated ACSF (30–31°C) at a constant flow rate of 2.5–3.0 ml/min. The slope of the field EPSPs (fEPSPs) was recorded from the apical dendrite layer of the CA1 pyramidal cells by means of saline-filled glass electrodes of ∼2–4 MΩ resistance. Stimulating monopolar electrodes were placed in Schaffer collateral/commissural afferents, and stimulation amplitude was adjusted so as to produce one-half of the maximal response. Signals were filtered at 3 kHz and digitized at 10 kHz. After the stabilization of the fEPSP, LTD was induced by low-frequency stimulation (1 Hz for 15 min) or following DHPG application (100 μm, 5 min). In some experiments DHPG was applied in the presence of the NMDA receptor antagonist d-AP5 (50 μm), the proteasome inhibitor UBE1-41 (50 μm), the protein tyrosine phosphatase inhibitor PAO (15 μm), the mammalian target of rapamycine (mTOR) inhibitor rapamycin (20 μm), the extracellular regulated kinase1/2 (ERK1/2) kinase inhibitor UO126 (20 μm), or the protein synthesis inhibitor anisomycin (20 μm). d-AP5 was applied 20 min before DHPG and maintained during the recording session; UBE1-41 (50 μm, from a mother solution of 50 mm in dimethyl sulfoxide) was applied to the slices during the recovery time for 60 min before placement in the recording chamber (Citri et al., 2009). All the other drugs were applied as indicated (see figures).
Immunoblotting.
Slices prepared as described for electrophysiological studies were allowed to recover for ≥3 h. Slices were then incubated with DHPG (100 μm) for 5 min and then snap frozen in liquid nitrogen. Samples were homogenized at 4°C in a lysis buffer composed of Tris-HCl 10 mm, pH 7.4; NaCl, 150 mm; EDTA, 5 mm; Igepal 1%; protease (Santa Cruz Biotechnology) and phosphatase (Sigma-Aldrich) inhibitor mixture. Five microliters of tissue extracts were used for protein determination. Proteins (30 μg) were resuspended in SDS-bromophenol blue reducing buffer with 40 mm DTT and used for protein analysis. Immunoblotting was performed with the following primary antibodies: Ube3A (Bethyl Laboratories), mGlu5 receptor (Millipore Biotechnology), p-ERK1/2 (Thr202/Tyr204; Santa Cruz Biotechnology), ERK (Cell Signaling Technology), p-Akt (Ser473; Cell Signaling Technology), Akt (Cell Signaling Technology), and Arc (activity-regulated cytoskeleton-associated protein; kindly provided by Prof. P. Worley, Department of Neuroscience, Johns Hopkins University School of Medicine, Baltimore, MD). After incubation in primary antibody overnight at 4°C, immunoblots were incubated with HRP-conjugated secondary antibodies (Calbiochem) and developed by ECL (Hybond ECL, GE Healthcare Europe).
Measurement of polyphosphoinositide hydrolysis in hippocampal slices.
Group I mGlu receptor-stimulated polyphosphoinositide (PI) hydrolysis was also measured in hippocampal slices obtained from postnatal day (P) 21–P30 Ube3Am−/p+ mice and their wild-type littermates as described previously (Nicoletti et al., 1986). Briefly, hippocampi were sliced (350 × 350 μm) using a McIlwain tissue chopper. Forty microliters of gravity-packed slices were then incubated for 60 min in 250 μl of buffer containing 1 μCi of myo-[3H]inositol. Slices were incubated with LiCl (10 mm for 10 min) followed by DHPG (100 μm). One hour later, the incubation was stopped by the addition of 900 μl of methanol/chloroform (2:1). After further addition of 300 μl of chloroform and 600 μl of water, samples were centrifuged at low speed to facilitate phase separation, and the upper aqueous phase was loaded into Dowex 1-X-8 columns for the separation and quantification of [3H]Inositolmonophosphate (InsP).
Gene expression analysis by real-time PCR.
Total RNA was isolated from hippocampi using TRIzol reagent (Invitrogen) according to the manufacturer's protocol and retrotranscribed into cDNA by using SuperScript III Reverse Transcriptase (Invitrogen). Real-time PCR was performed on the StepOnePlus (Applied Biosystems). PCR was performed by using Power SYBR Green PCR Master Mix Kit (Applied Biosystems) according to the manufacturer's instructions. Thermal cycler conditions were as follows: 10 min at 95°C, 40 cycles of denaturation (45 s at 95°C), and combined annealing/extension (1 min at 60°C). Sequences of primers used were as follows: Homer 1a: forward 5′-TCTTCAGTCTCCTTTGACACCA-3′ and reverse 5′-CATGATTGCTGAATTGAATGTG-3′; pan-Homer 1: forward 5′-TGGACTGGGATTCTCCTCTG-3′ and reverse 5′-TGTGTCACATCGGGTGTTCT-3′; mGlu5 receptor: forward 5′-ACGAAGACCAACCGTATTGC-3′ and reverse 5′-AGACTTCTCGGATGCTTGGA-3′; cyclophilin A: forward 5′-TCCAAAGACAGCAGAAAACTTTCG-3′ and reverse 5′-TCTTCTTGCTGGTCTTGCCATTCC-3′.
Concentrations of mRNA were calculated from serially diluted standard curves simultaneously amplified with the samples and normalized versus cyclophilin A mRNA levels.
Coimmunoprecipitation.
Hippocampi were homogenized at 4°C in a lysis buffer (as above) and 1 mg of total proteins were resuspended in a coimmunoprecipitation buffer (50 mm Tris, pH 7.4, 120 mm NaCl, 0.5% Nonidet P-40, 1 mm EDTA, 1 mm EGTA). Proteins were tumbled overnight at 4°C with 5 μg of antibody anti-Homer 1b/c or anti-pan-Homer (Santa Cruz Biotechnology). Protein A agarose bead slurry (GE Healthcare) was added for 2 h, and the beads were then washed with coimmunoprecipitation buffer. Western blotting was performed with antibodies against Homer 1b/c, pan-Homer, and mGlu5 receptor (Millipore Biotechnology). Protein extracts from the cerebral cortex of normal and mGlu5 receptor knock-out mice (stored in our laboratory) were used as positive and negative controls, respectively.
Statistical analysis.
Electrophysiological data were normalized to the averaged value of the initial slope of the fEPSP obtained during the 20 min period before the application of the conditioning stimulus or DHPG. Data are expressed as the means ± SEM. Significant differences between groups were determined using two-tailed unpaired Student's t test performed on a 10 min average taken 50 min after DHPG application. Statistical significance was set at p < 0.05. All experiments and the analysis of data were performed in a blind manner. For all statistical comparisons, the n used was the number of animals rather than number of slices. For biochemical experiments, statistical analysis was performed using two-way ANOVA plus Fisher's PLSD test or the Student's t test.
