

Abstract
Neuronal apoptosis is the main pathological feature of spinal cord injury (SCI), while autophagy contributes to ameliorating neuronal damage via inhibition of apoptosis. Here, we investigated the role of tectonic family member 2 (TCTN2) long non‐coding RNA on apoptosis and autophagy in SCI. TCTN2 was down‐regulated in the spinal cord tissues of a rat model of SCI and in oxygen–glucose deprivation‐induced hypoxic SY‐SH‐5Y cells, while microRNA‐216b (miR‐216b) was up‐regulated. Overexpression of TCTN2 reduced neuron apoptosis by inducing autophagy, and TCTN2 was observed to negatively regulate miR‐216b. Furthermore, TCTN2 promoted autophagy to repress apoptosis through the miR‐216b–Beclin‐1 pathway, and overexpression of TCTN2 improved neurological function in the SCI rat model. In summary, our data suggest that TCTN2 enhances autophagy by targeting the miR‐216b–Beclin‐1 pathway, thereby ameliorating neuronal apoptosis and relieving spinal cord injury.
Keywords: autophagy, Beclin‐1, miR‐216b, neuronal apoptosis, spinal cord injury, TCTN2
Abbreviations
- 3‐MA
3‐methyladenine
- AGO2
Argonaute2
- BBB scoring
Basso, Beattie, and Bresnahan scoring
- ECL
electrochemical luminescence
- lncRNA
long non‐coding RNA
- miR‐216b
microRNA‐216b
- OGD
oxygen–glucose deprivation
- qRT‐PCR
quantitative real‐time PCR
- RIP
RNA immunoprecipitation
- SCI
spinal cord injury
- TCTN2
tectonic family member 2
Apoptosis, also called programmed cell death, refers to an intrinsic pathway of cell death caused by various physiological or pathological signals, and inappropriate apoptosis is closely associated with human disorders 1, 2. Neuronal apoptosis is a main factor of neuronal loss in diverse neurological diseases, especially in neurodegenerative diseases 3. Neuronal apoptosis is one of the primary features observed in spinal cord injury (SCI), leading to a certain degree of neurological damage, necrosis, or autonomic nervous dysfunction 4, 5. However, apoptosis does not function alone in determining the fate of a cell 6. Autophagy, a dynamic process that triggers the self‐digestion of damaged organelles and misfolded proteins in cells, is believed to engage in a complex interplay with apoptosis 7.
In SCI model rats, autophagy has been verified to decrease neuronal damage and promote locomotor recovery, through suppressing neuronal apoptosis after SCI 8. Beclin‐1 protein has been proven to be associated with both autophagy and apoptosis 9. In spinal cord neurons, overexpression of Beclin‐1 remarkably enhances autophagy and reverses mechanical injury‐induced cell apoptosis, suggesting a role of Beclin‐1 in promoting autophagy and protecting neurons from apoptosis in SCI 10. In addition, up‐regulation of Beclin‐1 by vascular endothelial growth factor 165 contributes to increasing autophagy to attenuate SCI 11. These findings identify an essential role of Beclin‐1 in affecting autophagy, neuronal apoptosis, and the subsequent SCI development. Furthermore, Beclin‐1 has been identified as a target of microRNA‐216b (miR‐216b) and the miR‐216b–Beclin‐1 axis regulates both autophagy and apoptosis in human Tenon's capsule fibroblasts 12. The interaction between miR‐216b and Beclin‐1 and its effect on autophagy and apoptosis have not been investigated in SCI.
Tectonic family member 2 (TCTN2) belongs to the Tectonic (TCTN) family, and mutations in TCTN2 have been recently reported to disturb neural tube development in human and mouse, and loss of TCTN3 causes neuronal apoptosis 13. TCTN2 knockout shows neural tube defects in mouse 14, indicating a crucial influence of TCTN2 on neuronal apoptosis. The binding sites between miR‐216b and the long non‐coding RNA (lncRNA) TCTN2 (transcript: ENSMUST00000100706) predicted with the bioinformatics method imply a potential interaction between them. LncRNAs refer to RNA transcripts with more than 200 nucleotides that do not encode proteins, and they widely regulate gene expression in diverse biological and pathological processes. Based on the above findings, we infer that TCTN2 may participate in the neuronal apoptosis via interplay with miR‐216b to affect its target, Beclin‐1. Therefore, we undertook this study to explore whether lncRNA TCTN2 plays a role in autophagy and neuronal apoptosis, aiming to illuminate the function of this long non‐coding transcript in SCI progression for the first time.
Materials and methods
Animal treatment
All the animal experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals of the Chinese National Institutes of Health and approved by the Animal Care Committee of the Genernal Hospital, Tianjin Medical University. Sprague–Dawley rats (200–250 g) were used in the study and to ensure the animals did not suffer unnecessarily at any stage, they were housed in a clean environment with controlled temperature (23–25 °C) and provided with ad libitum food and water. SCI models (SCI group, n = 6) were constructed according to a previous protocol 15. To relieve pain, rats were anesthetized by intraperitoneal injection with 10% chloral hydrate (3 mL·kg−1), and then kept in prone position on the console. A laminectomy at the T9–T10 spinous process was conducted to thoroughly expose the spinal cord. Then spinal cord crush was performed with a 10 g weight dropped from a height of 5 cm to strike the spinal cord surface. The muscle and skin were sutured in layers. Paralysis of the lower limbs, tail swing, and spinal cord hematoma formation were observed in a successful SCI model. Only laminectomy was conducted in sham rats (sham, n = 6). After the SCI surgery, the bladder of the rats was massaged twice per day until the spontaneous voiding function of the bowel recovered.
