

Abstract
Activity-dependent protein synthesis at synapses is dysregulated in the Fragile X syndrome (FXS). This process contributes to dendritic spine dysmorphogenesis and synaptic dysfunction in FXS. Matrix Metalloproteinase 9 (MMP-9) is an enzyme involved in activity-dependent reorganization of dendritic spine architecture and was shown to regulate spine morphology in a mouse model of FXS, the Fmr1 knock-out mice. Here we show that MMP-9 mRNA is part of the FMRP complex and colocalizes in dendrites. In the absence of FMRP MMP-9 mRNA translation is increased at synapses, suggesting that this mechanism contributes to the increased metalloproteinase level at synapses of Fmr1 knock-out mice. We propose that such a local effect can contribute to the aberrant dendritic spine morphology observed in the Fmr1 knock-out mice and in patients with FXS.
Introduction
The Fragile X syndrome (FXS) is the most common form of inherited intellectual disability and also the most common known single gene cause of autism (Hagerman et al., 2005; Bagni et al., 2012). One of the characteristic features of FXS neurons is an abnormal dendritic spine morphology (Rudelli et al., 1985). Immature, long and thin dendritic spines are also observed in the Fmr1 knock-out (KO) mouse, a model of FXS (Comery et al., 1997). FXS results from the loss of FMRP, an RNA binding protein largely characterized for its role in regulating synaptic protein synthesis (Bassell and Warren, 2008; Bhakar et al., 2012; Darnell and Klann, 2013).
FMRP controls distinct aspects of mRNA metabolism in neurons, such as dendritic transport and translation at synapses or stability (Bagni et al., 2012). FMRP associates with a subset of mRNAs and is transported in a stimulus-dependent manner (Antar et al., 2004). At the synapse, it can regulate translation in at least two different levels: initiation through the neuronal eIF4E-BP CYFIP1 (Napoli et al., 2008) or other eIF4E-BPs (Sharma et al., 2010) and elongation through reversible stalling of ribosomes (Darnell et al., 2011). Local protein synthesis is elevated in Fmr1 KO mice and, importantly, it cannot be regulated by neuronal stimulation (Zalfa et al., 2003; Lu et al., 2004; Muddashetty et al., 2007). A number of FMRP target mRNAs identified to date encode for postsynaptic and presynaptic proteins, structural and scaffolding proteins, neurotransmitter receptors, and signaling molecules (Darnell et al., 2011).
In a recent study, it was shown that Matrix Metalloproteinase 9 (MMP-9) levels were increased in a mouse model for FXS and that reduction of MMP-9 by the antibiotic minocycline was sufficient to rescue dendritic spine dysmorphogenesis in vitro and in vivo (Bilousova et al., 2009). This effect was accompanied by behavioral improvement in young mice and ameliorated performance in general cognition, activity, and anxiety tests (Bilousova et al., 2009). This is an intriguing observation considering that MMP-9 is involved in activity-dependent reorganization of dendritic spine architecture (Michaluk et al., 2007, 2011; Dityatev et al., 2010). On the other hand, application of MMP-9 onto cultured neurons induces formation of filopodia-like immature dendritic spines that resemble FXS spines (Michaluk et al., 2011). Based on those findings clinical trials to test minocycline have been undertaken and there is evidence that minocycline provides some functional benefits to FXS patients (Paribello et al., 2010; Utari et al., 2010; Leigh et al., 2013). However, how absence of FMRP results in an increase of MMP-9 has never been investigated.
Recently, we have reported that MMP-9 mRNA is transported to the dendrites and locally translated in the synaptodendritic compartment after neuronal stimulation (Dziembowska et al., 2012). Here, we show that FMRP is involved in transport of MMP-9 mRNA to the synapses where it regulates its translation. Increased translation of the MMP-9 mRNA in the Fmr1 KO mice leads to excessive protein synthesis at the synapse. Since MMP-9 activity has an effect on the processing of extracellular matrix as well as proper function of adhesion molecules (Dziembowska and Wlodarczyk, 2012), we propose that increased MMP-9 levels in FXS can partially explain the dendritic spine dysmorphology observed in this syndrome.
Materials and Methods
Animals.
We have used 1- to 2-month-old Fmr1 KO mice on FVB background with age-matched FVB wild-type (WT) mice. Before the experiment, the animals were kept in the laboratory animal facility with ad libitum access to food and water with a 12 h light/dark cycle.
Gel zymography.
The gel zymography on synaptoneurosomes isolated from cortices and hippocampi of Fmr1 KO and WT mice (of either sex) was performed according to Szklarczyk et al. (2002). Protein concentrations were measured using BCA protein assay (Pierce). Synaptoneurosomes were lysed in 2× sample buffer and subjected to electrophoresis in SDS-PAGE Tris-glycine 8% acrylamide gels containing 0.5% gelatin (POCH) under nondenaturating, nonreducing conditions. Gels were washed twice for 30 min in 2.5% Triton X-100 and incubated in the zymography buffer (50 mm Tris, pH 7.5, 10 mm CaCl2, 1 μm ZnCl2, 1% Triton X-100, and 0.02% sodium azide) for 5 d at 37°C. Gels were stained with 0.5% Coomassie.
RNA coimmunoprecipitation.
Immunoprecipitation was performed according to the modified protocol of Brown et al. (2001). Synaptoneurosomes from WT and Fmr1 KO mice of either sex (1800 μg of total protein) were resuspended in 1200 μl of precipitation buffer (10 mm HEPES, pH 7.4, 400 mm NaCl, 30 mm EDTA, and 0.5% Triton X-100) supplemented with protease inhibitor cocktail (Sigma-Aldrich) and 100 U/ml RiboLock (Fermentas), then precleared with 180 μl of Dynabeads A for 2.5 h. Afterward, 1/3 of each supernatant was saved as input fraction for WB and RNA isolation. Precipitation was performed overnight in 4°C with 120 μl of antibody-bound Dynabeads Protein A, blocked beforehand with either anti-FMRP antibody (7G1–1 or rAM2) or normal mouse IgGs. Next, 1/6 of the beads was boiled with sample buffer for WB. From the remaining beads, total RNA was isolated with TRIzol (Invitrogen).
