

Abstract
Despite reducing the prevalent foodborne pathogen Campylobacter jejuni in chickens decreases campylobacteriosis, few effective approaches are available. The aim of this study was to use microbial metabolic product bile acids to reduce C. jejuni chicken colonization. Broiler chicks were fed with deoxycholic acid (DCA), lithocholic acid (LCA), or ursodeoxycholic acid (UDCA). The birds were also transplanted with DCA modulated anaerobes (DCA-Anaero) or aerobes (DCA-Aero). The birds were infected with human clinical isolate C. jejuni 81–176 or chicken isolate C. jejuni AR101. Notably, C. jejuni 81–176 was readily colonized intestinal tract at d16 and reached an almost plateau at d21. Remarkably, DCA excluded C. jejuni cecal colonization below the limit of detection at 16 and 28 days of age. Neither chicken ages of infection nor LCA or UDCA altered C. jejuni AR101 chicken colonization level, while DCA reduced 91% of the bacterium in chickens at d28. Notably, DCA diet reduced phylum Firmicutes but increased Bacteroidetes compared to infected control birds. Importantly, DCA-Anaero attenuated 93% of C. jejuni colonization at d28 compared to control infected birds. In conclusion, DCA shapes microbiota composition against C. jejuni colonization in chickens, suggesting a bidirectional interaction between microbiota and microbial metabolites.
Introduction
Campylobacter jejuni asymptomatically colonizes in poultry gut but is one of the prevalent food borne pathogens in the developed countries. Antibiotic resistant C. jejuni has been reported in children and adults in US and worldwide [1–5]. More than 13 campylobacteriosis cases per 100,000 population were recorded in 2014 in USA, which represents a 13% increase compared to 2006–2008 and was higher than the combined incidences by the following 8 bacterial pathogens [6]. A total of 1.3 million individuals are afflicted by the disease, resulting in 76 deaths every year [7]. Although antibiotics treatment has a marginal benefit (1.32 days) on the duration of campylobacteriosis symptoms [8], it is the only current available therapy for patients with severe campylobacteriosis or for those at high risk for severe disease [9]. However, ever increasing antimicrobial resistance [10] prompts the need for immediate and sustainable counter-actions from agricultural industry to medical field. Misuse or overuse of antimicrobial agents in medical and agricultural practice is contributing to exacerbating the episodes of emerging antimicrobial resistant microbes [10]. Hence, an effective and sustainable solution is to find antimicrobial alternatives in agricultural industry and medical field.
Tremendous efforts have been put forward to prevent campylobacteriosis by reducing C. jejuni contamination in animal food sources, particularly broiler chicken meat. The intervention approaches include on-farm biosecurity measures [11], vaccines [12], probiotics [13], phages [14], and post-slaughter decontamination of poultry carcasses [15]. Reducing carcass Campylobacter counts by 2 log is estimated to decrease a 30-fold in human campylobacteriosis [16]. Although those measures to prevent C. jejuni contamination have achieved some success, improvement is needed, as evidenced by the relative consistent rate of campylobacteriosis incidences in the Morbidity and Mortality Weekly Report at CDC infectious disease database from January,1996 to June, 2017 [17].
Sparse information is currently available on using microbiota to prevent C. jejuni colonization in poultry. Microbiota transplantation has shown tremendous success against recurrent Clostridium difficile infection [18]. The production of secondary bile acids by specific bacteria are attributed to inhibit C. difficile colonization and infection [19]. Bile acids at 3–30 mM in the human small intestine [20] are the byproducts of cholesterol and biotransformed from conjugated to unconjugated primary bile acids, and to secondary bile acids. Majority of bile acids (>95%) are effectively absorbed in intestine [21]. Primary bile acids of cholic acid (CA) and chenodeoxycholic acid (CDCA) are synthesized from cholesterol in hepatocytes and conjugated with glycine or taurine [22]. In the intestine, the conjugated primary bile acids are deconjugated by bacterial bile salt hydrolase (BSH) and further altered by microbiota to produce secondary bile acids including lithocholic acid (LCA), deoxycholic acid (DCA), and ursodeoxycholic acid (UDCA). Deconjugating enzyme BSH is present in all major bacterial divisions and archaeal species in the human gut including members of genus Lactobacilli, Bifidobacteria, Clostridium, and Bacteroides [21, 23–25]. Secondary bile acid producing bacteria consist of a small population of genus Clostridium, including C. scindens, C. hiranonis, C. hylemonae (Clostridium cluster XVIa), and C. sordelli (Clostridium cluster XI) [21]. The bile acid-metabolizing bacteria such as Lactobacilli and Bifidobacteria are probiotics and they enhance health by promoting host immune homeostasis [26]. Besides bile acids influence host response, their level is associated with microbial community dynamics in the gut [27]. Bile acids directly inhibit gut microbes [28] and indirectly modulate microbiota through FXR-induced antimicrobial peptides [29]. Mice fed CA have increased class Clostridia (70 vs. 39%) compared to control mice, and genus Blautia (including Ruminococcus spp. and Clostridium cluster XIVa) expands from 8.3 to 55–62% [30]. Bile acids, particularly secondary bile acid DCA, are associated with a variety of chronic diseases, such as obesity, diabetes, and colorectal tumorigenesis [31, 32]. Only very recently, new evidences shed light on the beneficial property of secondary bile acids in health and diseases, such as gut motility [33] and C. difficile infection [19]. We found that specific pathogen free (SPF) Il10-/- mice resisted against C. jejuni 81–176 induced colitis, while the mice were susceptible to campylobacteriosis after treated with antibiotic clindamycin which kills bile acid metabolizing bacteria [34]. 16S rDNA sequencing, bioinformatic, and HPLC/MS analysis showed that clindamycin depleted all secondary bile acids including DCA. Furthermore, anaerobe metabolite DCA prevents and treats C. jejuni-induced colitis in ex-Germ Free mice [34]. However, it remains unclear whether DCA regulates C. jejuni chicken colonization and transmission.
In this study, we hypothesized that DCA prevented C. jejuni chicken colonization. Our data indicate that DCA indeed prevented against chicken colonization of C. jejuni human clinical isolate 81–176 or chicken isolate AR101. Subsequent mechanistic studies found that DCA modulated intestinal microbiota and DCA-modulated anaerobes attenuated C. jejuni chicken colonization. Thus, the action of DCA against C jejuni chicken colonization represents unique bidirectional interaction between microbiota and microbial metabolites. It would be more effective against campylobacteriosis or the pathogen colonization through modulating both microbiota and microbial metabolites.
