

Assessment of socioenvironmental benefits and feasibility identifies priority areas for restoring global tropical rainforests.
Abstract
Over 140 Mha of restoration commitments have been pledged across the global tropics, yet guidance is needed to identify those landscapes where implementation is likely to provide the greatest potential benefits and cost-effective outcomes. By overlaying seven recent, peer-reviewed spatial datasets as proxies for socioenvironmental benefits and feasibility of restoration, we identified restoration opportunities (areas with higher potential return of benefits and feasibility) in lowland tropical rainforest landscapes. We found restoration opportunities throughout the tropics. Areas scoring in the top 10% (i.e., restoration hotspots) are located largely within conservation hotspots (88%) and in countries committed to the Bonn Challenge (73%), a global effort to restore 350 Mha by 2030. However, restoration hotspots represented only a small portion (19.1%) of the Key Biodiversity Area network. Concentrating restoration investments in landscapes with high benefits and feasibility would maximize the potential to mitigate anthropogenic impacts and improve human well-being.
INTRODUCTION
Less than 50% of the world’s tropical forests remain standing today, with much of the remaining forest cover seriously affected by logging, fires, fragmentation, mining, and hunting (1, 2). Loss and degradation of tropical forests bring strong negative consequences for biodiversity, climate regulation, and well-being of rural and urban populations (3, 4). Both conservation and restoration are urgently needed to mitigate anthropogenic impacts on tropical forests and their contributions to people in terms of ecosystem services of importance for human well-being (5, 6). The biodiversity hotspots for conservation priorities (hereafter conservation hotspots) approach (7) was a turning point for global-scale conservation policies, research, and actions over the past 18 years by identifying priority regions for conservation efforts as those with at least 0.5% or 1500 species of vascular plants as endemics and less than 30% of their primary native vegetation remaining. Since then, the primary approach to conserving biodiversity has shifted from narrowly focused restoration and species protection within ecosystems, to a broader approach incorporating landscape-scale restoration to achieve multiple objectives (8), including reducing species extinctions (9, 10), mitigating injurious climate change (11), and promoting sustainable livelihoods (12).
Global conservation and sustainable development commitments, such as the Aichi Targets of the Convention on Biological Diversity, United Nations Sustainable Development Goals—particularly Goals 14.2, 15.1, and 15.3—the intended Nationally Determined Contributions to the Paris Climate Agreement, and the New York Declaration on Forests, rely heavily on restoration to achieve their objectives (13–15). Restoration at a landscape scale is challenging, as restoration efforts occur in the context of intense competition for land, where agriculture already occupies 37.3% of the global ice-free land surface (16) and is still increasing in extent (17). Guidance is urgently needed to direct effort toward the most cost-efficient restoration outcomes—largest gain per investment of time, money, and effort—and to identify landscapes where levels of multiple restoration benefits can be maximized. Identifying restoration opportunities—areas that combine high potential for socioenvironmental benefits with high restoration feasibility—can be an essential tool for achieving the ambitious restoration commitments planned for the immediate future. Identification of restoration opportunities and particularly those areas where restoration opportunities achieve their highest level—restoration hotspots—can be further combined with other decision-making factors to define priorities for implementation and financing of the global restoration agenda.
Here, we identify global restoration opportunities in lowland tropical rainforest landscapes by overlaying recent, peer-reviewed global-scale spatial datasets that serve as proxies for socioenvironmental benefits and feasibility of restoration, with direct and indirect consequences for nature, economies, and human well-being. Our analysis supports the implementation of forest and landscape restoration, which relies on a balance of different restorative strategies to regain ecological functionality and enhance human well-being in degraded and deforested landscapes (15, 18). For restoration benefits, we include four variables (see details in Methods): biodiversity conservation (habitat provision for vulnerable species, determined as the number of threatened and range-restricted vertebrate species), climate change mitigation (contribution to reduce CO2 concentration in the atmosphere, determined as the carbon sequestration in aboveground tree biomass), climate change adaptation (restoration as adaptation measure in regions where climate will change faster), and human water security (potential for reduction of water security risks). For restoration feasibility (see details in Methods), we include three variables: land opportunity costs (the costs associated with land-use change from agriculture to restoration), landscape variation in forest restoration success (the variability associated with biodiversity recovery in restored forests, a proxy for investment risks, and implementation costs of restoration), and restoration persistence chance (i.e., the relative rate of recent tree cover loss, which represent the chances that restored forests persist over time without reconversion to alternative land uses and also serve as a proxy of investment risks).
We applied this approach to regions with available carbon density maps for old-growth forests (19), which were needed to determine the carbon sequestration potential of restored forests in aboveground tree biomass, within the global coverage of tropical moist broadleaf forests (hereafter referred to as tropical rainforests), which harbors most of the global biodiversity, has many densely populated areas (2), and encompasses most of the Bonn Challenge pledges (20). We limited our approach to the moist forest biome because dry and moist forests differ widely in potential carbon storage and biodiversity (21), and combining them in a single analysis could lead to anomalous results.
Within the distribution region of tropical rainforest biomes, we identified restorable areas (i.e., landscapes where restoration efforts could be implemented) as those with elevation below 1000 m, with <90% tree canopy cover, and not covered by urban areas, water bodies, and wetlands. The global restorable area in tropical rainforest landscapes is 863 Mha, slightly larger than the area of Brazil, and comprises close to 60% of the total study area. As finer-scale global maps of other nonforest native ecosystems are not available (e.g., grasslands in mountain outcrops, savannas, and coastal shrublands), it was not possible to remove them from the coarser-scale resolution map of tropical moist broadleaf forests (22). Consequently, patches of nonforest ecosystems may be embedded within our study region. Finer-scale vegetation maps should be applied within these regions before detailed restoration planning and implementation to avoid planting trees in native nonforest ecosystems [see (23)]. The methodological approach developed here can be applied to other biomes or vegetation types with urgent needs of restoration, such as tropical and subtropical dry forests and Mediterranean forests, complementing the knowledge basis required to leverage the implementation of national and global restoration commitments.
Whereas the conservation hotspot approach (24, 25) considered whole ecosystems as hotspot and defined a priori thresholds for including an ecosystem as a hotspot, we based our analyses in grid cells of 30 arc sec (approximately 1 km2 at the equator) to produce a gradient of values ranging from 0 to 1 [hereafter referred as restoration opportunity score (ROS)], representing the normalized opportunity for maximizing benefits and feasibility of restoration. We further considered areas within the top ~10% ROS in a landscape as restoration hotspots. We defined the hotspots relative to space rather than subjective thresholds. This provides users to identify priorities corresponding to their study areas. The restoration hotspots at the global scale had ROS above 0.62. We explored their global distribution according to different contexts for decision-making [country, ecoregion, Key Biodiversity Areas (hereafter KBAs), and conservation hotspots; (22, 24, 26)]. We did not define a priori what restorative actions should be used within each landscape nor the extent or precise location of interventions. These decisions need to be made by restoration practitioners based on the local socioecological context and negotiation among multiple stakeholders (27).
RESULTS
The combined analysis of benefits (Fig. 1A) and feasibility of restoration (Fig. 1B) identified landscapes with different ROS distributed across the global tropics (Fig. 1C). Global ROS were normally distributed (fig. S1), and only 11.8% of the area had a ROS ≥ 0.6 (hereafter referred as restoration hotspots). The top six countries with the highest mean ROS were found in Africa: Rwanda, Uganda, Burundi, Togo, South Sudan, and Madagascar (table S1). The top 15 countries with the largest areas of restoration hotspots were found across all the biogeographical realms (three in Neotropics, five in Afrotropics, seven in Indo-Malaysia, and part of Indonesia in Australasia). Brazil, Indonesia, India, Madagascar, and Colombia are the five countries with the largest hotspot areas (Fig. 2A and table S1). The top 15 ecoregions with the largest area of restoration hotspots were also well distributed across biogeographical realms (six in Neotropics, four in Afrotropics, and five in Indo-Malaysia), with a third of them in Brazil (Fig. 2B and table S2). Eight of the top 10 KBAs with the largest area of restoration hotspots were found in Neotropics (five of them in Brazil) and the other two in Africa (Fig. 2C and table S3). However, only a small proportion of the restorable area within KBAs were restoration hotspots (19.1%), and the average ROS of landscapes within KBAs was lower (0.47) than those outside these areas (0.51).