Basso, Beattie, and Bresnahan scoring
The motor function recovery of rat was estimated using Basso, Beattie, and Bresnahan (BBB) scoring with a 21‐point open field locomotor scale 8, in which a score of 0 indicates complete immobility and 21 indicates normal motor function. Double‐blinded observation of the rats’ locomotion was carried out respectively at 1, 6, 12, 24, and 48 h after SCI. The hind limb movements, stepping, stability, trunk position, coordination, paw placement, toe clearance, and tail position of the rats were observed and recorded. Two observers blinded to the groups obtained the BBB scores individually for mean value calculation.
Quantitative real‐time PCR
At 48 h post‐SCI, rats were sacrificed and the spinal cord tissues located 1 cm above and below the injury site were taken for total RNA extraction using Trizol reagent (Thermo Fisher Scientific, Waltham, MA, USA). RNA was quantified with a Nanodrop 2000 system and reverse‐transcribed with a Taqman Reverse Transcription Kit (Thermo Fisher Scientific). Subsequently, SYBR Green Master Mix (Applied Biosystems) was utilized for the quantitative RT‐PCR on an ABI PRISM 7500 sequence detection system (Thermo Fisher Scientific). Oligonucleotide primers were synthesized by Sangon Biotech (Shanghai, China). The relative expressions of TCTN2 and miR‐216b were calculated with the 2−∆∆Ct method.
Western blot analysis
Spinal cord tissues were lysed in RIPA lysis buffer (Beyotime, China) containing protease inhibitors. After centrifugation, the supernatant was collected and the protein extraction was separated with 12% SDS/PAGE. Protein was transferred to polyvinylidene difluoride membranes (Bio‐Rad Laboratories, Hercules, CA, USA) and then blocked in Tris‐buffered saline with 5% non‐fat milk for 2 h at room temperature. The membranes were probed with specific primary antibodies (Abcam, Cambridge, UK) against Beclin‐1 and β‐actin at 4 °C overnight. The membrane was washed and incubated in horseradish peroxidase‐bound secondary antibodies for 2 h at room temperature and imaged on a Tanon Chemiluminescence Imaging system with an ECL regent (Thermo Fisher Scientific).
Cell culture and hypoxia treatment
The human hippocampal neuron‐derived cell line SY‐SH‐5Y was cultured in Ham's F12 medium supplemented with Eagle's minimum essential medium (EMEM) (1 : 1), 1% non‐essential amino acids, 2 mm glutamine, 15% fetal bovine serum and maintained at 37 °C with 5% CO2. An oxygen–glucose deprivation (OGD) assay was performed to mimic the pathological conditions of ischemia for neuronal injury induction through a reduction in the supplement of oxygen and glucose 16. Neurons were incubated in OGD solution in a controlled hypoxic incubator (Thermo Forma, San Jose, CA, USA) with an environment of 94% N2/5%CO2/1% O2 at 37 °C for 1 h. The neurons were then cultured in OGD solution with 5.5 mm glucose at 37 °C with 5% CO2 for 24 h for reoxygenation.
Flow cytometry
After OGD treatment, SY‐SH‐5Y cell apoptosis was measured with an annexin V–FITC/propidium iodide cell apoptosis detection kit (Sigma‐Aldrich, St Louis, MO, USA) followed by flow cytometric analysis. SY‐SH‐5Y cells (2 × 105/well) were seeded in six‐well plates and grown to 70–80% confluence. The cells were harvested and stained with 200 μL annexin V‐FITC and 10 μL propidium iodide, followed by flow cytometric analysis on a flow cytometer (FACSCalibur, BD Biosciences, Franklin Lakes, NJ, USA).
Cell transfection
The pcDNA‐TCTN2 vector (with pcDNA acting as negative control), miR‐216b mimic and its negative control, pre‐NC, were synthesized by GenePharma (Shanghai, China). These plasmids were transfected into SY‐SH‐5Y cells using Lipofectamine 2000 (Invitrogen) according to the manufacture's manual; 48 h following cell transfection, the expression level of TCTN2, miR‐216b, and Beclin‐1 protein was analyzed.
RNA immunoprecipitation
Binding between TCTN2 and miR‐216b was estimated with RNA immunoprecipitation (RIP) assay using the RNA‐Binding Protein Immunoprecipitation Kit (Millipore, Boston, MA, USA). SY‐SH‐5Y cells were lysed and the lysate was incubated with Argonaute2 (AGO2) antibody or negative control (normal mouse IgG, Abcam). RNA–protein complexes were immunoprecipitated with protein A agarose magnetic beads and the unbound material was washed off. RNA was extracted by using Trizol reagent (Invitrogen) and quantitative real‐time PCR (qRT‐PCR) was performed to detect the expression of TCTN2. Immunoprecipitation–western blotting was conducted to measure the AGO2 protein level.