Materials and methods
Chicken experiment
Animal experiments performed were in accordance with the Animal Research: Reporting of In Vivo Experiments (https://www.nc3rs.org.uk/arrive-guidelines). The experiments were approved by the Care and Use Committee of the University of Arkansas. Cohorts of 13 to 18 one-day-old broiler chicks obtained from Cobb-Vantress (Siloam Springs, AR) were neck-tagged and randomly assigned to floor pens with a controlled age-appropriate environment. The birds were fed a corn-soybean meal-based starter diet during 0–10 days of age and a grower diet during 11–28 days of age. The basal diet was formulated as described before [35]. Bird were fed diet supplemented with 0 or 1.5 g/kg DCA, LCA, or UDCA (all from Alfa Aesar) from d 0. Before experiment, frozen stock of C. jejuni 81–176 and AR101 (isolated at Dr. Billy Hargis’s lab at University of Arkansas at Fayetteville) were cultured micro-aerobically at 42°C for 48 hours on C. jejuni selective blood plates with 5% lysed horse blood, five antibiotics (20 mg/L cefoperazone, 50 mg/L cycloheximide, 20 gm/L trimethoprim, 20 mg/L vancomycin and 0.35 mg/L polymyxin B), 500 mg/L ferrous sulfate, and 200 mg/L triphenyl-tetrazolium chloride (TTC). The motility of C. jejuni was ensured under a microscope and routinely examined on semisolid MH (0.4% agar) plates. C. jejuni in PBS was estimated as that OD600 of 1.872 meant 1010 CFU/ml, and the bacterium were also serially diluted, cultured on the select plates, and enumerated 48 hr later. Birds were infected with a single oral gavage of 109 CFU/bird human clinical isolate C. jejuni 81–176 at 14 days of age or chicken isolate AR101 at d 5, 10, or 14. AR101 was used mainly because 81–176 was failed to be isolated in some groups of infected chickens (details in result section) and C. jejuni transmitted through chickens of all groups had to be isolated for future virulence evaluation experiments using Il10-/- mice. Most birds were infected with C. jejuni AR101 at day 10 in order to have enough time for both C. jejuni colonization and microbiome development. Chicken body weight and feed intake were measured at 0, 14, 21, and 28 days of age. Birds were sacrificed from the same pens at 16 (4 birds/pen), 21 (4 birds/pen) or 28 (4–8 birds/pen) days of age to collect cecal samples for enumerating C. jejuni. Cecal digesta samples were collected for DNA isolation or were ten-fold serially diluted with sterile PBS and cultured at 42°C for 48 hours under microaerophilic atmosphere. Colonies were enumerated and CFU per gram was calculated. The limit of detection was 100 CFU/g cecal content, which was calculated based on the least dilution factor of the samples without enrichment. To specifically identify C. jejuni, TTC and ferrous sulfate were supplemented in the select plates and C. jejuni colonies showed distinctive dark red color. To isolate and validate chicken-transmitted C. jejuni, colonies grown on the plate were routinely examined on the selective plates, MacConkey plates, and Brain Heart Infusion (BHI) plates under aerobic and microaerobic conditions.
Microbiota transplantation and C. jejuni colonization
Because C. jejuni wasn’t detected in cecal digesta samples from 28 days old birds infected with C. jejuni 81–176 and fed 1.5g/kg DCA diet, the samples were used for isolating transplantation microbiota. Briefly, the cecal content from the birds were quickly squeezed into 50 ml conical tubes and PBS and glycerol were added at final concentration of 20%. The suspension was homogenized, aliquoted, and stored in -80°C freezer. Before experiments, the microbiota stock was cultured on brain heart infusion (BHI) plates under aerobic or anaerobic conditions for 48 hours at 42°C. For culturing anaerobic microbiota, the plates were put into GasPak Jar, briefly flushed with CO2, and incubated with GasPak EZ Anaerobe Pouch System (All from BD Diagnostics). The collected bacteria were labelled as DCA modulated aerobes (DCA-Aero) and DCA modulated Anaerobes (DCA-Anaero). Chickens were orally gavaged once with 108 CFU/bird DCA-Aero or DCA-Anaero at 0 days of age. The aerobes or anaerobes in PBS were estimated as that OD600 of 1 meant 109 CFU/ml, and the bacteria were then serially diluted, cultured on BHI plates, and enumerated 24 (aerobes) or 48 hr (anaerobes) later. At 10 days of age, the birds were infected with 109 CFU/bird chicken isolate C. jejuni AR101. Cecal digesta collected at days 21 and 28 were serially diluted and cultured for C. jejuni enumeration as described above.
In vitro C. jejuni growth with various bile acids
The impact of various species of bile acids on C. jejuni growth was measured. Briefly, frozen C. jejuni 81–176 or AR101 were cultured on the select plates for 48 hours. C. jejuni at 103 CFU was inoculated into 5 ml Campylobacter Enrichment Broth (Neogen Food Safety, MI) in the presence of DCA, taurocholic acid (TCA), or CA at various concentrations (0, 5, 25 mM) with triplication. The bacteria were cultured for 24 hours at 42 0C under microaerobic condition. C. jejuni growth was measured by serial dilution and plating on the select plates.
Microbiota composition at phylum level
Cecal digesta samples were collected and DNA was extracted using bead beater disruption and phenol: chloroform separation as describe before [34]. The levels of five phylum bacteria were determined using SYBR Green PCR Master mix (Bio-Rad) on a Bio-Rad 384-well Real-Time PCR System. The PCR reactions were performed according to the manufacturer’s recommendation. The following gene primers [34, 36] were used: Universal 16S: 16s357F: 5’-CTCCTACGGGGAGGCAGCAA-3’, 16s1392R: 5’-ACGGGCGGTGTGTRC-3’; α-proteobacteria: α682F 5’-CIAGTGTAGAGGTGAAATT-3’, 908αR 5’-CCCCGTCAATTCCTTTGAGTT-3’; γ-proteobacteria: 1080γF 5’-TCGTCAGCTCGTGTYGTGA-3’ γ1202R 5’-CGTAAGGGCCATGATG-3’; Bacteroidetes: 798cfbF 5’-5’-CRAACAGGATTAGATACCCT-3’ cfb967R 5’-GGTAAGGTTCCTCGCGTAT-3’; Firmicutes: 928F-Firm 5’-TGAAACTYAAAGGAATTGACG-3’ 1040 Firm R 5’-ACCATGCACCACCTGTC-3’; Actinobacteria: Act920F3 5’-TACGGCCGCAAGGCTA-3’ Act1200R 5’-TCRTCCCCACCTTCCTCCG-3’. The relative fold change of each phylum in one sample were normalized to universal 16S. The percentage of each phylum was then calculated as the phylum relative folds divided by total folds of all five phyla.
Statistical analysis
The data for bacterial colony forming unit (CFU) were transformed by log10(CFU+1). Values are shown as mean ± standard error of the mean as indicated. One-way ANOVA were first performed at each time point to ensure statistical significance when there were more than or equal to 3 groups. Differences between groups were analyzed using unpaired t-test between infected and treatment groups with Welch’s correction using Prism 7.0 software. For statistically test between infected birds and DCA birds with log10(CFU +1) values at 0s, One Sample t test was used against value of 0. Experiments were considered statistically significant if P values were <0.05.