Results
Enhancement of mGlu5 receptor-dependent LTD in the hippocampus of AS mice
We measured basal synaptic transmission and activity-dependent synaptic plasticity at the Schaffer collateral–CA1 synapses in hippocampal slices prepared from Ube3A maternal deficient mice (Ube3Am−/p+ AS mice) and wild-type (Ube3Am+/p+) littermates. No Ube3A was detected in hippocampal slices of AS mice, as expected (Fig. 1A). AS mice did not show alterations in basal synaptic transmission (Fig. 1B; p > 0.05; Jiang et al., 1998), and in LTD induced by low-frequency stimulation, which is known to be dependent on NMDA receptor activation (for review, see Manabe, 1997). Stimulation at 1 Hz for 15 min induced a similar depression of synaptic transmission in slices from wild-type and AS mice (wild-type: 85 ± 9%, n = 7; AS: 82 ± 11%, n = 8, p > 0.05; Fig. 1C). In contrast, LTD induced by bath application of DHPG (100 μm, 5 min) was amplified in AS mice (wild-type: 81 ± 8%, n = 9; AS: 61 ± 6%, n = 12, p < 0.05; Fig. 1D). The amplification was unaltered in the presence of the NMDA receptor antagonist, d-AP5 (50 μm; 60 ± 7%, n = 4), excluding any role for endogenous NMDA receptor activation in the DHPG/LTD phenotype of AS mice. As expected (Huber et al., 2001), two consecutive applications of DHPG produced maximal depression of fEPSPs in slices from wild-type mice. In contrast, only one application of DHPG was sufficient to achieve saturated levels of LTD in slices from AS mice, such that maximal depression did not differ between the two genotypes (wild-type: 67 ± 6%, n = 8; AS: 62 ± 5%, n = 8, p > 0.05; Fig. 1E). Paired-pulse facilitation (PPF), a presynaptic form of short-term synaptic plasticity (Zucker, 1989), did not differ between wild-type and AS mice at multiple interpulse intervals (Fig. 1F; p > 0.05). The increase in PPF induced by DHPG was also similar between the two genotypes, indicating no changes in the presynaptic component of group I mGlu-receptor-dependent LTD in AS mice (Fig. 1F; p > 0.05).
Figure 1.
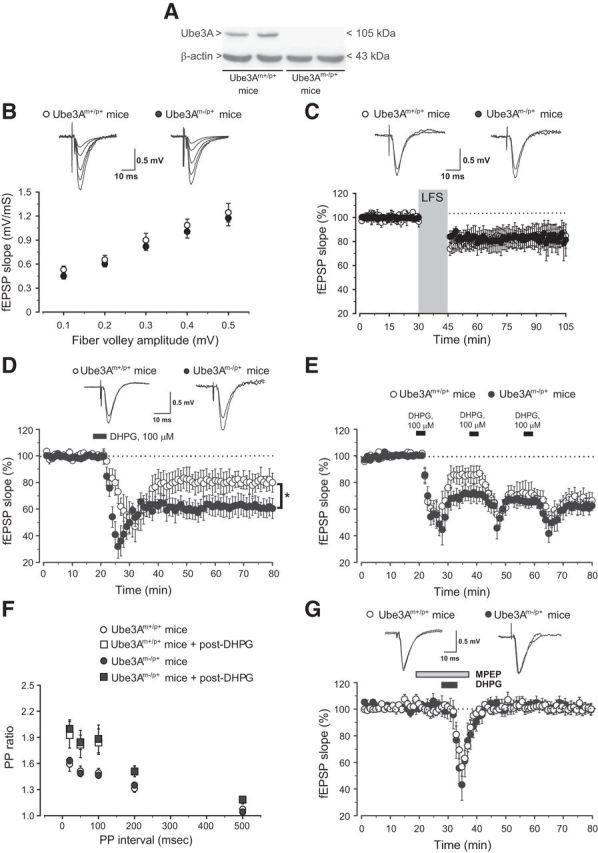
Enhanced mGlu5 receptor-dependent LTD in Ube3Am−/p+ mice. A, Immunoblot analysis of Ube3A in hippocampal slices from Ube3Am+/p+ (wild-type) mice and Ube3Am−/p+ mice. B, Input–output relation of fEPSPs as a function of presynaptic fiber volley size at the Schaffer collateral/CA1 pyramidal cell synapses. Each plot represents 7–8 separate recordings for each strain. Superimposed representative fEPSPs evoked in response to increasing stimulus intensity are shown. C, LTD induced by low-frequency stimulation (LFS; 1 Hz, 15 min) of Schaffer collaterals. The fEPSP slope (mean ± SEM) is plotted as percentage of the pre-LFS baseline. Insets show fEPSPs from a representative experiment during a baseline interval and 60 min after LTD. D, LTD induced by bath application of DHPG (100 μm, 5 min). Values are means ± SEM of data obtained from slices of 9–12 mice for each strain. *p < 0.05 (2-tailed unpaired Student's t test) versus values obtained in slices from wild-type mice. E, Depression of fEPSP induced by two consecutive applications of DHPG in slices from wild-type and Ube3Am−/p+ mice. Values are means ± SEM of data obtained from eight mice for each strain. F, PPF induced by pairs of stimulation delivered at several interstimulus intervals (20, 50, 100, 200, 500 ms) at baseline and 60 min after DHPG application. Data (means ± SEM) are expressed as the ratio between the second and the first response. G, Synaptic depression induced by DHPG (100 μm, 5 min) in the presence of MPEP (10 μm) in slices from wild-type and Ube3A mice. Values are means ± SEM of data obtained from 7–8 mice for each strain.
We performed pharmacological studies to dissect the relative contribution of mGlu1 and mGlu5 receptors in DHPG-induced LTD in the two genotypes. The mGlu5 receptor NAM MPEP (10 μm) abolished DHPG-induced LTD in both genotypes (wild-type: 98 ± 3%, n = 7; AS: 98 ± 4%, n = 8; Fig. 1G; for data with MPEP in normal mice, see Faas et al., 2002; Hou and Klann, 2004; Volk et al., 2006). In contrast, DHPG-induced LTD was unaffected by the mGlu1 receptor antagonist LY367385 (3 μm) in both wild-type and AS mice (data not shown). Thus, activation of mGlu5 receptors mediated DHPG-induced LTD in both genotypes.
Knowing that both DHPG-induced and NMDA receptor-dependent LTD are affected by ubiquitination inhibitors (Citri et al., 2009), we induced LTD in slices preincubated for 1 h with the proteasome inhibitor UBE1-41 (50 μm). DHPG-induced LTD was amplified by UBE1-41 in slices from wild-type mice during the first 40 min after DHPG (Fig. 2A). In contrast, UBE1-41 did not affect DHPG-induced LTD in slices from AS mice (Fig. 2B), indicating that the action of the proteasome inhibitor was occluded by the lack of Ube3A. We also examined NMDA receptor-dependent LTD induced by low-frequency stimulation in AS mice, finding no effect of UBE1-41 application (Fig. 2C).
