

Abstract
Coccidiosis is a parasitic disease of a wide variety of animals caused by coccidian protozoa. The coccidia are responsible for major economic losses of the livestock industry. For example, the annual cost due to coccidiosis to the global poultry industry has been estimated to exceed US$ 3 billion annually. Currently available drugs for the control of this disease are either polyether ionophorous antibiotics that are derived from fermentation products, or synthetic compounds, produced by chemical synthesis. Unfortunately, no new drugs in either category have been approved for use for decades. Resistance has been documented for all those of the drugs currently employed and therefore the discovery of novel drugs with unique modes of action is imperative if chemotherapy is to remain the principal means to control this disease. This chapter aims to give an overview of the efficacy and mode of action of the current compounds used to control coccidiosis in livestock and provides a brief outlook of research needs for the future.
Keywords: Coccidiosis, Anticoccidials, Livestock, Ionophores, Chemicals, Mode of action, Resistance
Introduction
Coccidiosis is an infectious disease of the intestinal tract of wild and domestic animals caused by different protozoa. These include but are not limited to Isospora, Neospora, Cryptosporidium, and Eimeria of the phylum Apicomplexa. Those parasites are widespread, especially where intensive production systems are used to raise livestock. They cause mortality, poor growth, and impaired performance. In addition to mortality and reduced growth, coccidiosis also affects meat yield and quality and increases susceptibility to ancillary infections (Nagi and Mathey 1972). In the chicken, at least seven Eimeria species are recognized that parasitize different regions of the intestine (Shirley et al. 1986); many species are recognized in ruminants. The parasites are transmitted from one host individual to others via the feces which shed the transmission stage of the life cycle (the oocyst) into the environment. Infection results from ingestion of sporulated oocysts in the litter. The global poultry industry is considered most affected by coccidiosis as it causes this industry annual losses that have been estimated to exceed US$ 3 billion per year (Williams 1999; Dalloul and Lillehoj 2006). Poultry production is projected to more than double by the year 2050 (Alexandratos and Bruinsma 2012) and control of coccidiosis will be essential if poultry and livestock meat is to fulfill the increasing need for protein by the growing world population (Kart and Bilgili 2008). Control of the disease by the use of drugs is indispensable if we are to achieve sustainable poultry production. Animal welfare is compromised by coccidial infections, and food safety is an important consideration (Kadykalo et al. 2018).
In addition to control programs based upon chemotherapy or vaccination, satisfactory control of coccidiosis in poultry requires strict attention to hygiene and sanitation, and biosecurity measures that limit human access to poultry facilities (Chapman 2018). Adequate ventilation and leak-free watering systems are important to reduce excessive moisture because wet litter aids sporulation of the infective stage of the life cycle (the oocyst). Nevertheless, despite such measures, eradication has not proved possible and the parasites persist in poultry flocks (Chapman et al. 2016). Preventative treatment may employ pharmaceutical ingredients in medicated food or drinking water, or immunization involving the use of live attenuated or non-attenuated vaccines (Klotz et al. 2005). By these means, it is estimated that most broiler chickens produced worldwide receive treatment with drugs or are vaccinated (Chapman et al. 2002). Prophylaxis has been the preferred method for the control of coccidiosis in poultry because treatment once clinical signs become apparent is often too late to prevent the pathological consequences of infection (Chapman 2009).
The concept of coccidiosis prevention in chickens by inclusion of drugs in the feed (prophylaxis) was first described in 1948 and involved the use of sulfaquinoxaline, the first feed additive for poultry (Grumbles et al. 1948; reviewed by Chapman 2009). In the years that followed, many other drugs were introduced, and until the introduction of ionophores in the 1970s, chemoprophylactic control of coccidiosis was based on the use of such synthetic anticoccidials (Ryley and Betts 1973). No new chemicals have been introduced for decades, and resistance has been documented for all the drugs approved for use in chickens (Chapman 1997), although the onset of resistance can be slowed by using rotation programs with different chemicals and/or ionophores (Chapman et al. 2010). Nevertheless, resistance to the available chemicals and ionophores has become widespread (Peek and Landman 2011). Drugs with novel molecular modes of action, and hence unprecedented targets, will be necessary if control of coccidiosis by chemotherapy is to be achievable in the future (Kinnaird et al. 2004; Scribner et al. 2009). Very little effort to discover new drugs has been undertaken in recent years, but this may change with the advent of genomics technology (Chapman et al. 2013). Examples of the successful application of novel drug discovery could be shown for other protozoa that are relevant for the animal health industry, e.g., for the pig parasite Cystoiospora suis (Shrestha et al. 2017a).
In this chapter, we aim to give an overview of the efficacy and mode of action of the current compounds used to control coccidiosis in livestock and provide a brief outlook of research needs for the future. Previous reviews of this subject include those by Chapman (1997) and McDougald (1982).
Drug categories
Anticoccidial drugs belong to one of two categories (Chapman 1997; Allen and Fetterer 2002):
Polyether antibiotics or ionophores, which are produced by the fermentation of Streptomyces spp. or Actinomadura spp. These drugs disrupt ion gradients across the cell membrane of the parasite:
Monovalent ionophores (monensin, narasin, salinomycin)
Monovalent glycosidic ionophores (maduramicin, semduramicin)
Divalent ionophore (lasalocid)
-
2.
Synthetic compounds popularly known as “chemicals”, produced by chemical synthesis, often with a specific mode of action:
Inhibition of parasite mitochondrial respiration (decoquinate, clopidol)
Inhibition of the folic acid pathway (sulfonamides)
Competitive inhibition of thiamine uptake (amprolium)
Unknown mode of action (e.g., diclazuril, halofuginone, nicarbazin, robenidine)
Combination products, consisting of either a synthetic compound and ionophore (e.g., nicarbazin/narasin — Maxiban®, Elanco) or two synthetic compounds (clopidol/methyl benzoquate — Lerbek®, Impextraco NV), are also available. Arsenical drugs such as roxarsone that has some anticoccidial efficacy, arsanilic acid, carbarsone, and combinations thereof have been discontinued in many countries since 2015, based on scientific reports that indicated organic arsenic could transform into inorganic, highly toxic arsenic (Nachman et al. 2013; Huang et al. 2014).
Ionophores
For many years, ionophores have been the principal choice to control coccidiosis because resistance develops slowly to them and because they do not completely suppress parasite development, thus allowing the development of immunity in the host after first exposure (Chapman 1999a, b; Chapman et al. 2010; Jeffers 1989). They are characterized by multiple tetrahydrofuran rings that are connected in the form of spiroketal moieties (Riddell 2002) and are effective against the asexual and sexual life cycle stages of coccidia, disturbing the normal transport of ions across surface membranes of sporozoites or early trophozoites (Augustine et al. 1992; Smith and Galloway 1983; Smith and Strout 1980; Smith et al. 1981). Ionophores are only used in livestock and are not employed for any purpose in human medicine. They are not active against most foodborne bacteria of poultry, e.g., Escherichia coli, Salmonella spp. and Campylobacter spp., and are not, therefore, included in the WHO list of medically important antimicrobials. Their use is not an issue for public health (Tang et al. 2017; WHO 2017).
These drugs have a rather narrow safety margin (Dowling 1992), and most are incompatible with several therapeutic antibiotics. Among those are tiamulin (Umemura et al. 1984a; Islam et al. 2009), chloramphenicol, erythromycin, oleandromycin (Umemura et al. 1984b; Mazlum et al. 1985; Mazlum and Pradella 1986; Perelman et al. 1986; Broz and Frigg 1987), and certain sulfonamides, leading to intoxication manifested by severe temporary clinical symptoms (Dowling 1992; Schuhmacher et al. 2006). In addition, ionophores are also incompatible with some antioxidants (Laczay 1988; Laczay et.al. 1988, 1989; Umemura et al. 1984b; Dowling 1992; Peek and Landman 2011).
Monovalent ionophores can form lipid-soluble complexes with sodium and potassium cations, whereas divalent ionophores can bind calcium and magnesium cations only. Polyether ionophores arrest the development of sporozoites by increasing the concentration of intracellular Na+ ions. In addition, they increase the activity of Na+/K+/ATPase (Wang et al. 2006) and affect merozoites by causing the cell membrane to burst (Mehlhorn et al. 1983). Toxic effects in horse, cattle, dogs, cats, rats, and avian species are thought to be mediated by disrupting ion gradients of cell membranes, leading to mitochondrial damage, and thus depletion of cellular energy. Well-known toxic effects are cardiac toxicity and muscle degeneration, and neuropathy, the latter one being manifested by myelin degeneration and ataxia (Chapman 2018; Kart and Bilgili 2008).
Monovalent ionophores
Monensin
In 1967, the structure of monensic acid (Fig. 1e), a fermentation product of Streptomyces cinnamonensis, was first described and the compound was reported to have a broad-spectrum effect against Eimeria (Agtarap et al. 1967). It forms lipid-soluble complexes with sodium and potassium cations, leading to increased permeability of the membrane for these ions. Monensin is able to transport sodium ions through membranes in both electrogenic and electroneutral manner (Mollenhauer et al. 1990). Horses are particularly susceptible to monensin poisoning (Matsuoka et al. 1996). Accidental deadly poisoning of horses with monensin has been published (Doonan et al. 1989; Bezerra et al. 1999; Bila et al. 2001).
Fig. 1.
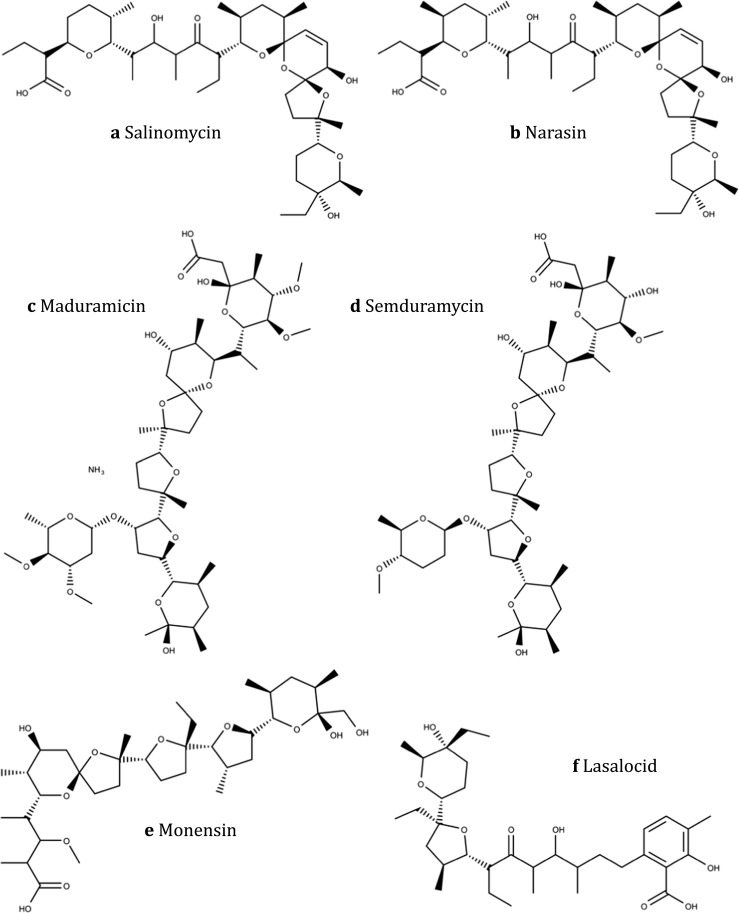
Ionophores used as anticoccidials. While salinomycin, narasin, maduramicin, semduramicin, and monensin (a–e) belong to the monovalent ionophores, lasalocid (f) is a divalent ionphore
Salinomycin
Salinomycin (Fig. 1a) was isolated from Streptomyces albus. It exhibits not only activity against Eimeria of poultry but also against gram-positive bacteria including mycobacteria and some filamentous fungi (Miyazaki et al. 1974). Salinomycin is an ionophore with strict selectivity for alkali ions and a strong preference for potassium, interfering with transmembrane potassium potential and promoting the efflux of K+ ions from mitochondria and cytoplasm. Recently, it has been shown to kill human cancer stem cells and to inhibit breast cancer growth and metastasis in mice (Naujokat et al. 2010). Salinomycin is the least toxic of all the ionophores (Oehme and Pickrell 1999).
