

ABSTRACT
B-type cyclins in association with Cdk1 mediate key steps of mitosis and meiosis, by phosphorylating a plethora of substrates. Progression through the meiotic cell cycle requires the execution of two cell divisions named meiosis I and II without intervening S-phase, to obtain haploid gametes. These two divisions are highly asymmetric in the large oocyte. Chromosome segregation in meiosis I and sister chromatid segregation in meiosis II requires the sharp, switch-like inactivation of Cdk1 activity, which is brought about by degradation of B-type cyclins and counteracting phosphatases. Importantly and contrary to mitosis, inactivation of Cdk1 must not allow S-phase to take place at exit from meiosis I. Here, we describe recent studies on the regulation of translation and degradation of B-type cyclins in mouse oocytes, and how far their roles are redundant or specific, with a special focus on the recently discovered oocyte-specific role of cyclin B3.
KEYWORDS: Cyclin-dependent kinases, meiosis, oocytes, B-type cyclins, cyclin specificities, cell cycle control
Introduction
The generation of healthy, haploid gametes depends on the correct execution of a specialized form of cell division named meiosis. Unlike mitosis, meiosis contains two steps of DNA division with only one round of DNA replication. These two divisions are called meiosis I and meiosis II. In oocytes, these two rounds of chromosome segregation are regulated temporally, are highly asymmetric and depending on the species, contain cell cycle arrest points that are crucial for female gametogenesis. In mammalian meiosis, after premeiotic S-phase and meiotic recombination, oocytes are arrested at prophase I from birth of the female onwards. Upon hormonal stimulation, this arrest is lifted and meiosis resumes with entry into the first meiotic division. Oocytes undergo the first metaphase-to-anaphase transition where chromosomes segregate and half the DNA is discarded in the smaller cell, the polar body. Oocytes enter meiosis II immediately following meiosis I exit. Oocytes must then arrest at metaphase II awaiting fertilization which triggers the second division with segregation of sister chromatids (Figure 1). Once the oocyte is fertilized, the embryo can begin its development [1, 2, 3].
Figure 1.
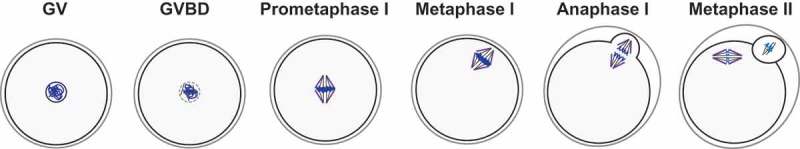
Mammalian oocyte maturation. Oocytes are arrested at prophase I, also known as germinal vesicle (GV) stage. Upon hormonal stimulation, fully-grown oocytes resume meiosis with germinal vesicle breakdown (GVBD). Chromosomes (in blue) condense, the bipolar spindle forms and microtubules attach to the chromosomes throughout prometaphase. In metaphase I chromosomes that are properly attached to the spindle and aligned at the metaphase plate migrate to the cortex. Chromosomes segregate during anaphase I with the extrusion of a first polar body. Oocytes immediately enter meiosis II and arrest at metaphase II awaiting fertilization.
It is primordial to understand the mechanisms that regulate the correct execution of female mammalian meiosis. Aneuploid embryos harbouring incorrect numbers of chromosomes, are mainly caused by maternal errors during female meiosis. 10–30% of all human oocytes are found to be aneuploid while only 1–2% of spermatozoa contain the wrong number of chromosomes [4–6]. This difference is increased in correlation with maternal age [5,7]. By age 42, the incidence of aneuploidy may increase up to 80%, according to [8]. Not all segregation errors are due to aberrant meiotic divisions though, and additionally, a high rate of segregation errors is observed in the following mitotic divisions at later stages of embryogenesis, up to the blastocyst stage [9,10]. Today, with more commonly used medically assisted reproduction and the increase of maternal age at first pregnancies, it is indispensable to gain a better comprehension of the regulatory mechanisms put in place to segregate the genetic material in meiosis and early mitotic divisions in the embryo. Insights into cell cycle regulation during meiotic maturation will help us understand what goes wrong so often in oocytes, which consequently leads to the generation of aneuploidies and hence, failure to generate healthy offspring.
Mammalian meiosis
Although meiotic divisions are composed of the same steps as mitosis, the fact that two divisions without intervening S-phase are executed with entirely different segregation patterns requires additional regulatory mechanisms. Detailed reviews on different aspects of chromosome and sister chromatid segregation in meiosis I and II, respectively, have been published and are beyond the scope of this review [2, 11, 12]. Here, we will focus on very recent results concerning the role of B-type cyclins for cell cycle progression in mammalian oocytes.