RNA pull‐down assay
RNA pull‐down assay was performed using a Magnetic RNA‐Protein Pull‐Down Kit (Thermo Fisher Scientific). A DNA probe complementary to TCTN2 was synthesized and biotinylated (GenePharma), and 3 μg of streptavidin‐labeled TCTN2 was mixed with protein in Protein‐RNA Binding Buffer. The biotinylated RNA was incubated with cell lysate and magnetic beads. After the beads were washed three times, the RNA‐binding protein complexes were finally eluted and the enriched RNA was determined by qRT‐PCR analysis.
Overexpression of TCTN2 in SCI model rat
To verify the role of TCTN2 in SCI progression in vivo, an SCI rat model was established. The vector highly expressing TCTN2 named pcDNA‐TCTN2 (volume of 5 μL) and its negative control pcDNA (volume of 5 μL) were injected into the spinal cord of rats (n = 6 in each group) using a glass micro‐pipette attached to a pico spritzer (Parker Instrumentation, Fairfield, NJ, USA) 17. BBB scoring was carried out at 1, 6, 12, 24, and 48 h following surgery. Rats were sacrificed at 48 h following SCI and the spinal cord tissues were obtained for the experiments that followed.
Statistical analysis
spss statistics 21.0 software (IBM Corp., Armonk, NY, USA) was used for statistical analyses. All experiments were repeated at least three times, and all the data are presented as mean ± standard deviation (SD). Two‐group comparisons were analyzed with Student's t test, and a value of P < 0.05 was considered significantly different.
Results
Down‐regulation of TCTN2 in SCI rat model
To evaluate the expression level of TCTN2 in SCI, an SCI rat model was constructed. The neurological function of the SCI rat model was assessed with BBB scoring at 1, 6, 12, 24, and 48 h after SCI. Compared with the rat in sham group (n = 6), the BBB score in SCI rat (SCI, n = 6) was clearly lower (Fig. 1A). Forty‐eight hours after SCI, the spinal tissue of the rats was isolated and the expression of TCTN2, miR‐216b, and Beclin‐1 protein in the tissue was determined. The expression of TCTN2 was dramatically diminished in spinal tissue of the SCI group (SCI, n = 6) when compared with the sham group (n = 6), while miR‐216b level was markedly elevated in the SCI group (Fig. 1B). However, the Beclin‐1 protein level was significantly decreased in spinal tissue of the SCI group (SCI, n = 6) (Fig. 1C). The results indicated the abnormal expression of TCTN2, miR‐216b, and Beclin‐1 protein in SCI, implying their potential influence on SCI initiation and development.
Figure 1.

Down‐regulation of TCTN2 in SCI rat model and OGD‐induced neurons and its influence on neuronal apoptosis. SCI rat model (SCI, n = 6) was constructed, and the rats in sham group (n = 6) served as control. (A) Neurological function of rats was assessed with BBB scoring at 1, 6, 12, 24, and 48 h after SCI establishment. (B) Forty‐eight hours following SCI, the spinal tissue of mice was taken and the expression of TCTN2 and miR‐216b in spinal tissue was quantified with qRT‐PCR. (C) The protein expression of Beclin‐1 was determined with western blot 48 h after SCI. *P < 0.05 compared with the sham group. The error bars indicated SD. The experiments were repeated three times. Comparisons between the two groups were analyzed with Student's t test.
Down‐regulation of TCTN2 in OGD‐induced neurons and its influence on neuronal apoptosis
SY‐SH‐5Y was treated with OGD to imitate a hypoxic environment, and the expression of TCTN2, miR‐216b, and Beclin‐1 protein in control and OGD‐treated cells was detected. Figure 2A shows that TCTN2 expression was suppressed in SY‐SH‐5Y cells treated with OGD, while the expression of miR‐216b was enhanced. Nevertheless, the protein expression of Beclin‐1 was inhibited in SY‐SH‐5Y cells treated with OGD (Fig. 2B). To assess the influence of TCTN2 on autophagy and neuronal apoptosis, SY‐SH‐5Y cells were divided into five groups: control, OGD, OGD+pcDNA, OGD+pcDNA‐TCTN2, and OGD+pcDNA‐TCTN2+3‐methyladenine (3‐MA), with 3‐MA serving as an autophagy inhibitor. The results indicated that transfection of pcDNA‐TCTN2 recovered the TCTN2 expression that was repressed by OGD, but no difference was noted after 3‐MA treatment (Fig. 2C). An evaluation of apoptotic status showed that OGD treatment induced apoptosis, pcDNA‐TCTN2 relieved apoptosis caused by OGD, but it was further reversed by 3‐MA treatment (Fig. 2D). Beclin‐1 and LC3II protein expression was restrained with OGD treatment but augmented by TCTN2 overexpression, and then inhibited after 3‐MA treatment (Fig. 2E). The protein expression of P62 was increased by OGD but decreased with pcDNA‐TCTN2 transfection, which was finally reversed by 3‐MA treatment; but the LC3I level remained unchanged (Fig. 2E). LC3 is the autophagy‐associated protein that is post‐translationally cleaved and localizes in the cytosol (LC3I) or in autophagosomal membranes (LC3II). Hence detection of LC3II was used to evaluate the abundance of autophagosomes. The above results revealed that overexpression of TCTN2 reduced neuronal apoptosis by inducing autophagy.
Figure 2.