Narasin
Narasin (Fig. 1b) is a polyether antibiotic obtained from Streptomyces aureofaciens (Jeffers et al. 1988). It is a derivative of salinomycin having an additional methyl group, therefore alternatively called (4S)-4-methyl salinomycin. When combining different ionophores with nicarbazin, Challey and Jeffers (1973) found that combinations of nicarbazin and narasin had synergistic activity. A combination product containing both active pharmaceutical ingredients (API) in 1:1 ratio was developed (Maxiban®). Very high levels of narasin caused death in sows, leg muscle weakness in turkeys, and cardiopulmonary clinical signs in 15% of the rabbits from Brazilian rabbit farms (Oehme and Pickrell 1999).
Monovalent glycosidic ionophores
Maduramicin
The ionophores maduramicin (also called Yumamycin; Fig. 1c) was first isolated from the bacterium Actinomadura yumaensis (Liu et al. 1983). It is a large heterocyclic compound with a series of electronegative crown ethers able to bind monovalent or divalent metal ions (Maron et al. 2015) and is widely used for commercial broiler production. Maduramicin is the most toxic of all the ionophores for non-target animals (Oehme and Pickrell 1999) and humans (Sharma et al. 2005). It might cause severe cardiovascular defects (Shlosberg et al. 1997), as it inhibits proliferation and induces apoptosis in myoblasts (Chen et al. 2014).
Semduramicin
Semduramicin (Fig. 1d) can be isolated from Actinomadura roserufa (Tynan et al. 1992). It is a highly effective drug against Eimeria and is well tolerated by chickens (Ricketts et al. 1992; Logan 1993).
Divalent ionophores
Lasalocid
Compound X-537A (later named lasalocid A; Fig. 1f) was isolated from Streptomyces lasaliensis. It was shown to have anticoccidial activity in chicken (Mitrovic and Schildknecht 1974) and to increase weight gain and feed conversion (Reid 1975).
With the exception of salinomycin, lasalocid is the one with lowest toxicity (Oehme and Pickrell 1999). Nevertheless, dogs appear to be more sensitive to lasalocid intoxication than other species, and accidental poisoning of dogs by lasalocid has been reported (Espino et al. 2003; Segev et al. 2004).
Synthetic compounds
Based on chemical structure, synthetic drugs include the quinolones, pyridones, alkaloids, guanidines, thiamine analogues, and triazine derivatives (Fig. 2). The mode of action of some synthetic anticoccidials has been described (Wang 1978, 1982) but for others their mode of action needs to be investigated (e.g., diclazuril, halofuginone, nicarbazin, robenidine). Such information and its relevance to the inhibition of specific developmental stages of the life cycle of the parasites are important in understanding toxicity and adverse effects of synthetic anticoccidials, and to obtain optimal control by correct timing of prophylaxis. Structures of synthetic anticoccidials are shown in Fig. 2.
Fig. 2.
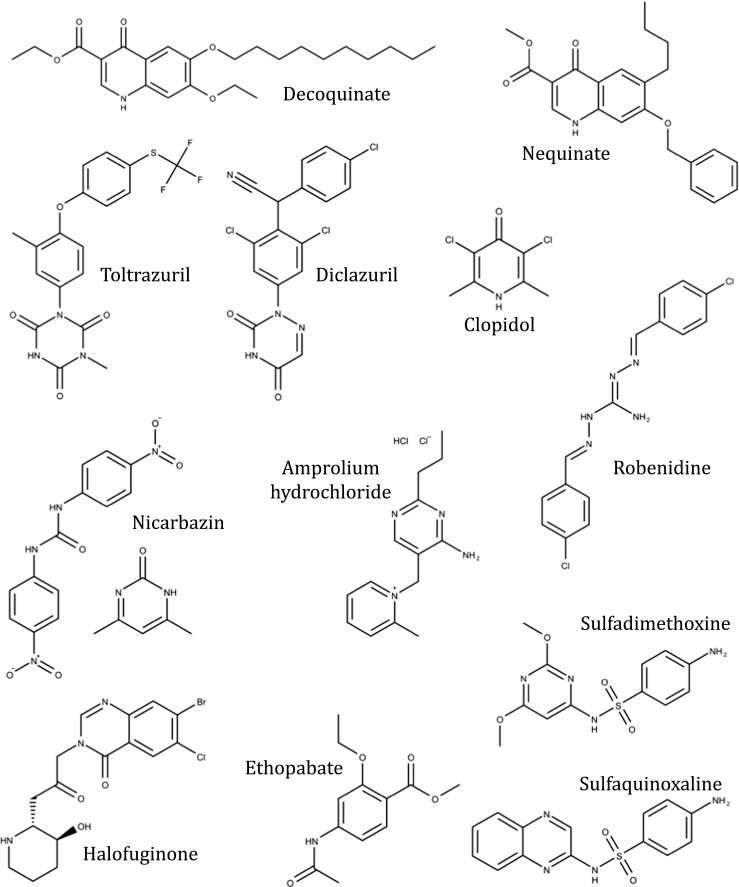
Synthetic anticoccidial APIs
Inhibition of parasite mitochondrial respiration
One of the main targets for anticoccidial drugs is the respiratory chain, which is different from vertebrates as Eimeria tenella oocysts predominantly use succinate or malate plus pyruvate to consume oxygen (Wang 1978). As the respiratory chain of protozoa is relatively insensitive to rotenone and amytal, it can be assumed that NADH dehydrogenase is less important than succinate dehydrogenase (Harder and Haberkorn 1989). Clopidol and quinolones inhibit mitochondrial energy production during the early stages of Eimeria development, but act on different strains of the coccidia (Anon 2010; Fry and Williams 1984).
Quinolones (decoquinate, nequinate (methyl benzoquate))
Quinolones were first discovered in 1962 and since then have undergone numerous modifications to their nucleus to improve spectrum as well as pharmacokinetics (Galarini et al. 2009). They arrest or kill sporozoites or early trophozoites, but even though they cover a broad spectrum, they are not able to fully control coccidiosis. Quinolone coccidiostats inhibit the respiration by blocking electron transport in the parasite mitochondrion reversibly, probably acting at a site near cytochrome b (Wang 1975, 1976).
Methyl benzoquate, an alkoxy-quinolone ester, acts synergistically with clopidol, a pyridone derivative to prevent Eimeria infections in chicken (Ryley 1975). Decoquinate (6-ethyl-(decycloxy)-7-ethoxy-4-hydroxy-3-quinolinecarboxylate) has been introduced in 1967 (Williams 2006). It also shows synergistic effects when combined with clopidol in low concentrations, inhibiting the electron transport more effectively than the sum of their individual actions (Fry and Williams 1984; Wang 1975; Williams 1997). Thus, both APIs are often used in combination products with clopidol. Nevertheless, quinolones and their combination with clopidol are used less frequently today because of the relatively rapid development of resistance (Chapman 1997; Kawazoe et al. 1991).
Clopidol
Clopidol, also known as meticlorpindol or clopindol, is a pyridinol with broad coccidiostatic activity against early development of Eimeria spp. by inhibiting mitochondrial energy production in sporozoites and trophozoites (Kant et al. 2013). A synergistic effect between meticlorpindol and 4-hydroxyquinolones has been described (Challey and Jeffers 1973; Jeffers and Challey 1973). To achieve complete control, combination products with quinolones are marketed.
Toltrazuril
Toltrazuril interacts with the mitochondrial pyrimidine biosynthesis linked to the respiratory chain; thus, it presumably inhibits mitochondrial dihydroorotate dehydrogenase (Harder and Haberkorn 1989; Jockel et al. 1998). In addition, toltrazuril might affect plastid-like organelles (Hackstein et al. 1995).
Toltrazuril is one of the triazines that acts on intracellular stages of the life cycle that are undergoing schizogony and gamogony (Haberkorn and Stoltefuss 1987), especially affecting mitochondria and the endoplasmatic reticulum (Mehlhorn et al. 1984). Respiratory chain enzymes like succinate-cytochrome C reductase, NADH oxidase and fumarate reductase as well as enzymes involved in pyrimidine synthesis are inhibited by toltrazuril. However, if one considers the high concentrations needed for inhibition of the latter enzymes, it is questionable whether this mechanism would translate into an anticoccidial effect (Harder and Haberkorn 1989). In addition, nuclear division in schizonts and microgamonts as well as the wall-forming bodies in macrogamonts are disturbed (Mehlhorn et al. 1984).
Inhibition of the folic acid pathway
The folic acid antagonists include sulfonamides, 2,4-diaminopyrimidines, and ethopabate, which are structural analogues of folic acid or of para-aminobenzoic acid (PABA), a precursor of folic acid. They interfere with the synthesis of folic acid by competing with PABA, thereby inhibiting folate synthetase, and thus preventing cellular replication (Lebkowska-Wieruszewska and Kowalski 2010). Diaveridine and ormetoprim are active against the protozoan enzyme dihydrofolate reductase (Lindsay et al. 1996). As coccidia rapidly synthesize nucleic acids, they have high requirements of folic acid — in contrast to their hosts, which are able to utilize folic acid from feed and thus, have no need for PABA (Zaionts et al. 1978).
Sulfonamides
Sulfonamides (sulfadimethoxine, sulfaquinoxaline) inhibit dihydropteroate synthetase (McCullough and Maren 1974). They have broad-spectrum activity against Gram-negative and Gram-positive bacteria as well as protozoa. Accidental human consumption of sulfonamide-contaminated products can cause central nervous system effects, gastrointestinal disturbances, and hypersensitivity reactions (Lebkowska-Wieruszewska and Kowalski 2010). Sulfonamides act on developing schizonts and on sexual stages.
Sulfonamides are only used very rarely in US broiler production because of the high potential for residues. On rare occasions only, a combination of sulfadimethoxine and ormetoprim is used in a “prestarter feed” for birds under 16 weeks of age to prevent mortality from coccidiosis and bacterial infections with a 5-day meat withdrawal period (United States Food and Drug Administration, FDA 2016). In Europe, sulfonamides are not approved for prevention of coccidiosis in poultry.
Ethopabate
Ethopabate is an antagonist of folic acid or of its precursor, PABA, thus inhibiting the synthesis of nucleic acid and limiting the production of new cells (Anon 2010). It is most active against Eimeria maxima and Eimeria brunetti (Peek and Landman 2011). As it lacks activity against E. tenella caecal stages, it is often used in combination products with amprolium.
Competitive inhibition of thiamine uptake
Amprolium
Amprolium hydrochloride (1-[(4-amino-2-propyl-5-pyrimidinyl)methyl]-2-methylpyridinium chloride monohydrochloride) is an analogue to thiamine (vitamin B1), but lacks the hydroxyethyl functionality that thiamine possesses and thus is not phosphorylated to a pyrophosphate analogue (Kart and Bilgili 2008). It inhibits the uptake of thiamine by second generation schizonts of E. tenella and prevents formation of thiamine pyrophosphate which is required for many essential metabolic reactions, e.g., as cofactor of several decarboxylase enzymes involved in cofactor synthesis (James 1980).
As amprolium is only poorly active against some Eimeria spp., it is largely used in combination products or mixtures with the folic acid antagonists ethopabate or sulfaquinoxaline to extend its spectrum of activity. The primary use of amprolium today is for water treatment during clinical outbreaks. Amprolium is the only active pharmaceutical ingredient approved for prevention and treatment in laying chicken. It has a large safety window (at least 5:1 when used at the recommended level in feed (125 ppm)) (Rychen et al. 2018).
Other modes of action
Nicarbazin
Nicarbazin is an equal molar complex of 4,4′-dinitrocarbanalide and 2-hydroxy-4,6-dimethylpyrimidine (Chapman 1994a). It was the first anticoccidial drug with a true broad-spectrum activity and has been in common use since 1955 (Anon 2010). The 4,4′-dinitrocarbanilide component of nicarbazin inhibits transglutaminase activity, whereas the 2-hydroxy-4,6-dimethylpyrimidine portion increases transglutaminase activity. In addition, nicarbazin increases lipoprotein lipase activity and acts as a calcium ionophore (Yoder et al. 2006). Nicarbazin and narasin show synergistic activity (Challey and Jeffers 1973) and a combination product of these drugs was developed. Nicarbazin has only a small safety window. As it disrupts the ion- and water equilibrium, medicated birds are at increased risk of heat stress under hot and humid weather conditions (Keshavarz and McDouglad 1981). In addition, it is highly toxic to layers — symptoms include bleaching of brown-shelled eggs, mottling of yolks, reduced hatchability, and decreased egg production (Jones et al. 1990).