Figure 2.
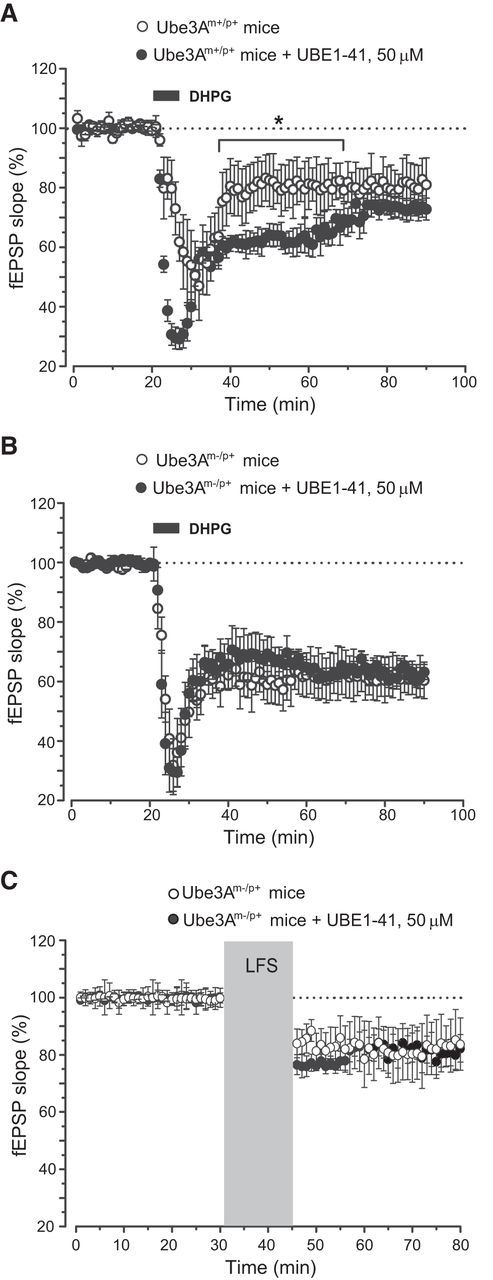
Pharmacological inhibition of proteasomal degradation did not affect synaptic plasticity in AS mice. Slices were preincubated with proteasome inhibitor UBE1-41 for 60 min before recording. A, Amplification of DHPG-induced LTD by UBE1-41 in hippocampal slices from wild-type mice. Values are means ± SEM of data obtained from four mice per group. *p < 0.05 (2-tailed unpaired Student's t test). B, C, The lack of effect of UBE1-41 on DHPG-induced (B) or low-frequency stimulation (LFS)-induced (C) LTD in slices from AS mice. Values are means ± SEM of data obtained from four mice per group.
Examination of the signaling pathways mediating the enhanced mGlu5 receptor-dependent LTD in AS mice
We first examined whether DHPG-induced LTD in wild-type and AS mice under our experimental conditions was sensitive to the protein synthesis inhibitor anysomicin (20 μm). This treatment abolished DHPG-induced LTD in both genotypes. (Fig. 3A).
Figure 3.
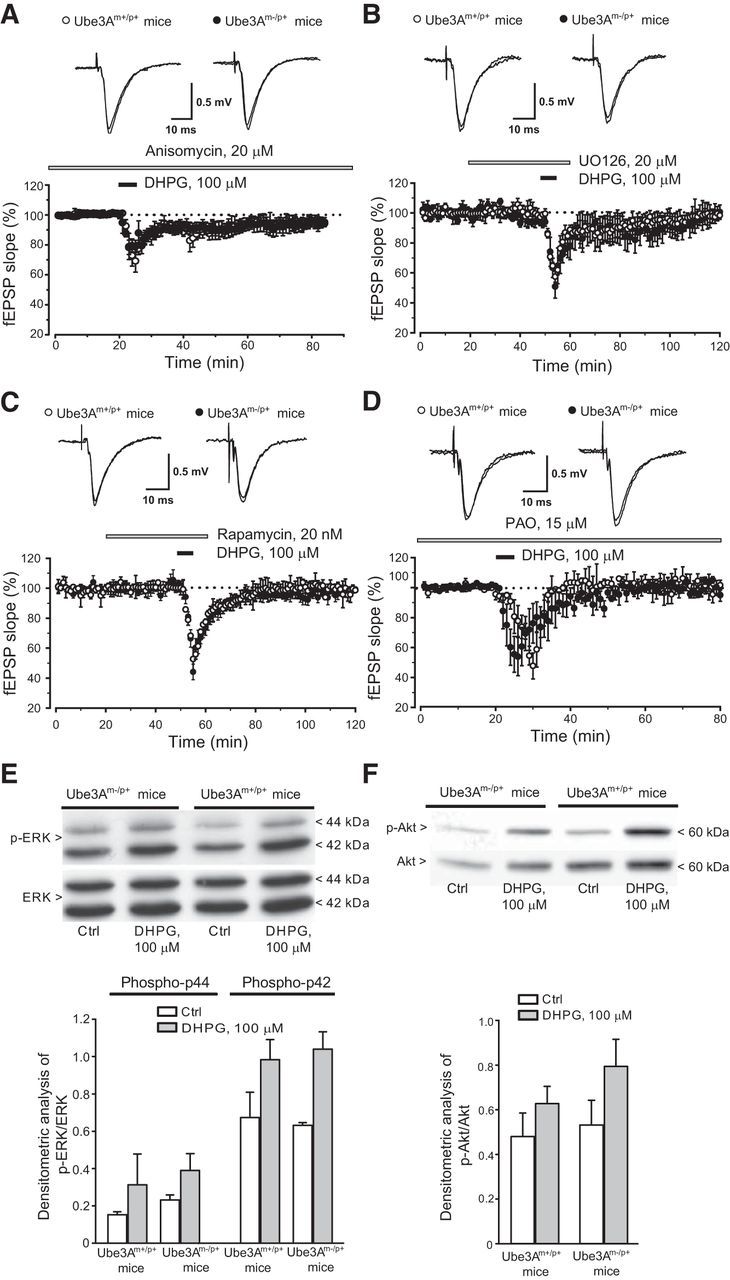
Examination of the intracellular signaling pathways mediating mGlu5 receptor-dependent LTD in hippocampal slices from wild-type and Ube3Am−/p+ mice. A–D, Depression of fEPSP induced by DHPG in the presence of anisomycin (A), UO126 (B), rapamycin (C), and PAO (D) in slices from wild-type and Ube3Am−/p+ mice. Values are means ± SEM of data obtained from 4–5 mice for each strain. E, F, DHPG-stimulated MAPK (E) and PI3K (F) pathways in slices from the two genotypes. Values are means ± SEM from slices obtained from 3–4 individual mice. Two-way ANOVA analysis of p-ERK and p-Akt data showed a drug effect (F(3,28) = 0.355 and F(3,12) = 0.396, respectively) but not genotype effect or drug–genotype interaction.