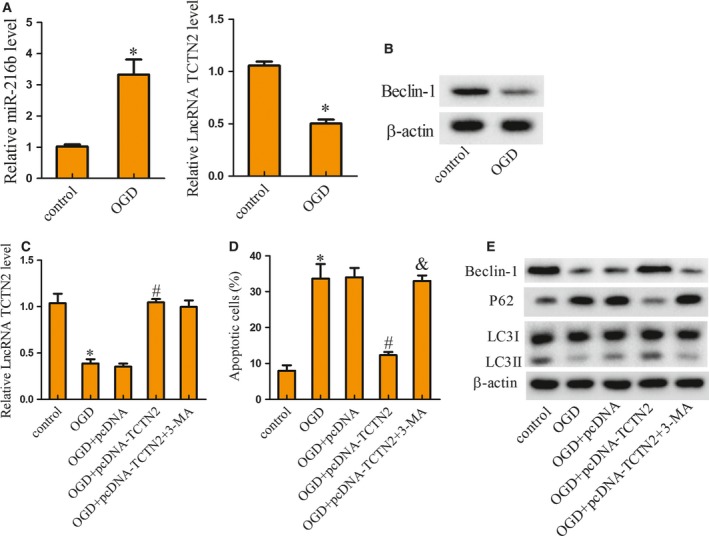
Cells of the human hippocampal neuron‐derived cell line, SY‐SH‐5Y were grouped into control and OGD for detection of TCTN2, miR‐216b, and Beclin‐1 protein expression. (A) After OGD treatment, the expression of TCTN2 and miR‐216b in SY‐SH‐5Y cells was examined by qRT‐PCR. (B) The protein level of Beclin‐1 was analyzed with western blot. (C) The expression of TCTN2 in SY‐SH‐5Y cells was estimated with qRT‐PCR. (D) SY‐SH‐5Y cell apoptosis was analyzed by flow cytometry. (E) The expression of Beclin‐1, P62, LC3I, and LC3II protein in SY‐SH‐5Y cells was assessed with western blot. *P < 0.05 compared with control; # P < 0.05 compared with OGD+pcDNA; & P < 0.05 compared with OGD+pcDNA‐TCTN2. The error bars indicate SD. The experiments were repeated three times. Comparisons between the two groups were analyzed with Student's t test.
Interaction between TCTN2 and miR‐216b
The putative binding site between TCTN2 and miR‐216b is shown in Fig. 3A. Their binding and interaction in SY‐SH‐5Y cells were assessed. AGO2 antibody was used in the RIP assay. Compared with IgG, a mass of TCTN2 and miR‐216b was detected with the AGO2 antibody (Fig. 3B). In the RNA pull‐down assay, AGO2 in the pull‐down complex of TCTN2 was analyzed, and miR‐216b was deposited extensively in the pull‐down complex of TCTN2, while a slight increase was observed in the negative control (NC) group (Fig. 3C). These data identified miR‐216b as a target of TCTN2 in SY‐SH‐5Y cells.
Figure 3.
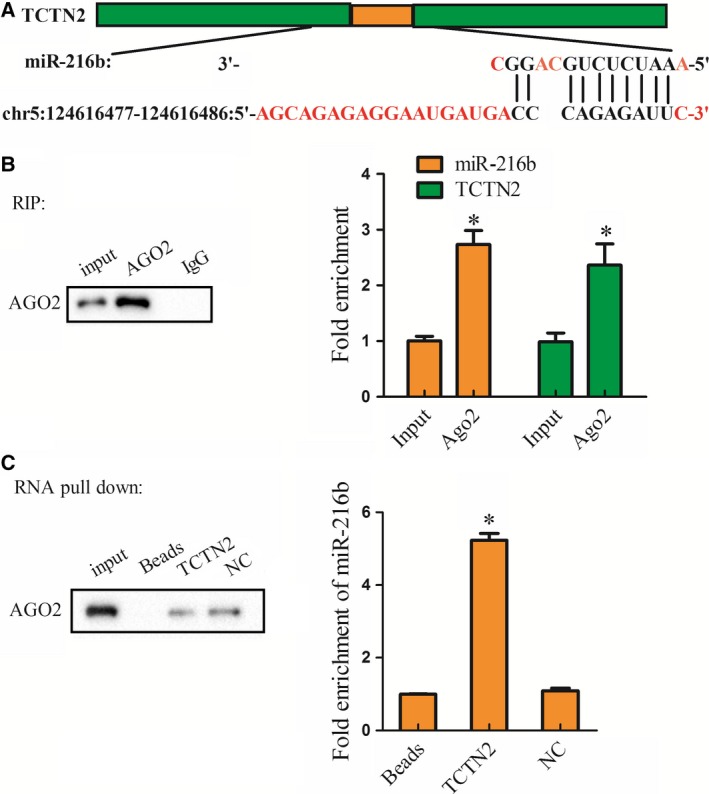
Relationship between TCTN2 and miR‐216b. RNA immunoprecipitation (RIP) and RNA pull‐down assay were performed to explore the interaction between TCTN2 and miR‐216b in SY‐SH‐5Y cells. (A) Bioinformatics analysis (DIANA) predicted the binding sites between TCTN2 and miR‐216b. (B) RIP assay was performed to determine the binding condition between TCTN2 and miR‐216b, and immunoprecipitation–western blotting was performed to determine whether TCTN2 and miR‐216b are present in the AGO2 protein complex. (C) RNA pull‐down assay was conducted to examine the interaction between TCTN2 and miR‐216b. *P < 0.05 compared with Input or negative control (NC). The error bars indicate SD. The experiments were repeated three times. Comparisons between the two groups were analyzed with Student's t test.