For the quantitative real-time (qRT)-PCR, RNA was suspended in 11 μl of RNase free H2O and its concentration was determined by NanoDrop. External control “spike” mRNA (LSM gene from Arabidopsis thaliana) was added into the reaction mix (1 pg per reaction). Then the RNA was reverse-transcribed using random hexamer primers (Fermentas).
Reverse transcription PCR and qRT-PCR.
RNA isolated from synaptoneurosomes, prepared from WT and Fmr1 KO female mice, was reverse transcribed using SuperScript III Reverse Transcriptase (Invitrogen) and random hexamer primers. Real-time PCR was performed using SYBR Green PCR Master Mix (Applied Biosystems) and primers: MMP-9F:gcgctgggcttagatcattc, MMP-9R:agccacgaccatacagatgc,αCaMKIIF:gcctgaagaagttcaatgcc, αCaMKIIR:ctcaatggtggtgttggtgc, PSD95(Dlg4)F:tgagctatgagacggtgacg, PSD95(Dlg4)R:cgcttaggacgtgtcgtatg, beta2mF:ctgaccggcctgtatgctat, beta2mR:cagtctcagtgggggtgaat. LSM(spike)F: tcttctctctccgtgtcca R:tgatcaattcgccaatgcg. In the coimmunoprecipitation experiment, for both Input fraction and immunoprecipitated RNA, TaqMan primer/probe set for MMP-9 (Applied Biosystems, Mm00442991_m1) was used. Values were calculated according to the ddCT method, using Input WT as a normalizer and spike mRNA as an external control.
3,4-dihydroxyphenylglycol stimulation, in situ hybridization, and immunostaining.
Primary rat hippocampal neurons (14 d in vitro (DIV)) were pulse stimulated with 50 μm 3,4-dihydroxyphenylglycol (DHPG) for 5 min, medium was then washed out, replaced with the conditioned medium, and cells were fixed at different time points at 0, 10, and 20 min. In situ hybridization and immunostaining with rabbit anti-FMR1 (sc-28739; Santa Cruz Biotechnology) were performed according to Dziembowska et al. (2012). We have counted MMP-9 mRNA-positive granules per 50 μm of dendrite in the minimum distance of 10 μm from the cell body and we have assayed their colocalization with FMRP.
Synaptoneurosomes and polysomes-mRNP assay.
Synaptoneurosomes prepared from cortices and hippocampi of 1- to 2-month-old WT and Fmr1 KO female mice as described previously (Dziembowska et al., 2012) were stimulated with 100 μm DHPG (Tocris Bioscience) for 15 min at 37°C. Next, synaptoneurosomes were lysed using buffer A (20 mm Tris-HCl, pH 7.4, 125 mm NaCl, 5 mm MgCl2, 2 mm dithiothreitol, 100 μg/ml cycloheximide, and protease and RNase inhibitors) containing 1.5% Triton X-100. Resulting supernatant was loaded on a 10–50% linear sucrose gradient prepared in buffer A and spun at 38,000 rpm for 2 h in a Beckman SW-41 rotor. Each gradient was separated into 11 fractions. Before the RNA extraction spike mRNA, an in vitro transcribed fragment of A. thaliana LSM gene was added to each of the fractions. RNA was next isolated by TRIzol (Invitrogen) and mRNAs of interest were analyzed by quantitative radioactive RT-PCR. Puromycin (Sigma) was added to synaptoneurosomes in 2 mm concentration for 30 min.
Western blotting.
Western blotting was performed as before (Dziembowska et al., 2012). Antibodies used were as follows: rabbit anti-FMRP H-120 (sc-28739; Santa Cruz Biotechnology) or D14F4 (#7104; Cell Signaling Technology), anti-ribosomal protein L13 (sc-98525; Santa Cruz Biotechnology), anti-GAPDH (MAB374; Millipore), anti-GFAP (sc-166458; Santa Cruz Biotechnology), and anti-β-DG (NCL-b-DG; Novocastra).
Results
MMP-9 protein levels are increased at synapses from Fmr1 KO mice
To investigate if the increase of MMP-9 protein level in the brain from the Fmr1 KO mice (Bilousova et al., 2009) was also detected at synapses, we studied the presence of MMP-9 gelatinolytic activity in synaptoneurosomes isolated from cortices and hippocampi of either WT or Fmr1 KO mice by gel zymography. As shown in Figure 1A, synaptoneurosomes (SN) from Fmr1 KO mice show an increase of MMP-9 activity. The enrichment in synaptoneurosomes was evaluated by Western blotting. In the SN fraction synaptic proteins such as postsynaptic density protein (PSD)-95 and β-dystroglycan (β-DG) were detected whereas the glia-expressed Glial fibrillary acidic protein (GFAP) was strongly reduced (Fig. 1B).
Figure 1.

MMP-9 level is increased in synaptoneurosomes from Fmr1 KO mice. A, Gel zymography on protein extracts from synaptoneurosomes prepared from WT and Fmr1 KO. The activity of MMP-9 was quantified by densitometry. ***p < 0.001, n = 7 males per group, unpaired t test. Error bars indicate SEM. B, Characterization of protein content of synaptoneurosomal fraction isolated from WT and Fmr1 KO mice. Crude homogenate (H), filtrate (F), synaptoneurosomes (SN), and supernatant (cytosolic, C) fractions were analyzed by Western blotting. Synaptoneurosomes were analyzed for the synaptic protein PSD-95, the transmembrane protein β-DG and the glial protein GFAP. Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) was a loading control.