In oocytes, the size of the cell, as well as the time frame of meiotic maturation to enter meiosis I and ensure fertilization takes place at the right moment of cell division, demands distinct cell cycle regulation. Below we introduce progression through the meiotic divisions (meiotic maturation) from a cell cycle point of view, before describing the roles of the three existing mammalian B-type cyclins B1, B2 and B3 [13–15], all three of which have now been studied by knock-out approaches in mouse oocytes [16–21].
Mammalian oocytes are formed during embryonic development and following DNA replication and recombination, oocytes arrest at prophase I. This is known as GV stage due to the presence of a germinal vesicle. This signifies that oocytes must stay arrested for extended periods of time (months for mice, decades for humans) before the meiotic divisions. Before resuming meiosis, selected oocytes undergo a short growth phase in the adult female. Upon hormonal stimulation, the GV breaks down (GVBD) followed by chromosome condensation and spindle formation [1, 2, 3]. In mammals, a prolonged prometaphase in meiosis I allows the spindle to fully form and microtubules (MT) to attach correctly to kinetochores (KT), hence chromosomes can align at the metaphase plate. Similarly to mitosis, the Spindle Assembly Checkpoint (SAC) is present and active during meiotic cell division. It imposes a delay in anaphase I onset by inhibiting the Anaphase Promoting Complex/Cyclosome (APC/C), an E3 ubiquitin ligase essential for this cell cycle transition, to allow time for establishing correct KT-MT attachments [7,22,23]. In order to allow metaphase-to-anaphase I transition to take place, the APC/C must be activated in order to ubiquitinate proteins that block separase activity, namely securin and cyclin B1. Once ubiquitinated, these proteins are degraded and separase becomes active leading to chromosome segregation [24–26]. Oocytes then enter meiosis II immediately, and crucially, enter meiosis II without carrying out a second S-phase. Chromosomes then align and arrest at metaphase II. APC/C activation is inhibited to maintain this second meiotic arrest (also called Cystostatic factor (CSF-) arrest) until anaphase II is triggered by fertilization [11,27,28].
M-phase promoting factor (MPF)
Upon hormonal stimulation, the protracted GV arrest is lifted and this depends on the increase in MPF (M-phase promoting factor) kinase activity, the main driver of mitosis and meiosis. MPF phosphorylates a large number of substrates, and is required for entry into and progression through mitosis and meiosis, chromosome condensation, dissolution of the nuclear membrane, spindle formation, and chromosome alignment. It has to be inactivated for anaphase onset and exit from cell division. What makes up this factor has been extensively debated, but in both mitosis and meiosis MPF is considered as corresponding to the complex cyclin B-Cdk1 [29]. However, MPF activity is not only due to cyclin B-Cdk1 complexes but also the kinase Greatwall (Mastl in mammals) that is essential to protect the phosphorylated substrates of cyclin B-Cdk1 from the counteracting phosphatase PP2A, allowing appropriate entry and progression through mitosis and meiosis [29–31].
During the prolonged GV arrest in oocytes, Cdk1 is kept inactive through phosphorylation established by the Wee1 and Myt1 kinases, and APC/C dependent degradation of cyclin B . Although the molecular mechanism is not completely known in mouse oocytes, depending on hormonal stimulation and release from surrounding follicle cells, activation of Cdk1 is triggered by the phosphatase Cdc25 that removes the inhibitory phosphorylation on Cdk1. To be active, Cdk1 needs to be associated with cyclins, in this case cyclin B1 or B2, to induce GVBD. Once Cdk1 is dephosphorylated and sufficient MPF activity is generated, GVBD occurs with resumption of meiosis [32,33].
Overview of M-phase B type cyclins
Most of the components needed for meiosis are also present during mitosis. This is the case for the main regulators of the cell cycle, cyclins. Several cyclins are expressed depending on the species and cell cycle phase. In order to fulfill their purpose, cyclins bind to their catalytic partners, Cdks (cyclin-dependent kinases). Progression through mitotic cell division is thought to require just one B-type cyclin together with Cdk1, as in both budding and fission yeast, Clb2 or Cdc13 respectively, is the only cyclin required to be paired up with Cdk1 to drive cells through mitosis. In S. pombe, it was even shown that Cdc13-Cdc2 (which corresponds to Cdk1) can -in principle- drive progression through all stages of the cell cycle, indicating that cyclin specificity is not determinant for ordering cell cycle events [34–36]. This has led to a model proposing that threshold levels of Cdk1-associated activity determine substrate specificities, and not individual cyclins. Importantly though, this model does not seem to apply to mitosis and meiosis in other organisms. At least in mammalian oocytes, specific B-type cyclins habour distinct roles. Individual knock-outs of all three B-type cyclins have been described in the mouse and provide useful insights to the specificities of each cyclin during the meiotic divisions. In this review, we will discuss the redundant and distinct roles that B-type cyclins occupy in mouse oocyte meiosis, highlighting the importance of each cyclin in the correct timing of cell cycle events during this specialized cell division.