Results
DCA prevents C. jejuni strain 81–176 cecal colonization in chickens
Secondary bile acid DCA prevents and treats C. jejuni-induced intestinal inflammation in germ free Il10-/- mice [34]. Because chickens are the natural reservoir of C. jejuni, we then interrogated the hypothesis that DCA modulated C. jejuni colonization in chickens. The birds fed diet supplemented with 1.5 g/kg DCA were orally infected with a single dose of 109 CFU/bird human clinical isolate C. jejuni strain 81–176 at 14 days of age. C. jejuni colonization level was determined by collecting and culturing cecal digesta of the birds at 16, 21, and 28 days of age using C. jejuni select medium. Notably, no C. jejuni in cecal digesta was detected from birds without the bacterial infection, suggesting the clean facility at our chicken farm and the success of strict biosecurity measurement during our experiments. C. jejuni was readily colonized the intestinal tract at a level of 105 CFU/g cecal digesta at 16 days of age, only 2 days post infection (Fig 1). C. jejuni colonization level then increased more than 100 folds and reached an almost plateau of 2.8 x107 CFU/g cecal digesta at 21 days of age. Remarkably, DCA excluded C. jejuni cecal colonization below the limit of detection (without enrichment) at 16 and 28 days of age compared to the infected control birds. These results suggest that the secondary bile acid DCA effectively reduces C. jejuni 81–176 colonization in the intestinal tract of broiler chickens.
Fig 1. DCA prevents against C. jejuni stain 81–176 colonization in chickens.
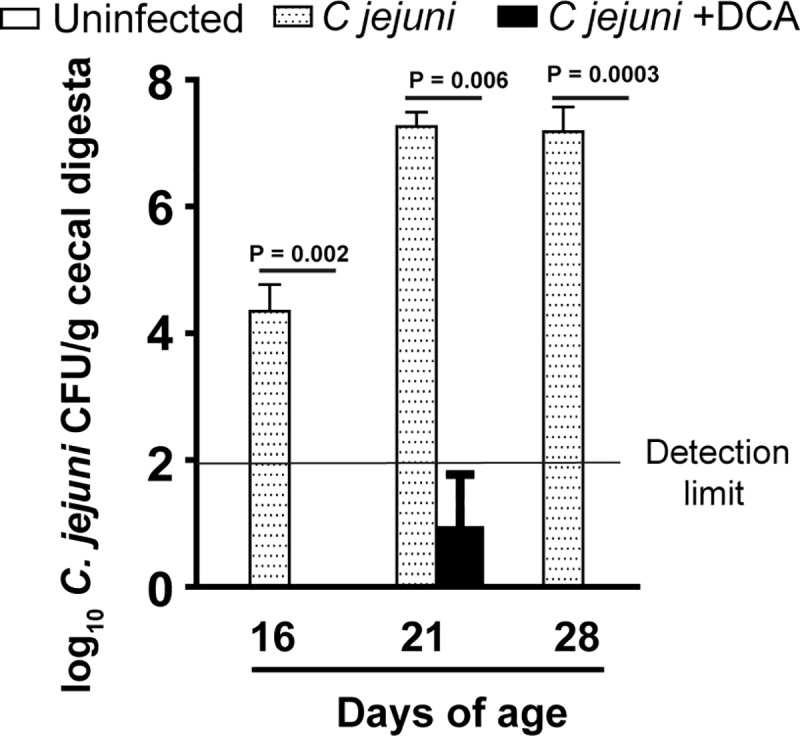
Cohorts of 13 one-day-old broiler chickens were fed 0 or 1.5 g/kg DCA diets. The birds were infected with C. jejuni 81–176 at 14 days of age. Cecal digesta were collected at 16, 21, and 28 days of age, serially diluted, and cultured on C. jejuni selective medium at 42°C. C. jejuni was counted after 48 hours of culture. All graphs depict mean ± SEM. Significant if P<0.05. Results are representative of 2 independent experiments.
DCA promotes bird growth performance
Increased level of secondary bile acids DCA has been associated with obesity [31], but the role of DCA on animal growth is unclear. To investigate the contribution of DCA on chicken growth, the bird average daily gain was measured at 14 and 28 days of age with or without C. jejuni infection. Unlike the outcome of severe intestinal diseases when infecting to human or mice [37, 38], C. jejuni infection in chickens neither induced diseases nor reduced the bird average daily gain compared to uninfected birds (Fig 2A). Remarkably, DCA promoted bird average daily gain by 36.3% at 14 days of age, compared to control birds. The average daily gain of birds fed DCA diets posed 12.7% increase compared to infected control birds at 28 days of age (Fig 2B). Interestingly, C. jejuni colonization increased chicken body weight gain during d 14–28. These findings suggest that DCA promotes bird average daily gain and C. jejuni colonization doesn’t induce adverse effect on bird health and growth.
Fig 2. DCA promotes broiler chicken growth performance.
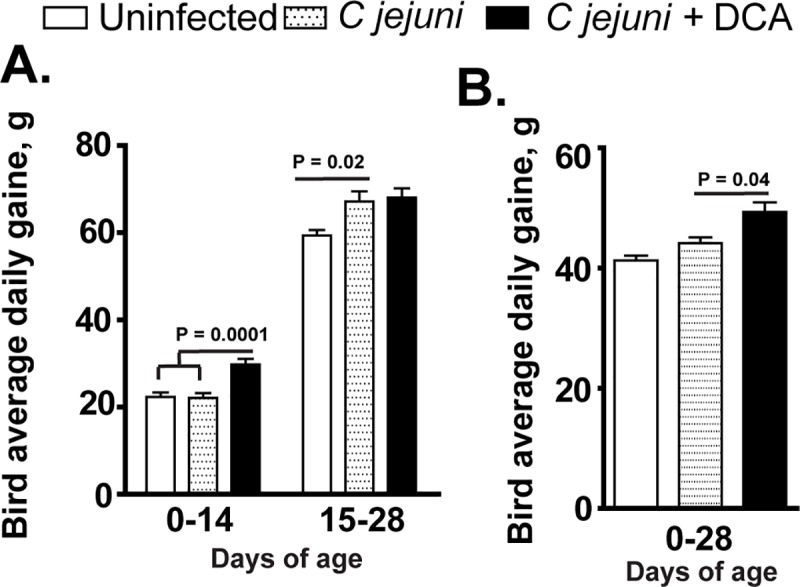
Cohorts of 13 birds were fed DCA diet and infected with C. jejuni as described in Fig 1. (A) Periodic bird average daily gain. (B) Accumulative bird average daily gain at 28 days of age. All graphs depict mean ± SEM. Significant if P<0.05. Results are representative of 2 independent experiments.
DCA fails to inhibit C. jejuni 81–176 in vitro growth
Since DCA prevented C. jejuni colonization in chickens, we then examine whether DCA would directly inhibit C. jejuni 81–176 growth. The bacterium was inoculated in Campylobacter Enrichment Broth in the presence of different concentrations (0, 5, and 25 mM) of bile acids DCA, CA, and TCA and cultured at 42°C for 24 hours under microaerobic condition. C. jejuni growth in 5 and 25 mM DCA broth induced gel formation and the OD600 reading was not accurate, therefore plate counting was used. Notably, primary bile acid CA at 25 mM reduced C. jejuni 81–176 growth (Fig 3A). Interestingly, conjugated primary bile acid TCA and secondary bile acid DCA didn’t reduce C. jejuni 81–176 in vitro growth.