FMRP interacts with MMP-9 mRNA in mouse synaptoneurosomes
In silico analysis of the murine MMP-9 mRNA sequence revealed a set of paired Gs within 3′UTR of the transcript. Paired Gs forming an intramolecular G-quartet, or G-rich sequences (Zalfa et al., 2007), are RNA sequence motifs bound by FMRP (Darnell et al., 2001; Schaeffer et al., 2001; Subramanian et al., 2011). To investigate whether MMP-9 mRNA interacts with FMRP in the brain, we performed RNA coimmunoprecipitation on synaptoneurosomes isolated from the cerebral cortex and hippocampi of WT and Fmr1 KO mice. As shown in Figure 2A, FMRP was precipitated by the 7G1–1 anti-FMRP antibody (Brown et al., 2001) from synaptoneurosomes prepared from WT hippocampi and cortex, while it was not detected from the Fmr1 KO immunoprecipitates (IPs). Next, we performed RT quantitative-PCR on both input and immunoprecipitated fractions, to assess levels of MMP-9 mRNA and control mRNAs. The mRNA levels of MMP-9, PSD-95, and β-2-microglobulin (B2M) mRNAs were not significantly different between WT and Fmr1 KO mice (Fig. 2B). On the contrary, MMP-9 mRNA was significantly enriched in the IPs from the WT mice compared with the Fmr1 KO. PSD-95 mRNA, a known target of FMRP (Muddashetty et al., 2007; Zalfa et al., 2007), was also significantly enriched in the IPs from the WT mice when compared with the Fmr1 KO. B2M mRNA levels did not differ between both IP fractions, further supporting the specificity of the MMP-9 mRNA-FMRP interaction (Fig. 2C). The coimmunoprecipitation was also performed independently with a different anti FMRP antibody (Ferrari et al., 2007) confirming these results (Fig. 2D,E).
Figure 2.
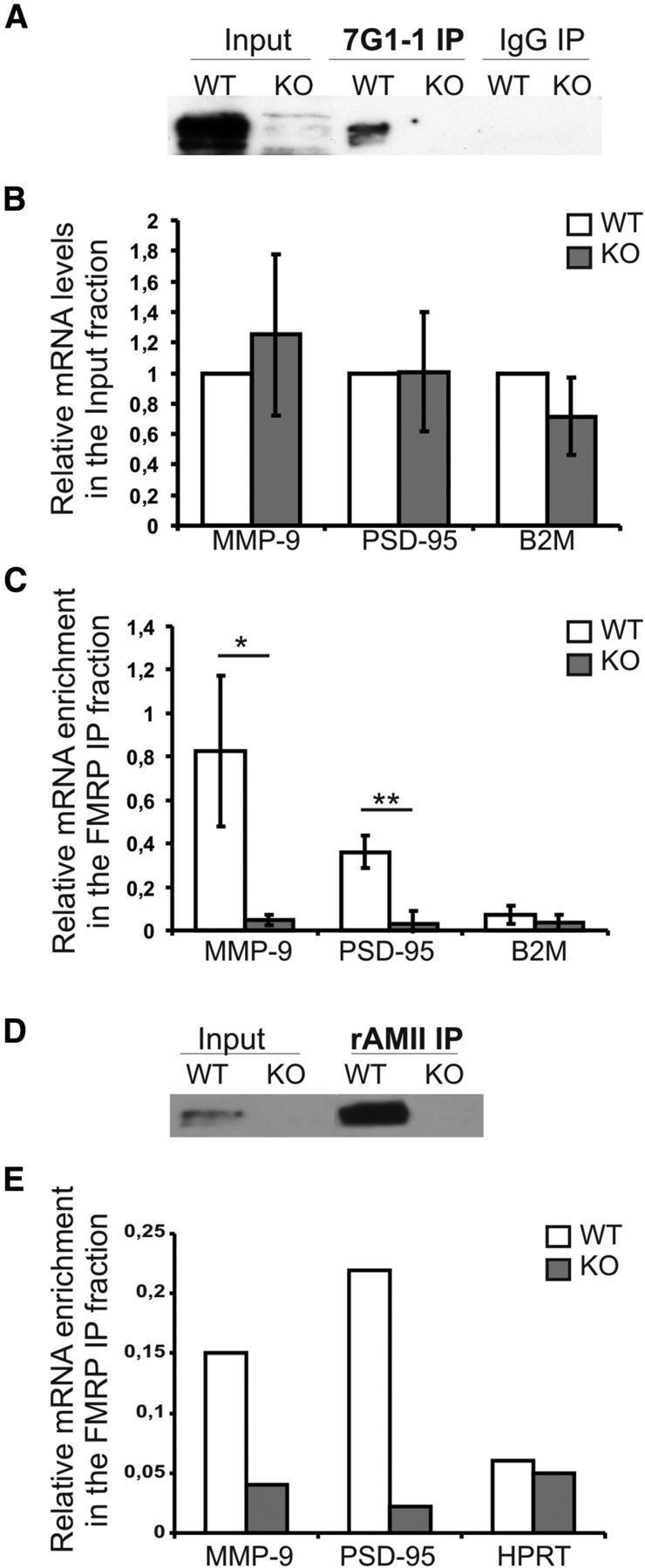
FMRP coprecipitates with MMP-9 mRNA in mouse synaptoneurosomes. A, Western blot analysis of the immunoprecipitated FMRP from mouse synaptoneurosomes shows FMRP precipitated by the anti-FMRP 7G1–1 (Brown et al., 2001) antibody in WT mice. Fmr1 KO extracts as well as IgG IPs were used as negative controls. B, Steady-state levels analyzed by RT-qPCR of PSD-95, MMP-9, and B2M mRNAs in synaptoneurosomes from WT and Fmr1 KO mice. p > 0.03, n = 3, one-sample t test. Values were normalized to WT. C, RT-qPCR analysis of mRNAs immunoprecipitated by anti-FMRP antibody. MMP-9, PSD-95, and B2M (negative control) mRNAs detected in the WT over Fmr1 KO IPs. Values were normalized to the external spike control gene. Error bars indicate SD, n = 3, *p < 0.03, **p < 0.01 by Student's t test. D, E, Same as in A and C using a different FMRP antibody: rAM2 antibody (Ferrari et al., 2007) analyzing PSD-95, MMP-9, and hypoxanthine phosphoribosyltransferase (HPRT; negative control) mRNAs, n = 1.