Fig. 1. ROS of tropical rainforest landscapes in lowlands.
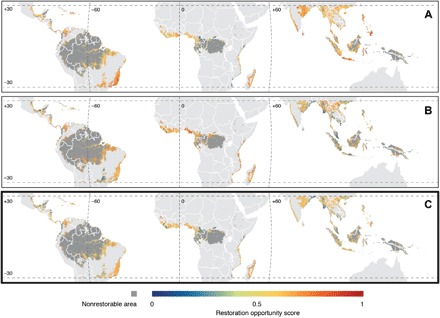
(A) Restoration benefits (biodiversity conservation, water security, climate change adaptation, and mitigation combined), (B) restoration feasibility (reduced land opportunity costs, reduced landscape variation in forest restoration success, and higher likelihood of forest persistence combined), and (C) benefits combined with feasibility of restoration. Higher ROS (values ranging from 0 to 1) represent landscapes with higher potential restoration benefits and feasibility. The depiction of boundaries and geographic names is simply for display purposes and does not imply views regarding the legal status of any territory or country.
Fig. 2. Top 10 countries, ecoregions, conservation hotspots, and KBAs with the largest area of restoration hotspots.
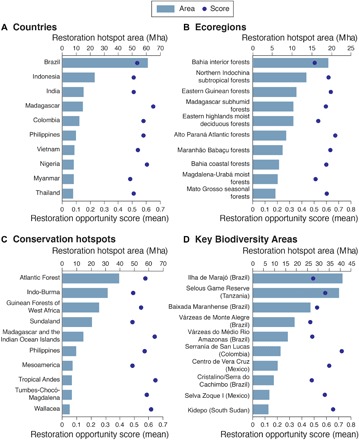
Total area of restoration hotspots [bars; million hectare meter (Mha)] and the mean ROS of all restorable areas (dots) within each country, ecoregion, conservation hotspot, and KBA in the study area.
The conservation hotspots with the highest mean ROS were Tropical Andes, Madagascar and the Indian Ocean Islands, and Eastern Afromontane (ROS ≥ 0.624; Fig. 2D and table S4). The Atlantic Forest, Indo-Burma, and Guinean Forests of West Africa were the conservation hotspots with the greatest restoration hotspot area (Figs. 2D and 3A and table S4), which also showed large areas of restoration hotspots within KBAs (Fig. 3B). The area of restoration hotspots was positively associated with the restorable area within countries (r2 = 0.87; P < 0.0001) and conservation hotspots (r2 = 0.84; P = 0.0001), but not for KBAs (r2 = 0.15; P = 0.14). The vast majority of restoration hotspots were found within conservation hotspots (88.6%) and within countries with Bonn Challenge commitments (73.0%; Fig. 3A). The renormalization of the ROS gradient for countries (fig. S2), biogeographical realms (fig. S3), and conservation hotspots (fig. S4) further enabled identification of priority tropical rainforest landscapes in lowlands for implementing restoration efforts at different scales and within different political, biogeographical, and conservation contexts. This renormalization process further permitted identification of the country area encompassing the top 15% ROS, which could guide implementation of the Aichi Target 15 of the Convention on Biological Diversity for rainforest biomes.
Fig. 3. Restoration hotspots, conservation hotspots, and Bonn Challenge commitments.
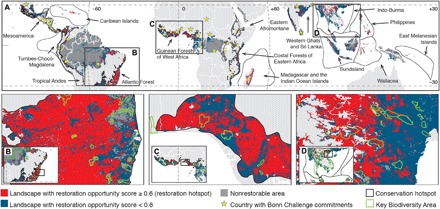
Spatial congruence between global hotspots for restoring tropical rainforest landscapes in lowlands and for biodiversity conservation in the global tropics (A), and between restoration hotspots and countries with restoration commitments to the Bonn Challenge (A). Expanded areas within the biodiversity conservation hotspots Atlantic Forest (B), Guinean Forests of West Africa (C), and Indo-Burma (D). The depiction of boundaries and geographic names is simply for display purposes and does not imply views regarding the legal status of any territory or country.
We found notable spatial matches and mismatches between benefits and feasibility of restoration at the global scale. The combined benefits and feasibility of restoration were weakly negatively correlated (r2 = −0.12; P < 0.0001; Fig. 4). Regarding restoration benefits, landscapes where forest restoration is expected to bring higher benefits for climate change mitigation tended to provide lower expected benefits for climate change adaptation (Fig. 4). Regarding restoration feasibility, landscapes with lower land opportunity costs and higher forest persistence chances showed higher landscape variation in forest restoration success (Fig. 4). When both benefits and feasibility of restoration are considered, landscapes where restoration has a higher potential to support biodiversity conservation mitigate and adapt to climate change and reduce water security risks generally correspond to areas of lower overall feasibility (Fig. 4).
Fig. 4. Spatial congruence among benefits and feasibility of restoration.
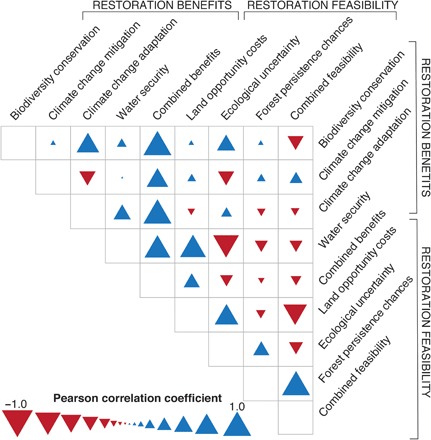
Pearson’s correlation coefficients for all combinations of benefits and feasibility of restoring tropical rainforest landscapes in lowlands.
DISCUSSION
Our multilayered approach to forest restoration opportunities signals a major advance toward identifying landscapes where interventions are expected to be more cost effective, based on locations where socioenvironmental benefits are maximized and investment costs and risks are minimized. Trade-offs between restoration benefits and feasibility are minimized in landscapes with higher ROS, particularly in the restoration hotspots. Landscapes with high ROS were ubiquitously distributed across the global tropics, highlighting the application of our fine-scale approach to guiding restoration implementation across different social, ecological, and political contexts. On the other hand, clusters of restoration hotspots emerged in a few global regions, notably within conservation hotspots (24). However, the KBAs showed a different pattern, with low mean ROS and only a minor proportion of their restorable area classified as restoration hotspots.
These contrasting results emerge from the different criteria used to designate conservation hotspots and KBAs. Although both classification types consider the levels and uniqueness of biodiversity within a region, the conservation hotspot approach consider only areas with very low habitat cover [<30% primary vegetation cover; (24)], whereas sites qualify as KBAs if they meet one or more thresholds for their significance for the global persistence of biodiversity (e.g., because they support threatened or geographically restricted species or ecosystems). As degraded habitats usually support lower abundance of these species and smaller extent of these ecosystems, they are less likely to qualify as KBAs (28). Consequently, many KBAs did not include restorable areas, as they presented >90% forest cover and were not even included in our analysis. In the KBAs that were included, areas had low ROS because our methodological approach indicated greater restoration benefits in areas with more severe degradation. On the other hand, our approach more closely matches that of conservation hotspots, explaining the remarkable congruence between conservation and restoration hotspots. Although biodiversity conservation hotspots would be, by definition, restoration hotspots when considering restoration benefits to prevent species loss, our approach considered six other independent layers in the assessment of restoration opportunities, so this congruence is an interesting demonstration that restoration in biodiversity conservation hotspots can also results in other associated co-benefits. Our results emphasize that approaches to minimizing extinctions of endemic species in these conservation hotspots will be strengthened by integrating well-planned restoration interventions within conservation programs (29).