Diclazuril
Like toltrazuril, diclazuril belongs to the chemical class of triazines, developed together with clazuril by Janssen Pharmaceuticals (Maes et al. 1988). Diclazuril is a nucleoside analogue thought to be involved in nucleic acid synthesis, possibly affecting later phases of coccidia differentiation (Verheyen et al. 1988). It has been shown to affect parasite wall synthesis resulting in the formation of an abnormally thickened, incomplete oocyst wall, and zygote necrosis in both E. brunetti and E. maxima (Verheyen et al. 1989). In addition, diclazuril has been shown to cause disruption of transmembrane potential of mitochondria and to induce ultrastructural changes in merozoites (Zhou et al. 2010). Nevertheless, it is not clear if this is the true mode of action or is just a consequence of cell death. Diclazuril was shown to downregulate mRNA expression of the serine/threonine protein phosphatase type 5 (PP5) significantly by 51.4% in E. tenella (Zhou et al. 2013). PP5s of many eukaryotic organisms have important regulatory functions in the cell cycle (Dobson et al. 2001; Lindenthal and Klinkert 2002) and are associated with the apoptosis signal-regulated kinase 1 (ASK1) (Kutuzov et al. 2005).
Unknown modes of action
Halofuginone
Halofuginone hydrobromide is a quinazolinone derivative related to the antimalarial drug febrifuginone. It was originally extracted from leafs and roots of the traditional Chinese herbal Cichroa febrifuga plant, which is used traditionally in Chinese medicine to treat malaria (Pines et al. 2000). It is effective against asexual stages of most species of Eimeria, delaying development (Zhang et al. 2012).
Robenidine
Robenidine (1-3-bis (p-chlorobenzylideneamino)-guanidine hydrochloride) is a synthetic derivative of guanidine introduced in 1972 (Kennett et al. 1974), which does not affect initial intracellular development of coccidian, but prevents formation of mature schizonts. Its MoA is presumed to interfere with energy metabolism by inhibition of respiratory chain phosphorylation and ATPases, and to inhibit oxidative phosphorylation (Wong et al. 1972; Kant et al. 2013).
Markets and market products
About 59 billion broilers, 5.8 billion layers, and 1400 billion eggs are produced each year worldwide. The global anticoccidial poultry market as estimated by the animal health industry is approx. US$ 1 billion (based on Boehringer Ingelheim internal analysis 2016). Academic estimates calculate that the global loss due to coccidiosis in poultry is in excess of US$ 3 billion (Williams 1999; Shirley et al. 2007). Specifying a value of the market and the cost of coccidiosis in poultry is not an easy if not an impossible task because small errors in such calculations can cause huge differences and there are many different estimates and continuously changing factors involved (e.g., currencies, inflation, energy cost) (Williams 1999). Nevertheless, it is reasonable to speculate how much of these costs can be exploited commercially. If only half of the estimates, it would still be in the range of US$ 0.5 billion and therefore deserving of further research. To illustrate distinctions in differently regulated markets, we will focus in the next sections on marketed products in Europe, the USA, and Australia (Fig. 3).
Fig. 3.

The global anticoccidial poultry market. a Endoparasiticides make up 85% of the total global anticoccidial poultry market; b the North American and European poultry market are currently the most relevant ones (Boehringer Ingelheim internal analysis 2016)
European Union
In the EU, chemicals are rarely used apart from the synthetic/ionophore combination product Maxiban®. In contrast, in the USA synthetic anticoccidials are often employed in rotational programs with ionophores. Nevertheless, they represent a minor part of the coccidiosis control program. Ionophores are most widely used in the EU, also due to their antibiotic affects in the intestine, e.g., against dysbacteriosis caused by clostridia. The ionophores salinomycin, narasin, monensin, lasalocid, maduramicin, and semduramicin and the chemical anticoccidial drugs robenidine, decoquinate, halofuginone, nicarbazin, and diclazuril are licensed in the EU as zootechnical feed additives under regulation 1831/2003/EC in species where coccidiosis is systematic for biological and zootechnical reasons, which is the case for poultry and rabbits. Systematic means that in these species, diagnosis of coccidiosis is not required and therefore, no prescription is necessary. By contrast, in species where coccidiosis is not systematic, anticoccidials are registered as veterinary medicines (e.g., for cattle; regulated in the European Union by Directive 2001/82/EC). In the UK, more than 40% of all antimicrobials sold for use in food and non-food animals are employed for the control of coccidia (277 tons of active ingredient in 2011; mostly for control of Eimeria) with ionophores representing more than 70% of these (Veterinary Medicines Directorate 2012) (Table 1).
Table 1.
Anticoccidial products and APIs approved in Europe for use in poultry (data retrieved from European Food Safety Authority http://www.efsa.europa.eu/, Department for Environment, Food & Rural Affairs https://www.vmd.defra.gov.uk/ProductInformationDatabase)
| Anticoccidial API | Trade name | Company |
|---|---|---|
| Decoquinate | Deccox | Alpharma BVBA; Zoetis SA |
| Diclazuril |
Clinacox Coxiril |
Eli Lilly and Company Ltd; Janssen Pharmaceutica NV Huvepharma NV |
| Halofuginone | Stenorol | Huvepharma NV |
| Lasalocid A | Avatec | Alpharma BVBA; Zoetis SA |
| Maduramicin | Cygro | Alpharma BVBA; Zoetis SA |
| Monensin | Coxidin | Huvepharma NV |
| Elancoban | Eli Lilly and Company Ltd | |
| Narasin | Monteban | Eli Lilly and Company Ltd |
| Narasin + nicarbazin | Maxiban | Eli Lilly and Company Ltd |
| Nicarbazin | Koffogran (Nicarb) | Phibro Animal Health SA |
| Robenidine | Robenz (Cycostst) | Alpharma BVBA; Zoetis SA |
| Salinomycin | Huvesal, Sacox | Huvepharma EOOD; Huvepharma NV |
| Salinomax | Alpharma BVBA | |
| Semduramicin | Aviax 5% | Phibro Animal Health SA |
| Toltrazuril | Baycox | Bayer AH |
USA
Rotation programs including ionophores and synthetic anticoccidials are the standard in intensive broiler production in the USA. These programs sometimes are also combined with vaccination. In contrast to the EU, the use of antibacterials for growth promotion is allowed in the USA. However, with the Guidance for Industry (GFI) 209 and GFI 213 (FDA 2013), the FDA has enhanced control of use of medically important antibacterials, eliminating the use of them for growth promotion. In addition, in 2017, the USA restricted the use of medically important antibiotics in feed to Veterinary Feed Directives (VFD) that require veterinary oversight (Castanon 2007; Federal Register 2015). The use of a VFD drug in feed is permitted only under the professional supervision of a licensed veterinarian (https://www.fda.gov/animalveterinary/developmentapprovalprocess/ucm455416.htm), while administration in drinking water still requires prescription. This restriction led to the withdrawal of some old anticoccidials, as for all, new VFD registrations were required, especially for combination products (Table 2).
Table 2.
Anticoccidial products and APIs approved by the FDA for use in poultry (data retrieved from U.S. Food & Drug Administration https://www.fda.gov/AnimalVeterinary/default.htm)
| Anticoccidial API | first approved | Trade names | Company | Combination products available with |
|---|---|---|---|---|
| Amprolium | 1960 | Amprol; Corid; Amprolium-P | Huvepharma EOOD | Bacitracin, |
| AmproMed P | Cross Vetpharm Group Ltd. | Bambermycins, | ||
| Cocciprol | Phibro Animal Health Corp. | Virginiamycin | ||
| Clopidol | 1968 | Coyden 25 | Huvepharma EOOD | Bacitracin, Bambermycins, Chlortetracycline |
| Decoquinate | 1970 | Deccox | Zoetis Inc. | Bacitracin, Chlortetracycline, Lincomycin |
| Diclazuril | 1999 | Clinacox | Huvepharma EOOD | Bacitracin, Bambermycins, Virginiamycin |
| Halofuginone | 1987 | Stenorol | Huvepharma EOOD | Bacitracin, Bambermycins |
| Lasalocid | 1976 | Avatec | Zoetis Inc. | Bacitracin, Bambermycins, Virginiamycin |
| Maduramicin | 1989 | Cygro | Zoetis Inc. | – |
| Monensin | 1971 | Coban 90 Coban 60 | Elanco US Inc. | Avilamycin, Bacitracin, Bambermycins, Chlortetracycline, Lincomycin, Ractopamine, Oxytetracycline, Virginiamycin, Tilmicosin |
| Narasin | 1988 | Monteban 45 | Elanco US Inc. | Avilamycin, Bacitracin, Bambermycins |
| Nicarbazin | 1955 | Nicarb 25% | Phibro Animal Health Corp. | Bacitracin, Bambermycins |
| Nicarbazin; Carbigran 25 | Elanco US Inc. | |||
| Nicarmix 25 | Planalquimica Industrial Ltda. | |||
| Robenidine | 1972 | Robenz | Zoetis Inc. | Bacitracin, Chlortetracycline, Lincomycin, Oxytetracycline |
| Salinomycin | 1983 |
Bio-Cox Type A Medicated Article Sacox 60 |
Huvepharma EOOD | Avilamycin, Bacitracin, Bambermycins, Chlortetracycline, Lincomycin, Oxytetracycline, Virginiamycin |
| Semduramicin | 1995 | Aviax | Phibro Animal Health Corp. | Bacitracin, Virginiamycin |
| Sulfachloropyrazine | ESB 3 | Zoetis Inc. | – | |
| Sulfamethazine | 1945 |
SMZ-Med 454 Sulmet Soluble Powder |
Cross Vetpharm Group Ltd. Huvepharma EOOD |
– |
| Sulfadimethoxine |
ALBON; AGRIBON SDM Sulfadimethoxine Concentrated Solution 12.5% Sulfadimethoxine Soluble Powder Sulfamed-G Sulfasol Soluble Powder; Sulforal DI-METHOX; Sulfadimethoxine 12.5% Oral Solution; Sulmet Drinking Water Solution, 12.5% |
Zoetis Inc. Cronus Pharma LLC Phibro Animal Health Corp. Cross Vetpharm Group Ltd. Med-Pharmex, Inc. Huvepharma EOOD |
– | |
| Sulfaquinoxaline | 1948 |
20% Sulfaquinoxaline Sodium Solution; 25% S.Q. Soluble; S.Q. 40%; Sul-Q-Nox Sulquin 6-50 Concentrate |
Huvepharma EOOD Zoetis Inc. |
– |
| Zoalene | 1960 |
Zoalene 90 Medicated Coccidiostat Zoamix Type A Medicated Article |
Zoetis Inc. | Bacitracin, Bambermycins, Lincomycin |
| Amprolium + ethopabate | 1997 |
Amprol Plus Amprol Hi-E Amprol Plus 3-Nitro |
Huvepharma EOOD | Bacitracin, Bambermycins, Chlortetracycline |
| Narasin + nicarbazin | 1989 | Maxiban 72 | Elanco US Inc. | Avilamycin, Bacitracin, Bambermycins |
| Ormetoprim + sulfadimethoxine | 1970 | Rofenaid 40 | Zoetis Inc. | – |
| Sulfamethazine + sulfaquinoxaline | 2006 | PoultrySulfa | Huvepharma EOOD | Sulfamerazine |
Australia
All agricultural and veterinary chemical products sold in Australia have to be registered by the Australian Pesticides and Veterinary Medicines Authority (APVMA). The “Guideline for the evaluation of the efficacy and safety of coccidiostats” (https://apvma.gov.au/node/427) has to be followed (Table 3).
Table 3.