FMRP and MMP-9 mRNA colocalize in dendrites and dissociate after mGluR stimulation
FMRP can act as an adaptor for kinesin light chain to promote stimulus-induced transport of specific mRNAs (Antar et al., 2005; Ferrari et al., 2007; Charalambous et al., 2013). Therefore we have next investigated whether MMP-9 mRNA is transported to the dendrites in FMRP-containing granules. To address this question, we first studied the colocalization of MMP-9 mRNA and FMRP in the dendrites of cultured hippocampal neurons (19—21 DIV). To directly visualize MMP-9 mRNA and FMRP in the dendrites, we transfected neurons with an MS2-GFP reporter system (Zenklusen et al., 2007). This system consists of three plasmids, one contains 24 MS2 binding sites fused with MMP-9 coding sequence and its 3′UTR (pMS2-BS-MMP9), containing the G-rich sequence possibly required for its dendritic localization (Subramanian et al., 2011). The second plasmid contains GFP fused with the MS2 protein able to bind the 24 MS2 binding sites (pMS2-GFP). The third plasmid contains FMRP-mCherry.
Neurons were cotransfected with pMS2-BS-MMP-9; pMS2-GFP and FMRP-mCherry, the latter expressing red-FMRP fusion protein. As shown in Figure 3A, GFP-labeled MMP-9 mRNA granules colocalize with FMRP-mCherry in dendrites consistent with the data generated upon immunoprecipitation. Several controls were performed to address the specificity of our findings: neurons were either transfected with FMRP-mCherry with the pMS2-GFP plasmid or with FMRP-mCherry and pMS2-BS-vector without MMP-9 mRNA sequence. GFP showed uniform diffuse staining in the controls and no colocalization with red fluorescent FMRP protein was detected (Fig. 3B).
Figure 3.
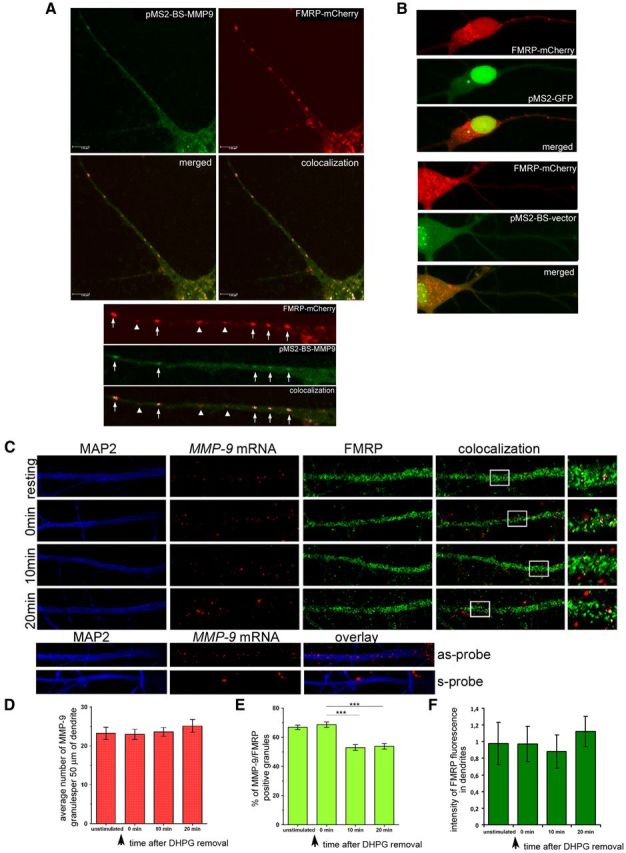
MMP-9 mRNA colocalizes with FMRP in dendrites and dissociates from FMRP-containing granules after mGluR stimulation. A, Hippocampal neurons were cotransfected with the MS2-GFP reporter system pMS2-BS-MMP9 to visualize MMP-9 mRNA (green) and FMRP-mCherry (red); the colocalization is indicated by white arrows. B, Control transfections of hippocampal neurons. FMRP-mCherry and pMS2-GFP vector; FMRP-mCherry and pMS2-BS-vector. C, In situ hybridization combined with immunofluorescence shows the MMP-9 mRNA (red) colocalization with FMRP (green) in primary dendrites of hippocampal neurons stained with MAP2 (blue). The insert is an enlarged area that shows colocalization (white granules). In situ hybridization performed with sense (s-probe) or antisense (as-probe) probe (bottom). D, E, Quantification of FMRP and MMP-9 mRNA granules. Fifty micrometer dendrite segments before (D) or after DHPG treatment (E) (10 and 20 min). ***p < 0.001, nonparametric ANOVA with Dunn's post-test. Data were analyzed from at least 10 dendrites in each group from four independent experiments. Error bars indicate SEM. F, As in D and E measuring the fluorescence intensity detected with anti-FMRP antibody. Error bars indicate SD.