Cyclin B1
In mammals, cyclin B1 is the main cyclin driving entry into and progression through mitosis and meiosis. Apart from phosphorylations impinging on Cdk1, protein levels of cyclin B1 directly correlate with Cdk1-associated kinase activity, regulating key steps of mitotic and meiotic cell cycle progression. In mouse oocytes, cyclin B1-Cdk1 activity gradually increases for the first hours after GVBD, as oocytes progress through prometaphase, to reach its highest levels in metaphase I. This is due to inactivation of the APC/C activator Cdh1, and concomitant SAC activation, preventing activation of the APC/C in a Cdc20-dependent manner [25,37–40]. Slow progression through prometaphase allows the establishment of KT-MT attachments and correct alignment of chromosomes at the metaphase plate. For anaphase I onset, cyclin B1-Cdk1 activity must decrease drastically for a switch-like transition from metaphase to anaphase, and this is brought about by inactivation of the SAC and cyclin B1 degradation [7,11,22]. Cyclin B1-Cdk1 activity has to rise again quickly to inhibit S-phase onset, and instead promote entry into meiosis II with metaphase II or CSF-arrest. Upon fertilization, cyclin B1-Cdk1 activity must drop again for anaphase II onset with sister chromatid segregation [11,28,41].
Although protein synthesis is not necessary for GVBD, Cyclin B synthesis is essential for correct progression through meiosis in mouse oocytes [42, 43]. Following GVBD, the increase of cyclin B1 synthesis is responsible for the rise in Cdk1 activity, inducing chromosome condensation and congression as well as alignment during metaphase I. Another specificity of meiosis in mouse oocytes is that -unlike in cultured human cells – cyclin B1 is expressed in excess compared to its partner Cdk1 [44,45], indicating that there is a distinct regulation of MPF activity in meiosis compared to mitosis. The large size of the cell and the extended time needed for prometaphase and metaphase might be the reason for this difference. Similarly to mitotic securin degradation which occurs preferentially when securin is not found in association with separase [46, 47], Levasseur et al demonstrated in a very recent study that there are different pools of cyclin B1; free cyclin B1 and Cdk1-bound cyclin B1, and that association with Cdk1 determines temporal control of cyclin B1 degradation. Free cyclin B1 is degraded before all chromosomes are correctly attached and the SAC is turned off, in an APC/C-dependent manner. It continues to be degraded afterwards, through only its destruction box (Dbox), which is a motif recognized by the APC/C in association with Cdc20. The first degradation wave of free cyclin B1 depends on a newly identified motif within the N-terminal helix of cyclin B1. However, when cyclin B1 is bound to Cdk1, it is somewhat protected from the first wave of degradation that depends on this new motif. The authors show that this newly identified degron-like motif in cyclin B1 is recognized by the APC/C leading to prometaphase degradation in mouse oocytes, and that the sequence is masked through binding of Cdk1. This would allow the cell to eliminate progressively excess cyclin B1 in prometaphase I in a SAC-independent manner, to allow the rapid inactivation of cyclin B1-Cdk1 needed for the anaphase switch to occur on time [44]. At the same time, this would provide a stock of cyclin B1 available for association with Cdk1, to make sure that cyclin B1 does not become a limiting factor for Cdk1 activity in oocytes (Figure 2). Indeed, reducing the amount of free cyclin B1 in oocytes leads to defects in chromosome alignment at the metaphase I plate and hence, missegregation events [44]. The authors propose that excess cyclin B1 compensates for weak SAC control in oocytes to ensure correct chromosome segregation [44]. It is interesting to note in this context that endogenous cyclin A2 degradation occurs also in two waves in oocytes, with most of cyclin A2 being degraded in prometaphase I, and a small fraction of cyclin A2 localized to kinetochores escaping degradation until anaphase I onset [48]. Future studies will show whether a similar motif as the one identified in cyclin B1 can be identified in the N-terminal helix of cyclin A2, or other B-type cyclins.
Figure 2.