TCTN2 regulates autophagy and apoptosis through the miR‐216b–Beclin‐1 pathway
To investigate the role of TCTN2 and miR‐216b and their interaction in neuronal apoptosis, SY‐SH‐5Y cells were assigned to six groups: control, OGD, OGD+pcDNA, OGD+pcDNA‐TCTN2, OGD+pcDNA‐TCTN2+pre‐NC (the negative control of miR‐216 mimic), and OGD+pcDNA‐TCTN2+miR‐216b mimic. Compared with the control group, miR‐216b expression was promoted by OGD treatment but then inhibited with pcDNA‐TCTN2 transfection, which was further inverted by miR‐216b mimic, while the Beclin‐1 protein expression change was the contrary of that of miR‐216b (Fig. 4A). The protein level of P62 was increased by OGD but decreased with pcDNA‐TCTN2 transfection, which was finally reversed by up‐regulation of miR‐216b. The expression of LC3II protein was decreased with OGD treatment but increased after pcDNA‐TCTN2 transfection, which was finally reversed with overexpressed miR‐216b mimic (Fig. 4B). Neuronal apoptosis induced by OGD was mitigated by pcDNA‐TCTN2 transfection but then inverted by miR‐216b mimic (Fig. 4C). Herein, we showed that TCTN2 regulated autophagy and apoptosis through miR‐216b.
Figure 4.
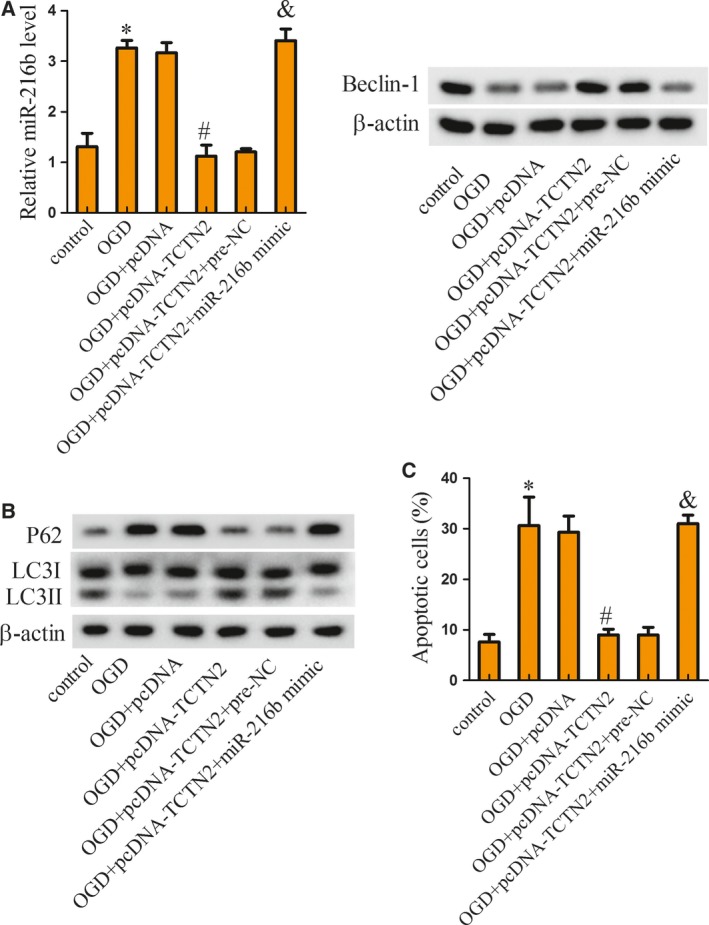
TCTN2 regulated autophagy and apoptosis through miR‐216b–Beclin‐1 pathway. SY‐SH‐5Y cells were assigned to six groups: control, OGD, OGD+pcDNA, OGD+pcDNA‐TCTN2, OGD+pcDNA‐TCTN2+pre‐NC, and OGD+pcDNA‐TCTN2+miR‐216b mimic. (A) Forty‐eight hours after cell transfection, the expression of miR‐216b and Beclin‐1 protein was evaluated with qRT‐PCR and western blot, separately. (B) The expression of P62, LC3I, and LC3II protein in SY‐SH‐5Y cells was assessed with western blot. (C) SY‐SH‐5Y cell apoptosis was estimated with flow cytometric analysis. *P < 0.05 compared with control; # P < 0.05 compared with OGD+pcDNA; & P < 0.05 compared with OGD+pcDNA‐TCTN2+pre‐NC. The error bars indicate SD. The experiments were repeated three times. Comparisons between the two groups were analyzed with Student's t test.
Overexpression of TCTN2 improved neurological function in SCI rat model
To validate the influence of TCTN2 expression on SCI progression in vivo, the SCI rat model was intrathecally injected with pcDNA (n = 6) or pcDNA‐TCTN2 (n = 6) vector. BBB scoring was performed to estimate the neurological function at 1, 6, 12, 24, and 48 h after SCI. Compared with the pcDNA (n = 6) group, the BBB score of rats with pcDNA‐TCTN2 (n = 6) was clearly elevated from 12 h to 48 h (Fig. 5A). Notably, the expression of TCTN2 was also increased with pcDNA‐TCTN2 introduction, while the miR‐216b expression was reduced (Fig. 5B). P62 protein level was decreased in the pcDNA‐TCTN2 group, while the expression of LC3II protein was clearly elevated (Fig. 5C), implying that autophagy was facilitated by overexpression of TCTN2. These findings demonstrated that overexpression of TCTN2 improved neurological function in the SCI rat model.