To investigate if neuronal stimulation causes a change in the association of MMP-9 mRNA with FMRP, hippocampal neurons were pulse stimulated with a Gp1 agonist of mGluRs (DHPG) for 5 min, the cell culture medium was then washed out, replaced with the conditioned medium, and cells were fixed at 10 and 20 min. Endogenous MMP-9 mRNA was revealed by in situ hybridization combined with immunodetection of FMRP. MMP-9 mRNA was localized in granules along the dendrites and exhibited abundant colocalization with FMRP (Fig. 3C). The number of double-positive MMP-9 mRNA and FMRP granules was counted in control conditions and after stimulation with DHPG. We observed a significant decrease in the number of double-positive (MMP-9 and FMRP) granules, 10 and 20 min after DHPG stimulation, whereas the total number of MMP-9 granules and the intensity of fluorescence detected in the dendrites with the anti-FMRP antibody did not change (Fig. 3C–F). The percentage of MMP-9 mRNA-positive granules, which colocalized with FMRP protein in WT resting neurons, was 66.8% (SEM = 1.5) for unstimulated and 68.6 (SEM = 1.9). After mGluR stimulation with DHPG (10–20 min) we observed a significant decrease in the number of MMP-9-positive granules that colocalized with FMRP: 10 min after DHPG 52.9% (SEM = 2.0), 20 min after DHPG 53.8% (SEM = 1.9).
Analysis of the polysomes-mRNP distribution of MMP-9 mRNA at synapses
To investigate if MMP-9 mRNA is efficiently delivered to the synapses of Fmr1 KO mice, we detected the level of MMP-9 mRNA, as well as other dendritically localized FMRP target mRNAs, calcium/calmodulin-dependent protein kinase II alpha (αCaMKII) and postsynaptic density protein PSD-95, in synaptoneurosomes isolated from cortices and hippocampi of WT and Fmr1 KO mice (Hou et al., 2006; Muddashetty et al., 2007; Zalfa et al., 2007). No significant difference in the level of MMP-9 mRNA was detected between WT and Fmr1 KO synaptoneurosomes (Fig. 4A). Considering the elevated level of MMP-9 protein detected in the synaptoneurosomes of Fmr1 KO mice (Fig. 1A), we hypothesized that a deregulated local synthesis of MMP-9 occurs in the Fmr1 KO mice at synapses.
Figure 4.

Translational regulation of MMP-9 mRNA at synapses. A, Quantitative analysis (RT-qPCR) of MMP-9, PSD-95, and αCaMKII mRNA levels in synaptoneurosomes isolated from WT and Fmr1 KO mice (unpaired t test, p > 0.05; n = 4 males per group). B, Polysome-mRNP distribution of MMP-9 mRNA in WT and Fmr1 KO synaptoneurosomes. Radioactive RT-PCR showing the level of MMP-9 mRNA and control (LSM) in five fractions without and after DHPG stimulation. Polysomal incorporation of MMP-9 mRNA was compared between WT and Fmr1 KO synaptoneurosomal fractions in basal and DHPG stimulated conditions. The distribution of MMP-9 mRNA across the fractions is expressed as a percentage of mRNA in each fraction. n = 3 for each group; **p < 0.01, *p < 0.03, two-way ANOVA with Fisher's LSD post hoc test. Histograms represent mean values ± SEM. C, The effect of puromycin on the MMP-9 mRNA distribution in the gradient fractions (left). Western blotting with anti-ribosomal protein L13 on protein extracts prepared from the gradient fractions (right).
To investigate if local production of MMP-9 protein is regulated by FMRP, we analyzed the polysome-mRNP profile of MMP-9 mRNA in synaptoneurosomes from WT and Fmr1 KO mice under basal and stimulated conditions. Lysed synaptoneurosomes were separated by ultracentrifugation on linear sucrose gradient. Collected fractions were divided into five groups, which correspond, from the top to the bottom of the gradient, to free messenger ribonucleoprotein complexes (mRNP), monosomal fraction, light polysomes, heavy polysomes, corresponding to the actively translating polyribosomal fraction, and RNA granules (Dziembowska et al., 2012). Total RNA was extracted from the gradient fractions and then analyzed by nonsaturating radioactive RT-PCR. For normalization the same amount of a spike mRNA was added to each fraction before RNA extraction. After DHPG stimulation of WT synaptoneurosomes MMP-9 mRNA shifted to actively translating heavy polyribosomal fraction, from ∼30 to ∼40% (Fig. 4B), in agreement with the fine-tuned local protein synthesis. Fmr1 KO mice showed instead already 45% of MMP-9 mRNA associated with heavy polysomal fraction in basal status. After DHPG stimulation we could not observe an increase of MMP-9 mRNA translation in synaptoneurosomes from Fmr1 KO, indicating a lack of response to synaptic stimulation. These findings suggest that absence of FMRP leads to increased basal translation and impaired activity-mediated translation of MMP-9 mRNA at the synapse.
To reveal the fractions that contain polysomes actively translating MMP-9 mRNA we studied the effect of puromycin on MMP-9 mRNA distribution along the gradient. Puromycin is an amino-acyl tRNA analog that inhibits active translation on polyribosomes. Synaptoneurosomes from WT mice brains were stimulated with 100 μm DHPG and then treated with 2 mm puromycin or 100 μm cycloheximide (CHX). Puromycin treatment resulted in a shift of MMP-9 mRNA from the heavy and light polysomal fractions to the fraction containing monosomes (translationally silent; Fig. 4C). The cosedimentation of the ribosomal protein rpL13 detected by Western blot shifted after puromycin treatment from heavy to lighter polysomal fractions (Fig. 4C, right).