Anticoccidial products and APIs approved in Australia for use in poultry (data retrieved from Australian Pesticides and Veterinary Medicines Authority https://apvma.gov.au)
| Anticoccidial API | First approved | Trade names | Company |
|---|---|---|---|
| 3,5-Dinitro-O-toluamide | 1994 | Dot | Dox-Al Australia PTY Ltd |
| Dot premix | Bec Feed Solutions PTY Ltd | ||
| Doteco | International Animal Health Products PTY Ltd | ||
| Nutridot | Nutriment Health PTY Ltd | ||
| Phibrodot | Phibro Animal Health PTY Limited | ||
| Amprolium | 1996 | Amprolium | Parafarm PTY Ltd |
| Decoquinate | 2016 | Deccox | Zoetis Australia PTY Ltd |
| Lasalocid | 2001 | Avatec | Zoetis Australia PTY Ltd |
| Maduramicin | 1997 | CyGro | Zoetis Australia PTY Ltd |
| Maduradox | Dox-Al Australia PTY Ltd | ||
| Monensin | 1994 | CCD Monensin, Rumensin | Elanco Australasia PTY Ltd |
| Coxidin | Huvepharma EOOD | ||
| Doxaban | Dox-Al Italia S.P.A. | ||
| Monendox | Dox-Al Australia PTY Ltd | ||
| Moneco | International Animal Health Products PTY Ltd | ||
| Neove Monensin | Nutriment Health PTY Ltd | ||
| Phibromonensin | Phibro Animal Health PTY Limited | ||
| Narasin | 1984 | Elanco Narasin, Monteban | Elanco Australasia PTY Ltd |
| Nicarbazin | 1996 | Carbidox | Dox-Al Australia PTY Ltd |
| Cycarb | Zoetis Australia PTY Ltd | ||
| Elanco Nicarbazin, Carbigran | Elanco Australasia PTY Ltd | ||
| Keymix | International Animal Health Products PTY Ltd | ||
| Nutrinicarb | Nutriment Health PTY Ltd | ||
| Phicarb | Phibro Animal health PTY limited | ||
| Robenidine | 2003 | Cycostat | Zoetis Australia PTY Ltd |
| Nutrirob | Nutriment Health PTY Ltd | ||
| Salinomycin | 1996 | Bio-Cox, Sacox | Huvepharma EOOD |
| CCD Salinomycin | CCD Animal Health PTY Ltd | ||
| Coxistac | Phibro Animal Health PTY Limited | ||
| Neove | Nutriment Health PTY Ltd | ||
| Sadox, Salindox | Dox-Al Australia PTY Ltd | ||
| Doxalino | Dox-Al Italia S.P.A | ||
| Saleco | International Animal Health Products PTY Ltd | ||
| Semduramicin | 1998 | Aviax | Phibro Animal health PTY limited |
| Sulfaquinoxaline | 1983 | Inca Sulpha-Quin | Inca (Flight) Co PTY Ltd |
| Toltrazuril | 1993 | Baycox, Toltracox Poultry | Bayer Australia Ltd. (Animal Health) |
| Coxi-Stop | Abbey Laboratories PTY Ltd | ||
| Diaveridine + sulfaquinoxaline | 1990 | Keymix Solquin | International Animal Health Products PTY Ltd |
| Amprolium + ethopabate | 1989 | Keymix Keystat | International Animal Health Products PTY Ltd |
| Maduramicin + nicarbazin | 2005 | Gromax | Zoetis Australia PTY Ltd |
| Methyl benzoquate + clopidol | 1993 | Lerbek | Feedworks PTY Ltd |
Control of coccidiosis in ruminants and swine
As with poultry, Eimeria infections in ruminants are ubiquitously present in the environment and occur wherever animals are raised, especially in heavily stocked pasture with intensive grazing and crowded conditions such as feedlots (Daugschies and Najdrowski 2005; Keeton and Navarre 2018; Taylor and Catchpole 1994). In a study on first-year grazing cattle in Germany, up to 90% of all animals showed Eimeria-positive fecal samples (von Samson-Himmelstjerna et al. 2006). Probing the scientific literature, it seems that only little research in ruminants has been undertaken (or at least published) for many years, and only a few drugs are currently approved for use. Many of the difficulties in the interpretation of drug efficacy have been described (Gregory et al. 1982). For example, a decrease in oocyst production may follow treatment but this may not be associated with better weight gain and clinical condition. Many field studies show the effects of drugs upon oocyst production but effects upon clinical criteria require artificial inoculation of very large numbers of oocysts. To provide a framework for evaluation of efficacy of anticoccidials in mammals, a respective WAAVP guideline has been published recently (Joachim et al. 2018). This guideline proposes how to conduct both experimental and field studies for dose determination, dose confirmation, and assessment of field effectiveness to obtain solid efficacy data. In addition, guidance on selection of animals, diagnostic techniques, statistical evaluation, and methods concerning preparation, maintenance, and application of parasites is provided.
There are several anticoccidial drugs available for treatment and prevention of coccidiosis in ruminants, both from the class of synthetic drugs (e.g., sulfonamides, amprolium, decoquinate, the triazines diclazuril and toltrazuril) and ionophores (monensin, lasalocid). These drugs are used either therapeutically for the treatment of young animals showing signs of infection (e.g., amprolium, sulfonamides; triazines) or for prevention by inclusion in the feed (e.g., decoquinate, monensin, lasalocid). As most of the damage to the intestine is already present before diagnosis of coccidiosis occurs, therapy is mostly not sufficient to cure, but rather useful to hinder further spread of disease. Therapy has to be combined with palliative application of electrolytes, glucose, and anti-diarrheals to help to maintain hemostasis (Daugschies and Najdrowski 2005).
Only three APIs are currently used for treatment of porcine coccidiosis: narasin, salinomycin, and toltrazuril. Resistance against toltrazuril has been observed not only for Eimeria isolates from chicken (Stephan et al. 1997) and ovine Eimeria field isolates (Odden et al. 2018), but also in a field isolate of Cystoisospora suis, the usual cause of coccidiosis in very young pigs (Shrestha et al. 2017b).
Ionophores
The divalent ionophore lasalocid has been shown to decrease oocyst production in naturally infected lambs and cattle and in a few cases improve performance and reduce clinical signs of disease when included in the feed (Foreyt et al. 1986; Horton and Stockdale 1981). The efficacy of the monovalent ionophore monensin, employed as a preventive drug by incorporation in the feed of livestock, is well documented (e.g., Bergstrom and Maki 1976; Fitzgerald and Mansfield 1978; Gregory et al. 1983; Leek et al. 1976; McDougald 1978).
Amprolium
Amprolium has been used for many years for the control coccidiosis in sheep and cattle. The drug is principally used for the treatment of animals showing clinical signs of disease but may be employed for prevention by inclusion in the feed. It is available as an oral solution, soluble powder, or as a pelleted feed additive. The drug was shown to reduce oocyst production in lambs when given as an in-feed medication and an outbreak of clinical coccidiosis was successfully controlled by single drenching followed by medication (Talmon et al. 1989). Amprolium was effective for the control of coccidiosis in feedlot lambs (Baker et al. 1972) and in cattle (Norcross et al. 1974).
Decoquinate
This drug is used for the prevention of coccidiosis in young ruminants by incorporation in the feed. Its efficacy has been demonstrated in sheep, goats, and cattle (Fitzgerald and Mansfield 1989; Foreyt 1987; Miner and Jensen 1976).
Triazines
Numerous studies have shown that diclazuril and toltrazuril, when administered orally to young cattle, lambs, or pigs prior to the onset of clinical signs (referred to as metaphylactic treatment), decreases oocyst production in natural and artificial infections with Eimeria species (Bohrmann 1991; Mundt et al. 2003a, 2005, 2009; Epe et al. 2005; Daugschies et al. 2007) or Isospora suis (Driesen et al. 1995; Mundt et al. 2003b, 2007; Kreiner et al. 2011). In some cases, improvements in performance have been demonstrated (e.g., Ruiz et al. 2012; Scala et al. 2009; Rypula et al. 2012).
Sulfonamides
Various sulfonamides, such as sulfaquinoxaline and sulfaguanidine, have been used for many years for the treatment of livestock showing clinical signs of coccidiosis (e.g. Hammond et al. 1956). As appetite is depressed in infected animals then inclusion in the drinking water is preferable for treatment (Shumard 1957).
Registered APIs in different markets
European Union
In the European Union, only few anticoccidial APIs are registered for use in ruminants and/or pigs (mostly piglets): decoquinate, diclazuril, lasalocid, monensin, and toltrazuril (European Medicines Agency www.EMA.Europa.eu).
USA
In the USA, anticoccidials for use in ruminants and pigs are mainly used in combinations as feed additives, combined with antibacterials and growth promoters. Registered APIs are as follows:
| Amprolium | Clopidol | Decoquinate | Dexamethasone |
| Diclazuril | Ethopabate | Halofuginone | Lasalocid |
| Maduramicin | Monensin | Narasin | Nicarbazin |
| Ormetoprim | Robenidine | Salinomycin | Semduramicin |
| Sulfachloropyrazine | Sulfadimethoxine | Sulfamerazine | Sulfamethazine |
| Sulfaquinoxaline | Tylosin | Zoalene |
(Source: U.S. Food & Drug Administration https://www.fda.gov/AnimalVeterinary/default.htm).
Australia
In Australia, only lasalocid, monensin, narasin, salinomycin, and toltrazuril are registered anticoccidials for treatment of ruminants and/or pigs (Australian Pesticides and Veterinary Medicines Authority https://apvma.gov.au).
Resistance and research for new anticoccidials
Development of resistance is a threat for all drugs that are used extensively for a prolonged time. A consequence of this is documented resistance for all drugs in intensive poultry production (Table 4) (Chapman 1997). In a survey conducted in the USA from 1995 to 1999, it was found that anticoccidials were universally used by 99% of commercial broiler operations (Chapman 2001).
Table 4.
Summary of reported resistance to anticoccidials in field strains of Eimeria (adapted from Chapman 1997)
| Drug | Year of introduction | Country where resistance was first described | Year resistance was first described | Speciesa |
|---|---|---|---|---|
| Sulphaquinoxaline | 1948 | USA | 1954 | Et |
| Nitrofurazone | 1948 | USA | 1955 | Not given |
| Nicarbazin | 1955 | Britain | 1964 | Et |
| Dinitolmide | 1960 | Britain | 1964 | Et, En |
| Amprolium | 1960 | Britain | 1964 | Eb |
| Clopidol | 1966 | Britain | 1969 | Ea, Em, Et |
| Buquinolate | 1967 | USA | 1968 | Not given |
| Methyl benzoquate | 1967 | Britain | 1970 | Et |
| Decoquinate | 1967 | Britain | 1970 | Et |
| Monensin | 1971 | USA | 1974 | Em |
| Robenidine | 1972 | USA | 1974 | Em |
| Halofuginone | 1975 | France | 1986 | Ea, Et |
| Lasalocid | 1976 | USA | 1977 | Ea |
| Arprinocid | 1980 | Britain | 1982 | Et |
| Salinomycin | 1983 | USA | 1984 | Various |
| Diclazuril | 1990 | Brazil | 1994 | Ea, Em, Et |
| Toltrazuril | 1986 | Netherlands | 1993 | Not given |
aSpecies: Ea, E. acervulina; Eb, E. brunetti; Em, E. maxima; En, E. necatrix; Et, E. tenella
The polyether ionophores became the drug of choice in 1972 and remain the most extensively used drugs in poultry as of today. While the development of resistance to ionophores is rather slow probably due to their unique mode of action, resistance development in synthetic drugs that have a specific mode of action seems to appear more rapidly, involving genetic mechanisms. Sometimes resistance was reported shortly after marketing a new drug, an example being arprinocid (Fig. 4) (Chapman 1983). Another example is the quinolone buquinolate (Fig. 4) that was “commercially dead” within 6 months of its introduction due to the sudden appearance of drug resistance. Development of resistance against buquinolate was found to take place after a single experimental passage of Eimeria (Chapman 1975). By comparison, drug resistance against toltrazuril did not occur in at least five successive drug exposures in field studies (Claeskens et al. 2007).
Fig. 4.