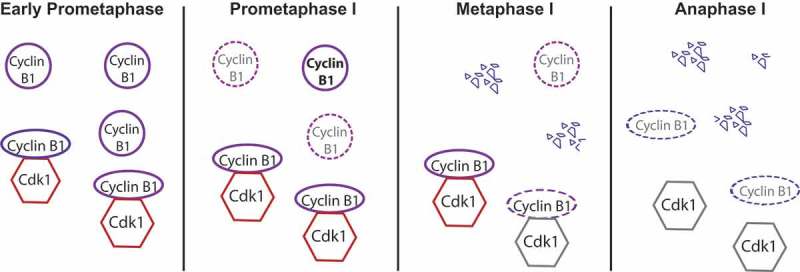
Regulation of cyclin B1 degradation during meiosis I. At early prometaphase, different pools of cyclin B1 are present in the oocyte: free cyclin B1 and cyclin B1 bound to Cdk1. During prometaphase, free cyclin B1 starts to be degraded while the SAC is on, whereas Cdk1-bound cyclin B1 is protected from APC/C-dependent ubiquitination and degradation through its binding. Most of the free cyclin B1 is degraded once oocytes progress into metaphase I, while bound cyclin B1 begins to be degraded only once the SAC is satisfied and Cdc20-APC/C under SAC control becomes active. Once the APC/C is fully active, cyclin B1 bound to Cdk1 is degraded leading to a sharp decrease of Cdk1 activity and anaphase I onset.
Additionally, the location of cyclin B1 and binding to separase may also play a role in regulating its ubiquitination by the APC/C. In vitro cultured mitotic cells seem to harbour a pool of cyclin B1 that preferentially inhibits separase near chromosomes, compared to cytoplasmic cyclin B1. The degradation of this pool of cyclin B1 inhibiting separase is slower compared to the cytoplasmic pool [49], indicating that APC/C dependent degradation is determined by the partners cyclin B1 is associated with. Whether location of cyclin B1 and binding to separase regulate cyclin B1 ubiquitination also in oocytes is currently unknown.
Altogether, these studies indicate that additional regulation of cyclin B1 in meiosis and mitosis is accomplished through differential APC/C affinity for the separate pools present in the cell in prometaphase. When the SAC is satisfied, APC/C with its co-activator Cdc20, targets cyclin B1 for degradation through ubiquitination. This brings about a sharp decrease of Cdk1 activity and exit from meiosis I. Once vertebrate oocytes exit meiosis I, they continue into meiosis II and arrest at metaphase II awaiting fertilization. In order for this arrest to be maintained, Cdk1 activity must be kept at high enough levels. Cyclin B1 is necessary for this very dynamic arrest, with an equilibrium of cyclin B1 degradation and synthesis to keep Cdk1 activity at an appropriate threshold [50,51]. In Xenopus oocytes, it was found that cyclin B1 and B2 synthesis continues during CSF arrest while Cdk1 activity level seems to be constant. It is thought that once a high amount of B-type cyclins is synthesized this leads to an increase in Cdk1 activity past a specific threshold. The APC/C with its co-activator Cdc20 becomes active inducing degradation of B-type cyclins. As a consequence, Cdk1 activity drops until another regulatory pathway, the Mos-MAPK pathway, counteracts APC/C activation allowing once again synthesis of cyclin B1 and B2. Therefore, CSF-arrest is dynamic, brought about by an equilibrium between cyclin B1 and B2 synthesis and APC/C activation [52]. Additionally, Cdc25 phosphatase is required during the arrest to remove inhibitory phosphorylation on Cdk1. Upon fertilization, the Mos-MAPK pathway is inhibited, allowing full APC/C activation and cyclin B1 and B2 degradation, while Cdk1 is additionally downregulated by the kinase Wee1B, which phosphorylates Cdk1 for further inhibition[53, 54, 55, 56]. Oocytes exit meiosis II and form a female pronucleus, which fuses with the male pronucleus provided by the sperm, to re-establish the diploid chromosome content in the zygote [57].
Cyclin B1 vs cyclin B2
Overall, cyclin B1 expression seemed to be necessary for development and fertility in mice while cyclin B2 is dispensable [20]. Cyclin B1 null mice are not viable while cyclin B2 null mice are viable and fertile, although litters are smaller in size and numbers [20]. Lack of cyclin B1 in embryos induces an arrest after only two divisions at the 4-cell stage, underlining its significance in embryonic development [58]. Both cyclins are expressed during mitosis although they are localized differently, next to microtubules (cyclin B1) and the Golgi region (cyclin B2), respectively, and Cyclin B1 is translocated to the nucleus during prophase unlike cyclin B2 [59]. In mouse oocytes, the distribution of exogenous cyclin B1 evolves before GVBD with a cytoplasmic localization at first followed by its import into the nucleus [60]. Nuclear targeting of exogenous cyclin B1 significantly increased its capacity to induce entry into meiosis I[61], indicating that localization is an important factor regulating cyclin B1’s capacity to induce GVBD. But as the importance of cyclin B1 and B2 subcellular localization for its role during meiotic maturation has not been further studied in mouse oocytes, it will not be subject of further discussion here.