Discussion
In the present study we identify MMP-9 mRNA as a specific target of FMRP and provide evidence that FMRP regulates its dendritic translation at the synapse. Our data support a model in which synaptic MMP-9 is translationally regulated by FMRP. In the case of FXS the lack of FMRP leads to excessive synaptic synthesis of MMP-9 protein. We propose that such a local effect can contribute to the aberrant dendritic spine morphology observed in patients with FXS.
Recently 842 FMRP target transcripts have been identified by the CLIP method (Darnell et al., 2011). However, MMP-9 mRNA was not identified in this approach, probably due to its low abundance in neurons when compared with other transcripts. Also in a second study, which identified a larger number of FMRP target mRNAs, MMP-9 was not detected (Ascano et al., 2012). Of note, in a recent study performed in cancer cells MMP9 mRNA appears as a strong FMRP target (Lucá et al., 2013).
Our work in neurons shows that MMP-9 mRNA is detected at low levels in the dendrites compared with Arc and its level increases after long-term potentiation induction (Dziembowska et al., 2012).
Enhanced MMP-9 activity has been described in hippocampal extracts of Fmr1 KO mice (Bilousova et al., 2009). In agreement with those findings we also observed an increase in MMP-9 activity in the cerebral cortex and more importantly in synaptoneurosomes isolated from the Fmr1 KO mice (Fig. 1).
Sequence analysis of the 3′UTR of MMP-9 mRNA revealed the presence of a G-rich sequence, similar to that found in several FMRP-associated mRNAs, such as MAP1b (Darnell et al., 2011) and PSD-95 (Zalfa et al., 2007). RNA-IP with two independent FMRP antibodies confirmed the presence of the MMP-9 mRNA in the complex with FMRP (Fig. 2). These findings were further supported by colocalization of the MMP-9 mRNA with FMRP in the dendrites (Fig. 3A). However, to confirm direct interaction between FMRP and MMP-9 mRNA further experiments are needed.
In an earlier study Kao et al. (2010) show that FMRP can mediate spatial mRNA delivery for local protein synthesis in response to synaptic stimulation. After DHPG stimulation they found that the total number of CaMKIIα mRNA granules in the dendrite did not change; however, the fraction of mRNA localized in dendritic spine versus dendrite increased (Kao et al., 2010). We also observed no change in the number of MMP-9 mRNA granules in the dendrite after DHPG stimulation, but the number of MMP-9 mRNA and FMRP colocalizing granules decreases from 66.8% (SEM = 1.5) in unstimulated and 68.6 (SEM = 1.9) in control neurons to 52.9% (SEM = 2.0) 10 min and 53.8% (SEM = 1.9) 20 min after DHPG stimulation (Fig. 3). RNA granules can disassemble in response to neuronal activity, which leads to release of translationally silenced mRNAs and their association into actively translating polysomes (Krichevsky and Kosik, 2001). This would allow MMP-9 mRNA to dissociate from its transporting, translationally silent FMRP-mRNP and to be loaded on the polysomes.
Previous studies indicated that mRNAs transported in FMRP containing mRNPs (such as αCaMKII, PSD-95, GluR1, and NR1) can be properly delivered to the synapse of Fmr1 KO mice since their level in synaptoneurosomes is not different from in WT mice. (Muddashetty et al., 2007). In agreement with previously published work, our data show that dendritic transport of MMP-9 mRNA does not decrease in the Fmr1 KO mice (Fig. 4A).
It is well established that one of the major consequences of FMRP loss is deregulated synaptic protein synthesis (Bassell and Warren, 2008; Bhakar et al., 2012). Here, we show impaired mGluR-dependent translation of MMP-9 in synaptoneurosomes from Fmr1 KO mice. Stimulation with DHPG leads to significantly increased association of MMP-9 mRNA with polyribosomes. In the case of Fmr1 KO mice, the fraction of MMP-9 cosedimenting with the polysomes does not change following DHPG stimulation (Fig. 4B). This is consistent with previous studies that show other transcripts regulated by FMRP behave in the same way (Zalfa et al., 2003; Muddashetty et al., 2007).
Activity-dependent local translation of MMP-9 mRNA at the synapse is important for structural and functional dendritic spine remodeling. Our findings for dysregulated synaptic translation of MMP-9 mRNA in the Fmr1 KO explain higher activity of MMP-9 at the synapse of Fmr1 KO mice contributing to the aberrant morphology of FXS dendritic spines (Michaluk et al., 2011). Hence, our data further support clinical trials aimed at reducing MMP-9 activity in FXS.
Footnotes
This work was supported by NCN Harmonia grant 2013/08/M/NZ4/00650 to M.D.; Telethon GGP10150, Compagnia San Paolo, Associazione Italiana Sindrome X Fragile, and VIB to C.B.; and Foundation for Polish Science, FNP TEAM to L.K.