Fig 3. Bile acids fail to consistently inhibit C. jejuni in vitro growth.
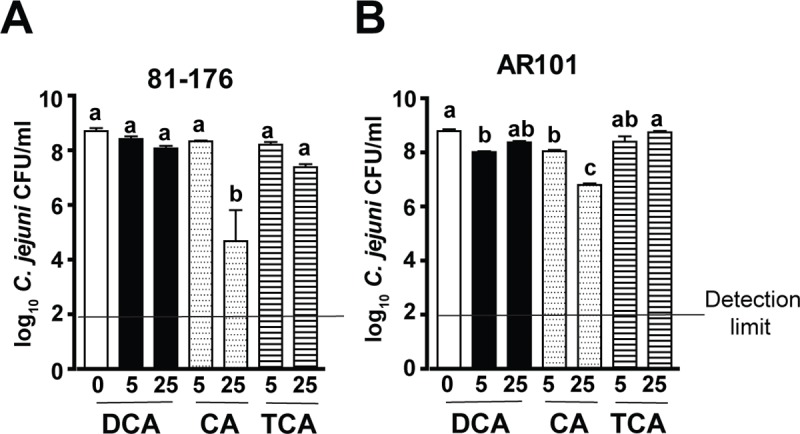
C. jejuni 81–176 or AR101 at 103 CFU was inoculated into 5 ml Campylobacter broth under microaerophilic condition and cultured at 42°C for 24 hours. The broth was supplemented with various concentrations (0, 5, or 25 mM) of secondary bile acid DCA, primary bile acid CA, or conjugated primary bile acid TCA. (A) Bile acids on 81–176 growth. (B) Bile acids on AR101 growth. Different letters represent significance (P<0.05) between treatments. Results are representative of 3 independent experiments.
Our long-term goal is to use microbiome to reduce C. jejuni transmission in chickens and to attenuate transmitted campylobacteriosis using mouse Il10-/- model as reported before [34, 38]. Hence, C. jejuni colonized and transmitted in chickens with microbiome treatment is required to be isolated for subsequently infecting Il10-/- mice. It was problematic when C. jejuni 81–176 couldn’t be isolated from DCA treated chickens as showed in Fig 1. Different C. jejuni strains show variable colonization ability in chickens [39]. Based on this knowledge, we then assessed the impact of bile acids on growth of C. jejuni strain AR101 which was isolated from chickens at Dr. Billy Hargis’s laboratory. Notably, 5 mM DCA, 5 mM CA, and 25 mM CA significantly reduced C. jejuni AR101 growth but not TCA (Fig 3B). Because DCA didn’t consistently reduce C. jejuni 81–176 or AR101 in vitro growth, it was concluded that DCA against C. jejuni chicken colonization results from factors other than DCA direct inhibition on C. jejuni growth.
Other secondary bile acids don’t reduce C. jejuni AR101 colonization in chickens
Secondary bile acids DCA, LCA, and UDCA were frequently used in assaying their ability against bacterial pathogens including Clostridium difficile [19] or C. jejuni [34] in mice and in vitro. Since DCA reduced C. jejuni 81–176 colonization in chickens, it was possible that other secondary bile acids also decreased the bacterial chicken colonization. To assess this possibility, birds were fed diets supplemented with 0 or 1.5 g/kg DCA, LCA, or UDCA from d 0. Because the chicken ages of C. jejuni AR101 infection didn’t influence its chicken colonization (S1 Fig), birds were infected once with C. jejuni AR101 at d10 instead of d14 to have longer time to interact with the bile acids. Consistent with previous findings in Fig 1, DCA reduced 91% of C. jejuni AR101 colonization compared to infected control birds (2.06x106 vs. 2.39x107 CFU/g), while LCA and UDCA failed to decrease AR101 chicken colonization (2.05x107 and 1.40x107 CFU/g, respectively) (Fig 4). Together, these data suggest that only secondary bile acid DCA effectively reduces C. jejuni chicken colonization.
Fig 4. LCA and UDCA fail to prevent C. jejuni colonization in chickens.
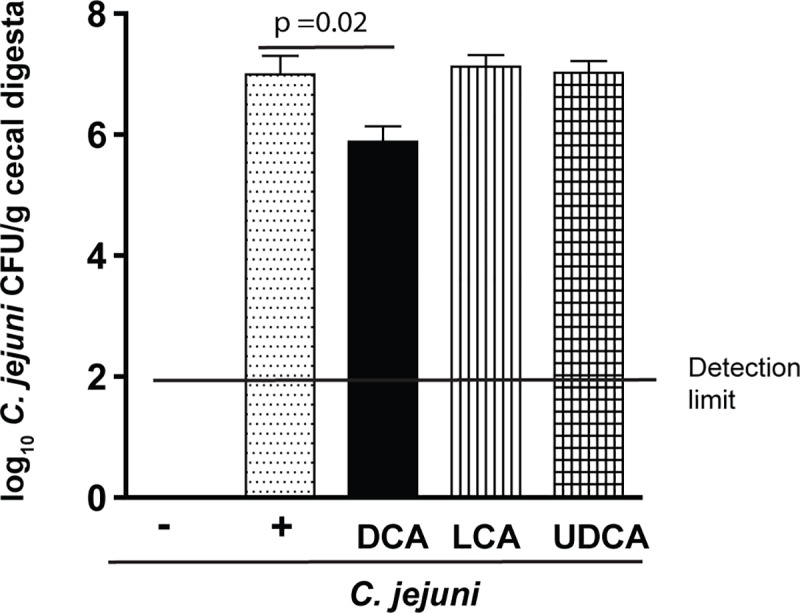
Cohorts of 18 chicks were fed diets supplemented with 0 or 1.5 g/kg of DCA, UDCA, or LCA. and were infected with C. jejuni AR 101 at day 10 of age. All birds were sacrificed and cecal digesta were collected at 28 days of age, serially diluted, and cultured on C. jejuni selective medium at 42°C. C. jejuni was counted after 48 hours. All graphs depict mean ± SEM. Significant if P<0.05. Results are representative of 2 independent experiments.
DCA modulates bird cecal microbiota
Relative abundance of phylum Firmicutes is dramatically expanded from 54 to 93% in intestine of rat fed primary bile acid CA [30]. Since DCA didn’t inhibit C. jejuni in vitro growth, we then reasoned that DCA might modify microbiota against C. jejuni chicken colonization. To examine this hypothesis, cecal digesta from birds infected with C. jejuni 81–176 and fed DCA (Fig 1) were used to exact DNA. We selected these samples because C. jejuni wasn’t detected in the samples and the samples would be used for following experiment of isolating transplantation microbiota. Phylum specific primers were used to analyze the microbiota composition. Interestingly, C. jejuni infection didn’t change microbial composition in microbiota of infected vs. uninfected birds (Fig 5). Remarkably, DCA reduced phylum Firmicutes (82.7 vs. 98.8%) compared to infected control birds, while increased Bacteroidetes (16.9 vs. 0.8%). These results indicate that DCA is able to alter the chicken gut microbiota.