Multiple intracellular signaling pathways, including the mitogen-activated protein kinase (MAPK) pathway, the phosphatidylinositol-3-kinase (PI3K)/Akt/mTOR pathway, and tyrosine phosphatase (PTP)-dependent pathways, are involved in mGlu5-receptor-dependent LTD in the hippocampus (for review, see Gladding et al., 2009; Collingridge et al., 2010; Lüscher and Huber, 2010). We examined the involvement of these three pathways by inducing mGlu5 receptor-dependent LTD in the presence of the PTP inhibitor PAO (15 μm), the ERK1/2 kinase inhibitor U0126 (20 μm), or the mTOR inhibitor rapamycin (20 nm). Treatment of hippocampal slices with each of these inhibitors had no effect on basal synaptic transmission but fully blocked DHPG-induced LTD in both wild-type and AS mice (Fig. 3B–D; p > 0.05). In addition, all these treatments did not reverse changes in PPF induced by DHPG (data not shown). These data suggest that mGlu5 receptor-dependent LTD has the same molecular requirements in the two genotypes. In addition, DHPG-induced phosphorylation of ERK1/2 and Akt in hippocampal slices did not differ significantly between wild-type and AS mice (Fig. 3E,F). It was still possible that Ube3A-target proteins that are regulated by the MAPK or PI3K/Akt/mTOR pathways in response to mGlu5 receptor activation could be altered in AS mice. We measured the expression of Arc, the product of an early inducible gene that has been implicated in mechanisms of mGlu5 receptor-dependent LTD (Park et al., 2008; Waung et al., 2008). Basal Arc protein levels did not change in hippocampal slices from AS mice (Fig. 4; Greer et al., 2010). A 5 min exposure of hippocampal slices to DHPG (100 μm) increased Arc protein levels to the same extent in wild-type and AS mice (Fig. 4).
Figure 4.

Stimulation of Arc expression by DHPG in hippocampal slices from wild-type and Ube3Am−/p+ mice. Representative immunoblots are shown. Values are means ± SEM of data obtained from slices of three mice. Two-way ANOVA showed a drug effect at 5 min (F(3,12) = 0.412) and not genotype effect or drug–genotyping interaction. DHPG had no significant effect at 60 min.
Enhanced coupling of mGlu5 receptors with the long isoforms of Homer proteins in the hippocampus of AS mice
We next examined the possibility that the enhancement of mGlu5 receptor-dependent LTD could rely on mechanisms that lie upstream in the signal propagation. mGlu5 receptor mRNA and protein levels were not altered in the hippocampus of AS mice (Fig. 5A,B). We extended the analysis to agonist-stimulated PI hydrolysis, which represents the canonical signal transduction pathway activated by mGlu5 receptors (for review, see Nicoletti et al., 2011). DHPG enhanced [3H]InsP formation to the same extent in hippocampal slices prepared from wild-type and AS mice (Fig. 5C). Interaction between mGlu5 receptors and Homer proteins (Tu et al., 1998; Xiao et al., 1998) has been implicated in LTD induction (Ronesi and Huber, 2008; Takayasu et al., 2010; Ronesi et al., 2012). Interestingly, mRNA and protein levels of the short, activity-induced Homer 1a isoform were reduced in the hippocampus of AS mice, whereas levels of the long, constitutive Homer 1b/c isoforms were unchanged (Fig. 5D,E). We also measured mGlu5 receptor protein levels in hippocampal protein immunoprecipitated with either pan-Homer or Homer 1b/c antibodies. mGlu5 receptor levels were significantly increased in pan-Homer immunoprecipitates from AS mice, compared with wild-type mice (Fig. 5F). A significant increase in mGlu5 receptor levels was also found in Homer 1b/c immunoprecipitates from AS mice (Fig. 5G).
Figure 5.
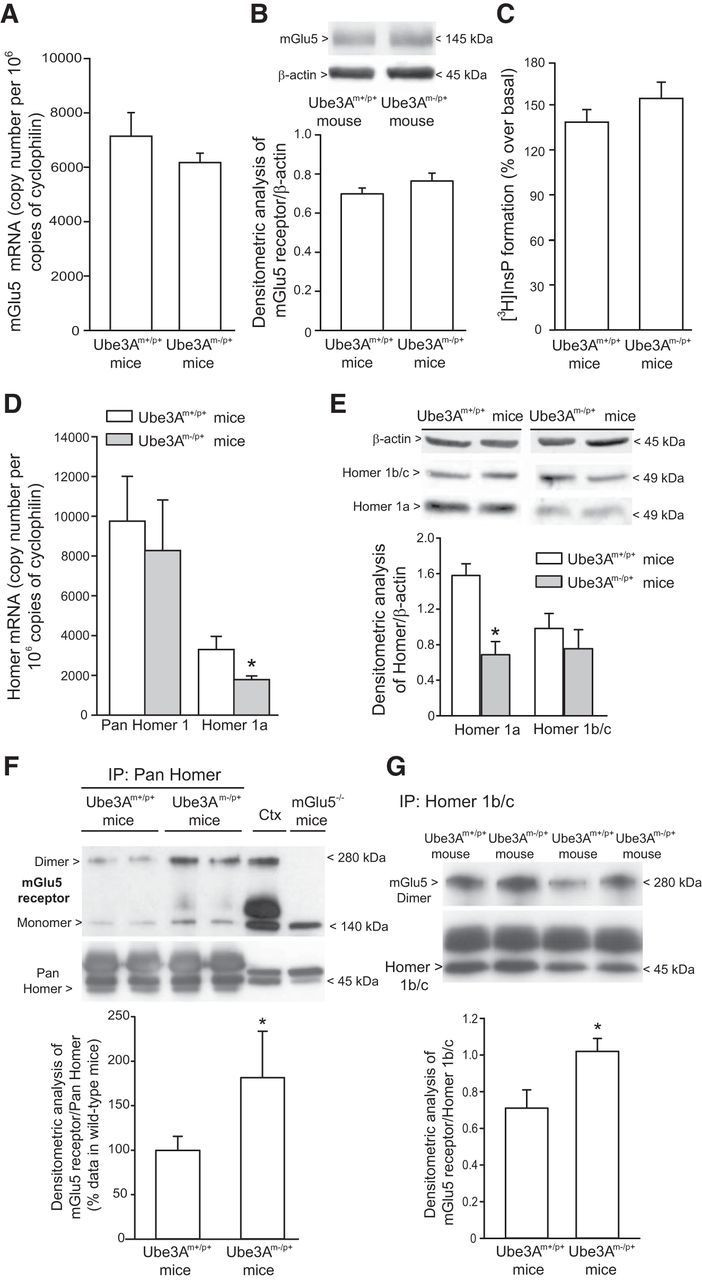
Changes in mGlu5 receptor coupling to Homer proteins in the hippocampus of Ube3Am−/p+ mice. A, B, mGlu5 receptor mRNA and protein levels in the hippocampus of wild-type (A) and Ube3Am−/p+ (B) mice. Values are means ± SEM of 4–8 mice per group. C, DHPG-stimulated inositol phospholipid hydrolysis in hippocampal slices. Values are means ± SEM and were obtained from slices obtained from five mice (here, slices were pooled and the experiment was performed in triplicate). The experiment was repeated twice with identical results. D, mRNA levels of pan-Homer and Homer 1a. E, Homer 1b/c and Homer 1a protein levels. Values are means ± SEM of four mice per group. *p < 0.05 versus the respective wild-type (Ube3Am+/p+) mice (Student's t test; t values: D, 4.42; E, 3.915). F, G, Levels of mGlu5 receptors in pan-Homer (F) and Homer 1b/c (G) immunoprecipitates. In F, values (n = 6–7) were calculated from two independent experiments and data are expressed as percentage of values (means ± SEM) obtained in wild-type mice. *p < 0.05 versus wild-type mice values (Student's t test; t values, −2.2895). In G, values (n = 8 mice per group) are means ± SEM *p < 0.05 (Student's t test; t value, −2.64).