Translational regulation is specific to each cyclin in mouse oocytes. The translation of cyclin B1 is tightly regulated in mouse oocytes since at different time points, cyclin B1 has to be rapidly synthesized. mRNA granule formation plays a role in inhibiting translation while their dispersion allows quick synthesis of cyclin B1 in mouse oocyte meiosis I [62]. Additionally, timing of cyclin B1 translation is regulated through polyadenylation mediated by specific lengths of the 3ʹUTR. Long and intermediate 3ʹUTR length of cyclin B1 mRNA leads to repression of translation in GV arrested oocytes, and to their translation after GVBD while cyclin B1 mRNA with short 3ʹUTR is constitutively translated already in GV oocytes [63]. Following GVBD, the synthesis and amount of cyclin B1 increases while cyclin B2 remains at stable levels (Figure 3). Unlike cyclin B2, cyclin B1 transcripts are regulated through ribosome loading, dependent on the RNA-binding protein CPEB1. This loading of ribosomes on cyclin B1 increases after GVBD while ribosome loading on cyclin B2 does not change after resumption of meiosis. Intriguingly, Cdk1-associated activity was found to play a role in the phosphorylation of CPEB1 and therefore, to regulate cyclin B1 translation [64]. It is attractive to speculate that cyclin A2, a strong inducer of GVBD when expressed exogenously [48], may act as a trigger for entry into meiosis I by mediating phosphorylation of CPEB1, but this has not been addressed yet.
Figure 3.
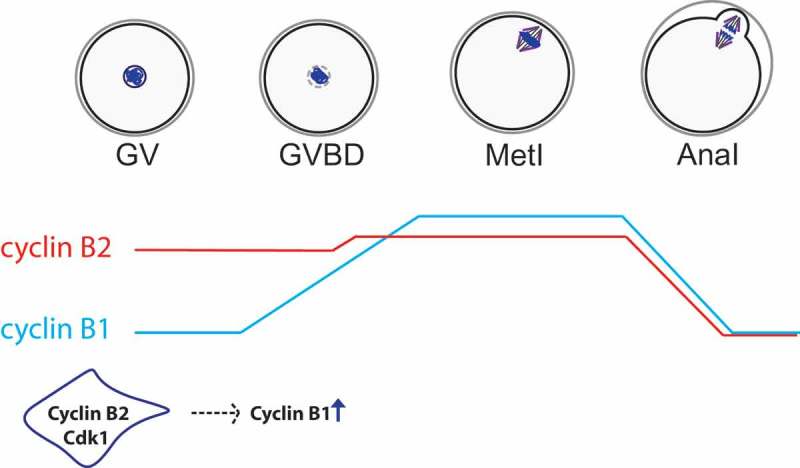
Protein levels of cyclin B1 vs. cyclin B2 in meiosis I. According to 64, preceding resumption of meiosis, cyclin B2 (in red) is translated at higher levels than cyclin B1 (in blue) while their degradation rates are similar. Therefore, at GV stage, cyclin B2 protein is more abundant than cyclin B1. After entry into meiosis I, protein levels of cyclin B2 remain similar with only a slight increase in translation. However, cyclin B1 is actively translated leading to an increase of protein levels following GVBD reaching its maximum at metaphase I. Cyclin B1 translation depends on cyclin B2 protein at entry into meiosis I. Hence, protein levels of both cyclins are regulated differentially and temporally in mouse oocytes.
Complete loss of either cyclin B1 or B2 in mouse oocytes
To elucidate the contributions of cyclin B1 and B2 for meiotic progression in oocytes, two recent studies examined meiotic maturation in mice harbouring oocytes devoid of either cyclin with knock-out approaches targeting the gene encoding cyclin B1 (Ccnb1) or cyclin B2 (Ccnb2). In mitotic tissues, only cyclin B1 seems to be essential, and a conditional knock-out approach using a Gdf9-dependent deletion strategy was necessary to determine its role in oocytes. Surprisingly though, oocytes without cyclin B1 are still able to enter meiosis I and extrude a polar body. No abnormalities as far as chromosome alignment and spindle formation is concerned, were observed in oocytes devoid of cyclin B1. However, once these oocytes exit meiosis I, they fail to enter meiosis II and to establish a CSF arrest. At entry into meiosis I, cyclin B2 is upregulated and promotes GVBD in these Ccnb1-/- oocytes, but after meiosis I, except when over-expressed, cyclin B2 cannot substitute for cyclin B1. Because oocytes fail to re-activate Cdk1 after meiosis I, Ccnb1-/- oocytes decondense chromosomes and seem to enter an interphase-like state. Importantly, injection of mRNA coding for cyclin B2 can rescue meiosis II in Ccnb1-/- oocytes, indicating that the failure of cyclin B1 oocytes to progress beyond meiosis I may be due to a failure to re-accumulate sufficient levels of cyclin B2 to compensate for loss of cyclin B1 such as in meiosis I, and not due to a specific role of cyclin B1 in meiosis II. Collectively these data indicate that cyclin B2 can substitute for cyclin B1 in oocyte meiosis [16,20].