Fig 5. DCA modulates cecal microbiota.
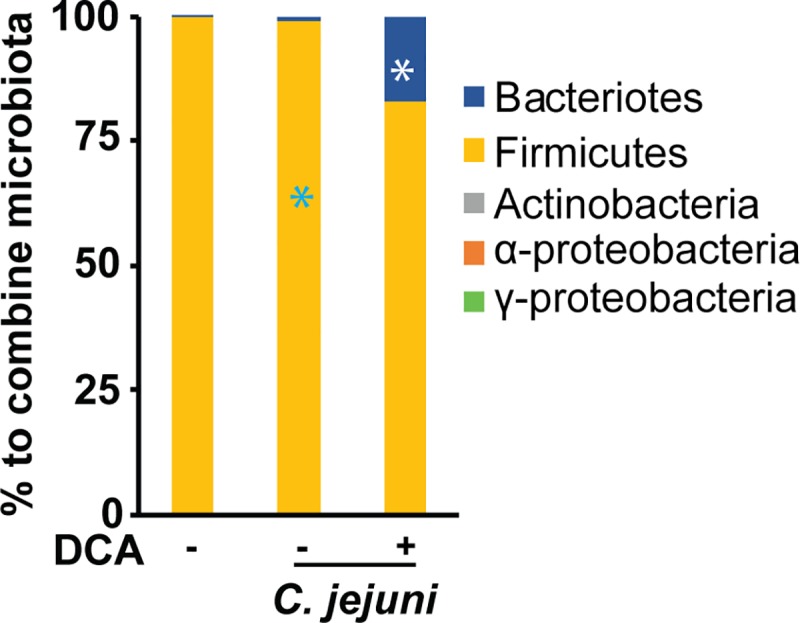
Cohorts of 13 chicks were fed 0 or 1.5 g/kg DCA diets and infected with C. jejuni 81–176 as described in Fig 1. Cecal digesta samples were collected and DNA was isolated. Real time PCR was performed to calculate bacterial composition at phylum level. All graphs depict mean ± SEM. *, P<0.05. Results are representative of 2 independent experiments.
DCA modulated-anaerobes attenuate C. jejuni AR101 chicken colonization
Since DCA-modulated chicken cecal microbiota, we then hypothesized that the altered microbiota contributed to the reduction of C. jejuni in chickens. In our previous studies, we found that only anaerobes prevent C. jejuni-induced intestinal inflammation in mice [34]. Based on this knowledge, cecal digesta from the birds fed DCA and infected with C. jejuni 81–176 in Fig 1 were used to culture bacteria on BHI plates under aerobic or anaerobic conditions. The resulted bacteria were labeled as DCA-modulated aerobes (DCA-Aero) or DCA-modulated anaerobes (DCA-Anaero), respectively. To functionally dissect the role of these newly isolated microbiota, birds were gavaged once with DCA-Aero or DCA-Anaero microbiota at 0 day of age. The birds were then infected with 109 CFU/bird C. jejuni AR101 at 10 days of age. Consistent with previous observations, C. jejuni colonization in chicken ceca reached a plateau at a level of 2.80x107 CFU/g cecal digesta at 21 days of age (Fig 6A), or 11 days post infection. Importantly, DCA-Anaero significantly attenuated 93% of C. jejuni cecal colonization at 28 days of age compared to infected control birds (1.79x106 vs. 2.52x107 CFU/bird), while DCA-Aero only reduced 34.46% of C. jejuni (1.65x107 vs. 2.52x107) colonization. To examine if the transplanted microbiota colonized the chickens, cecal samples from the birds were collected. DNA extracted was performed phylum level PCR and the bacterial DNA was also planned for 16S rDNA sequencing. Consistent with the inoculum DCA microbiota, the microbiota of DCA-Anaero birds showed increased Bacteroidetes (23.6 vs. 3.8%) but reduced Firmicutes (76.3 vs. 95.8%) compared to infected control birds (Fig 6B), while DCA-Aero birds shared similar microbiota composition to infected control birds. These data supported the notion of our successful DCA microbiota collection, inoculation, and chicken colonization. Taken together, these findings suggest that DCA modulates microbiota and the modulated anaerobes contribute to the reduction of C. jejuni colonization in chickens.
Fig 6. DCA-modulated microbiota attenuates C. jejuni colonization in chickens.

Cohorts of 18 broiler chickens were orally transplanted with 108 CFU/bird DCA-Aero or DCA-Anaero at d 0. The birds were then infected with 109 CFU/bird C. jejuni AR 101 at 10 days of age. Cecal digesta were collected at 21 and 28 days of age, serially diluted, and cultured on C. jejuni selective medium at 42°C. C. jejuni was counted after 48 hours. (A) C. jejuni cecal colonization. (B) Cecal microbiota composition at d28 at phylum level. *, P<0.05 between DCA-Anaero and infected control birds. All graphs depict mean ± SEM.; significant if P<0.05. Results are representative of 3 independent experiments.
Discussion
Although C. jejuni is one of the prevalent foodborne pathogens in developed countries, a paucity of information is available regarding reducing the pathogen in the main food animal source of chickens. Moreover, the microbiota and cellular events implicated in host resistance/susceptibility to C. jejuni infection remain elusive [40, 41]. Here we report that microbial metabolic product DCA prevented colonization of both human clinical isolate C. jejuni 81–176 and chicken isolate AR101 in chickens. Interestingly, bile acids of DCA, TCA, or low concentration CA failed to reduce C. jejuni in vitro growth. Furthermore, neither bird ages of C. jejuni infection at d 5, 10 or 14, nor other bile acids of LCA or UDCA influenced the bacterial chicken colonization levels. Mechanistic studies revealed that DCA modified chicken cecal microbiota with increased phylum Bacteroidetes and reduced Firmicutes. Importantly, DCA-modulated anaerobes prevented C. jejuni chicken colonization. Altogether, these findings identified a novel mechanism that DCA shapes microbiota composition against C. jejuni colonization in chickens, suggesting a bidirectional interaction between microbiota and microbial metabolites.