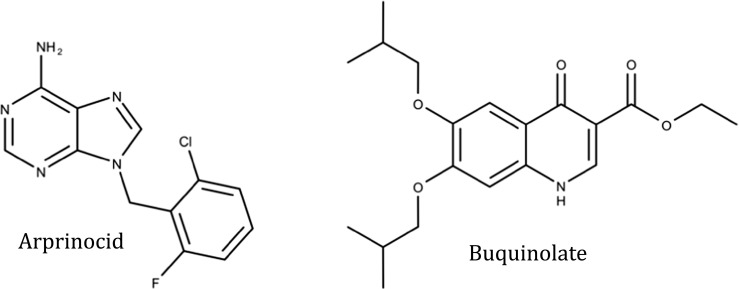
Chemical structures of arprinocid and buquinolate
A study on ten Eimeria field isolates from northern Germany detected resistance against in nine out of the ten isolates of Eimeria, with seven out of the ten having developed multiple resistance (Stephan et al. 1997). Similar results have been shown in isolates from broiler farms in the UK (Chapman 1993) and the Netherlands (Peek and Landman 2006), showing the enormous threat of development of broad resistance against all classes of anticoccidial drugs. As drug sensitivity in a population of coccidia can be altered by the introduction of drug-sensitive coccidia, e.g., through the use of coccidiosis vaccines, or by the use of drug-sensitive laboratory-maintained lines or other reservoirs (Ball 1966; Jeffers 1976; McLoughlin and Chute 1978), these measures have to be combined in an attempt to control coccidiosis. Restoration of sensitivity to drugs following the use of vaccines comprising drug-sensitive strains of Eimeria has been demonstrated for the ionophores, monensin, and salinomycin, and the synthetic drug diclazuril (Chapman 1994b; Chapman and Jeffers 2014, 2015; Jenkins et al. 2010; Peek and Landman 2006; Mathis and Broussard 2006). Partial restoration of sensitivity to diclazuril and monensin was also observed following use of attenuated vaccine (Peek and Landman 2006). Therefore, a yearly rotation program has been proposed in which use of ionophores is alternated in successive flocks with vaccination (Chapman et al. 2010).
The development of resistance led to increasing efforts for the identification of new, resistance-breaking drugs. One recent example is nitromezuril, a new triazine anticoccidial. It shows only limited cross-resistance with diclazuril or toltrazuril (Fei et al. 2013), but these results have to be further evaluated before this drug might finally enter the market.
Conclusion
In the light of the continuously expanding livestock industry and its growing significance for global food production, control of coccidiosis, perhaps the most widespread and intractable disease of poultry and other livestock, is of great importance; however, relatively little effort has been made on Eimeria infections of domestic livestock (Engels et al. 2010; Fernandez et al. 2012; Marhöfer et al. 2013; Müller and Hemphill 2016). In view of advances in biotechnology, modern approaches towards the discovery of novel resistance-breaking drug candidates may be anticipated. Genomic analysis of all seven Eimeria species that cause coccidiosis in poultry has been accomplished (Reid et al. 2014) and this may allow the identification and validation of species-specific protein targets. Novel drug discovery rationales including high throughput screening, structural biology, and the elucidation of the mode of action of active compounds can be envisioned. Combining target-based approaches with parasite in vitro and in vivo testing and medicinal chemistry generates a comprehensive view on the genotype-to-phenotype-to-compound correlation, which could allow for the design of novel drug candidates. Unfortunately, resistance develops rapidly following the introduction of drugs in the field. The current approach to delay the onset of resistance is to employ rotation programs combined with good husbandry, chemoprophylaxis, and/or live parasite vaccination (Blake et al. 2017). However, for the control of coccidiosis in the future, both novel cost-effective preventative chemotherapy and subunit or recombinant vaccines are desperately needed.
Compliance with ethical standards
Conflict of interest
SN and PS are employees of Boehringer Ingelheim Vetmedica GmbH, an organization with commercial interest in the animal health market. HDC is a member of the University of Arkansas, declaring no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Agtarap A, Chamberlin JW, Pinkerton M, Steinrauf L. The structure of monensic acid, a new biologically active compound. J Am Chem Soc. 1967;89:5737–5739. doi: 10.1021/ja00998a062. [DOI] [PubMed] [Google Scholar]
- Alexandratos N, Bruinsma J (2012) World agriculture towards 2030/2050: the 2012 revision (ESA working paper no. 12-03). FAO. Agricultural Development Economics Division, Global Perspective Studies Team http://www.fao.org/fileadmin/templates/esa/Global_persepctives/world_ag_2030_50_2012_rev.pdf
- Allen PC, Fetterer RH. Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clin Microbiol Rev. 2002;15(1):58–65. doi: 10.1128/CMR.15.1.58-65.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anon . The Merck veterinary manual, Vol. 10. New Jersey: Merck & Co., Inc; 2010. [Google Scholar]
- Augustine PC, Watkins KL, Danforth HD. Effect of monensin on ultrastructure and cellular invasion by the turkey coccidia Eimeria adenoeides and Eimeria meleagrimitis. Poult Sci. 1992;71(6):970–978. doi: 10.3382/ps.0710970. [DOI] [PubMed] [Google Scholar]
- Baker NF, Walters GT, Fisk RA. Amprolium for control of coccidiosis in feedlot lambs. Am J Vet Res. 1972;33(1):83–86. [PubMed] [Google Scholar]
- Ball SJ. The development of resistance to glycarbylamide and 2-chloro-4-nitrobenzamide in Eimeria tenella in chicks. Parasitology. 1966;56:25–37. doi: 10.1017/s0031182000071067. [DOI] [PubMed] [Google Scholar]
- Bergstrom RC, Maki LR. Coccidiostatic action of monensin fed to lambs: body weight gains and feed conversion efficacy. Am J Vet Res. 1976;37(1):79–81. [PubMed] [Google Scholar]
- Bezerra PS, Driemeier D, Loretti AP, Riet-Correa F, Kamphues J, de Barros CS. Monensin poisoning in Brazilian horses. Vet Hum Toxicol. 1999;41(6):383–385. [PubMed] [Google Scholar]
- Bila CG, Perreira CL, Gruys E. Accidental monensin toxicosis in horses in Mozambique. J South African Vet Assoc. 2001;72(3):163–164. doi: 10.4102/jsava.v72i3.641. [DOI] [PubMed] [Google Scholar]
- Blake DP, Pastor-Fernandez I, Nolan MJ, Tomley FM. Recombinant anticoccidial vaccines—a cup half full? Infect Genet Evol. 2017;55:358–365. doi: 10.1016/j.meegid.2017.10.009. [DOI] [PubMed] [Google Scholar]
- Bohrmann R. Treatment with toltrazuril in a natural outbreak of coccidiosis in calves. Dtsch Tierarztl Wochenschr. 1991;98(9):343–345. [PubMed] [Google Scholar]
- Broz J, Frigg M. Incompatibility between lasalocid and chloramphenicol in broiler chicks after a long-term simultaneous administration. Vet Res Commun. 1987;11(2):159–172. doi: 10.1007/BF00344947. [DOI] [PubMed] [Google Scholar]
- Castanon JI. History of the use of antibiotic as growth promoters in European poultry feeds. Poult Sci. 2007;86(11):2466–2471. doi: 10.3382/ps.2007-00249. [DOI] [PubMed] [Google Scholar]
- Challey JR, Jeffers TK. Synergism between 4-hydroxyquinoline and pyridone coccidiostats. J Parasitol. 1973;59(3):502–504. [PubMed] [Google Scholar]
- Chapman HD. Eimeria tenella in chickens: development of resistance to quinolone anticoccidial drugs. Parasitology. 1975;71:41–49. doi: 10.1017/s0031182000053130. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Field isolated of Eimeria resistant to arprinocid. Vet Parasitol. 1983;12(1):45–50. doi: 10.1016/0304-4017(83)90086-9. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Resistance to anticoccidial drugs in fowl. Parasitol Today. 1993;9:159–162. doi: 10.1016/0169-4758(93)90137-5. [DOI] [PubMed] [Google Scholar]
- Chapman HD. A review of the biological activity of the anticoccidial drug nicarbazin and its application for the control of coccidiosis in poultry. Poult Sci Rev. 1994;5:231–243. [Google Scholar]
- Chapman HD. Sensitivity of field isolates of Eimeria to monensin following the use of a coccidiosis vaccine in broiler chickens. Poult Sci. 1994;73:476–478. doi: 10.3382/ps.0730476. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Biochemical, genetic and applied aspects of drug resistance in Eimeria parasites of the fowl. Avian Pathol. 1997;26:221–244. doi: 10.1080/03079459708419208. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Anticoccidial drugs and their effects upon the development of immunity to Eimeria infections in poultry. Avian Pathol. 1999;28:521–535. doi: 10.1080/03079459994317. [DOI] [PubMed] [Google Scholar]
- Chapman HD. The development of immunity to Eimeria species in broilers given anticoccidial drugs. Avian Pathol. 1999;28:155–162. doi: 10.1080/03079459994885. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Use of anticoccidial drugs in broiler chickens in the USA: analysis for the years 1995 to 1999. Poult Sci. 2001;80:572–580. doi: 10.1093/ps/80.5.572. [DOI] [PubMed] [Google Scholar]
- Chapman HD. A landmark contribution to poultry science—prophylactic control of coccidiosis in poultry. Poult Sci. 2009;88:813–815. doi: 10.3382/ps.2008-00316. [DOI] [PubMed] [Google Scholar]
- Chapman HD. Applied strategies for the control of coccidiosis in poultry. CAB Rev. 2018;13:1–11. [Google Scholar]
- Chapman HD, Jeffers TK. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int J Parasitol Drugs Drug Resist. 2014;4:214–217. doi: 10.1016/j.ijpddr.2014.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapman HD, Jeffers TK. Restoration of sensitivity to salinomycin in Eimeria following 5 flocks of broiler chickens reared in floor-pens using drug programs and vaccination to control coccidiosis. Poult Sci. 2015;94:943–946. doi: 10.3382/ps/pev077. [DOI] [PubMed] [Google Scholar]
- Chapman HD, Cherry TE, Danforth HD, Richards G, Shirley MW, Williams RB. Sustainable coccidiosis control in poultry production: the role of live vaccines. Int J Parasitol. 2002;32:617–629. doi: 10.1016/s0020-7519(01)00362-9. [DOI] [PubMed] [Google Scholar]
- Chapman HD, Jeffers TK, Williams RB. Forty years of monensin for the control of coccidiosis in poultry. Poult Sci. 2010;89:1788–1801. doi: 10.3382/ps.2010-00931. [DOI] [PubMed] [Google Scholar]
- Chapman HD, Barta JR, Blake D, Gruber A, Jenkins M, Smith NC, Suo X, Tomley FM. A selective review of advances in coccidiosis research. Adv Parasitol. 2013;83:93–171. doi: 10.1016/B978-0-12-407705-8.00002-1. [DOI] [PubMed] [Google Scholar]
- Chapman HD, Barta JR, Hafeez MA, Matsler P, Rathinam T, Raccoursier M. The epizootiology of Eimeria infections in commercial broiler chickens where anticoccidial drug programs were employed in six successive flocks to control coccidiosis. Poult Sci. 2016;95:1774–1778. doi: 10.3382/ps/pew091. [DOI] [PubMed] [Google Scholar]
- Chen X, Gu Y, Singh K, Shang C, Barzegar M, Jiang S, Huang S. Maduramicin inhibits proliferation and induces apoptosis in myoblast cells. PLoS One. 2014;9(12):e115652. doi: 10.1371/journal.pone.0115652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claeskens M, Verdonck W, Heesen H, Froyman R, Torres A. A field study assessing control of broiler coccidiosis by Paracox™ vaccination or by Toltrazuril (Baycox®) stand-alone treatment. Parasitol Res. 2007;101:105–112. [Google Scholar]