Further analysis of the benefits and feasibility layers can be performed to evaluate the distribution of restoration hotspots based on differential weightings of layers to emphasize particular benefits or reduce costs of restoration interventions, depending on particular restoration objectives and stakeholder needs. The restoration hotspots approach can be adapted to different contexts, however, depending on project objectives and conservation priorities.
The restoration hotspots approach could also be used in combination with additional criteria at regional and national scales to guide the implementation of ecological corridors to mitigate effects of climate change and avoid biodiversity loss in the tropics (30), as well for avoiding afforestation in nonforest native ecosystems (23). Establishing restoration “hubs” in the emerging clusters of restoration hotspots may allow for a better optimization of restoration infrastructure and restoration supply chains (e.g., nursery facilities, capacity building and training, and development of markets for restoration products), fixed costs reduction, more efficient logistics, and the promotion of effective governance (31, 32).
We anticipate that our findings can be readily used to identify more cost-effective landscapes for implementation of the ambitious forest and landscape restoration commitments in the global tropics. For instance, the Bonn Challenge and the New York Declaration on Forests have accumulated 57 commitments including national and subnational commitments to restore 170 Mha by 2030 (15), with over 80% of these commitments in tropical developing countries. The methodological approach presented here could also contribute to the implementation of the Aichi Target 15 of the Convention on Biological Diversity, which aims to restore 15% of all degraded terrestrial ecosystems by 2020. A comprehensive analysis on the progress toward the achievement of Aichi targets was already made (33) but did not assess Target 15 due to the lack of appropriate indicators. This caveat highlights the challenge for planning, implementing, and monitoring Aichi Target 15 and reinforces the potential contribution of this work toward this goal. Moreover, our work goes beyond the assessment of this area-based target (34) to include other important variables to assess the potential contribution of restoration to biodiversity conservation. Our analysis could contribute to the post-2020 biodiversity framework, which is under negotiation through the Convention on Biological Diversity to replace the current 2011–2020 Strategic Plan.
Viewing restoration as a means to achieve certain goals and not as an end in itself, restoration commitments should ideally include the potential level of benefits that restoration can provide as criteria for identifying target areas. By doing so, the limited time and resources available to invest in restoration activities can be optimized, and high levels of benefits could be provided even in cases where restoration is implemented in only a portion of the area committed (35). Thus, our findings could be used to optimize the implementation of restoration efforts in the context of the Aichi Target 15, making better use of the limited time left before its expiration in 2020, and to guide the post-2020 restoration plans of the Convention on Biological Diversity.
The implementation of forest and landscape restoration commitments and targets also relies on many other socioenvironmental factors that were not included in this study, such as land tenure security, local disturbance factors, or legal instruments (5, 36), which demand attention when planning restoration at the local level. The benefits of restoration could also be improved if complementary criteria are included in the analysis, such as the use of KBAs to identify areas where restoration could be more relevant to support the persistence of unique biodiversity groups (37). Recognizing the rights and livelihoods of local peoples is critically important when implementing restoration projects (38, 39). Other types of land-use changes geared toward either protection (i.e., creating protected areas) or production (i.e., industrialized agriculture or forestry plantations) have often failed to recognize traditional land rights of local people, resulting in displacement and loss of livelihoods in local populations (40, 41). This risk is substantial for restoration interventions in highly populated areas.
Although forest and landscape restoration is viewed as a win-win solution for multiple environmental and socioeconomic benefits (18), our study highlights spatial matches and mismatches between benefits and feasibility of restoration across global tropics, pointing to potential trade-offs and synergies. For instance, we found a positive, but weak, association between locations of greater potential biodiversity benefit and locations with greater potential for delivering climate change mitigation, as previously found for tropical forest remnants (42). Our results showed a positive and robust spatial congruence between locations of greater importance for biodiversity conservation and for climate change adaptation, as well as between the locations of these two benefits and those of greater importance for water security. As water security has been one of the major benefits targeted in restoration projects (43, 44), decision makers may take advantage of restoration projects implemented to achieve watershed services to obtain these important associated co-benefits. In contrast, the most valuable landscapes for promoting biodiversity conservation and water security through restoration are associated with unfavorable economic conditions for implementing restoration projects, as a consequence of higher land opportunity costs and greater landscape variation in forest restoration success. These trade-offs may be a direct consequence of low remnant forest cover and the high degree of land degradation in highly populated regions submitted to intensive land uses, where watersheds are more degraded (45) and native species are threatened by anthropogenic impacts (46).
Although these challenges may be particularly relevant when implementing restoration at the local level, the global forest and landscape restoration movement is advancing rapidly and has been gradually incorporated as part of the solutions for promoting the United Nations Sustainable Development Goals. Analyses like ours, which go beyond the simplistic definition of a total area to be restored and explore which kind of benefits can be obtained and limitations to be faced by restoring landscapes in different locations, provide help to achieve existing targets. Newly established restoration targets could be accompanied with a set of potential priority landscapes right from the start. The time has come to go beyond setting targets based on how much land to restore, and advance toward prioritizing where and how to restore, decisions with utmost importance for maximizing benefits and minimizing implementation barriers. Although it may not be feasible to restore the large areas committed by many national and subnational pledges, more realistic targets could focus on restoring these priority areas within moist forest biomes and within other biomes. Concentrating investments of time, money, and effort in areas with higher potential return of benefits and feasibility of restoration would maximize the potential of restoration to repair anthropogenic impacts and offer a better future for all.
METHODS
Study area and restorable areas
We defined our study area (fig. S5A) within the distribution region of tropical moist broadleaf forests [fig. S6B; (22)], including only areas with altitude below 1000 m above sea level (asl; fig. S5C) and with available carbon density maps [figure 6D of (19)]. We excluded higher elevation areas because our approach to modeling the aboveground biomass (hereafter AGB) stocks (47)—a step in the assessment of restoration benefits on climate change mitigation—relied on the distribution of the secondary forest plots used to develop the model (42), and these plots were limited to below 1000 m asl. The equation used to estimate AGB stocks based on lowland forests cannot be reliably used for areas above 1000 m asl (47), so we excluded these areas from our analysis.
Within the study area, we identified restorable landscapes (fig. S5E). We excluded (i.e., we masked) areas that were not considered to be targets for forest and landscape restoration (i) with >90% tree canopy cover (48), using the updated version of the global tree cover map from 2016. These areas were excluded to avoid including areas that are essentially covered by continuous forests but did not reach 90 to 100% tree cover potentially due to natural conditions (e.g., tree fall gaps, small patches of nonforested ecosystems, and rock outcrops), as well as to focus our analysis on landscapes with a reasonable area to be restored to justify human interventions; (ii) urban areas and water bodies; and (iii) wetlands (49, 50). Data on urban areas, water bodies, and wetlands were obtained from the ESA Global Land Cover Dataset for 2015 (51). Nonforest ecosystems other than wetlands could not be excluded because of the lack of global maps presenting their original cover at a finer scale than (22), which is an unresolved caveat of our definition of restorable moist forest landscapes. We highlight that restrictions of the methodological approach used to define the study area and restorable landscapes excluded important areas for tropical rainforest restoration, as the Tropical Andes conservation hotspot and the Queensland tropical rainforests of Australia. We then assigned a ROS (i.e., normalized potential to maximize benefits and feasibility of restoration) to each of these restorable landscapes (i.e., the portion of the study area not covered by continuous forests, urban areas, water bodies, and wetlands) based on benefits and feasibility of restoration, as described below.