A remarkable observation from our study is that DCA but not LCA or UDCA prevented C. jejuni colonization in chickens. The logic reasoning would be that the reduction would be through DCA directly impairing C. jejuni growth. DCA at 1.2 mM inhibits C. jejuni 81–176 in vitro growth after 12 hour incubation [42] but Lin and colleague found that the MICs of DCA and CA for C. jejuni 81–176 are 24 and 14 mM, respectively [43]. Interestingly, DCA at 48 mM fails to reduce C. jejuni 43431 growth at 6, 22, 25 and 30 hours of incubation but not at 16 hours [44]. Consistent with the latter reports, we found that conjugated primary bile acid TCA or secondary bile acid DCA at as high as 25 mM didn’t significantly inhibit C. jejuni 81–176 or AR101 in vitro growth. Consistent with Lin’s report [43], CA at 25 mM significantly reduced C. jejuni in vitro growth, suggesting a potential to manipulate this bile acid against C. jejuni chicken colonization. In animals, DCA reduces C. jejuni induced intestinal inflammation in ex-Germ Free mice without altering C. jejuni colonization level in colon [34]. It was obvious that DCA reduced chicken colonization of C. jejuni 81–176 greater than AR101. This could result from human clinical isolate 81–176 passed for many generations in our lab, while AR101 was isolated from chickens and passed less in lab. It could be that different C. jejuni strains have different colonization ability in chickens [39].Together, the knowledge and data suggest that DCA reduces C. jejuni colonization possibly through mechanisms other than directly inhibiting the bacterial growth.
Comprehensive database analysis showed that chicken microbiota at phylum level is mainly comprised of 13 phyla including Firmicutes (70%), Bacteroidetes (12.3%), Proteobacteria (9.3%), and other small proportion of Actinobacteria, Cyanobacteria, Spirochaetes, Synergisteles, Fusobacteria, Tenericutes, and Verrucomicrobia [45]. The first three phyla account for more than 91% of the 13 phyla, and we then measured five phyla using the reported real time PCR primers [36]. The lack of the rest 8 phyla of small proportion could influence the interpretation of our microbiota results. Our phylum level analysis of microbiota has found that birds fed DCA were colonized with reduced Firmicutes (82.7 vs. 98.8%) and increased Bacteroidetes (16.9 vs. 0.8%), which is associated with less C. jejuni colonization compared to control infected birds. Notably, this finding is consistent with a field survey report that birds from the farm with the highest Campylobacter counts are associated with the highest percentage of Firmicutes and the lowest percentage of Bacteroidetes, although microbiota composition is highly variable between inter- or intra-farms [46]. Interestingly, the microbiota in mice fed CA expands phylum Firmicutes (54 to 99%), class Clostridia (70 to 39%), and genus Blautia (8.3 to 55–62%), in the expense of phylum Bacteroidetes (30 to 0.39%) [30]. The levels of bile acids are associated with microbial community dynamics in the gut [27]. Bile acids directly inhibit gut microbes [28] and ileal bacterial overgrowth through FXR-induced antimicrobial peptides [29]. However, it remains elusive how various bile acid species differentially influence intestinal microbiota composition. In the future, we will investigate the microbiota composition using 16S rDNA sequencing in birds fed DCA, LCA, and UDCA.
Based on the results of DCA altering microbiota, we hypothesized that the microbiota played roles on protecting against C. jejuni chicken colonization. To functionally dissect the protection of the DCA-modulated microbiota, microbiota culture and chicken microbiota transplantation were performed. Indeed, DCA modulated anaerobes reduced C. jejuni chicken colonization, while DCA modulated aerobes failed to do so. Phylum level microbiota composition analysis by real time PCR strongly support the successful microbiota collection, inoculation and chicken colonization. Microbiota prevents against C. jejuni chicken colonization because C. jejuni colonizes at higher level in germ free or antibiotics pre-treated chickens compared to conventional birds [47]. Specific members of microbiota play an important role against C. jejuni induced diseases. Microbiota diversity and the relative abundances of genera Dorea and Coprococcus in family Lachnospiraceae were higher in healthy travelers compared to individuals suffered with campylobacteriosis [41]. Mouse microbiota with higher level of genera Clostridium XI, Bifidobacterium, and Lactobacillus is associated with resistance to C. jejuni induced colitis [34]. The three genera bacteria metabolize conjugated bile acids into secondary forms [21] which prevent campylobacteriosis in mice [34]. Interestingly, probiotics Bifidobacterium longum PCB133 and a xylo-oligosaccharide fail to reduce C. jejuni chicken colonization using plate enumeration [48]. We are processing the cecal samples to extract bacterial DNA to run 16S rDNA sequencing and we expect that we will find specific bacteria in DCA-modulated anaerobes responsible for protection against C. jejuni chicken colonization.
Finally, we found that C. jejuni colonization levels was independent on the ages of birds infected. It remains controversial whether chicken ages of inflection play any role on C. jejuni chicken colonization. C. jejuni is detected in 40% of broiler chicken flocks at 4 weeks of age and in 90% of flocks at 7 weeks of age [49]. This infection pattern could be attributed to age-related resistance or the necessary timing for C. jejuni transmission within house. Birds from parent breeders colonized with high level C. jejuni strain 99/308 are resistant to 99/308 only at 8 days of age but not at 1 or 22 days of age [50], suggesting specific and limited protection from the breeders. However, Han and colleagues showed that C. jejuni strains Lior6 and 0097 colonize less in birds of d 22 compared to d 1, after 14 days post infection [43]. Because bird microbiota starts to assemble after hatch, the older birds in some farms might develop microbiota resistant to C. jejuni, as in Han and colleagues’ report [43]. Similarly, germ free mice transferred to SPF housing for 14 days resist against C. jejuni induced colitis [34]. In those research reports showing no age difference on C. jejuni bird colonization, the resistant microbiota assembly in the birds might be blocked by strict biosecurity measurements or no available resistant microbiota in the environment.
In conclusion, the microbiota metabolite secondary bile acid DCA decreases C. jejuni counts and modulates microbiota composition in the chicken intestine. At mechanistic level, DCA-modulated anaerobic microbiota may be responsible for protecting against C. jejuni colonization in chickens. These findings identified a novel mechanism that DCA shapes microbiota composition against C. jejuni colonization in chickens, suggesting a bidirectional interaction between microbiota and microbial metabolites. Simultaneously reconstituting both microbiota and microbial metabolites may render better therapeutic effect against campylobacteriosis or the pathogen colonization.
Supporting information
Cohorts of 18 one-day-old broiler chicks were fed basal diet and orally gavaged with 109 CFU/bird C. jejuni AR 101 at 5, 10, or 14 days of age. At d 28, birds were humanely sacrificed and cecal samples were collected. Cecal digesta samples were serially diluted and cultured on Campylobacter selective media. Colonies were enumerated and C. jejuni colonization levels were calculated. All graphs depict mean ± SEM. Results are representative of 2 independent experiments.