Ccnb2 knock-out mice are viable, allowing the analysis of meiosis in oocytes devoid of cyclin B2. Without cyclin B2, entry into meiosis I is significantly delayed [16,21,65], because oocytes fail to activate Cdk1 efficiently for GVBD. Unlike cyclin B2 in Ccnb1-/- oocytes, no upregulation of cyclin B1 to compensate for loss of cyclin B2 was observed. Quite the opposite, translation of mRNA coding for cyclin B1 was affected in the absence of cyclin B2 in a proportion of oocytes, indicating that this cyclin has a specific role in translational control of cyclin B1 and other, oocyte-specific mRNAs such as coding for Mos [21]. How cyclin B2 brings about translational control of meiotic transcripts is currently unknown, but may involve phosphorylation of CPEB1, which occurs in a Cdk1-dependent manner [64]. If cyclin B2 together with Cdk1 is able to phosphorylate CPEB1, it will be important to determine whether this is due to specific substrate specificity of cyclin B2, or due to the fact that cyclin B2 translation occurs independently of the CPEB1. Because of missing cyclin B2 protein in GV oocytes and failure to accumulate sufficiently high levels of cyclin B1, overall MPF activity is lower in Ccnb2-/- oocytes, resulting in defects in spindle formation, delayed and inefficient APC/C activation, delay in anaphase I onset and reduced number of oocytes that succeed to extrude a polar body and exit meiosis I. These defects were rescued by overexpression of cyclin B1 [21]. The authors suggest that the metaphase I arrest observed in a significant proportion of Ccnb2-/- oocytes is due to SAC activation because inhibition of the essential SAC kinase Mps1 can rescue polar body extrusion. Ccnb2-/- oocytes that fail to extrude a polar body show kinetochore recruitment of the SAC component Mad2 indicating prolonged SAC activation, even though under normal conditions the SAC is activated only very transiently in meiosis I [21]. A small caveat of this assay is the fact that without a functional SAC, oocytes undergo metaphase-to-anaphase transition precociously and with much lower Cdk1 activity [66,67]. Therefore, oocytes devoid of cyclin B2 may harbour just enough cyclin B1-Cdk1 activity to undergo metaphase-to-anaphase transition in the absence of a functional SAC, explaining the rescue. Overall, the data obtained from complete loss of either cyclin indicates that cyclin B1 and B2 harbour specific roles during meiotic maturation mainly because of their expression profiles brought about in part by a specific role of cyclin B2 for translational control of cyclin B1. They can substitute for each other when expressed at the required times during meiotic maturation [16,21], indicating that they can phosphorylate the same substrates throughout meiosis I and II.
Cyclin B3
Evidence from the nucleotide sequence, cyclin box conservation, expression pattern and Cdk binding site pointed to cyclin B3 being a distinct and unique cyclin. Cyclin B3 was first described in chicken and found to exhibit 33% identity with cyclin B2 and 30% identity with A-type cyclins showing that it resembles both types of cyclins. Cyclin B3 was found to be located in the nucleus in interphase, similarly to A-type cyclins, while cyclin B1 is located in the cytoplasm [68, 69, 70]. In C. elegans, cyclin B3 was found to be close to chicken cyclin B3 with 50% identity in the cyclin box. Cyclin B3 is mainly expressed in maternal germ cells and conserved in other worms [71]. In humans, cyclin B3 mRNA was detected by RT-PCR in testis and 8 other tissues, but only in testis by northern blot analysis, suggesting low abundance in tissues other than germ cells. Protein levels and mRNA expression was found at highest levels in prepachytene spermatocytes and in oocytes during embryonic development [70]. This particular expression hinted at a role in early meiosis I (prophase) in both male and female meiosis.