References
- Antar LN, Afroz R, Dictenberg JB, Carroll RC, Bassell GJ. Metabotropic glutamate receptor activation regulates fragile x mental retardation protein and FMR1 mRNA localization differentially in dendrites and at synapses. J Neurosci. 2004;24:2648–2655. doi: 10.1523/JNEUROSCI.0099-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antar LN, Dictenberg JB, Plociniak M, Afroz R, Bassell GJ. Localization of FMRP-associated mRNA granules and requirement of microtubules for activity-dependent trafficking in hippocampal neurons. Genes Brain Behav. 2005;4:350–359. doi: 10.1111/j.1601-183X.2005.00128.x. [DOI] [PubMed] [Google Scholar]
- Ascano M, Jr, Mukherjee N, Bandaru P, Miller JB, Nusbaum JD, Corcoran DL, Langlois C, Munschauer M, Dewell S, Hafner M, Williams Z, Ohler U, Tuschl T. FMRP targets distinct mRNA sequence elements to regulate protein expression. Nature. 2012;492:382–386. doi: 10.1038/nature11737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagni C, Tassone F, Neri G, Hagerman R. Fragile X syndrome: causes, diagnosis, mechanisms, and therapeutics. J Clin Invest. 2012;122:4314–4322. doi: 10.1172/JCI63141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassell GJ, Warren ST. Fragile X syndrome: loss of local mRNA regulation alters synaptic development and function. Neuron. 2008;60:201–214. doi: 10.1016/j.neuron.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhakar AL, Dölen G, Bear MF. The pathophysiology of fragile X (and what it teaches us about synapses) Annu Rev Neurosci. 2012;35:417–443. doi: 10.1146/annurev-neuro-060909-153138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bilousova TV, Dansie L, Ngo M, Aye J, Charles JR, Ethell DW, Ethell IM. Minocycline promotes dendritic spine maturation and improves behavioural performance in the fragile X mouse model. J Med Genet. 2009;46:94–102. doi: 10.1136/jmg.2008.061796. [DOI] [PubMed] [Google Scholar]
- Brown V, Jin P, Ceman S, Darnell JC, O'Donnell WT, Tenenbaum SA, Jin X, Feng Y, Wilkinson KD, Keene JD, Darnell RB, Warren ST. Microarray identification of FMRP-associated brain mRNAs and altered mRNA translational profiles in fragile X syndrome. Cell. 2001;107:477–487. doi: 10.1016/S0092-8674(01)00568-2. [DOI] [PubMed] [Google Scholar]
- Charalambous DC, Pasciuto E, Mercaldo V, Pilo Boyl P, Munck S, Bagni C, Santama N. KIF1Bbeta transports dendritically localized mRNPs in neurons and is recruited to synapses in an activity-dependent manner. Cell Mol Life Sci. 2013;70:335–356. doi: 10.1007/s00018-012-1108-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Comery TA, Harris JB, Willems PJ, Oostra BA, Irwin SA, Weiler IJ, Greenough WT. Abnormal dendritic spines in fragile X knockout mice: maturation and pruning deficits. Proc Natl Acad Sci U S A. 1997;94:5401–5404. doi: 10.1073/pnas.94.10.5401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darnell JC, Klann E. The translation of translational control by FMRP: therapeutic targets for FXS. Nat Neurosci. 2013;16:1–7. doi: 10.1038/nn.3379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darnell JC, Jensen KB, Jin P, Brown V, Warren ST, Darnell RB. Fragile X mental retardation protein targets G quartet mRNAs important for neuronal function. Cell. 2001;107:489–499. doi: 10.1016/S0092-8674(01)00566-9. [DOI] [PubMed] [Google Scholar]
- Darnell JC, Van Driesche SJ, Zhang C, Hung KY, Mele A, Fraser CE, Stone EF, Chen C, Fak JJ, Chi SW, Licatalosi DD, Richter JD, Darnell RB. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell. 2011;146:247–261. doi: 10.1016/j.cell.2011.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dityatev A, Schachner M, Sonderegger P. The dual role of the extracellular matrix in synaptic plasticity and homeostasis. Nat Rev Neurosci. 2010;11:735–746. doi: 10.1038/nrn2898. [DOI] [PubMed] [Google Scholar]
- Dziembowska M, Wlodarczyk J. MMP9: a novel function in synaptic plasticity. Int J Biochem Cell Biol. 2012;44:709–713. doi: 10.1016/j.biocel.2012.01.023. [DOI] [PubMed] [Google Scholar]
- Dziembowska M, Milek J, Janusz A, Rejmak E, Romanowska E, Gorkiewicz T, Tiron A, Bramham CR, Kaczmarek L. Activity-dependent local translation of matrix metalloproteinase-9. J Neurosci. 2012;32:14538–14547. doi: 10.1523/JNEUROSCI.6028-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari F, Mercaldo V, Piccoli G, Sala C, Cannata S, Achsel T, Bagni C. The fragile X mental retardation protein-RNP granules show an mGluR-dependent localization in the post-synaptic spines. Mol Cell Neurosci. 2007;34:343–354. doi: 10.1016/j.mcn.2006.11.015. [DOI] [PubMed] [Google Scholar]
- Hagerman RJ, Ono MY, Hagerman PJ. Recent advances in fragile X: a model for autism and neurodegeneration. Curr Opin Psychiatry. 2005;18:490–496. doi: 10.1097/01.yco.0000179485.39520.b0. [DOI] [PubMed] [Google Scholar]
- Hou L, Antion MD, Hu D, Spencer CM, Paylor R, Klann E. Dynamic translational and proteasomal regulation of fragile X mental retardation protein controls mGluR-dependent long-term depression. Neuron. 2006;51:441–454. doi: 10.1016/j.neuron.2006.07.005. [DOI] [PubMed] [Google Scholar]
- Kao DI, Aldridge GM, Weiler IJ, Greenough WT. Altered mRNA transport, docking, and protein translation in neurons lacking fragile X mental retardation protein. Proc Natl Acad Sci U S A. 2010;107:15601–15606. doi: 10.1073/pnas.1010564107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krichevsky AM, Kosik KS. Neuronal RNA granules: a link between RNA localization and stimulation-dependent translation. Neuron. 2001;32:683–696. doi: 10.1016/S0896-6273(01)00508-6. [DOI] [PubMed] [Google Scholar]