Restoration benefits
Overall, we considered that the higher the extent of degradation (deviation from continuous forest areas) in a given landscape, the higher is the anticipated benefit of restoration (i.e., ROS will be higher). Restoration benefits were defined within restorable landscapes according to four independent benefits, all equally weighted in the analysis: biodiversity conservation, climate change mitigation, climate change adaptation, and water security, as described below.
Biodiversity conservation
The restoration of tropical rainforest landscapes can critically contribute to conserve biodiversity because tropical forests harbor half of all Earth’s species (2) and because this activity may help to prevent species extinctions by increasing habitat cover and landscape connectivity (10, 52). We specifically focused our analysis on small-ranged (i.e., with a geographic range size smaller than the global median) and threatened (i.e., included in the International Union for the Conservation of Nature’s Red List) mammal, bird, and amphibian species, because restoration could potentially have a greater benefit for their conservation compared to generalist, widespread species and because there were no consolidated global maps for other biodiversity groups. We used the global database downloaded from biodiversitymapping.org for the analyses, which is underlying data from the International Union for the Conservation of Nature, BirdLife International, and NatureServe, comprising distribution maps for each species (53, 54). Mammals, birds, and amphibians, as well as small-ranged and threatened species, were equally considered in the analysis. To do so, first, we summed the number of small-ranged or threatened species of each biodiversity group separately and generated three individual maps with the total number of small-ranged and threatened species of mammals, birds, and amphibians. Second, each map was rescaled from 0 to 1 (1 being the highest number of species). Last, the three maps were intercepted and rescaled from 0 to 1 (fig. S6A). Then, the landscapes with higher normalized number of small-ranged and threatened species of mammals, birds, and amphibians received a higher restoration score.
Climate change mitigation
Restored tropical forests can critically contribute to reduction of CO2 in the atmosphere and, thus, mitigate global climate change (55, 56). To include this restoration benefit in the analyses, we first built a map of the total potential AGB based on the interpolation of existing old-growth forests to restorable areas, using climate and soil variables, and an existing equation developed for the Neotropics based on over 1500 field plots (47). Specifically, we mapped the 1-ha old-growth plot locations described in (42), which are widely distributed across our study area, but exclude many tropical rainforest regions, as Central America, the Caribbean, Atlantic Forest, Madagascar, and mainland Southeast Asia (only one plot), so we recognize that there is high likelihood that the results in these regions will not be accurate. Since equation selected to model old-growth carbon potential was derived from (47), which does include a large network of old-growth and secondary forest areas across the Neotropics, the accuracy of the modeled AGB potential was expected to be higher in these regions. We then buffered the old-growth plots by a radius of 500 m to provide a stable area, large enough to encompass any spatial resolution sensor that we decided to include in the study, which was not completely determined at the time of this selection process. For example, if we used a 500-m MODIS pixel, then it would still be certain to be within only intact areas of old-growth forests versus being a mixed land use pixel. We then manually reviewed each plot using Google Earth Pro high-resolution recent (2012+) year satellite imagery and removed those plots having degraded or converted forest visible within the buffer. A multivariate linear regression equation was developed to predict maximum total AGB that included annual average, minimum, and maximum temperature, long-term climatic water deficit (CWD), soil cation exchange capacity (CEC), and elevation [Digital Elevation Model (DEM)], based on (3). We used a forward stepwise regression to identify significant variables (all with P < 0.0001) from a larger pool of potential bioclimatic predictor variables, with the final equation (Adj. R2 = 0.55, N = 259) provided below
Maximum potential AGB = −358 − 68 × avg. temp. + 44 × min. temp. + 48 × max. temp. + 0.15 × CWD − 2.9 × CEC + 0.21 × DEM.
We then subtracted from this maximum potential AGB the present day values (e.g., which includes intact and deforested or degraded areas), as provided by Baccini et al. (19), to calculate the potential AGB that could be sequestered by restoring a forest to local reference values within each pixel. We used the asymptote of the curve describing the temporal accumulation of AGB by each forest area to calculate its potential AGB stock. The estimated time required to achieve this potential is around 40 years (21) but varies depending on the bioclimatic characteristics of the site. AGB values of restorable areas were rescaled from 0 to 1 (1 being the highest AGB potential), resulting in a final map of the maximum potential AGB uptake by restored forests (fig. S6B). Then, the landscapes with higher normalized potential for new AGB sequestration in restored forests received a higher ROS for climate change mitigation.
Climate change adaptation
The restoration of tropical rainforest landscapes can contribute to climate change adaptation because (i) forest restoration increases landscape connectivity and may support species migration to more favorable conditions within human-modified landscapes (57); (ii) forest restoration can protect soils against extreme rainfall events (58); (iii) new forests can help reduce heat waves and movement of dust storms in populated regions (59); and (iv) restored forests can help protect watersheds and increase their resilience to extreme events (44). Briefly, forest restoration may help people and biodiversity to face climate change where its consequences are expected to be most severe. We then assumed that restoration benefits as adaptive measure to climate change would be maximized where climate will change faster. We used the database developed by Loarie et al. (60) in the analyses. First, we rescaled climate change speed (km year−1) from 0 to 1 (1 being the highest climate speed) to generate a single map (fig. S6C). Then, the landscapes with higher normalized velocity of climate change at the global scale received a higher restoration score for climate change adaptation.
Water security
The restoration of tropical rainforest landscapes can contribute to mitigate water supply risks as consequence of the (i) protection of water courses from siltation, (ii) protection of water channels from fluvial erosion, (iii) enhancement of rainfall infiltration into soils and then recharge of water table and aquifers, (iv) regulation of water supply distribution across the year, and (v) enhancement of moisture recycling (61). We are aware that forest restoration can reduce water yield in catchments (44) and eventually increase water security risk. However, we considered that the overall benefits of restorative interventions in degraded and deforested landscapes would compensate for those potential negative and short-term outcomes of restoration. We then assumed that restoration benefit would be maximized where incident water security risk is higher and based our analyses in the map of incident water security risks (map A of Fig. 1) developed by Vorosmarty et al. (45). This map was already scaled from 0 to 1, so it was directly integrated into our analysis (fig. S6D). Then, the landscapes with higher normalized incident water security risk received a higher restoration score for water security.
Restoration feasibility
Restoration feasibility consists of enabling socioenvironmental factors of forest and landscape restoration in tropical rainforest landscapes. We considered that restoration has higher likelihood of effective implementation and long-term sustainability in regions where (i) land opportunity costs are lower, as the chances of land use conversion to forests are higher when farmers get lower economic benefits from their lands; (ii) the landscape variation in forest restoration is reduced, and biodiversity has a higher and more predictable chance to recolonize landscapes undergoing restoration, which would lower the risks for investors to financially support restoration initiatives; and (iii) the chances of restoration persistence over time are higher, which estimates the chances of a forest undergoing restoration to be reconverted to agricultural land uses. The development of the maps associated with each of these variables was described below.
Land opportunity costs
Land opportunity cost is an important component of forest and landscape restoration feasibility because restoration has higher chances to occur in lands yielding reduced economic gains to farmers. Landscapes with lower land opportunity costs may offer less suitable conditions for intensive, mechanized agriculture, so the chances for increasing tree cover in productive landscapes tend to be higher, as well the chances of abandonment of marginal agricultural lands for further regeneration of second-growth native forests. We used the map on economic return from agricultural lands produced by Naidoo and Iwamura (62) as a proxy of land opportunity costs (i.e., the higher is the economic return, the higher is the land opportunity cost). We normalized the monetary values of economic return from agricultural lands from 0 to 1 (1 being the lowest value). Then, since extreme high values were resulting in a generalized homogenization of land opportunity cost scores globally, we “clamped” the extreme 5% high and 5% low values (5%), so all pixels with top 5% of values received a score of 0, and all pixels with the lowest values received a score of 1. Then, the landscapes with higher normalized economic return from agricultural lands received a lower restoration score.