(TIF)
Abbreviations
- DCA
Deoxycholic acid
- CFU
colony forming unit
- DCA-Anaero
DCA modulated anaerobes
- DCA-Aero
DCA modulated aerobes
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This research was supported grants of Arkansas Biosciences Institute, USDA National Institute of Food and Agriculture (NIFA) Hatch project 1012366, USDA NIFA Hatch/Multi State project 1018699, USDA NIFA project 2018-06686 to X. Sun. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Pham NT, Thongprachum A, Tran DN, Nishimura S, Shimizu-Onda Y, Trinh QD, et al. Antibiotic Resistance of Campylobacter jejuni and C. coli Isolated from Children with Diarrhea in Thailand and Japan. Jpn J Infect Dis. 2016;69(1):77–9. 10.7883/yoken.JJID.2014.582 . [DOI] [PubMed] [Google Scholar]
- 2.Bae J, Oh E, Jeon B. Enhanced transmission of antibiotic resistance in Campylobacter jejuni biofilms by natural transformation. Antimicrob Agents Chemother. 2014;58(12):7573–5. 10.1128/AAC.04066-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hou FQ, Sun XT, Wang GQ. Clinical manifestations of Campylobacter jejuni infection in adolescents and adults, and change in antibiotic resistance of the pathogen over the past 16 years. Scand J Infect Dis. 2012;44(6):439–43. 10.3109/00365548.2011.652163 . [DOI] [PubMed] [Google Scholar]
- 4.Szczepanska B, Kaminski P, Andrzejewska M, Spica D, Kartanas E, Ulrich W, et al. Prevalence, virulence, and antimicrobial resistance of Campylobacter jejuni and Campylobacter coli in white stork Ciconia ciconia in Poland. Foodborne Pathog Dis. 2015;12(1):24–31. 10.1089/fpd.2014.1793 . [DOI] [PubMed] [Google Scholar]
- 5.Zhao S, Mukherjee S, Chen Y, Li C, Young S, Warren M, et al. Novel gentamicin resistance genes in Campylobacter isolated from humans and retail meats in the USA. The Journal of antimicrobial chemotherapy. 2015;70(5):1314–21. 10.1093/jac/dkv001 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.CDC. Foodborne Diseases Active Surveillance Network (FoodNet) Fast: Centers for Disease Control and Prevention; https://wwwn.cdc.gov/foodnetfast/; 2018. [updated May 19, 2018; cited 2018 May 19]. Available from: https://wwwn.cdc.gov/foodnetfast/. [Google Scholar]
- 7.CDC. Campylobacter general inflammation 2014. Available from: http://www.cdc.gov/nczved/divisions/dfbmd/diseases/campylobacter/. [Google Scholar]
- 8.Ternhag A, Asikainen T, Giesecke J, Ekdahl K. A meta-analysis on the effects of antibiotic treatment on duration of symptoms caused by infection with Campylobacter species. Clin Infect Dis. 2007;44(5):696–700. Epub 2007/02/06. 10.1086/509924 [pii] 10.1086/509924. . [DOI] [PubMed] [Google Scholar]
- 9.CDC. Campylobacter: General Information. Available at http://www.cdc.gov/nczved/divisions/dfbmd/diseases/campylobacter/. Accessed October 19, 2010. 2010. [Google Scholar]
- 10.Neu HC. The crisis in antibiotic resistance. Science. 1992;257(5073):1064–73. Epub 1992/08/21. 10.1126/science.257.5073.1064 . [DOI] [PubMed] [Google Scholar]
- 11.Gibbens JC, Pascoe SJ, Evans SJ, Davies RH, Sayers AR. A trial of biosecurity as a means to control Campylobacter infection of broiler chickens. Preventive veterinary medicine. 2001;48(2):85–99. . [DOI] [PubMed] [Google Scholar]
- 12.Hodgins DC, Barjesteh N, St Paul M, Ma Z, Monteiro MA, Sharif S. Evaluation of a polysaccharide conjugate vaccine to reduce colonization by Campylobacter jejuni in broiler chickens. BMC Res Notes. 2015;8:204 10.1186/s13104-015-1203-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aguiar VF, Donoghue AM, Arsi K, Reyes-Herrera I, Metcalf JH, de los Santos FS, et al. Targeting motility properties of bacteria in the development of probiotic cultures against Campylobacter jejuni in broiler chickens. Foodborne Pathog Dis. 2013;10(5):435–41. 10.1089/fpd.2012.1302 . [DOI] [PubMed] [Google Scholar]
- 14.Kittler S, Fischer S, Abdulmawjood A, Glunder G, Klein G. Colonisation of a phage susceptible Campylobacter jejuni population in two phage positive broiler flocks. PLoS One. 2014;9(4):e94782 10.1371/journal.pone.0094782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hertwig S, Hammerl JA, Appel B, Alter T. Post-harvest application of lytic bacteriophages for biocontrol of foodborne pathogens and spoilage bacteria. Berl Munch Tierarztl Wochenschr. 2013;126(9–10):357–69. . [PubMed] [Google Scholar]
- 16.Rosenquist H, Nielsen NL, Sommer HM, Norrung B, Christensen BB. Quantitative risk assessment of human campylobacteriosis associated with thermophilic Campylobacter species in chickens. Int J Food Microbiol. 2003;83(1):87–103. . [DOI] [PubMed] [Google Scholar]
- 17.CDC. Morbidity and Mortality Weekly Report at https://wonder.cdc.gov/mmwr/mmwrmort.asp 1996–2016 [updated 2016 week 53]. Available from: https://wonder.cdc.gov/mmwr/mmwrmort.asp.