- Leigh MJ, Nguyen DV, Mu Y, Winarni TI, Schneider A, Chechi T, Polussa J, Doucet P, Tassone F, Rivera SM, Hessl D, Hagerman RJ. A randomized double-blind, placebo-controlled trial of minocycline in children and adolescents with fragile x syndrome. J Dev Behav Pediatr. 2013;34:147–155. doi: 10.1097/DBP.0b013e318287cd17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu R, Wang H, Liang Z, Ku L, O'donnell WT, Li W, Warren ST, Feng Y. The fragile X protein controls microtubule-associated protein 1B translation and microtubule stability in brain neuron development. Proc Natl Acad Sci U S A. 2004;101:15201–15206. doi: 10.1073/pnas.0404995101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucá R, Averna M, Zalfa F, Vecchi M, Bianchi F, Fata GL, Del Nonno F, Nardacci R, Bianchi M, Nuciforo P, Munck S, Parrella P, Moura R, Signori E, Alston R, Kuchnio A, Farace MG, Fazio VM, Piacentini M, De Strooper B, et al. The Fragile X Protein binds mRNAs involved in cancer progression and modulates metastasis formation. EMBO Mol Med. 2013;5:1523–1536. doi: 10.1002/emmm.201302847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaluk P, Kolodziej L, Mioduszewska B, Wilczynski GM, Dzwonek J, Jaworski J, Gorecki DC, Ottersen OP, Kaczmarek L. Beta-dystroglycan as a target for MMP-9, in response to enhanced neuronal activity. J Biol Chem. 2007;282:16036–16041. doi: 10.1074/jbc.M700641200. [DOI] [PubMed] [Google Scholar]
- Michaluk P, Wawrzyniak M, Alot P, Szczot M, Wyrembek P, Mercik K, Medvedev N, Wilczek E, De Roo M, Zuschratter W, Muller D, Wilczynski GM, Mozrzymas JW, Stewart MG, Kaczmarek L, Wlodarczyk J. Influence of matrix metalloproteinase MMP-9 on dendritic spine morphology. J Cell Sci. 2011;124:3369–3380. doi: 10.1242/jcs.090852. [DOI] [PubMed] [Google Scholar]
- Muddashetty RS, Kelić S, Gross C, Xu M, Bassell GJ. Dysregulated metabotropic glutamate receptor-dependent translation of AMPA receptor and postsynaptic density-95 mRNAs at synapses in a mouse model of fragile X syndrome. J Neurosci. 2007;27:5338–5348. doi: 10.1523/JNEUROSCI.0937-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Napoli I, Mercaldo V, Boyl PP, Eleuteri B, Zalfa F, De Rubeis S, Di Marino D, Mohr E, Massimi M, Falconi M, Witke W, Costa-Mattioli M, Sonenberg N, Achsel T, Bagni C. The fragile X syndrome protein represses activity-dependent translation through CYFIP1, a new 4E-BP. Cell. 2008;134:1042–1054. doi: 10.1016/j.cell.2008.07.031. [DOI] [PubMed] [Google Scholar]
- Paribello C, Tao L, Folino A, Berry-Kravis E, Tranfaglia M, Ethell IM, Ethell DW. Open-label add-on treatment trial of minocycline in fragile X syndrome. BMC Neurol. 2010;10:91. doi: 10.1186/1471-2377-10-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudelli RD, Brown WT, Wisniewski K, Jenkins EC, Laure-Kamionowska M, Connell F, Wisniewski HM. Adult fragile X syndrome. Clinico-neuropathologic findings. Acta Neuropath. 1985;67:289–295. doi: 10.1007/BF00687814. [DOI] [PubMed] [Google Scholar]
- Schaeffer C, Bardoni B, Mandel JL, Ehresmann B, Ehresmann C, Moine H. The fragile X mental retardation protein binds specifically to its mRNA via a purine quartet motif. EMBO J. 2001;20:4803–4813. doi: 10.1093/emboj/20.17.4803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma A, Hoeffer CA, Takayasu Y, Miyawaki T, McBride SM, Klann E, Zukin RS. Dysregulation of mTOR signaling in fragile X syndrome. J Neurosci. 2010;30:694–702. doi: 10.1523/JNEUROSCI.3696-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian M, Rage F, Tabet R, Flatter E, Mandel JL, Moine H. G-quadruplex RNA structure as a signal for neurite mRNA targeting. EMBO Rep. 2011;12:697–704. doi: 10.1038/embor.2011.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szklarczyk A, Lapinska J, Rylski M, McKay RD, Kaczmarek L. Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. J Neurosci. 2002;22:920–930. doi: 10.1523/JNEUROSCI.22-03-00920.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Utari A, Chonchaiya W, Rivera SM, Schneider A, Hagerman RJ, Faradz SM, Ethell IM, Nguyen DV. Side effects of minocycline treatment in patients with fragile X syndrome and exploration of outcome measures. Am J Intellect Dev Disabil. 2010;115:433–443. doi: 10.1352/1944-7558-115.5.433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zalfa F, Giorgi M, Primerano B, Moro A, Di Penta A, Reis S, Oostra B, Bagni C. The fragile X syndrome protein FMRP associates with BC1 RNA and regulates the translation of specific mRNAs at synapses. Cell. 2003;112:317–327. doi: 10.1016/S0092-8674(03)00079-5. [DOI] [PubMed] [Google Scholar]
- Zalfa F, Eleuteri B, Dickson KS, Mercaldo V, De Rubeis S, di Penta A, Tabolacci E, Chiurazzi P, Neri G, Grant SG, Bagni C. A new function for the fragile X mental retardation protein in regulation of PSD-95 mRNA stability. Nat Neurosci. 2007;10:578–587. doi: 10.1038/nn1893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zenklusen D, Wells AL, Condeelis JS, Singer RH. Imaging real-time gene expression in living yeast. CSH Prot prot4870. 2007 doi: 10.1101/pdb.prot4870. [DOI] [PMC free article] [PubMed] [Google Scholar]