Discussion
Drug treatment of AS remains an unmet clinical need, and pharmacological options to control symptoms of the disease have been only partially effective. Ube3A has been implicated in the regulation of activity-dependent synaptic plasticity (Jiang et al., 1998; Weeber et al., 2003), but its role in mGlu receptor-dependent forms of synaptic plasticity is unexplored. Here, we have shown that mGlu5 receptor-dependent LTD was enhanced in the hippocampus of AS mice, and this was associated with alterations in mGlu5 receptor coupling with Homer proteins. mGlu5 receptor-dependent LTD was also enhanced by the proteasome inhibitor UBE1-41, as expected (Citri et al., 2009), and the action of UBE1-41 was occluded in AS mice. Thus, the enhancement of mGlu5 receptor-dependent LTD in AS mice can be ascribed to the impairment of the ubiquitin/proteasome system. Changes in mGlu receptor-dependent LTD in the hippocampus of AS mice were specific because LTD induced by low-frequency stimulation (e.g., NMDA receptor-dependent LTD) was unaltered. This contrasts with the finding of a reduced NMDA receptor-dependent LTD in the visual cortex of AS mice (Yashiro et al., 2009). We highlight that DHPG-induced LTD under our experimental conditions was insensitive to NMDA receptor blockade, as expected.
LTD mediated by group I mGlu receptors at the Schaffer collateral–CA1 synapses requires dendritic protein synthesis (Waung and Huber, 2009). In FXS mice, LTD is enhanced and becomes independent of new protein synthesis because of the lack of FMRP, which normally restrains translation of LTD-related proteins (Huber et al., 2002; Hou et al., 2006; Nosyreva and Huber, 2006). In apparent contrast with these findings, the mGlu5 receptor-dependent LTD in AS mice was sensitive to the protein synthesis inhibitor anisomycin to the same extent as in control mice. Thus, although the defect of Ube3A is expected to prolong the half-life of postsynaptic proteins, mGlu receptor-dependent LTD in AS mice retains its sensitivity to de novo protein synthesis.
Arc, which is the product of an immediate early gene, has been directly related to mechanisms of LTD mediated by group I mGlu receptors. Rapid translation of Arc mediates mGlu1/5 receptor-dependent LTD in hippocampal neurons through a persistent increase in the rate of AMPA receptor endocytosis (Waung et al., 2008). FXS mice show increased Arc levels in dendrites, and lentiviral-mediated expression of FMRP in these mice normalizes both Arc levels and LTD in the hippocampus (Niere et al., 2012). We expected to find changes in Arc levels in the hippocampus of AS mice because Arc is a substrate for Ube3A (Greer et al., 2010). In contrast, Arc levels did not differ between wild-type and AS hippocampal slices under the same conditions used for the induction of LTD. However, we cannot exclude the possibility that differences in Arc levels between wild-type and AS mice are present, but anatomically restricted and too small to be revealed by immunoblot analysis.
Recent data suggest that in addition to triggering protein degradation, ubiquitination can modify protein–protein interactions and protein localization and activity (Hicke, 2001; DiAntonio and Hicke, 2004; Chen and Sun, 2009). We therefore took steps to find at which level the lack of Ube3A could affect the propagation of mGlu5 receptor signaling. Using specific pharmacological inhibitors, we showed that mGlu5 receptor-dependent LTD in AS mice relied on the same signaling pathways that mediate LTD in wild-type mice, i.e., the PTP, MAPK, and PI3K/mTOR pathways (Gladding et al., 2009; Collingridge et al., 2010; Lüscher and Huber, 2010). Stimulation of at least the MAPK and PI3K pathways by DHPG was unaltered in AS mice, suggesting that changes in the activity of these pathways are not responsible for the enhanced mGlu5 receptor-dependent LTD. mGlu5 receptor expression and mGlu5 receptor-mediated PI hydrolysis were also unaltered in AS mice.
AS mice differed from wild-type mice in the coupling mechanism of mGlu5 receptors to Homer proteins. Long, constitutive isoforms of Homer proteins (Homer 1b, 1c, 2, and 3) multimerize through their C-terminal coiled-coil domains and target mGlu1a and mGlu5 receptors to the postsynaptic density through interactions with SHANK (SH3 and multiple ankyrin repeat domains protein). In addition, long isoforms of Homer link mGlu1a and mGlu5 receptors to signaling molecules, such as PIKE (phosphoinositide-3 kinase enhancer), EF2K (the elongation factor 2 kinase), the inositol-1,4,5-trisphosphate receptors TRPC1 and TRPC3, N-type calcium channels, and M-type potassium channels (Brakeman et al., 1997; Tu et al., 1998, 1999; Xiao et al., 1998; Kammermeier et al., 2000; Yuan et al., 2003; Kim et al., 2006). In contrast, Homer 1a, a short and activity-inducible form of Homer lacking the coiled-coil domain, acts as a dominant negative isoform by uncoupling mGlu1a or mGlu5 receptors from postsynaptic effectors (Kammermeier and Worley, 2007).