Landscape variation in forest restoration success
Restoration feasibility is negatively affected when biodiversity recolonization of restoration sites is more variable and, then, unpredictable because (i) restoration becomes more risky, so investors may not be attracted to financially support restoration programs; (ii) restoration implementation costs tend to be higher, since expensive tree plantings may be required in landscapes with higher landscape variation in forest restoration success; and (iii) biodiversity recovery may be compromised and so too the potential of restored forests to deliver many of the benefits expected from restoration, such as pollination and pest control in agriculture. To estimate the potential variation of biodiversity recovery in restorable landscapes, we first performed a meta-analysis based on 135 study forest landscapes distributed globally and measured the response ratio of biodiversity recovery (i.e., species richness, abundance, diversity, and/or similarity) in restored forests versus reference forests (i.e., old-growth/less-disturbed forests). Then, we calculated the response ratio variance of biodiversity recovery in restored forests, using as a benchmark, the “full restoration success” (i.e., the value of the diversity metric in reference forests within the same study landscape). For more details and equations, see (63). The evidence ratio of the top-ranked model [Akaike weights (wi) = 0.4)] was 400 times higher than the null model [ΔAICc (Δ Akaike information criterion, with correction for small sizes) – 11.47; wi = 0.001]. We found a negative exponential relationship between the amount of forest cover within landscapes with a buffer size of 5-km radius and the response ratio variance (63). Then, we built a global map of forest cover in 2016 for 5-km radio landscapes using data from (48) (but updated to 2016). The forest cover map was built by using tree canopy cover for 2000 excluding global forest loss up to 2016 and resampled at 300-m pixel size. Last, the equation describing the response ratio variance in relation to forest cover in the landscape (used here as a surrogate of the landscape variation in forest restoration success) was applied across the restorable landscapes identified in this work, which had their forest cover values determined as described above, to assign a value ranging from 0 to 1 to each landscape, 1 being the lowest value (lower landscape variation in forest restoration success; fig. S8B). Then, the landscapes with higher normalized landscape variation in forest restoration success received a lower restoration score.
Persistence chances of restored forests
The persistence of regenerating tropical forests over time has been considered one of the most critical factors for successful large-scale restoration, as most young forests may persist for only few years (64). Low rates of forest persistence may reflect that the socioeconomic drivers of land-use change in the landscape are still directed toward the conversion of forest ecosystems and tree cover to agricultural lands and that the enabling conditions for a forest transition are not yet realized. We estimated the persistence chances of restored forests using the relative rate of recent tree cover loss as surrogate. To do so, we summed forest cover loss from 2001 to 2015 and divided it by forest cover in 2000, using data from (48). We applied a threshold of 20% tree canopy cover for year 2000 data to produce a binary map of forest (1)/nonforest (0) on its original spatial resolution (fig. S8C). Then, the landscapes with higher normalized chances for restored forests to persist over time received a higher restoration score.
Harmonization and geospatial analyses
The spatial resolution of all input layers were 0.5 km × 0.5 km or finer, except for biodiversity conservation (~10 km × 10 km) and water security (~10 km × 10 km). We defined the spatial resolution of our work at 1 km × 1 km. The layer with finer spatial resolution was aggregated by mean, and those with coarser spatial resolution were resampled using bilinear interpolation to the target resolution of 1 km × 1 km. We assessed the robustness of the ROS to the changes in the input layers. We performed the robustness analysis by randomly changing the cell values within 5, 10, and 20% and then used a Monte Carlo procedure with 500 iterations for each of the input layers. We then calculated the SD of the score of each combination for each pixel. The maximum deviations from mean were 0.0087, 0.0164, and 0.0299 when inputs changed 5, 10, and 20%, respectively.
We performed all the geospatial analyses in R version 3.4.4 (65) using raster (66), rdgal (67), and ncdf4 (68). The visualization and cartography of the maps were conducted in ArcMap (69).
Spatial congruence among different benefits and feasibility of restoration
We used Pearson correlations to investigate the spatial association among all benefits and feasibility of restoration. We did not control for spatial autocorrelation because we assumed that it is an inherent component of the definition of restoration hotspots, so we expected that the association between the variables we analyzed would inevitably reflect the regional context of restoration implementation.
Supplementary Material
Acknowledgments
Funding: P.H.S.B. thanks the National Council for Scientific and Technological Development of Brazil (CNPq, grant no. 304817/2015-5). P.H.S.B. and R.L.C. received funding from the Coordination for the Improvement of Higher Education Personnel of Brazil (CAPES, grant no. 88881.064976/2014-01). S.G. acknowledges support from NASA grants NNX17AG51G and NNL15AA03C. This work is a product of the PARTNERS Research Coordination Network grant no. DEB1313788 from the U.S. NSF Coupled Natural and Human Systems Program. Author contributions: P.H.S.B. conceived the idea, designed the study, and led the writing. R.L.C. co-led the writing. P.H.S.B. and A.N. decided on data analysis and integration. A.N., E.B., R.C., F.S.M.B., and A.M.A.Z. performed geospatial analysis. A.B. and S.G. provided data on biomass stocks. E.B. and A.M.A.Z. developed the biomass equation. R.C. and F.S.B. provided the data on forest restoration success meta-analysis and developed an equation for the global tropics. All authors discussed the idea and study design and revised the manuscript. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors. All data and materials are available at https://doi.org/10.5281/zenodo.3233495.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/7/eaav3223/DC1
Fig. S1. Distribution of the total restoration area in relation to the ROS in tropical rainforest landscapes.
Fig. S2. ROS of tropical rainforest landscapes of countries, with data renormalized for each country.
Fig. S3. ROS of tropical rainforest landscapes of biogeographical realms, with data renormalized for each realm (A, Neotropics; B, Afrotropics; C, Indo-Malaysia; D, Australasia).
Fig. S4. ROS of tropical rainforest landscapes of global hotspots for biodiversity conservation, with data renormalized for each hotspot.
Fig. S5. Identification of restorable areas.
Fig. S6. Restoration benefits.
Fig. S7. Restoration feasibility factors.
Table S1. Country’s ROS, restorable area, and restorable area with a restoration score of >0.6 of tropical rainforest landscapes and their national pledges to the Bonn Challenge.
Table S2. Mean ROS, total area, restorable area, and restorable area with a restoration score of >0.6 of ecoregions within tropical rainforest landscapes.
Table S3. Mean ROS, study area, restorable area, and restorable area with a restoration score of >0.6 of KBAs within tropical rainforest landscapes.
Table S4. Mean ROS, total area, restorable area, and restorable area with a restoration score of >0.6 of tropical rainforest landscapes among hotspots for biodiversity conservation.