- Dalloul RA, Lillehoj HS. Poultry coccidiosis: recent advancements in control measures and vaccine development. Expert Rev Vaccines. 2006;5(1):143–163. doi: 10.1586/14760584.5.1.143. [DOI] [PubMed] [Google Scholar]
- Daugschies A, Najdrowski M. Eimeriosis in cattle: current understanding. J Vet Med B Infect Dis Vet Public Health. 2005;52(10):417–427. doi: 10.1111/j.1439-0450.2005.00894.x. [DOI] [PubMed] [Google Scholar]
- Daugschies A, Agneessens J, Goossens L, Mengel H, Veys P. The effect of a metaphylactic treatment with diclazuril (Vecoxan) on the oocyst excretion and growth performance of calves exposed to a natural Eimeria infection. Vet Parasitol. 2007;149(3–4):199–206. doi: 10.1016/j.vetpar.2007.08.003. [DOI] [PubMed] [Google Scholar]
- Dobson S, Kar B, Kumar R, Adams B, Barik S. A novel tetratricopeptide repeat (TPR) containing PP5 serine/threonine protein phosphatase in the malaria parasite, Plasmodium falciparum. BMC Microbiol. 2001;1:31. doi: 10.1186/1471-2180-1-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doonan GR, Brown CM, Mullaney TP, Brooks DB, Ulmanis EG, Slanker MR. Monensin poisoning in horses—an international incident. Can Vet J. 1989;30(2):165–169. [PMC free article] [PubMed] [Google Scholar]
- Dowling L. Ionophore toxicity in chickens: a review of pathology and diagnosis. Avian Pathol. 1992;21(3):355–368. doi: 10.1080/03079459208418854. [DOI] [PubMed] [Google Scholar]
- Driesen SJ, Fahy VA, Carland PG. The use of toltrazuril for the prevention of coccidiosis in piglets before weaning. Aust Vet J. 1995;72(4):139–141. doi: 10.1111/j.1751-0813.1995.tb15034.x. [DOI] [PubMed] [Google Scholar]
- Engels K, Beyer C, Suárez Fernández ML, Bender F, Gassel M, Unden G, Marhöfer RJ, Mottram JC, Selzer PM. Inhibition of Eimeria tenella CDK-related kinase 2: from target identification to lead compounds. Chem Med Chem. 2010;5(8):1259–1271. doi: 10.1002/cmdc.201000157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epe C, von Samson-Himmelstjerna G, Wirtherle N, von der Heyden V, Welz C, Beening J, Radeloff I, Hellmann K, Schnieder T, Krieger K. Efficacy of toltrazuril as a metaphylactic and therapeutic treatment of coccidiosis in first-year grazing calves. Parasitol Res. 2005;97:S127–s133. doi: 10.1007/s00436-005-1456-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espino L, Suarez ML, Mino N, Goicoa A, Fidalgo LE, Santamarina G. Suspected lasalocid poisoning in three dogs. Vet Hum Toxicol. 2003;45(5):241–242. [PubMed] [Google Scholar]
- FDA (2013) CVM GFI #213 New Animal Drugs and New Animal Drug Combination Products Administered in or on Medicated Feed or Drinking Water of Food-Producing Animals: Recommendations for Drug Sponsors for Voluntarily Aligning Product Use Conditions with GFI #209 http://www.fda.gov/downloads/AnimalVeterinary/GuidanceComplianceEnforcement/GuidanceforIndustry/UCM299624.pdf. Accessed 11 Apr 2019
- FDA (2016) 21CFR558.575 Sulfadimethoxine and ormetoprim. https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=558.575. Accessed 11 April 2019
- Fei C, Fan C, Zhao Q, Lin Y, Wang X, Zheng W, Wang M, Zhang K, Zhang L, Li T, Xue F. Anticoccidial effects of a novel triazine nitromezuril in broiler chickens. Vet Parasitol. 2013;198(1–2):39–44. doi: 10.1016/j.vetpar.2013.08.024. [DOI] [PubMed] [Google Scholar]
- Fernandez ML, Engels KK, Bender F, Gassel M, Marhöfer RJ, Mottram JC, Selzer PM. High-throughput screening with the Eimeria tenella CDC2-related kinase2/cyclin complex EtCRK2/EtCYC3a. Microbiology. 2012;158:2262–2271. doi: 10.1099/mic.0.059428-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitzgerald PR, Mansfield ME. Ovine coccidiosis: effect of the antibiotic monensin against Eimeria ninakohlyakimovae and other naturally occurring coccidia of sheep. Am J Vet Res. 1978;39(1):7–10. [PubMed] [Google Scholar]
- Fitzgerald PR, Mansfield ME. Effects of intermittent and continuous administration of decoquinate on bovine coccidiosis in male calves. Am J Vet Res. 1989;50(6):961–964. [PubMed] [Google Scholar]
- Foreyt WJ. Coccidiosis in sheep and goats. Vet Hum Toxicol. 1987;29(Suppl 1):60–64. [PubMed] [Google Scholar]
- Foreyt WJ, Rice DH, Wescott RB. Evaluation of lasalocid as a coccidiostat in calves: titration, efficacy, and comparison with monensin and decoquinate. Am J Vet Res. 1986;47(9):2031–2035. [PubMed] [Google Scholar]
- Fry M, Williams RB. Effects of decoquinate and clopidol on electron transport in mitochondria of Eimeria tenella (Apicomplexa: coccidia) Biochem Pharmacol. 1984;33(2):229–240. doi: 10.1016/0006-2952(84)90480-5. [DOI] [PubMed] [Google Scholar]
- Galarini R, Fioroni L, Angelucci F, Tovo GR, Cristofani E. Simultaneous determination of eleven quinolones in animal feed by liquid chromatography with fluorescence and ultraviolet absorbance detection. J Chromatogr A. 2009;1216(46):8158–8164. doi: 10.1016/j.chroma.2009.06.076. [DOI] [PubMed] [Google Scholar]
- Gregory MW, Joyner LP, Catchpole J. Medication against ovine coccidiosis—a review. Vet Res Commun. 1982;5(4):307–325. doi: 10.1007/BF02215000. [DOI] [PubMed] [Google Scholar]
- Gregory MW, Catchpole J, Joyner LP, Parker BN. Observations on the epidemiology of coccidial infections in sheep under varying conditions of intensive husbandry including chemoprophylaxis with monensin. Parasitology. 1983;87:421–427. doi: 10.1017/s0031182000082949. [DOI] [PubMed] [Google Scholar]
- Grumbles LC, Delaplane JP, Higgins TC (1948) Continuous feeding of low concentrations of sulfaquinoxaline for the control of coccidiosis in poultry. Poult Sci 27(5). 10.3382/ps.0270605
- Haberkorn A, Stoltefuss J. Studies on the activity spectrum of toltrazuril, a new anticoccidial agent. Vet Med Rev. 1987;1:22–32. [Google Scholar]
- Hackstein JH, Mackenstedt U, Mehlhorn H, Meijerink JP, Schubert H, Leunissen JA. Parasitic apicomplexans harbor a chlorophyll a-D1 complex, the potential target for therapeutic triazines. Parasitol Res. 1995;81(3):207–216. doi: 10.1007/BF00937111. [DOI] [PubMed] [Google Scholar]
- Hammond DM, Shupe JL, Johnson AE, Fitzgerald PR, Thorne JL. Sulfaquinoxaline and sulfamerazine in the treatment of experimental infections with Eimeria bovis in calves. Am J Vet Res. 1956;17(64):463–470. [PubMed] [Google Scholar]
- Harder A, Haberkorn A. Possible mode of action of toltrazuril: studies on two Eimeria species and mammalian and Ascaris suum enzymes. Parasitol Res. 1989;76(1):8–12. doi: 10.1007/BF00931064. [DOI] [PubMed] [Google Scholar]
- Horton GM, Stockdale PH. Lasalocid and monensin in finishing diets for early weaned lambs with naturally occurring coccidiosis. Am J Vet Res. 1981;42(3):433–436. [PubMed] [Google Scholar]
- Huang L, Yao L, He Z, Zhou C, Li G, Yang B, Deng X. Roxarsone and its metabolites in chicken manure significantly enhance the uptake of As species by vegetables. Chemosphere. 2014;100:57–62. doi: 10.1016/j.chemosphere.2013.12.074. [DOI] [PubMed] [Google Scholar]
- Islam KM, Klein U, Burch DG. The activity and compatibility of the antibiotic tiamulin with other drugs in poultry medicine—a review. Poult Sci. 2009;88(11):2353–2359. doi: 10.3382/ps.2009-00257. [DOI] [PubMed] [Google Scholar]
- James S. Thiamine uptake in isolated schizonts of Eimeria tenella and the inhibitory effects of amprolium. Parasitology. 1980;80(02):313–322. doi: 10.1017/s0031182000000779. [DOI] [PubMed] [Google Scholar]
- Jeffers TK. Reduction of anticoccidial drug resistance by massive introduction of drug-sensitive coccidia. Avian Dis. 1976;20(4):649–653. [PubMed] [Google Scholar]
- Jeffers T (1989) Anticoccidial drug resistance: a review with emphasis on the polyether ionophores. Pages 295–308 In: Coccidia and Intestinal Coccidiomorphs. P Yvoré, ed. Proc. 5th International Coccidiosis Conference, Tours, France. INRA, Paris, France
- Jeffers TK, Challey JR. Collateral sensitivity to 4-hydroxyquinolines in Eimeria acervulina strains resistant to meticlorpindol. J Parasitol. 1973;59(4):624–630. [PubMed] [Google Scholar]
- Jeffers TK, Tonkinson LV, Callender ME. Anticoccidial efficacy of narasin in battery cage trials. Poult Sci. 1988;67(7):1043–1049. doi: 10.3382/ps.0671043. [DOI] [PubMed] [Google Scholar]
- Jenkins M, Klopp S, Ritter D, Miska K, Fetterer R. Comparison of Eimeria species distribution and salinomycin resistance in commercial broiler operations utilizing different coccidiosis control strategies. Avian Dis. 2010;54(3):1002–1006. doi: 10.1637/9137-111109-Reg.1. [DOI] [PubMed] [Google Scholar]
- Joachim A, Altreuther G, Bangoura B, Charles S, Daugschies A, Hinney B, Lindsay DS, Mundt HC, Ocak M, S10 S (2018) W A A V P guideline for evaluating the efficacy of anticoccidials in mammals (pigs, dogs, cattle, sheep). Vet Parasitol 253:102–119. 10.1016/j.vetpar.2018.02.029 [DOI] [PubMed]
- Jockel J, Wendt B, Loffler M. Structural and functional comparison of agents interfering with dihydroorotate, succinate and NADH oxidation of rat liver mitochondria. Biochem Pharmacol. 1998;56(8):1053–1060. doi: 10.1016/s0006-2952(98)00131-2. [DOI] [PubMed] [Google Scholar]
- Jones JE, Solis J, Hughes BL, Castaldo DJ, Toler JE. Production and egg-quality responses of White Leghorn layers to anticoccidial agents. Poult Sci. 1990;69(3):378–387. doi: 10.3382/ps.0690378. [DOI] [PubMed] [Google Scholar]
- Kadykalo S, Roberts T, Thompson M, Wilson J, Lang M, Espeisse O. The value of anticoccidials for sustainable global poultry production. Int J Antimicrob Agents. 2018;51(3):304–310. doi: 10.1016/j.ijantimicag.2017.09.004. [DOI] [PubMed] [Google Scholar]
- Kant V, Singh P, Verma PK, Bais I, Parmar MS, Gopal A, Gupta V. Anticoccidial drugs used in the poultry: an overview. Sci Int. 2013;1(7):261–265. [Google Scholar]
- Kart A, Bilgili A. Ionophore antibiotics: toxicity, mode of action and neurotoxic aspect of carboxylic ionophores. J Anim Vet Adv. 2008;7(6):748–751. [Google Scholar]
- Kawazoe U, Chapman HD, Shaw M. Sensitivity of field isolates of Eimeria acervulina to salinomycin, maduramicin, and a mixture of clopidol and methyl benzoquate in the chicken. Avian Pathol. 1991;20(3):439–446. doi: 10.1080/03079459108418782. [DOI] [PubMed] [Google Scholar]
- Keeton STN, Navarre CB. Coccidiosis in large and small ruminants. Vet Clin N Am Food Anim Pract. 2018;34(1):201–208. doi: 10.1016/j.cvfa.2017.10.009. [DOI] [PubMed] [Google Scholar]
- Kennett RL, Kantor S, Gallo A. Efficacy studies with robenidine, a new type of anticoccidial, in the diet. Poult Sci. 1974;53(3):978–986. doi: 10.3382/ps.0530978. [DOI] [PubMed] [Google Scholar]