Concerning Cyclin B3’s sequence encoded by Ccnb3 in mice, an important extension of exon 8 is observed in placental mammals and not in other species, suggesting that the resulting 3-fold increase in the size of the protein occurred during evolution of placental mammals. The extension and increase in size makes placental mammals’ cyclin B3 of substantial size compared to other species; human cyclin B3 is 1395 aa long while chicken and Xenopus cyclin B3 are 403 aa and 416 aa long, respectively. This extended sequence could mediate new protein-protein interactions that may be important for its function specifically in these species [72]. Contrary to human and mouse cyclin B3, Drosophila and C.elegans cyclin B3 is expressed in both meiotic and mitotic cells during embryogenesis [71,73–77]. Although it is expressed in mitotic Drosophila cells, cyclin B3 was found to be dispensable for mitosis but essential for female meiosis [74]. This differs from C. elegans cyclin B3 which is essential for embryonic development. In humans, aberrant expression of cyclin B3 during mitosis has been observed in a specific type of cancer, Ewing-like sarcoma, where a fusion of cyclin B3 and a ubiquitously expressed protein has been found [78–80], indicating that untimely expression of this cyclin can have serious consequences on cell fate.
The importance and role of cyclin B3 has been established in several species although the molecular mechanism is not fully understood. In Drosophila and C. elegans oocytes and embryos, cyclin B3 was found to be a partner of Cdk1 although a possible interaction with Cdk2 was not examined [74, 81]. In chicken mitotic cells, cyclin B3 was able to bind to both Cdk1 and Cdk2 [68]. Surprisingly, human cyclin B3 did not co-immunoprecipitate with Cdk1 but was found to interact with Cdk2 resulting in very low in vitro kinase activity [82]. Mouse cyclin B3 purified from insect cells is able to be partnered up with Cdk1 and associated in vitro kinase activity was detected [18].
In C. elegans, cyclin B3 knockdown is lethal and detrimental to development. Cyclin B3 deficient embryos show several phenotypic defects: in meiosis, cyclin B3 depleted oocytes are not able to segregate sister chromatids during anaphase II although other cell cycle steps occur. Once both male and female pronuclei are formed, their migration is slower in mutant embryos. During the first mitosis, metaphase was found to be delayed and chromosomes do not segregate. This phenotype is partially rescued by simultaneously knocking down SAC components indicating that cyclin B3’s role in C. elegans is SAC dependent and essential for embryonic development [77]. As previously discussed, cyclin B3 in Drosophila is dispensable for viability and male meiosis but essential for female meiosis. Cyclin B3 deficient Drosophila females are sterile [74]. In Drosophila oocytes, cyclin B3 shares some functions with other cyclins, cyclin A and cyclin B, but also harbours a unique role in promoting anaphase onset in both oocytes and embryos, contrary to cyclin B1 that inhibits anaphase onset [69,73–75]. Furthermore, together with cyclin A, cyclin B3 inhibits entry into S phase between meiosis I and II [75]. Although cells progress slower through the cell cycle, cyclin B1 is dispensable for mitosis in Drosophila. Only double knockdown of cyclin B3 and B1 leads to embryonic lethality once maternal reserves are depleted [74,75]. Surprisingly, a different role of cyclin B3 was uncovered in the ascidian Ciona intestinalis. Ciona intestinalis cyclin B3 was found to be essential for maternal to zygotic transition, where zygotic genome activation (ZGA) occurs. The authors described a correlation between the decrease of cyclin B3 mRNA and start of ZGA. Knockdown of cyclin B3 led to advanced initiation of ZGA [83] demonstrating yet another role for this cyclin.
Through the analysis of Ccnb3 knock-out male mice it was found that although expressed in testis in mice, its role is dispensable during spermatogenesis [17,18]. Furthermore, its protein and mRNA levels are low in adult ovaries; nevertheless, its role is essential for correct meiotic maturation, as oocytes cannot progress beyond metaphase I without cyclin B3 [18,19,84]. Complete loss of cyclin B3 leads to a SAC-independent metaphase I arrest, with high Cdk1 activity and a failure to correctly degrade endogenous APC/C substrates [18,19], even though exogenously expressed substrates are degraded [18]. This may indicate that cyclin B3 is required for targeting specific pools of cyclin B1 and securin for APC/C dependent ubiquitination, such as cyclin B1 associated with Cdk1, and securin in association with separase [44,46,49]. Alternatively, cyclin B3 may be involved in regulating substrate specificity of the APC/C by phosphorylating one of its subunits, or by activating phosphatases that determine substrate specificity through some yet unknown mechanism (Figure 4). Lastly, cyclin B3 may be required for translational regulation of cyclin B1 and securin mRNAs, and in its absence, translation is upregulated leading to the observed failure to degrade endogenous substrates and metaphase I arrest. A cyclin B3 mutant that does not yield in vitro kinase activity with Cdk1 was also not able to rescue oocytes devoid of cyclin B3 [18], strongly suggesting that Cdk1 activity associated with cyclin B3 is essential for its role in female meiosis. Most importantly, cyclin B1 cannot substitute for cyclin B3, showing that cyclin B3 has a non-redundant and specific role in oocytes, indicating that it confers distinct substrate specificity to Cdk1 than cyclin A2, B1, or B2 [18]. As mentioned above, cyclin B3 in placental mammalians is three times bigger than from other vertebrates[72]. It came therefore as a surprise that the function of cyclin B3 seems to be conserved between species, because expression of cyclin B3 derived from Drosophila, zebrafish and Xenopus was able to rescue the meiotic defects of Ccnb3 knock-out mouse oocytes [18]. The extended exon 8 in placental mammals seems therefore not absolutely required for cyclin B3 function in meiosis.