REFERENCES AND NOTES
- 1.Malhi Y., Gardner T. A., Goldsmith G. R., Silman M. R., Zelazowski P., Tropical forests in the Anthropocene. Annu. Rev. Env. Resour. 39, 125–159 (2014). [Google Scholar]
- 2.Lewis S. L., Edwards D. P., Galbraith D., Increasing human dominance of tropical forests. Science 349, 827–832 (2015). [DOI] [PubMed] [Google Scholar]
- 3.Baccini A., Walker W., Carvalho L., Farina M., Sulla-Menashe D., Houghton R. A., Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 358, 230–234 (2017). [DOI] [PubMed] [Google Scholar]
- 4.Watson J. E. M., Evans T., Venter O., Williams B., Tulloch A., Stewart C., Thompson I., Ray J. C., Murray K., Salazar A., McAlpine C., Potapov P., Walston J., Robinson J. G., Painter M., Wilkie D., Filardi C., Laurance W. F., Houghton R. A., Maxwell S., Grantham H., Samper C., Wang S., Laestadius L., Runting R. K., Silva-Chávez G. A., Ervin J., Lindenmayer D., The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2, 599–610 (2018). [DOI] [PubMed] [Google Scholar]
- 5.Barlow J., Lennox G. D., Ferreira J., Berenguer E., Lees A. C., Nally R. M., Thomson J. R., Ferraz S. F., Louzada J., Oliveira V. H. F., Parry L., R. Solar R., Vieira I. C. G., Aragão L. E., Begotti R. A., Braga R. F., Cardoso T. M., de Oliveira R. C. Jr., Souza Jr C. M., Moura N. G., Nunes S. S., Siqueira J. V., Pardini R., Silveira J. M., Vaz-de-Mello F. Z., Veiga R. C. S., Venturieri A., Gardner T. A., Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 535, 144–147 (2016). [DOI] [PubMed] [Google Scholar]
- 6.Díaz S., Pascual U., Stenseke M., Martín-López B., Watson R. T., Molnár Z., Hill R., Chan K. M. A., Baste I. A., Brauman K. A., Polasky S., Church A., Lonsdale M., Larigauderie A., Leadley P. W., van Oudenhoven A. P. E., van der Plaat F., Schröter M., Lavorel S., Aumeeruddy-Thomas Y., Bukvareva E., Davies K., Demissew S., Erpul G., Failler P., Guerra C. A., Hewitt C. L., Keune H., Lindley S., Shirayama Y., Assessing nature’s contributions to people. Science 359, 270–272 (2018). [DOI] [PubMed] [Google Scholar]
- 7.Myers J. A., Chase J. M., Jiménez I., Jørgensen P. M., Araujo-Murakami A., Paniagua-Zambrana N., Seidel R., Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly. Ecol. Lett. 16, 151–157 (2013). [DOI] [PubMed] [Google Scholar]
- 8.Mace G. M., Whose conservation? Science 345, 1558–1560 (2014). [DOI] [PubMed] [Google Scholar]
- 9.Possingham H. P., Bode M., Klein C. J., Optimal conservation outcomes require both restoration and protection. PLOS Biol. 13, e1002052 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Newmark W. D., Jenkins C. N., Pimm S. L., McNeally P. B., Halley J. M., Targeted habitat restoration can reduce extinction rates in fragmented forests. Proc. Natl. Acad. Sci. U.S.A. 114, 9635–9640 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Houghton R. A., Byers B., Nassikas A. A., A role for tropical forests in stabilizing atmospheric CO2. Nat. Clim. Change 5, 1022–1023 (2015). [Google Scholar]
- 12.Sunderlin W. D., Angelsen A., Belcher B., Burgers P., Nasi R., Santoso L., Wunder S., Livelihoods, forests, and conservation in developing countries: An overview. World Dev. 33, 1383–1402 (2005). [Google Scholar]
- 13.Brancalion P. H. S., Chazdon R. L., Beyond hectares: Four principles to guide reforestation in the context of tropical forest and landscape restoration. Restor. Ecol. 25, 491–496 (2017). [Google Scholar]
- 14.Holl K. D., Restoring tropical forests from the bottom up. Science 355, 455–456 (2017). [DOI] [PubMed] [Google Scholar]
- 15.Chazdon R. L., Brancalion P. H. S., Lamb D., Laestadius L., Calmon M., Kumar C., A policy-driven knowledge agenda for global forest and landscape restoration. Conserv. Lett. 10, 125–132 (2017). [Google Scholar]
- 16.WorldBank (World Bank, 2016), vol. 2018.
- 17.Godfray H. C. J., Beddington J. R., Crute I. R., Haddad L., Lawrence D., Muir J. F., Pretty J., Robinson S., Thomas S. M., Toulmin C., Food Security: The challenge of feeding 9 billion people. Science 327, 812–818 (2010). [DOI] [PubMed] [Google Scholar]
- 18.Sabogal C., Besacier C., McGuire D., Forest and landscape restoration: Concepts, approaches and challenges for implementation. Unasylva 66, 3–10 (2015). [Google Scholar]
- 19.Baccini A., Goetz S. J., Walker W. S., Laporte N. T., Sun M., Sulla-Menashe D., Hackler J., Beck P. S. A., Dubayah R., Friedl M. A., Samanta S., Houghton R. A., Estimated carbon dioxide emissions from tropical deforestation improved by carbon-density maps. Nat. Clim. Change 2, 182–185 (2012). [Google Scholar]
- 20.NewYorkDeclarationonForests (2018).
- 21.Poorter L., Bongers F., Aide T. M., Almeyda Zambrano A. M., Balvanera P., Becknell J. M., Boukili V., Brancalion P. H. S., Broadbent E. N., Chazdon R. L., Craven D., de Almeida-Cortez J. S., Cabral G. A. L., de Jong B. H. J., Denslow J. S., Dent D. H., DeWalt S. J., Dupuy J. M., Durán S. M., Espírito-Santo M. M., Fandino M. C., César R. G., Hall J. S., Hernandez-Stefanoni J. L., Jakovac C. C., Junqueira A. B., Kennard D., Letcher S. G., Licona J. C., Lohbeck M., Marín-Spiotta E., Martínez-Ramos M., Massoca P., Meave J. A., Mesquita R., Mora F., Muñoz R., Muscarella R., Nunes Y. R. F., Ochoa-Gaona S., de Oliveira A. A., Orihuela-Belmonte E., Peña-Claros M., Pérez-García E. A., Piotto D., Powers J. S., Rodríguez-Velázquez J., Romero-Pérez I. E., Ruíz J., Saldarriaga J. G., Sanchez-Azofeifa A., Schwartz N. B., Steininger M. K., Swenson N. G., Toledo M., Uriarte M., van Breugel M., van der Wal H., Veloso M. D. M., Vester H. F. M., Vicentini A., Vieira I. C. G., Bentos T. V., Williamson G. B., Rozendaal D. M. A., Biomass resilience of Neotropical secondary forests. Nature 530, 211–214 (2016). [DOI] [PubMed] [Google Scholar]
- 22.Olson D. M., Dinerstein E., Wikramanayake E. D., Burgess N. D., Powell G. V. N., Underwood E. C., D’amico J. A., Itoua I., Strand H. E., Morrison J. C., Loucks C. J., Allnutt T. F., Ricketts T. H., Kura Y., Lamoreux J. F., Wettengel W. W., Hedao P., Kassem K. R., Terrestrial ecoregions of the world: A new map of life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51, 933–938 (2001). [Google Scholar]
- 23.Veldman J. W., Overbeck G. E., Negreiros D., Mahy G., Le Stradic S., Fernandes G. W., Durigan G., Buisson E., Putz F. E., Bond W. J., Where tree planting and forest expansion are bad for biodiversity and ecosystem services. Bioscience 65, 1011–1018 (2015). [Google Scholar]
- 24.Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J., Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000). [DOI] [PubMed] [Google Scholar]
- 25.Roberts C. M., McClean C. J., Veron J. E., Hawkins J. P., Allen G. R., McAllister D. E., Mittermeier C. G., Schueler F. W., Spalding M., Wells F., Vynne C., Werner T. B., Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295, 1280–1284 (2002). [DOI] [PubMed] [Google Scholar]
- 26.Tordoff A., et al. , Key Biodiversity Areas as Site Conservation Targets. Bioscience 54, 1110–1118 (2004). [Google Scholar]
- 27.IUCN/WRI, “A guide to the Restoration Opportunities Assessment Methodology (ROAM): Assessing forest landscape restoration opportunities at the national or sub-national level” (2014).
- 28.K. S. A. A. Committee, Guidelines for using a Global Standard for the Identification of Key Biodiversity Areas (International Union for the Conservation of Nature, 2019), pp. 148.