Recent evidence links Homer proteins to mGlu5 receptor-mediated synaptic plasticity and autism. Disruption of mGlu5 interaction with Homer proteins blocks mGlu5 receptor-dependent LTD and protein synthesis in normal mice (Ronesi et al., 2012). In FXS mice, mGlu5 receptors are less associated with the long Homer isoforms and more associated with Homer 1a (Giuffrida et al., 2005; Ronesi et al., 2012). Genetic deletion of Homer 1a corrects several phenotypes in FXS mice, but not the enhancement of mGlu5 receptor-dependent LTD in the hippocampus (Ronesi et al., 2012). The gene encoding for Homer 1 has been identified as a novel risk gene for nonsyndromic autism. Rare Homer 1 gene variants that potentially affect protein function cosegregate closely with autism among children of affected families (Kelleher et al., 2012).
In AS mice, changes in the coupling of mGlu5 receptors to Homer proteins were opposite to those seen in FXS mice. AS mice showed reduced Homer 1a mRNA and protein levels, and increased association of mGlu5 receptors with Homer proteins in immunoprecipitates. The reduction of Homer 1a in AS mice was unexpected because Homer 1a is a substrate for ubiquitination, and proteasome inhibitors are known to enhance Homer 1a levels (Ageta et al., 2001). Perhaps the lack of Ube3A enhances the stability of a negative regulator of Homer 1a, which may function at transcriptional or translational levels. Alternatively, in AS mice the Homer 1a phenotype may lay upstream of the proteasome system, which may help to explain why Homer 1a levels are not enhanced despite the lack of Ube3A. It will be interesting to examine whether an enhanced coupling of mGlu5 receptors to long Homer isoforms has any influence on the efficiency of the ubiquitine/proteasomal system. Whatever the mechanism, the reduction of Homer 1a suggests that mGlu5 receptors are more efficiently coupled to postsynaptic effectors in AS mice. This fits nicely with the enhanced mGlu5 receptor-dependent LTD found in AS mice, although the precise mechanism that is ultimately responsible for the amplification of LTD is unknown. It is intriguing that in FXS and AS mice opposite changes in mGlu5/Homer coupling are associated with an enhanced mGlu5 receptor-dependent LTD. One should take into account the fact that a reduced association of mGlu5 receptors to long Homer isoforms restrains receptor coupling to postsynaptic effectors on one side (Kammermeier and Worley, 2007), but enhances the agonist-independent “constitutive” activity of mGlu5 receptors on the other side (Ango et al., 2001). Whether changes in the constitutive activity of mGlu5 receptors have any role in the pathological phenotype of AS mice is unknown.
In conclusion, we have described for the first time the association of AS with abnormalities in the interaction between mGlu5 receptors and Homer proteins, and an enhancement of mGlu5 receptor-dependent LTD in the hippocampus. These findings lay the groundwork for the use of mGlu5 receptor antagonists in models of AS.
Footnotes
We thank Prof. P. Worley, Department of Neuroscience, Johns Hopkins University School of Medicine, Baltimore, MD, for kindly providing Arc antibody.
The authors declare no competing financial interests.
References
- Ageta H, Kato A, Hatakeyama S, Nakayama K, Isojima Y, Sugiyama H. Regulation of the level of Vesl-1S/Homer-1a proteins by ubiquitin-proteasome proteolytic systems. J Biol Chem. 2001;276:15893–15897. doi: 10.1074/jbc.M011097200. [DOI] [PubMed] [Google Scholar]
- Ango F, Prézeau L, Muller T, Tu JC, Xiao B, Worley PF, Pin JP, Bockaert J, Fagni L. Agonist-independent activation of metabotropic glutamate receptors by the intracellular protein Homer. Nature. 2001;411:962–965. doi: 10.1038/35082096. [DOI] [PubMed] [Google Scholar]
- Auerbach BD, Osterweil EK, Bear MF. Mutations causing syndromic autism define an axis of synaptic pathophysiology. Nature. 2011;480:63–68. doi: 10.1038/nature10658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bear MF, Huber KM, Warren ST. The mGluR theory of fragile X mental retardation. Trends Neurosci. 2004;27:370–377. doi: 10.1016/j.tins.2004.04.009. [DOI] [PubMed] [Google Scholar]
- Bhakar AL, Dölen G, Bear MF. The pathophysiology of fragile X (and what it teaches us about synapses) Annu Rev Neurosci. 2012;35:417–443. doi: 10.1146/annurev-neuro-060909-153138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brakeman PR, Lanahan AA, O'Brien R, Roche K, Barnes CA, Huganir RL, Worley PF. Homer: a protein that selectively binds metabotropic glutamate receptors. Nature. 1997;386:284–288. doi: 10.1038/386284a0. [DOI] [PubMed] [Google Scholar]
- Chen ZJ, Sun LJ. Nonproteolytic functions of ubiquitin in cell signaling. Mol Cell. 2009;33:275–286. doi: 10.1016/j.molcel.2009.01.014. [DOI] [PubMed] [Google Scholar]
- Citri A, Soler-Llavina G, Bhattacharyya S, Malenka RC. N-methyl-d-aspartate receptor- and metabotropic glutamate receptor-dependent long-term depression are differentially regulated by the ubiquitin-proteasome system. Eur J Neurosci. 2009;30:1443–1450. doi: 10.1111/j.1460-9568.2009.06950.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collingridge GL, Peineau S, Howland JG, Wang YT. Long-term depression in the CNS. Nat Rev Neurosci. 2010;11:459–473. doi: 10.1038/nrn2867. [DOI] [PubMed] [Google Scholar]
- DiAntonio A, Hicke L. Ubiquitin-dependent regulation of the synapse. Annu Rev Neurosci. 2004;27:223–246. doi: 10.1146/annurev.neuro.27.070203.144317. [DOI] [PubMed] [Google Scholar]
- Dölen G, Osterweil E, Rao BS, Smith GB, Auerbach BD, Chattarji S, Bear MF. Correction of fragile X syndrome in mice. Neuron. 2007;56:955–962. doi: 10.1016/j.neuron.2007.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong C, Upadhya SC, Ding L, Smith TK, Hegde AN. Proteasome inhibition enhances the induction and impairs the maintenance of late-phase long-term potentiation. Learn Mem. 2008;15:335–347. doi: 10.1101/lm.984508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehlers MD. Activity level controls postsynaptic composition and signaling via the ubiquitin-proteasome system. Nat Neurosci. 2003;6:231–342. doi: 10.1038/nn1013. [DOI] [PubMed] [Google Scholar]