- Keshavarz K, McDougald LR. Influence of anticoccidial drugs on losses of broiler chickens from heat stress and coccidiosis. Poult Sci. 1981;60(11):2423–2428. doi: 10.3382/ps.0602423. [DOI] [PubMed] [Google Scholar]
- Kinnaird JH, Bumstead JM, Mann DJ, Ryan R, Shirley MW, Shiels BR, Tomley FM. EtCRK2, a cyclin-dependent kinase gene expressed during the sexual and asexual phases of the Eimeria tenella life cycle. Int J Parasitol. 2004;34(6):683–692. doi: 10.1016/j.ijpara.2004.01.003. [DOI] [PubMed] [Google Scholar]
- Klotz C, Marhöfer RJ, Selzer PM, Lucius R, Pogonka T. Eimeria tenella: identification of secretory and surface proteins from expressed sequence tags. Exp Parasitol. 2005;111(1):14–23. doi: 10.1016/j.exppara.2005.04.005. [DOI] [PubMed] [Google Scholar]
- Kreiner T, Worliczek HL, Tichy A, Joachim A. Influence of toltrazuril treatment on parasitological parameters and health performance of piglets in the field—an Austrian experience. Vet Parasitol. 2011;183(1–2):14–20. doi: 10.1016/j.vetpar.2011.07.019. [DOI] [PubMed] [Google Scholar]
- Kutuzov MA, Andreeva AV, Voyno-Yasenetskaya TA. Regulation of apoptosis signal-regulating kinase 1 (ASK1) by polyamine levels via protein phosphatase 5. J Biol Chem. 2005;280(27):25388–25395. doi: 10.1074/jbc.M413202200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laczay P. [Compatibility of ionophore antibiotics.] Zur Kompatibilität der ionophoren Antibiotika. Monatshefte für Veterinärmedizin. 1988;43(22):786–788. [Google Scholar]
- Laczay P, Simon F, Szurop I, Voros G, Lehel J (1988) Study of the compatibility of CH-402 antioxidant with ionophore antibiotics and other chemotherapeutics in broilers. Magyar Allatorvosok Lapja (Hungary)
- Laczay P, Simon F, Mora Z, Lehel J. The compatibility of the new ionophore-coccidiostats with other chemotherapeutics in broilers. Dtsch Tierarztl Wochenschr. 1989;96(9):449–451. [PubMed] [Google Scholar]
- Lebkowska-Wieruszewska BI, Kowalski CJ. Sulfachlorpyrazine residues depletion in turkey edible tissues. J Vet Pharmacol Ther. 2010;33(4):389–395. doi: 10.1111/j.1365-2885.2009.01147.x. [DOI] [PubMed] [Google Scholar]
- Leek RG, Fayer R, McLoughlin DK. Effect of monensin on experimental infections of Eimeria ninakohlyakimovae in lambs. Am J Vet Res. 1976;37(3):339–341. [PubMed] [Google Scholar]
- Lindenthal C, Klinkert MQ. Identification and biochemical characterisation of a protein phosphatase 5 homologue from Plasmodium falciparum. Mol Biochem Parasitol. 2002;120(2):257–268. doi: 10.1016/s0166-6851(02)00007-5. [DOI] [PubMed] [Google Scholar]
- Lindsay DS, Butler JM, Rippey NS, Blagburn BL. Demonstration of synergistic effects of sulfonamides and dihydrofolate reductase/thymidylate synthase inhibitors against Neospora caninum tachyzoites in cultured cells, and characterization of mutants resistant to pyrimethamine. Am J Vet Res. 1996;57(1):68–72. [PubMed] [Google Scholar]
- Liu CM, Hermann TE, Downey A, Prosser BL, Schildknecht E, Palleroni NJ, Westley JW, Miller PA. Novel polyether antibiotics X-14868A, B, C, and D produced by a Nocardia. Discovery, fermentation, biological as well as ionophore properties and taxonomy of the producing culture. J Antibiot. 1983;36(4):343–350. doi: 10.7164/antibiotics.36.343. [DOI] [PubMed] [Google Scholar]
- Logan NB, McKenzie ME, Conway DP, Chappel LR, Hammet NC (1993) Anticoccidial efficacy of semduramicin. 2. Evaluation against field isolates including comparisons with salinomycin, maduramicin, and monensin in battery tests. Poult Sci 72(11):2058–2063 [DOI] [PubMed]
- Maes L, Coussement W, Vanparijs O, Marsboom R. In vivo action of the anticoccidial diclazuril (Clinacox) on the developmental stages of Eimeria tenella: a histological study. J Parasitol. 1988;74(6):931–938. [PubMed] [Google Scholar]
- Marhöfer RJ, Mottram JC, Selzer PM (2013) Protein kinases as suitable targets for combating Eimeria spp. protein phosphorylation in parasites. In: Doerig C, Späth G, Wiese M (eds) Protein phosphorylation in parasites. Wiley-VCH Verlag GmbH & Co. KGaA, pp 317–336. 10.1002/9783527675401.ch15
- Maron MI, et al. Maduramicin rapidly eliminates malaria parasites and potentiates the gametocytocidal activity of the pyrazoleamide PA21A050. Antimicrob Agents Chemother. 2015;60(3):1492–1499. doi: 10.1128/AAC.01928-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathis GF, Broussard C. Increased level of Eimeria sensitivity to diclazuril after using a live coccidial vaccine. Avian Dis. 2006;50(3):321–324. doi: 10.1637/7455-101305R1.1. [DOI] [PubMed] [Google Scholar]
- Matsuoka T, Novilla MN, Thomson TD, Donoho AL. Review of monensin toxicosis in horses. J Equine Vet Sci. 1996;16(1):8–15. [Google Scholar]
- Mazlum Z, Pradella G. Interactions between narasin and monensin with some antibiotics and chemotherapeutics in broiler chicks. Clin Vet. 1986;109:62–63. [Google Scholar]
- Mazlum Z, Pickles RW, Pradella G, Pagnani R. Interaction between monensin, narasin or salinomycin and the antibiotics erythromycin, chloramphenicol or tylosin in broiler chicks. Clinica Vet. 1985;108:95–104. [Google Scholar]
- McCullough JL, Maren TH. Dihydropteroate synthetase from Plasmodium berghei: isolation, properties, and inhibition by dapsone and sulfadiazine. Mol Pharmacol. 1974;10(1):140–145. [PubMed] [Google Scholar]
- McDougald LR. Monensin for the prevention of coccidiosis in calves. Am J Vet Res. 1978;39(11):1748–1749. [PubMed] [Google Scholar]
- McLoughlin DK, Chute MB. Robenidine resistance in Eimeria tenella. J Parasitol. 1978;64(5):874–877. [PubMed] [Google Scholar]
- Mehlhorn H, Pooch H, Raether W. The action of polyether ionophorous antibiotics (monensin, salinomycin, lasalocid) on developmental stages of Eimeria tenella (Coccidia, Sporozoa) in vivo and in vitro: study by light and electron microscopy. Ztsch Parasitenk. 1983;69(4):457–471. doi: 10.1007/BF00927702. [DOI] [PubMed] [Google Scholar]
- Mehlhorn H, Ortmann-Falkenstein G, Haberkorn A. The effects of sym. Triazinones on developmental stages of Eimeria tenella, E. maxima and E. acervulina: a light and electron microscopical study. Ztschr Parasitenk. 1984;70(2):173–182. doi: 10.1007/BF00942219. [DOI] [PubMed] [Google Scholar]
- Miner ML, Jensen JB. Decoquinate in the control of experimentally induced coccidiosis of calves. Am J Vet Res. 1976;37(9):1043–1045. [PubMed] [Google Scholar]
- Mitrovic M, Schildknecht EG. Anticoccidial activity of Lasalocid (X-537A) in chicks. Poult Sci. 1974;53(4):1448–1455. doi: 10.3382/ps.0531448. [DOI] [PubMed] [Google Scholar]
- Miyazaki Y, Shibuya M, Sugawara H, Kawaguchi O, Hirsoe C. Salinomycin, a new polyether antibiotic. J Antibiotics. 1974;27(11):814–821. doi: 10.7164/antibiotics.27.814. [DOI] [PubMed] [Google Scholar]
- Mollenhauer HH, James Morré D, Rowe LD. Alteration of intracellular traffic by monensin; mechanism, specificity and relationship to toxicity. Biochim Biophys Acta Rev Biomembr. 1990;1031(2):225–246. doi: 10.1016/0304-4157(90)90008-Z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller J, Hemphill A. Drug target identification in protozoan parasites. Expert Opin Drug Discovery. 2016;11(8):815–824. doi: 10.1080/17460441.2016.1195945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mundt HC, Daugschies A, Uebe F, Rinke M. Efficacy of toltrazuril against artificial infections with Eimeria bovis in calves. Parasitol Res. 2003;90(Suppl 3):S166–S167. doi: 10.1007/s00436-003-0929-z. [DOI] [PubMed] [Google Scholar]
- Mundt HC, Daugschies A, Wüstenberg S, Zimmermann M. Studies on the efficacy of toltrazuril, diclazuril and sulphadimidine against artificial infections with Isospora suis in piglets. Parasitol Res. 2003;90(Suppl 3):S160–S162. doi: 10.1007/s00436-003-0927-1. [DOI] [PubMed] [Google Scholar]
- Mundt HC, Bangoura B, Mengel H, Keidel J, Daugschies A. Control of clinical coccidiosis of calves due to Eimeria bovis and Eimeria zuernii with toltrazuril under field conditions. Parasitol Res. 2005;97(Suppl 1):S134–s142. doi: 10.1007/s00436-005-1457-9. [DOI] [PubMed] [Google Scholar]
- Mundt HC, Mundt-Wüstenberg S, Daugschies A, Joachim A. Efficacy of various anticoccidials against experimental porcine neonatal isosporosis. Parasitol Res. 2007;100(2):401–411. doi: 10.1007/s00436-006-0314-9. [DOI] [PubMed] [Google Scholar]
- Mundt HC, Dittmar K, Daugschies A, Grzonka E, Bangoura B. Study of the comparative efficacy of toltrazuril and diclazuril against ovine coccidiosis in housed lambs. Parasitol Res. 2009;105(Suppl 1):S141–S150. doi: 10.1007/s00436-009-1505-y. [DOI] [PubMed] [Google Scholar]
- Nachman KE, Baron PA, Raber G, Francesconi KA, Navas-Acien A, Love DC. Roxarsone, inorganic arsenic, and other arsenic species in chicken: a U.S.-based market basket sample. Environ Health Perspect. 2013;121(7):818–824. doi: 10.1289/ehp.1206245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagi MS, Mathey WJ. Interaction of Escherichia coli and Eimeria brunetti in chickens. Avian Dis. 1972;16(4):864–873. [PubMed] [Google Scholar]
- Naujokat C, Fuchs D, Opelz G. Salinomycin in cancer: a new mission for an old agent. Mol Med Rep. 2010;3(4):555–559. doi: 10.3892/mmr_00000296. [DOI] [PubMed] [Google Scholar]
- Norcross MA, Siegmund OH, Fraser CM. Amprolium for coccidiosis in cattle: a review of efficacy and safety. Vet Med Small Anim Clin. 1974;69(4):459 passim. [PubMed] [Google Scholar]
- Odden A, et al. Controlled efficacy trial confirming toltrazuril resistance in a field isolate of ovine Eimeria spp. Parasit Vectors. 2018;11(1):394. doi: 10.1186/s13071-018-2976-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oehme FW, Pickrell JA. An analysis of the chronic oral toxicity of polyether ionophore antibiotics in animals. Vet Hum Toxicol. 1999;41(4):251–257. [PubMed] [Google Scholar]
- Peek HW, Landman WJ. Higher incidence of Eimeria spp. field isolates sensitive for diclazuril and monensin associated with the use of live coccidiosis vaccination with paracox-5 in broiler farms. Avian Dis. 2006;50(3):434–439. doi: 10.1637/7486-121205R.1. [DOI] [PubMed] [Google Scholar]
- Peek HW, Landman WJM. Coccidiosis in poultry: anticoccidial products, vaccines and other prevention strategies. Vet Q. 2011;31(3):143–161. doi: 10.1080/01652176.2011.605247. [DOI] [PubMed] [Google Scholar]
- Perelman B, Abarbanel JM, Gur-Lavie A, Meller Y, Elad T. Clinical and pathological changes caused by the interaction of lasalocid and chloramphenicol in broiler chickens. Avian Pathol. 1986;15(2):279–288. doi: 10.1080/03079458608436288. [DOI] [PubMed] [Google Scholar]
- Pines M, Vlodavsky I, Nagler A, Gurwitz D. Halofuginone: from veterinary use to human therapy. Drug Dev Res. 2000;50(3–4):371–378. [Google Scholar]