- 29.Brooks T. M., Mittermeier R. A., Mittermeier C. G., Da Fonseca G. A. B., Rylands A. B., Konstant W. R., Flick P., Pilgrim J., Oldfield S., Magin G., Hilton-Taylor C., Habitat loss and extinction in the hotspots of biodiversity. Conserv. Biol. 16, 909–923 (2002). [Google Scholar]
- 30.Jantz P., Goetz S., Laporte N., Carbon stock corridors to mitigate climate change and promote biodiversity in the tropics. Nat. Clim. Change 4, 138–142 (2014). [Google Scholar]
- 31.Brancalion P. H. S., Lamb D., Ceccon E., Boucher D., Herbohn J., Strassburg B., Edwards D. P., Using markets to leverage investment in forest and landscape restoration in the tropics. Forest Policy Econ. 85, 103–113 (2017). [Google Scholar]
- 32.Menz M. H. M., Dixon K. W., Hobbs R. J., Hurdles and opportunities for landscape-scale restoration. Science 339, 526–527 (2013). [DOI] [PubMed] [Google Scholar]
- 33.Tittensor D. P., Walpole M., Hill S. L. L., Boyce D. G., Britten G. L., Burgess N. D., Butchart S. H. M., Leadley P. W., Regan E. C., Alkemade R., Baumung R., Bellard C., Bouwman L., Bowles-Newark N. J., Chenery A. M., Cheung W. W. L., Christensen V., Cooper H. D., Crowther A. R., Dixon M. J. R., Galli A., Gaveau V., Gregory R. D., Gutierrez N. L., Hirsch T. L., Hoft R., Januchowski-Hartley S. R., Karmann M., Krug C. B., Leverington F. J., Loh J., Lojenga R. K., Malsch K., Marques A., Morgan D. H. W., Mumby P. J., Newbold T., Noonan-Mooney K., Pagad S. N., Parks B. C., Pereira H. M., Robertson T., Rondinini C., Santini L., Scharlemann J. P. W., Schindler S., Sumaila U. R., Teh L. S. L., van Kolck J., Visconti P., Ye Y., A mid-term analysis of progress toward international biodiversity targets. Science 346, 241–244 (2014). [DOI] [PubMed] [Google Scholar]
- 34.Barnes M., Protect biodiversity, not just area. Nature 526, 195 (2015). [DOI] [PubMed] [Google Scholar]
- 35.Strassburg B. B. N., Beyer H. L., Crouzeilles R., Iribarrem A., Barros F., de Siqueira M. F., Sánchez-Tapia A., Balmford A., Sansevero J. B. B., Brancalion P. H. S., Broadbent E. N., Chazdon R. L., Filho A. O., Gardner T. A., Gordon A., Latawiec A., Loyola R., Metzger J. P., Mills M., Possingham H. P., Rodrigues R. R., Scaramuzza C. A. M., Scarano F. R., Tambosi L., Uriarte M., Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nat. Ecol. Evol. 3, 62–70 (2019). [DOI] [PubMed] [Google Scholar]
- 36.Latawiec A. E., Strassburg B. B. N., Brancalion P. H. S., Rodrigues R. R., Gardner T., Creating space for large-scale restoration in tropical agricultural landscapes. Front. Ecol. Environ. 13, 211–218 (2015). [Google Scholar]
- 37.C. R. Beatty, N. A. Cox, M. E. Kuzee, Biodiversity Guidelines for Forest Landscape Restoration Opportunities Assessments (International Union for the Conservation of Nature, 2018).
- 38.Lazos-Chavero E., Zinda J., Bennett-Curry A., Balvanera P., Bloomfield G., Lindell C., Negra C., Stakeholders and tropical reforestation: Challenges, trade-offs, and strategies in dynamic environments. Biotropica 48, 900–914 (2016). [Google Scholar]
- 39.Erbaugh J. T., Oldekop J. A., Forest landscape restoration for livelihoods and well-being. Curr. Opin. Environ. Sustain. 32, 76–83 (2018). [Google Scholar]
- 40.Fairhead J., Leach M., Scoones I., Green Grabbing: A new appropriation of nature? J. Peasant Stud. 39, 237–261 (2012). [Google Scholar]
- 41.Rulli M. C., Saviori A., D'Odorico P., Global land and water grabbing. Proc. Natl. Acad. Sci. U.S.A. 110, 892–897 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sullivan M. J. P., Talbot J., Lewis S. L., Phillips O. L., Qie L., Begne S. K., Chave J., Cuni-Sanchez A., Hubau W., Lopez-Gonzalez G., Miles L., Monteagudo-Mendoza A., Sonké B., Sunderland T., ter Steege H., White L. J. T., Affum-Baffoe K., Aiba S. I., de Almeida E. C., de Oliveira E. A., Alvarez-Loayza P., Dávila E. Á., Andrade A., Aragão L. E. O. C., Ashton P., Aymard C. G. A., Baker T. R., Balinga M., Banin L. F., Baraloto C., Bastin J. F., Berry N., Bogaert J., Bonal D., Bongers F., Brienen R., Camargo J. L. C., Cerón C., Moscoso V. C., Chezeaux E., Clark C. J., Pacheco Á. C., Comiskey J. A., Valverde F. C., Coronado E. N. H., Dargie G., Davies S. J., de Canniere C., Djuikouo K. M. N., Doucet J. L., Erwin T. L., Espejo J. S., Ewango C. E. N., Fauset S., Feldpausch T. R., Herrera R., Gilpin M., Gloor E., Hall J. S., Harris D. J., Hart T. B., Kartawinata K., Kho L. K., Kitayama K., Laurance S. G. W., Laurance W. F., Leal M. E., Lovejoy T., Lovett J. C., Lukasu F. M., Makana J. R., Malhi Y., Maracahipes L., Marimon B. S., Junior B. H. M., Marshall A. R., Morandi P. S., Mukendi J. T., Mukinzi J., Nilus R., Vargas P. N., Camacho N. C. P., Pardo G., Peña-Claros M., Pétronelli P., Pickavance G. C., Poulsen A. D., Poulsen J. R., Primack R. B., Priyadi H., Quesada C. A., Reitsma J., Réjou-Méchain M., Restrepo Z., Rutishauser E., Salim K. A., Salomão R. P., Samsoedin I., Sheil D., Sierra R., Silveira M., Slik J. W. F., Steel L., Taedoumg H., Tan S., Terborgh J. W., Thomas S. C., Toledo M., Umunay P. M., Gamarra L. V., Vieira I. C. G., Vos V. A., Wang O., Willcock S., Zemagho L., Diversity and carbon storage across the tropical forest biome. Sci. Rep. 7, 39102 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Murcia C., Guariguata M. R., Andrade Á., Andrade G. I., Aronson J., Escobar E. M., Etter A., Moreno F. H., Ramírez W., Montes E., Challenges and prospects for scaling-up ecological restoration to meet international commitments: Colombia as a case Study. Conserv. Lett. 9, 213–220 (2016). [Google Scholar]
- 44.Filoso S., Bezerra M. O., Weiss K. C. B., Palmer M. A., Impacts of forest restoration on water yield: A systematic review. PLOS ONE 12, e0183210 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Vorosmarty C. J., McIntyre P. B., Gessner M. O., Dudgeon D., Prusevich A., Green P., Glidden S., Bunn S. E., Sullivan C. A., Liermann C. R., Davies P. M., Global threats to human water security and river biodiversity. Nature 467, 555–561 (2010). [DOI] [PubMed] [Google Scholar]
- 46.Moreno-Mateos D., Barbier E. B., Jones P. C., Jones H. P., Aronson J., López-López J. A., McCrackin M. L., Meli P., Montoya D., Rey Benayas J. M., Anthropogenic ecosystem disturbance and the recovery debt. Nat. Commun. 8, 14163 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chazdon R. L., Broadbent E. N., Rozendaal D. M. A., Bongers F., Zambrano A. M. A., Aide T. M., Balvanera P., Becknell J. M., Boukili V., Brancalion P. H. S., Craven D., Almeida-Cortez J. S., Cabral G. A. L., de Jong B., Denslow J. S., Dent D. H., DeWalt S. J., Dupuy J. M., Durán S. M., Espírito-Santo M. M., Fandino M. C., César R. G., Hall J. S., Hernández-Stefanoni J. L., Jakovac C. C., Junqueira A. B., Kennard D., Letcher S. G., Lohbeck M., Martínez-Ramos M., Massoca P., Meave J. A., Mesquita R., Mora F., Muñoz R., Muscarella R., Nunes Y. R. F., Ochoa-Gaona S., Orihuela-Belmonte E., Peña-Claros M., Pérez-García E. A., Piotto D., Powers J. S., Rodríguez-Velazquez J., Romero-Pérez I. E., Ruíz J., Saldarriaga J. G., Sanchez-Azofeifa A., Schwartz N. B., Steininger M. K., Swenson N. G., Uriarte M., van Breugel M., van der Wal H., Veloso M. D. M., Vester H., Vieira I. C. G., Bentos T. V., Williamson G. B., Poorter L., Carbon sequestration potential of second-growth forest regeneration in the Latin American tropics. Sci. Adv. 2, e1501639 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hansen M. C., Potapov P. V., Moore R., Hancher M., Turubanova S. A., Tyukavina A., Thau D., Stehman S. V., Goetz S. J., Loveland T. R., Kommareddy A., Egorov A., Chini L., Justice C. O., Townshend J. R. G., High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013). [DOI] [PubMed] [Google Scholar]
- 49.Prigent C., Papa F., Aires F., Rossow W. B., Matthews E., Global inundation dynamics inferred from multiple satellite observations, 1993–2000. J. Geophys. Res. Atmos. 112, 10.1029/2006JD007847 (2007). [Google Scholar]
- 50.Fluet-Chouinard E., Lehner B., Rebelo L.-M., Papa F., Hamilton S. K., Development of a global inundation map at high spatial resolution from topographic downscaling of coarse-scale remote sensing data. Remote Sens. Environ. 158, 348–361 (2015). [Google Scholar]
- 51.ESA. (2017), vol. 2017.