- Faas GC, Adwanikar H, Gereau RW, 4th, Saggau P. Modulation of presynaptic calcium transients by metabotropic glutamate receptor activation: a differential role in acute depression of synaptic transmission and long-term depression. J Neurosci. 2002;22:6885–6890. doi: 10.1523/JNEUROSCI.22-16-06885.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng Y, Zhang F, Lokey LK, Chastain JL, Lakkis L, Eberhart D, Warren ST. Translational suppression by trinucleotide repeat expansion at FMR1. Science. 1995;268:731–734. doi: 10.1126/science.7732383. [DOI] [PubMed] [Google Scholar]
- Giuffrida R, Musumeci S, D'Antoni S, Bonaccorso CM, Giuffrida-Stella AM, Oostra BA, Catania MV. A reduced number of metabotropic glutamate subtype 5 receptors are associated with constitutive Homer proteins in a mouse model of fragile X syndrome. J Neurosci. 2005;25:8908–89016. doi: 10.1523/JNEUROSCI.0932-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gladding CM, Fitzjohn SM, Molnár E. Metabotropic glutamate receptor-mediated long-term depression: molecular mechanisms. Pharmacol Rev. 2009;61:395–412. doi: 10.1124/pr.109.001735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer PL, Hanayama R, Bloodgood BL, Mardinly AR, Lipton DM, Flavell SW, Kim TK, Griffith EC, Waldon Z, Maehr R, Ploegh HL, Chowdhury S, Worley PF, Steen J, Greenberg ME. The Angelman syndrome protein Ube3A regulates synapse development by ubiquitinating arc. Cell. 2010;140:704–716. doi: 10.1016/j.cell.2010.01.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagerman R, Lauterborn J, Au J, Berry-Kravis E. Fragile X syndrome and targeted treatment trials. Results Probl Cell Differ. 2012;54:297–335. doi: 10.1007/978-3-642-21649-7_17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hicke L. Protein regulation by monoubiquitin. Nat Rev Mol Cell Biol. 2001;2:195–201. doi: 10.1038/35056583. [DOI] [PubMed] [Google Scholar]
- Hou L, Klann E. Activation of the phosphoinositide 3-kinase-Akt-mammalian target of rapamycin signaling pathway is required for metabotropic glutamate receptor-dependent long-term depression. J Neurosci. 2004;24:6352–6361. doi: 10.1523/JNEUROSCI.0995-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou L, Antion MD, Hu D, Spencer CM, Paylor R, Klann E. Dynamic translational and proteasomal regulation of fragile X mental retardation protein controls mGluR-dependent long-term depression. Neuron. 2006;51:441–454. doi: 10.1016/j.neuron.2006.07.005. [DOI] [PubMed] [Google Scholar]
- Huber KM, Kayser MS, Bear MF. Role for rapid dendritic protein synthesis in hippocampal mGluR-dependent long-term depression. Science. 2000;288:1254–1257. doi: 10.1126/science.288.5469.1254. [DOI] [PubMed] [Google Scholar]
- Huber KM, Roder JC, Bear MF. Chemical induction of mGluR5- and protein synthesis-dependent long-term depression in hippocampal area CA1. J Neurophysiol. 2001;86:321–325. doi: 10.1152/jn.2001.86.1.321. [DOI] [PubMed] [Google Scholar]
- Huber KM, Gallagher SM, Warren ST, Bear MF. Altered synaptic plasticity in a mouse model of fragile X mental retardation. Proc Natl Acad Sci U S A. 2002;99:7746–7750. doi: 10.1073/pnas.122205699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang YH, Armstrong D, Albrecht U, Atkins CM, Noebels JL, Eichele G, Sweatt JD, Beaudet AL. Mutation of the Angelman ubiquitin ligase in mice causes increased cytoplasmic p53 and deficits of contextual learning and long-term potentiation. Neuron. 1998;21:799–811. doi: 10.1016/S0896-6273(00)80596-6. [DOI] [PubMed] [Google Scholar]
- Kammermeier PJ, Worley PF. Homer 1a uncouples metabotropic glutamate receptor 5 from postsynaptic effectors. Proc Natl Acad Sci U S A. 2007;104:6055–6060. doi: 10.1073/pnas.0608991104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kammermeier PJ, Xiao B, Tu JC, Worley PF, Ikeda SR. Homer proteins regulate coupling of group I metabotropic glutamate receptors to N-type calcium and M-type potassium channels. J Neurosci. 2000;20:7238–7245. doi: 10.1523/JNEUROSCI.20-19-07238.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelleher RJ, 3rd, Bear MF. The autistic neuron: troubled translation? Cell. 2008;135:401–406. doi: 10.1016/j.cell.2008.10.017. [DOI] [PubMed] [Google Scholar]
- Kelleher RJ, 3rd, Geigenmüller U, Hovhannisyan H, Trautman E, Pinard R, Rathmell B, Carpenter R, Margulies D. High-throughput sequencing of mGluR signaling pathway genes reveals enrichment of rare variants in autism. PLoS One. 2012;7:e35003. doi: 10.1371/journal.pone.0035003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim JY, Zeng W, Kiselyov K, Yuan JP, Dehoff MH, Mikoshiba K, Worley PF, Muallem S. Homer 1 mediates store- and inositol 1,4,5-trisphosphate receptor-dependent translocation and retrieval of TRPC3 to the plasma membrane. J Biol Chem. 2006;281:32540–32549. doi: 10.1074/jbc.M602496200. [DOI] [PubMed] [Google Scholar]
- Kishino T, Lalande M, Wagstaff J. UBE3A/E6-AP mutations cause Angelman syndrome. Nat Genet. 1997;15:70–73. doi: 10.1038/ng0197-70. [DOI] [PubMed] [Google Scholar]
- Krueger DD, Bear MF. Toward fulfilling the promise of molecular medicine in fragile X syndrome. Annu Rev Med. 2011;62:411–429. doi: 10.1146/annurev-med-061109-134644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lüscher C, Huber KM. Group 1 mGluR-dependent synaptic long-term depression: mechanisms and implications for circuitry and disease. Neuron. 2010;65:445–459. doi: 10.1016/j.neuron.2010.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manabe T. Two forms of hippocampal long-term depression, the counterpart of long-term potentiation. Rev Neurosci. 1997;8:179–193. doi: 10.1515/REVNEURO.1997.8.3-4.179. [DOI] [PubMed] [Google Scholar]