- Reid WM. Progress in the control of coccidiosis with anticoccidials and planned immunization. Am J Vet Res. 1975;36(4):593–596. [PubMed] [Google Scholar]
- Reid AJ, et al. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. 2014;24(10):1676–1685. doi: 10.1101/gr.168955.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricketts AP, Glazer EA, Migaki TT, Olson JA. Anticoccidial efficacy of semduramicin in battery studies with laboratory isolates of coccidia. Poult Sci. 1992;71(1):98–103. doi: 10.3382/ps.0710098. [DOI] [PubMed] [Google Scholar]
- Riddell FG. Structure, conformation, and mechanism in the membrane transport of alkali metal ions by ionophoric antibiotics. Chirality. 2002;14(2–3):121–125. doi: 10.1002/chir.10052. [DOI] [PubMed] [Google Scholar]
- Ruiz A, et al. Control strategies using diclazuril against coccidiosis in goat kids. Parasitol Res. 2012;110(6):2131–2136. doi: 10.1007/s00436-011-2746-0. [DOI] [PubMed] [Google Scholar]
- Rychen G, et al. Safety and efficacy of COXAM® (amprolium hydrochloride) for chickens for fattening and chickens reared for laying. EFSA J. 2018;16(7):e05338. doi: 10.2903/j.efsa.2018.5338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryley JF. Lerbek, a synergistic mixture of methyl benzoquate and clopidol for the prevention of chicken coccidiosis. Parasitology. 1975;70(Part 3):377–384. doi: 10.1017/s0031182000052148. [DOI] [PubMed] [Google Scholar]
- Ryley JF, Betts MJ. Chemotherapy of chicken coccidiosis. Adv Pharmacol Chemother. 1973;11:221–293. doi: 10.1016/s1054-3589(08)60459-7. [DOI] [PubMed] [Google Scholar]
- Rypula K, Porowski M, Kaba J, Gorczykowski M, Deniz A. Effect of isosporiasis prevention with toltrazuril on long-term pig performance. Sci World J. 2012;2012:486324. doi: 10.1100/2012/486324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scala A, Demontis F, Varcasia A, Pipia AP, Poglayen G, Ferrari N, Genchi M. Toltrazuril and sulphonamide treatment against naturally Isospora suis infected suckling piglets, is there an actual profit? Vet Parasitol. 2009;163:362–365. doi: 10.1016/j.vetpar.2009.04.028. [DOI] [PubMed] [Google Scholar]
- Schuhmacher A, et al. Tiamulin and semduramicin: effects of simultaneous administration on performance and health of growing broiler chickens. Poult Sci. 2006;85(3):441–445. doi: 10.1093/ps/85.3.441. [DOI] [PubMed] [Google Scholar]
- Scribner A, et al. Synthesis and biological activity of anticoccidial agents: 2,3-diarylindoles. Bioorg Med Chem Lett. 2009;19(5):1517–1521. doi: 10.1016/j.bmcl.2009.01.001. [DOI] [PubMed] [Google Scholar]
- Segev G, Baneth G, Levitin B, Shlosberg A, Aroch I. Accidental poisoning of 17 dogs with lasalocid. Vet Rec. 2004;155(6):174–176. doi: 10.1136/vr.155.6.174. [DOI] [PubMed] [Google Scholar]
- Sharma N, Bhalla A, Varma S, Jain S, Singh S. Toxicity of maduramicin. Emerg Med J. 2005;22(12):880–882. doi: 10.1136/emj.2004.020883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirley MW, Smith AL, Blake DP (2007) Challenges in the successful control of the avian coccidia. Vaccine 26;25(30):5540-7. 10.1016/j.vaccine.2006.12.030 [DOI] [PubMed]
- Shirley MW, McDonald V, Bellatti MA (1986) Eimeria brunetti: selection and characteristics of a precocious (and attenuated) line. Avian Pathol 15(4):705-17. 10.1080/03079458608436333 [DOI] [PubMed]
- Shlosberg A, Perl S, Harmelin A, Hanji V, Bellaiche M, Bogin E, Cohen R, Markusfeld-Nir O, Shpigel N, Eisenberg Z, Furman M, Brosh A, Holzer Z, Aharoni Y. Acute maduramicin toxicity in calves. Vet Rec. 1997;140(25):643–646. doi: 10.1136/vr.140.25.643. [DOI] [PubMed] [Google Scholar]
- Shrestha A, Freudenschuss B, Jansen R, Hinney B, Ruttkowski B, Joachim A. Experimentally confirmed toltrazuril resistance in a field isolate of Cystoisospora suis. Parasit Vectors. 2017;10(1):317. doi: 10.1186/s13071-017-2257-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrestha A, Palmieri N, Abd-Elfattah A, Ruttkowski B, Pages M, Joachim A. Cloning, expression and molecular characterization of a Cystoisospora suis specific uncharacterized merozoite protein. Parasit Vectors. 2017;10(1):68. doi: 10.1186/s13071-017-2003-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shumard RF. Ovine coccidiosis; incidence, possible endotoxin, and treatment. J Am Vet Med Assoc. 1957;131(12):559–561. [PubMed] [Google Scholar]
- Smith CK, Galloway RB. Influence of monensin on cation influx and glycolysis of Eimeria tenella sporozoites in vitro. J Parasitol. 1983;69(4):666–670. [PubMed] [Google Scholar]
- Smith C, II, Strout R. Eimeria tenella: effect of narasin, a polyether antibiotic on the ultrastructure of intracellular sporozoites. Exp Parasitol. 1980;50(3):426–436. doi: 10.1016/0014-4894(80)90045-4. [DOI] [PubMed] [Google Scholar]
- Smith CK, Galloway RB, White SL. Effect of ionophores on survival, penetration, and development of Eimeria tenella sporozoites in vitro. J Parasitol. 1981;67(4):511–516. [PubMed] [Google Scholar]
- Stephan B, Rommel M, Daugschies A, Haberkorn A. Studies of resistance to anticoccidials in Eimeria field isolates and pure Eimeria strains. Vet Parasitol. 1997;69(1):19–29. doi: 10.1016/s0304-4017(96)01096-5. [DOI] [PubMed] [Google Scholar]
- Talmon P, Jager LP, de Leeuw WA, Timmer WJ. Coccidiosis in lambs: observations in the preventive use of an amprolium-containing medicated feed. Tijdschr Diergeneeskd. 1989;114(11):611–617. [PubMed] [Google Scholar]
- Tang KL, Caffrey NP, Nóbrega DB, Cork SC, Ronksley PE, Barkema HW, Polachek AJ, Ganshorn H, Sharma N, Kellner JD, Ghali WA. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: a systematic review and meta-analysis. Lancet Planet Health. 2017;1(8):e316–e327. doi: 10.1016/S2542-5196(17)30141-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor MA, Catchpole J. Review article: coccidiosis of domestic ruminants. Appl Parasitol. 1994;35(2):73–86. [PubMed] [Google Scholar]
- Tynan EJ, 3rd, Nelson TH, Davies RA, Wernau WC. The production of semduramicin by direct fermentation. J Antibiot (Tokyo) 1992;45(5):813–815. doi: 10.7164/antibiotics.45.813. [DOI] [PubMed] [Google Scholar]
- Umemura T, Nakamura H, Goryo M, Itakura C. Histopathology of monensin-tiamulin myopathy in broiler chicks. Avian Pathol. 1984;13(3):459–467. doi: 10.1080/03079458408418547. [DOI] [PubMed] [Google Scholar]
- Umemura T, Nakamura H, Goryo M, Itakura C. Ultrastructural changes of monensin-oleandomycin myopathy in broiler chicks. Avian Pathol. 1984;13(4):743–751. doi: 10.1080/03079458408418571. [DOI] [PubMed] [Google Scholar]
- Verheyen A, Maes L, Coussement W, Vanparijs O, Lauwers F, Vlaminckx E, Borgers M, Marsboom R. In vivo action of the anticoccidial diclazuril (Clinacox) on the developmental stages of Eimeria tenella: an ultrastructural evaluation. J Parasitol. 1988;74(6):939–949. [PubMed] [Google Scholar]
- Verheyen A, Maes L, Coussement W, Vanparijs O, Lauwers F, Vlaminckx E, Marsboom R. Ultrastructural evaluation of the effects of diclazuril on the endogenous stages of Eimeria maxima and E. brunetti in experimentally inoculated chickens. Parasitol Res. 1989;75(8):604–610. doi: 10.1007/BF00930956. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G, et al. Clinical and epidemiological characteristics of Eimeria infections in first-year grazing cattle. Vet Parasitol. 2006;136(3–4):215–221. doi: 10.1016/j.vetpar.2005.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang CC. Studies of the mitochondria from Eimeria tenella and inhibition of the electron transport by quinolone coccidiostats. Biochim Biophys Acta. 1975;396(2):210–219. doi: 10.1016/0005-2728(75)90035-3. [DOI] [PubMed] [Google Scholar]
- Wang CC. Inhibition of the respiration of Eimeria tenella by quinolone coccidiostats. Biochem Pharmacol. 1976;25(3):343–349. doi: 10.1016/0006-2952(76)90225-2. [DOI] [PubMed] [Google Scholar]
- Wang CC (1978) Biochemical and nutritional aspects of coccidia. Avian Coccidiosis, pp 135–184
- Wang CC. Biochemistry and physiology of coccidia. p167–228 in: The biology of the coccidia. P.L. Long. Baltimore: University Park Press; 1982. [Google Scholar]
- Wang Z, Suo X, Xia X, Shen J. Influence of monensin on cation influx and Na+-K+-ATPase activity of Eimeria tenella sporozoites in vitro. J Parasitol. 2006;92(5):1092–1096. doi: 10.1645/GE-783R.1. [DOI] [PubMed] [Google Scholar]
- WHO (2017) WHO guidelines on use of medically important antimicrobials in food-producing animals. ISBN: 978-92-4-155,013-0. https://www.who.int/foodsafety/publications/cia_guidelines/en/ [PubMed]
- Williams RB. The mode of action of anticoccidial quinolones (6-decyloxy-4-hydroxyquinoline-3-carboxylates) in chickens. Int J Parasitol. 1997;27(1):101–111. doi: 10.1016/s0020-7519(96)00156-7. [DOI] [PubMed] [Google Scholar]
- Williams RB. A compartmentalised model for the estimation of the cost of coccidiosis to the world’s chicken production industry. Int J Parasitol. 1999;29(8):1209–1229. doi: 10.1016/s0020-7519(99)00086-7. [DOI] [PubMed] [Google Scholar]
- Williams RB. Tracing the emergence of drug-resistance in coccidia (Eimeria spp.) of commercial broiler flocks medicated with decoquinate for the first time in the United Kingdom. Vet Parasitol. 2006;135(1):1–14. doi: 10.1016/j.vetpar.2005.10.012. [DOI] [PubMed] [Google Scholar]
- Wong DT, Horng JS, Wilkinson JR. Robenzidene, an inhibitor of oxidative phosphorylation. Biochem Biophys Res Commun. 1972;46(2):621–627. doi: 10.1016/s0006-291x(72)80185-2. [DOI] [PubMed] [Google Scholar]
- Yoder CA, Graham JK, Miller LA. Molecular effects of nicarbazin on avian reproduction. Poult Sci. 2006;85(7):1285–1293. doi: 10.1093/ps/85.7.1285. [DOI] [PubMed] [Google Scholar]
- Zaionts VI, Krylov MV, Loskot VI, Kirillov AI. Biosynthesis of folic acid in Eimeria tenella (Coccidia) Parazitologiia. 1978;12(1):3–8. [PubMed] [Google Scholar]
- Zhang DF, et al. Anticoccidial effect of halofuginone hydrobromide against Eimeria tenella with associated histology. Parasitol Res. 2012;111(2):695–701. doi: 10.1007/s00436-012-2889-7. [DOI] [PubMed] [Google Scholar]
- Zhou BH, Wang HW, Wang XY, Zhang LF, Zhang KY, Xue FQ. Eimeria tenella: effects of diclazuril treatment on microneme genes expression in second-generation merozoites and pathological changes of caeca in parasitized chickens. Exp Parasitol. 2010;125(3):264–270. doi: 10.1016/j.exppara.2010.01.022. [DOI] [PubMed] [Google Scholar]
- Zhou BH, Wang HW, Zhao ZS, Liu M, Yan WC, Zhao J, Zhang Z, Xue FQ. A novel serine/threonine protein phosphatase type 5 from second-generation merozoite of Eimeria tenella is associated with diclazuril-induced apoptosis. Parasitol Res. 2013;112(4):1771–1780. doi: 10.1007/s00436-013-3336-0. [DOI] [PubMed] [Google Scholar]