Figure 5.
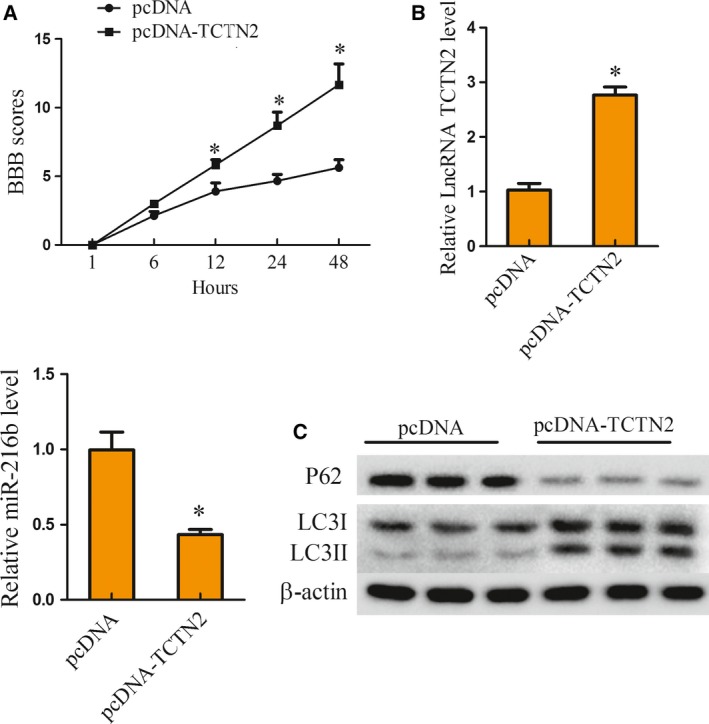
Overexpression of TCTN2 improved neurological function in SCI rat model. SCI rat model was intrathecally injected with pcDNA (n = 6) or pcDNA‐TCTN2 (n = 6) vector. (A) BBB scoring was used to estimate the neurological function at 1, 6, 12, 24, and 48 h after SCI induction. (B) Forty‐eight hours later, the spinal tissue of mice was taken and the expression of TCTN2 and miR‐216b in spinal tissue of rats was examined by qRT‐PCR. (C) The P62, LC3I, and LC3II protein level in spinal tissue was analyzed with western blot 48 h after SCI. *P < 0.05 compared with pcDNA. The error bars indicate SD. The experiments were repeated three times. Comparisons between the two groups were analyzed with Student's t test.
Discussion
SCI is a serious neurological trauma, but its pathogenesis and the key influencing factors have not been precisely illuminated 18. Neuronal apoptosis is a main factor in neuronal loss and is widely considered to be implicated in the development of SCI 19. In addition, neuronal autophagy has also been shown to decrease neuronal damage and promote locomotor restoration by restraining of apoptosis after SCI in rats 8, 20. In the current study, we utilized an in vivo SCI model and human neurons to clarify the role of autophagy in SCI and its interrelationship with apoptosis, in which the function of lncRNA TCTN2 is initially highlighted. It can be summarized that TCTN2 increases autophagy by targeting the miR‐216b–Beclin‐1 pathway to control neuronal apoptosis, thus relieving spinal cord injury. Our study may provide new promising targets for SCI treatment and prognostic evaluation.
A regulatory effect of lncRNAs on cell autophagy or apoptosis has been validated in human disease. For instance, lncRNA MALAT1 promotes autophagy resulting in suppression of tumor cell apoptosis in multiple myeloma 21. LncRNAs also have been reported to regulate SCI initiation and progression, mostly by affecting cell autophagy or apoptosis. LncRNA SCIR1 down‐regulation has been proven to play a detrimental role in the pathophysiology of traumatic SCI through stimulating astrocyte proliferation and migration 22. In acute SCI, knockdown of lncRNA BDNF‐AS suppresses neuronal cell apoptosis 23, while lncRNA XIST contributes to neuronal apoptosis in rat SCI 17. Suppression of lncRNA Sox2ot decreases apoptosis and autophagy to inhibit H2O2‐induced PC‐12 cell injury and injury in spinal cord 24. The present study illustrates that lncRNA TCTN2 represses neuronal apoptosis by increasing autophagy, revealing a neuroprotective role of TCTN2 in SCI to benefit locomotor recovery of SCI rats. This result is in line with the significance of the TCTN2 gene in neural tube development 14.
Most lncRNAs show regulatory effects in disease initiation and progression by serving as the competing endogenous RNAs of miRNAs 25. By competitively binding miR‐494, lncRNA XIST contributes to neuronal apoptosis in rat SCI, participating in the pathogenesis of SCI 17. MALAT1 exhibits a neuroprotective role in spinal cord ischemia–reperfusion injury through miR‐204 regulation 26. The present study innovatively identifies miR‐216b as a down‐stream target of lncRNA TCTN2, uncovering the critical relationship between TCTN2 and miR‐216b in human disease. MiR‐216b is a member of the miRNAs that negatively regulates gene expression to take part in human diseases such as nasopharyngeal carcinoma 27, cervical cancer 28, and hepatocellular carcinoma 29, acting as a tumor suppressor. Moreover, miR‐216b has been noted to modulate autophagy and reduce LC3II in hepatocellular carcinoma cells, and miR‐216b represses autophagy and apoptosis through Beclin‐1 in human Tenon's capsule fibroblasts induced by hydroxycamptothecin 12, 30. Beclin‐1 is a key autophagy‐related protein and apoptosis regulator 31, and Beclin‐1‐mediated autophagy has been reported to protect spinal cord neurons from apoptosis 10, demonstrating its neuroprotective role in SCI. Our study confirms once more the function of Beclin‐1 in autophagy, apoptosis, and SCI progression, but the role of miR‐216b in affecting neuronal apoptosis and SCI, achieved by controlling Beclin‐1, was discovered for the first time.