- Michalon A, Sidorov M, Ballard TM, Ozmen L, Spooren W, Wettstein JG, Jaeschke G, Bear MF, Lindemann L. Chronic pharmacological mGlu5 inhibition corrects fragile X in adult mice. Neuron. 2012;74:49–56. doi: 10.1016/j.neuron.2012.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moult PR, Corrêa SA, Collingridge GL, Fitzjohn SM, Bashir ZI. Co-activation of p38 mitogen-activated protein kinase and protein tyrosine phosphatase underlies metabotropic glutamate receptor-dependent long-term depression. J Physiol. 2008;586:2499–2510. doi: 10.1113/jphysiol.2008.153122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicoletti F, Iadarola MJ, Wroblewski JT, Costa E. Excitatory amino acid recognition sites coupled with inositol phospholipid metabolism: developmental changes and interaction with α1-adrenoceptors. Proc Natl Acad Sci U S A. 1986;83:1931–1935. doi: 10.1073/pnas.83.6.1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicoletti F, Bockaert J, Collingridge GL, Conn PJ, Ferraguti F, Schoepp DD, Wroblewski JT, Pin JP. Metabotropic glutamate receptors: from the workbench to the bedside. Neuropharmacology. 2011;60:1017–1041. doi: 10.1016/j.neuropharm.2010.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niere F, Wilkerson JR, Huber KM. Evidence for a fragile X mental retardation protein-mediated translational switch in metabotropic glutamate receptor-triggered Arc translation and long-term depression. J Neurosci. 2012;32:5924–5936. doi: 10.1523/JNEUROSCI.4650-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nisticò R, Mango D, Mandolesi G, Piccinin S, Berretta N, Pignatelli M, Feligioni M, Musella A, Gentile A, Mori F, Bernardi G, Nicoletti F, Mercuri NB, Centonze D. Inflammation subverts hippocampal synaptic plasticity in experimental multiple sclerosis. PLoS One. 2013;8:e54666. doi: 10.1371/journal.pone.0054666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nosyreva ED, Huber KM. Metabotropic receptor-dependent long-term depression persists in the absence of protein synthesis in the mouse model of fragile X syndrome. J Neurophysiol. 2006;95:3291–3295. doi: 10.1152/jn.01316.2005. [DOI] [PubMed] [Google Scholar]
- Park S, Park JM, Kim S, Kim JA, Shepherd JD, Smith-Hicks CL, Chowdhury S, Kaufmann W, Kuhl D, Ryazanov AG, Huganir RL, Linden DJ, Worley PF. Elongation factor 2 and fragile X mental retardation protein control the dynamic translation of Arc/Arg3.1 essential for mGluR-LTD. Neuron. 2008;59:70–83. doi: 10.1016/j.neuron.2008.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronesi JA, Huber KM. Homer interactions are necessary for metabotropic glutamate receptor-induced long-term depression and translational activation. J Neurosci. 2008;28:543–547. doi: 10.1523/JNEUROSCI.5019-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronesi JA, Collins KA, Hays SA, Tsai NP, Guo W, Birnbaum SG, Hu JH, Worley PF, Gibson JR, Huber KM. Disrupted Homer scaffolds mediate abnormal mGluR5 function in a mouse model of fragile X syndrome. Nat Neurosci. 2012;15:431–440. S1. doi: 10.1038/nn.3033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato M, Stryker MP. Genomic imprinting of experience-dependent cortical plasticity by the ubiquitin ligase gene Ube3a. Proc Natl Acad Sci U S A. 2010;107:5611–5616. doi: 10.1073/pnas.1001281107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steffenburg S, Gillberg CL, Steffenburg U, Kyllerman M. Autism in Angelman syndrome: a population-based study. Pediatr Neurol. 1996;14:131–136. doi: 10.1016/0887-8994(96)00011-2. [DOI] [PubMed] [Google Scholar]
- Takayasu Y, Takeuchi K, Kumari R, Bennett MV, Zukin RS, Francesconi A. Caveolin-1 knockout mice exhibit impaired induction of mGluR-dependent long-term depression at CA3-CA1 synapses. Proc Natl Acad Sci U S A. 2010;107:21778–21783. doi: 10.1073/pnas.1015553107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tu JC, Xiao B, Yuan JP, Lanahan AA, Leoffert K, Li M, Linden DJ, Worley PF. Homer binds a novel proline-rich motif and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron. 1998;21:717–726. doi: 10.1016/S0896-6273(00)80589-9. [DOI] [PubMed] [Google Scholar]
- Tu JC, Xiao B, Naisbitt S, Yuan JP, Petralia RS, Brakeman P, Doan A, Aakalu VK, Lanahan AA, Sheng M, Worley PF. Coupling of mGluR/Homer and PSD-95 complexes by the Shank family of postsynaptic density proteins. Neuron. 1999;23:583–592. doi: 10.1016/S0896-6273(00)80810-7. [DOI] [PubMed] [Google Scholar]
- Volk LJ, Daly CA, Huber KM. Differential roles for group 1 mGluR subtypes in induction and expression of chemically induced hippocampal long-term depression. J Neurophysiol. 2006;95:2427–2438. doi: 10.1152/jn.00383.2005. [DOI] [PubMed] [Google Scholar]
- Waung MW, Huber KM. Protein translation in synaptic plasticity: mGluR-LTD, Fragile X. Curr Opin Neurobiol. 2009;19:319–326. doi: 10.1016/j.conb.2009.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waung MW, Pfeiffer BE, Nosyreva ED, Ronesi JA, Huber KM. Rapid translation of Arc/Arg3.1 selectively mediates mGluR-dependent LTD through persistent increases in AMPAR endocytosis rate. Neuron. 2008;59:84–97. doi: 10.1016/j.neuron.2008.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weeber EJ, Jiang YH, Elgersma Y, Varga AW, Carrasquillo Y, Brown SE, Christian JM, Mirnikjoo B, Silva A, Beaudet AL, Sweatt JD. Derangements of hippocampal calcium/calmodulin-dependent protein kinase II in a mouse model for Angelman mental retardation syndrome. J Neurosci. 2003;23:2634–2644. doi: 10.1523/JNEUROSCI.23-07-02634.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams CA. Neurological aspects of the Angelman syndrome. Brain Dev. 2005;27:88–94. doi: 10.1016/j.braindev.2003.09.014. [DOI] [PubMed] [Google Scholar]
- Xiao B, Tu JC, Petralia RS, Yuan JP, Doan A, Breder CD, Ruggiero A, Lanahan AA, Wenthold RJ, Worley PF. Homer regulates the association of group 1 metabotropic glutamate receptors with multivalent complexes of Homer-related, synaptic proteins. Neuron. 1998;21:707–716. doi: 10.1016/S0896-6273(00)80588-7. [DOI] [PubMed] [Google Scholar]
- Yashiro K, Riday TT, Condon KH, Roberts AC, Bernardo DR, Prakash R, Weinberg RJ, Ehlers MD, Philpot BD. Ube3a is required for experience-dependent maturation of the neocortex. Nat Neurosci. 2009;12:777–783. doi: 10.1038/nn.2327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan JP, Kiselyov K, Shin DM, Chen J, Shcheynikov N, Kang SH, Dehoff MH, Schwarz MK, Seeburg PH, Muallem S, Worley PF. Homer binds TRPC family channels and is required for gating of TRPC1 by IP3 receptors. Cell. 2003;114:777–789. doi: 10.1016/S0092-8674(03)00716-5. [DOI] [PubMed] [Google Scholar]
- Zucker RS. Short-term synaptic plasticity. Annu Rev Neurosci. 1989;12:13–31. doi: 10.1146/annurev.ne.12.030189.000305. [DOI] [PubMed] [Google Scholar]