- 52.Banks-Leite C., Pardini R., Tambosi L. R., Pearse W. D., Bueno A. A., Bruscagin R. T., Condez T. H., Dixo M., Igari A. T., Martensen A. C., Metzger J. P., Using ecological thresholds to evaluate the costs and benefits of set-asides in a biodiversity hotspot. Science 345, 1041–1045 (2014). [DOI] [PubMed] [Google Scholar]
- 53.Jenkins C. N., Pimm S. L., Joppa L. N., Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. U.S.A. 110, E2602–E2610 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pimm S. L., Jenkins C. N., Abell R., Brooks T. M., Gittleman J. L., Joppa L. N., Raven P. H., Roberts C. M., Sexton J. O., The biodiversity of species and their rates of extinction, distribution, and protection. Science 344, 1246752 (2014). [DOI] [PubMed] [Google Scholar]
- 55.Griscom B. W., Adams J., Ellis P. W., Houghton R. A., Lomax G., Miteva D. A., Schlesinger W. H., Shoch D., Siikamäki J. V., Smith P., Woodbury P., Zganjar C., Blackman A., Campari J., Conant R. T., Delgado C., Elias P., Gopalakrishna T., Hamsik M. R., Herrero M., Kiesecker J., Landis E., Laestadius L., Leavitt S. M., Minnemeyer S., Polasky S., Potapov P., Putz F. E., Sanderman J., Silvius M., Wollenberg E., Fargione J., Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rogelj J., den Elzen M., Höhne N., Fransen T., Fekete H., Winkler H., Schaeffer R., Sha F., Riahi K., Meinshausen M., Paris agreement climate proposals need a boost to keep warming well below 2 °C. Nature 534, 631–639 (2016). [DOI] [PubMed] [Google Scholar]
- 57.Noss R. F., Beyond Kyoto: Forest management in a time of rapid climate change. Conserv. Biol. 15, 578–590 (2001). [Google Scholar]
- 58.Zheng H., Chen F., Ouyang Z., Tu N., Xu W., Wang X., Miao H., Li X., Tian Y., Impacts of reforestation approaches on runoff control in the hilly red soil region of Southern China. J. Hydrol. 356, 174–184 (2008). [Google Scholar]
- 59.Tan M., Li X., Does the Green Great Wall effectively decrease dust storm intensity in China? A study based on NOAA NDVI and weather station data. Land Use Policy 43, 42–47 (2015). [Google Scholar]
- 60.Loarie S. R., Duffy P. B., Hamilton H., Asner G. P., Field C. B., Ackerly D. D., The velocity of climate change. Nature 462, 1052–1055 (2009). [DOI] [PubMed] [Google Scholar]
- 61.Ellison D., Morris C. E., Locatelli B., Sheil D., Cohen J., Murdiyarso D., Gutierrez V., Noordwijk M., Creed I. F., Pokorny J., Gaveau D., Spracklen D. V., Tobella A. B., Ilstedt U., Teuling A. J., Gebrehiwot S. G., Sands D. C., Muys B., Verbist B., Springgay E., Sugandi Y., Sullivan C. A., Trees, forests and water: Cool insights for a hot world. Global Environ. Change 43, 51–61 (2017). [Google Scholar]
- 62.Naidoo R., Iwamura T., Global-scale mapping of economic benefits from agricultural lands: Implications for conservation priorities. Biol. Conserv. 140, 40–49 (2007). [Google Scholar]
- 63.Crouzeilles R., Curran M., Ferreira M. S., Lindenmayer D. B., Grelle C. E. V., Benayas J. M. R., A global meta-analysis on the ecological drivers of forest restoration success. Nat. Commun. 7, 11666 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Reid J. L., Wilson S. J., Bloomfield G. S., Cattau M. E., Fagan M. E., Holl K. D., Zahawi R. A., How long do restored ecosystems persist? Ann. Mo. Bot. Gard. 102, 258–265 (2017). [Google Scholar]
- 65.RCoreTeam (R Foundation for Statistical Computing, 2018).
- 66.R. J. Hijmans, Geographic Data Analysis and Modeling [R package raster version 2.5-8], (2017); available at http://cran.r-project.org/package=raster.
- 67.R. Bivand, T. Keitt, B. Rowlingson, rgdal: Bindings for the “Geospatial” Data Abstraction Library [R package version 1.4-3] (2018); available at https://CRAN.R-project.org/package=rgdal.
- 68.D. Pierce, ncdf4: Interface to Unidata netCDF (Version 4 or Earlier) Format Data. [R package version 1.15] (2017); available at https://cran.r-project.org/package=ncdf4.
- 69.ESRI (Environmental Systems Research Institute, 2017).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/7/eaav3223/DC1
Fig. S1. Distribution of the total restoration area in relation to the ROS in tropical rainforest landscapes.
Fig. S2. ROS of tropical rainforest landscapes of countries, with data renormalized for each country.
Fig. S3. ROS of tropical rainforest landscapes of biogeographical realms, with data renormalized for each realm (A, Neotropics; B, Afrotropics; C, Indo-Malaysia; D, Australasia).
Fig. S4. ROS of tropical rainforest landscapes of global hotspots for biodiversity conservation, with data renormalized for each hotspot.
Fig. S5. Identification of restorable areas.
Fig. S6. Restoration benefits.
Fig. S7. Restoration feasibility factors.
Table S1. Country’s ROS, restorable area, and restorable area with a restoration score of >0.6 of tropical rainforest landscapes and their national pledges to the Bonn Challenge.
Table S2. Mean ROS, total area, restorable area, and restorable area with a restoration score of >0.6 of ecoregions within tropical rainforest landscapes.
Table S3. Mean ROS, study area, restorable area, and restorable area with a restoration score of >0.6 of KBAs within tropical rainforest landscapes.
Table S4. Mean ROS, total area, restorable area, and restorable area with a restoration score of >0.6 of tropical rainforest landscapes among hotspots for biodiversity conservation.