In summary, these findings highlight the regulatory effects of lncRNA TCTN2 on neuronal autophagy and apoptosis by targeting the miR‐216b–Beclin‐1 pathway, which is beneficial for SCI improvement, offering novel insights for SCI amelioration and neurological function recovery after SCI. However, some limitations still exist in this work. In the first place, more potential targets of lncRNA TCTN2 need to be explored to seek the optimal miRNA that has the maximal influence on neuronal autophagy, apoptosis, and SCI. In addition, many other targeting genes of miR‐216b and the other competing endogenous RNAs of miR‐216 also deserve further investigation in the future.
Conflict of interest
The authors declare no conflict of interest.
Author contributions
CW conceived and designed this study, and contributed to the acquisition of the data and the preparation of the manuscript. XR designed the study, analyzed the data, drafted and made crucial revision of the manuscript for important intellectual content. YN contributed to the conception of the study, collection and analysis of the data, manuscript editing and revision. All the authors approved the final version.
Contributor Information
Chun‐xiao Wan, Email: cnuoe1@163.com.
Ya‐li Niu, Email: lojil1@163.com.
References
- 1. Hassan M, Watari H, AbuAlmaaty A, Ohba Y and Sakuragi N (2014) Apoptosis and molecular targeting therapy in cancer. Biomed Res Int 2014, 150845. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 2. Elmore S (2007) Apoptosis: a review of programmed cell death. Toxicol Pathol 35, 495–516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Radi E, Formichi P, Battisti C and Federico A (2014) Apoptosis and oxidative stress in neurodegenerative diseases. J Alzheimers Dis 42 (Suppl 3), S125–S152. [DOI] [PubMed] [Google Scholar]
- 4. Liu XJ, Zheng XP, Zhang R, Guo YL and Wang JH (2015) Combinatorial effects of miR‐20a and miR‐29b on neuronal apoptosis induced by spinal cord injury. Int J Clin Exp Pathol 8, 3811–3818. [PMC free article] [PubMed] [Google Scholar]
- 5. Chang CK, Chou W, Lin HJ, Huang YC, Tang LY, Lin MT and Chang CP (2014) Exercise preconditioning protects against spinal cord injury in rats by upregulating neuronal and astroglial heat shock protein 72. Int J Mol Sci 15, 19018–19036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Booth LA, Tavallai S, Hamed HA, Cruickshanks N and Dent P (2014) The role of cell signalling in the crosstalk between autophagy and apoptosis. Cell Signal 26, 549–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Fernandez A, Ordonez R, Reiter RJ, Gonzalez‐Gallego J and Mauriz JL (2015) Melatonin and endoplasmic reticulum stress: relation to autophagy and apoptosis. J Pineal Res 59, 292–307. [DOI] [PubMed] [Google Scholar]
- 8. Tang P, Hou H, Zhang L, Lan X, Mao Z, Liu D, He C, Du H and Zhang L (2014) Autophagy reduces neuronal damage and promotes locomotor recovery via inhibition of apoptosis after spinal cord injury in rats. Mol Neurobiol 49, 276–287. [DOI] [PubMed] [Google Scholar]
- 9. Kang R, Zeh HJ, Lotze MT and Tang D (2011) The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ 18, 571–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Wang ZY, Lin JH, Muharram A and Liu WG (2014) Beclin‐1‐mediated autophagy protects spinal cord neurons against mechanical injury‐induced apoptosis. Apoptosis 19, 933–945. [DOI] [PubMed] [Google Scholar]
- 11. Wang H, Wang Y, Li D, Liu Z, Zhao Z, Han D, Yuan Y, Bi J and Mei X (2015) VEGF inhibits the inflammation in spinal cord injury through activation of autophagy. Biochem Biophys Res Commun 464, 453–458. [DOI] [PubMed] [Google Scholar]
- 12. Xu X, Fu Y, Tong J, Fan S, Xu K, Sun H, Liang Y, Yan C, Yuan Z and Ge Y (2014) MicroRNA‐216b/Beclin 1 axis regulates autophagy and apoptosis in human Tenon's capsule fibroblasts upon hydroxycamptothecin exposure. Exp Eye Res 123, 43–55. [DOI] [PubMed] [Google Scholar]
- 13. Wang B, Zhang Y, Dong H, Gong S, Wei B, Luo M, Wang H, Wu X, Liu W, Xu X et al (2018) Loss of Tctn3 causes neuronal apoptosis and neural tube defects in mice. Cell Death Dis 9, 520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Sang L, Miller JJ, Corbit KC, Giles RH, Brauer MJ, Otto EA, Baye LM, Wen X, Scales SJ, Kwong M et al (2011) Mapping the NPHP‐JBTS‐MKS protein network reveals ciliopathy disease genes and pathways. Cell 145, 513–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Guo Y, Wang F, Li H, Liang H, Li Y, Gao Z and He X (2018) Metformin protects against spinal cord injury by regulating autophagy via the mTOR signaling pathway. Neurochem Res 43, 1111–1117. [DOI] [PubMed] [Google Scholar]