- 18.Silverman MS, Davis I, Pillai DR. Success of self-administered home fecal transplantation for chronic Clostridium difficile infection. Clin Gastroenterol Hepatol. 2010;8(5):471–3. 10.1016/j.cgh.2010.01.007 . [DOI] [PubMed] [Google Scholar]
- 19.Buffie CG, Bucci V, Stein RR, McKenney PT, Ling L, Gobourne A, et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature. 2015;517(7533):205–8. 10.1038/nature13828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Darkoh C, Lichtenberger LM, Ajami N, Dial EJ, Jiang ZD, DuPont HL. Bile acids improve the antimicrobial effect of rifaximin. Antimicrob Agents Chemother. 2010;54(9):3618–24. 10.1128/AAC.00161-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ridlon JM, Kang DJ, Hylemon PB. Bile salt biotransformations by human intestinal bacteria. Journal of lipid research. 2006;47(2):241–59. 10.1194/jlr.R500013-JLR200 . [DOI] [PubMed] [Google Scholar]
- 22.Chiang JY. Regulation of bile acid synthesis: pathways, nuclear receptors, and mechanisms. Journal of hepatology. 2004;40(3):539–51. 10.1016/j.jhep.2003.11.006 . [DOI] [PubMed] [Google Scholar]
- 23.Archer RH, Chong R, Maddox IS. Hydrolysis of bile acid conjugates by Clostridium bifermentans. European journal of applied microbiology and biotechnology. 1982;14(1):41–5. 10.1007/bf00508002 [DOI] [Google Scholar]
- 24.Gilliland SE, Speck ML. Deconjugation of bile acids by intestinal lactobacilli. Applied and environmental microbiology. 1977;33(1):15–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jones BV, Begley M, Hill C, Gahan CG, Marchesi JR. Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(36):13580–5. 10.1073/pnas.0804437105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vlasova AN, Chattha KS, Kandasamy S, Liu Z, Esseili M, Shao L, et al. Lactobacilli and bifidobacteria promote immune homeostasis by modulating innate immune responses to human rotavirus in neonatal gnotobiotic pigs. PLoS One. 2013;8(10):e76962 10.1371/journal.pone.0076962 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ridlon JM, Kang DJ, Hylemon PB, Bajaj JS. Bile acids and the gut microbiome. Current opinion in gastroenterology. 2014;30(3):332–8. 10.1097/MOG.0000000000000057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Begley M, Gahan CG, Hill C. The interaction between bacteria and bile. FEMS microbiology reviews. 2005;29(4):625–51. 10.1016/j.femsre.2004.09.003 . [DOI] [PubMed] [Google Scholar]
- 29.Inagaki T, Moschetta A, Lee YK, Peng L, Zhao G, Downes M, et al. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(10):3920–5. 10.1073/pnas.0509592103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Islam KB, Fukiya S, Hagio M, Fujii N, Ishizuka S, Ooka T, et al. Bile acid is a host factor that regulates the composition of the cecal microbiota in rats. Gastroenterology. 2011;141(5):1773–81. 10.1053/j.gastro.2011.07.046 . [DOI] [PubMed] [Google Scholar]
- 31.Ma H, Patti ME. Bile acids, obesity, and the metabolic syndrome. Best practice & research Clinical gastroenterology. 2014;28(4):573–83. 10.1016/j.bpg.2014.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bernstein C, Holubec H, Bhattacharyya AK, Nguyen H, Payne CM, Zaitlin B, et al. Carcinogenicity of deoxycholate, a secondary bile acid. Archives of toxicology. 2011;85(8):863–71. 10.1007/s00204-011-0648-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015;161(2):264–76. 10.1016/j.cell.2015.02.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sun X, Winglee K, Gharaibeh RZ, Gauthier J, He Z, Tripathi P, et al. Microbiota-Derived Metabolic Factors Reduce Campylobacteriosis in Mice. Gastroenterology. 2018;154(6):1751–63 e2. 10.1053/j.gastro.2018.01.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Galarza-Seeber R, Latorre JD, Bielke LR, Kuttappan VA, Wolfenden AD, Hernandez-Velasco X, et al. Leaky Gut and Mycotoxins: Aflatoxin B1 Does Not Increase Gut Permeability in Broiler Chickens. Frontiers in veterinary science. 2016;3:10 10.3389/fvets.2016.00010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bacchetti De Gregoris T, Aldred N, Clare AS, Burgess JG. Improvement of phylum- and class-specific primers for real-time PCR quantification of bacterial taxa. Journal of microbiological methods. 2011;86(3):351–6. 10.1016/j.mimet.2011.06.010 . [DOI] [PubMed] [Google Scholar]
- 37.van Spreeuwel JP, Duursma GC, Meijer CJ, Bax R, Rosekrans PC, Lindeman J. Campylobacter colitis: histological immunohistochemical and ultrastructural findings. Gut. 1985;26(9):945–51. Epub 1985/09/01. 10.1136/gut.26.9.945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lippert E, Karrasch T, Sun X, Allard B, Herfarth HH, Threadgill D, et al. Gnotobiotic IL-10; NF-kappaB mice develop rapid and severe colitis following Campylobacter jejuni infection. PLoS One. 2009;4(10):e7413 10.1371/journal.pone.0007413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pielsticker C, Glunder G, Rautenschlein S. Colonization properties of Campylobacter jejuni in chickens. Eur J Microbiol Immunol (Bp). 2012;2(1):61–5. 10.1556/EuJMI.2.2012.1.9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Luethy PM, Huynh S, Ribardo DA, Winter SE, Parker CT, Hendrixson DR. Microbiota-Derived Short-Chain Fatty Acids Modulate Expression of Campylobacter jejuni Determinants Required for Commensalism and Virulence. mBio. 2017;8(3). 10.1128/mBio.00407-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kampmann C, Dicksved J, Engstrand L, Rautelin H. Composition of human faecal microbiota in resistance to Campylobacter infection. Clinical microbiology and infection: the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 2016;22(1):61 e1– e8. 10.1016/j.cmi.2015.09.004 . [DOI] [PubMed] [Google Scholar]
- 42.Negretti NM, Gourley CR, Clair G, Adkins JN, Konkel ME. The food-borne pathogen Campylobacter jejuni responds to the bile salt deoxycholate with countermeasures to reactive oxygen species. Sci Rep. 2017;7(1):15455 10.1038/s41598-017-15379-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Han Z, Pielsticker C, Gerzova L, Rychlik I, Rautenschlein S. The influence of age on Campylobacter jejuni infection in chicken. Dev Comp Immunol. 2016;62:58–71. 10.1016/j.dci.2016.04.020 . [DOI] [PubMed] [Google Scholar]
- 44.Lertpiriyapong K, Gamazon ER, Feng Y, Park DS, Pang J, Botka G, et al. Campylobacter jejuni type VI secretion system: roles in adaptation to deoxycholic acid, host cell adherence, invasion, and in vivo colonization. PLoS One. 2012;7(8):e42842 10.1371/journal.pone.0042842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wei S, Morrison M, Yu Z. Bacterial census of poultry intestinal microbiome. Poult Sci. 2013;92(3):671–83. 10.3382/ps.2012-02822 . [DOI] [PubMed] [Google Scholar]
- 46.Sakaridis I, Ellis RJ, Cawthraw SA, van Vliet AHM, Stekel DJ, Penell J, et al. Investigating the Association Between the Caecal Microbiomes of Broilers and Campylobacter Burden. Front Microbiol. 2018;9:927 10.3389/fmicb.2018.00927 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Han Z, Willer T, Li L, Pielsticker C, Rychlik I, Velge P, et al. Influence of the Gut Microbiota Composition on Campylobacter jejuni Colonization in Chickens. Infect Immun. 2017;85(11). 10.1128/IAI.00380-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Baffoni L, Gaggia F, Garofolo G, Di Serafino G, Buglione E, Di Giannatale E, et al. Evidence of Campylobacter jejuni reduction in broilers with early synbiotic administration. Int J Food Microbiol. 2017;251:41–7. 10.1016/j.ijfoodmicro.2017.04.001 . [DOI] [PubMed] [Google Scholar]
- 49.Evans SJ, Sayers AR. A longitudinal study of campylobacter infection of broiler flocks in Great Britain. Preventive veterinary medicine. 2000;46(3):209–23. . [DOI] [PubMed] [Google Scholar]
- 50.Cawthraw SA, Newell DG. Investigation of the presence and protective effects of maternal antibodies against Campylobacter jejuni in chickens. Avian Dis. 2010;54(1):86–93. 10.1637/9004-072709-Reg.1 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Cohorts of 18 one-day-old broiler chicks were fed basal diet and orally gavaged with 109 CFU/bird C. jejuni AR 101 at 5, 10, or 14 days of age. At d 28, birds were humanely sacrificed and cecal samples were collected. Cecal digesta samples were serially diluted and cultured on Campylobacter selective media. Colonies were enumerated and C. jejuni colonization levels were calculated. All graphs depict mean ± SEM. Results are representative of 2 independent experiments.
(TIF)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.