- 16. Li L, Jiang HK, Li YP and Guo YP (2015) Hydrogen sulfide protects spinal cord and induces autophagy via miR‐30c in a rat model of spinal cord ischemia‐reperfusion injury. J Biomed Sci 22, 50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Gu S, Xie R, Liu X, Shou J, Gu W and Che X (2017) Long coding RNA XIST contributes to neuronal apoptosis through the downregulation of AKT phosphorylation and is negatively regulated by miR‐494 in rat spinal cord injury. Int J Mol Sci 18, 732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Fagoe ND, van Heest J and Verhaagen J (2014) Spinal cord injury and the neuron‐intrinsic regeneration‐associated gene program. Neuromolecular Med 16, 799–813. [DOI] [PubMed] [Google Scholar]
- 19. Chen M, Xia X, Zhu X, Cao J, Xu D, Ni Y, Liu Y, Yan S, Cheng X, Liu Y et al (2014) Expression of SGTA correlates with neuronal apoptosis and reactive gliosis after spinal cord injury. Cell Tissue Res 358, 277–288. [DOI] [PubMed] [Google Scholar]
- 20. Yao J, Zheng K and Zhang X (2015) Rosiglitazone exerts neuroprotective effects via the suppression of neuronal autophagy and apoptosis in the cortex following traumatic brain injury. Mol Med Rep 12, 6591–6597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Gao D, Lv AE, Li HP, Han DH and Zhang YP (2017) LncRNA MALAT‐1 elevates HMGB1 to promote autophagy resulting in inhibition of tumor cell apoptosis in multiple myeloma. J Cell Biochem 118, 3341–3348. [DOI] [PubMed] [Google Scholar]
- 22. Wang J, Hu B, Cao F, Sun S, Zhang Y and Zhu Q (2015) Down regulation of lncSCIR1 after spinal cord contusion injury in rat. Brain Res 1624, 314–320. [DOI] [PubMed] [Google Scholar]
- 23. Zhang H, Li D, Zhang Y, Li J, Ma S, Zhang J, Xiong Y, Wang W, Li N and Xia L (2018) Knockdown of lncRNA BDNF‐AS suppresses neuronal cell apoptosis via downregulating miR‐130b‐5p target gene PRDM5 in acute spinal cord injury. RNA Biol 15, 1071–1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Yin D, Zheng X, Zhuang J, Wang L, Liu B and Chang Y (2018) Downregulation of long noncoding RNA Sox2ot protects PC‐12 cells from hydrogen peroxide‐induced injury in spinal cord injury via regulating the miR‐211‐myeloid cell leukemia‐1 isoform2 axis. J Cell Biochem 119, 9675–9684. [DOI] [PubMed] [Google Scholar]
- 25. Qi X, Zhang DH, Wu N, Xiao JH, Wang X and Ma W (2015) ceRNA in cancer: possible functions and clinical implications. J Med Genet 52, 710–718. [DOI] [PubMed] [Google Scholar]
- 26. Qiao Y, Peng C, Li J, Wu D and Wang X (2018) LncRNA MALAT1 is neuroprotective in a rat model of spinal cord ischemia‐reperfusion injury through miR‐204 regulation. Curr Neurovasc Res 15, 211–219. [DOI] [PubMed] [Google Scholar]
- 27. Deng M, Tang H, Zhou Y, Zhou M, Xiong W, Zheng Y, Ye Q, Zeng X, Liao Q, Guo X et al (2011) miR‐216b suppresses tumor growth and invasion by targeting KRAS in nasopharyngeal carcinoma. J Cell Sci 124, 2997–3005. [DOI] [PubMed] [Google Scholar]
- 28. He S, Liao B, Deng Y, Su C, Tuo J, Liu J, Yao S and Xu L (2017) MiR‐216b inhibits cell proliferation by targeting FOXM1 in cervical cancer cells and is associated with better prognosis. BMC Cancer 17, 673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Liu FY, Zhou SJ, Deng YL, Zhang ZY, Zhang EL, Wu ZB, Huang ZY and Chen XP (2015) MiR‐216b is involved in pathogenesis and progression of hepatocellular carcinoma through HBx‐miR‐216b‐IGF2BP2 signaling pathway. Cell Death Dis 6, e1670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Yuan P, Cao W, Zang Q, Li G, Guo X and Fan J (2016) The HIF‐2alpha‐MALAT1‐miR‐216b axis regulates multi‐drug resistance of hepatocellular carcinoma cells via modulating autophagy. Biochem Biophys Res Commun 478, 1067–1073. [DOI] [PubMed] [Google Scholar]
- 31. Katagiri H, Nakayama K, Razia S, Nakamura K, Sato E, Ishibashi T, Ishikawa M, Iida K, Ishikawa N, Otsuki Y et al (2015) Loss of autophagy‐related protein Beclin 1 may define poor prognosis in ovarian clear cell carcinomas. Int J Oncol 47, 2037–2044. [DOI] [PMC free article] [PubMed] [Google Scholar]