Figure 4.
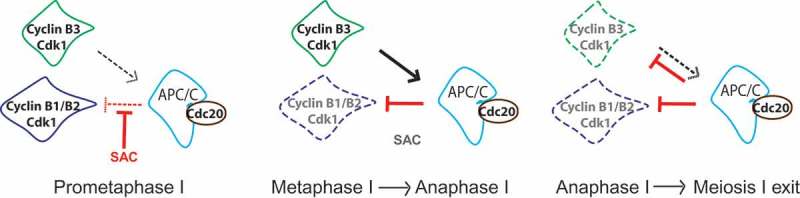
Model of how cyclin B3 may promote anaphase I onset in mouse oocytes. Cyclin B1/B2-Cdk1 (in blue) activity increases as oocytes progress through prometaphase into metaphase, because the APC/C (light blue) in association with its activator Cdc20 (brown) is kept in check by the SAC. At the metaphase-to-anaphase transition, the SAC is satisfied and inactivated, allowing full APC/C activity, ubiquitination and hence degradation of cyclin B1 and B2 in association with Cdk1. In oocytes, full APC/C activity requires the function of cyclin B3-Cdk1. Cyclin B3-Cdk1 (in green) promotes APC/C activity leading to chromosome segregation and exit from meiosis I. Therefore cyclin B3, a late substrate of the APC/C, shows an opposing role to cyclin B1 and B2 during mouse oocyte meiosis I.
Conclusion
Recent studies using knock-out mouse models and sophisticated imaging approaches have provided important insights on the roles of B-type cyclins during female mammalian meiosis. Cyclin B2 occupies a specific role in oocytes, regulating translation of meiotic mRNAs, and this role of cyclin B2 is essential for accumulating sufficient levels of MPF activity for meiotic cell cycle progression. Cyclin B1 and B2 seem to be largely redundant as far as MPF substrate phosphorylation for progression through meiosis I and meiosis II are concerned, and their individual roles in oocyte meiosis are most likely due to differences in expression and localization. Cyclin B3 clearly occupies a distinct role in meiotic cell cycle progression, specific to oocytes. Cyclin B3 is required for metaphase-to-anaphase transition in meiosis I, whereas cyclin B1 and B2 inhibit anaphase I onset. Recent data indicate that cyclin B3-Cdk1 substrates are not shared with other A- and B-type cyclins, because none of them can substitute for cyclin B3, putting this cyclin apart. Future work will aim at identifying the targets of cyclin B3 to elucidate why this cyclin is required specifically in oocytes. It is attractive to speculate that the size of the oocyte requires the additional contribution of cyclin B3 for ensuring switch-like transitions in meiosis.
Funding Statement
This work was supported by the Fondation pour la Recherche Médicale [Equipe DEQ20160334921]; Agence National de la Recherche [ANR-16-CE92-0007-01].
Acknowledgments
We thank members of the Wassmann lab for discussion, and S.A. Touati (Francis Crick Institute, London) for valuable comments. NB received a 3-year PhD fellowship from the French Ministère de la Recherche, followed by a 1-year PhD fellowship by the Fondation ARC pour la Recherche sur le Cancer. The lab of K.W. is financed through a grant “Equipe FRM” by the Fondation de la Recherche Médicale (Equipe DEQ20160334921, to KW), an international ANR grant (ANR-16-CE92-0007-01 to KW), and core funding from Paris Sorbonne Université and the CNRS.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- [1].El Yakoubi W, Wassmann K. Meiotic divisions: no place for gender equality In: Gotta M, Meraldi P, editors. Cell division machinery and disease. Cham: Springer International Publishing; 2017. p. 1–17. Advances in Experimental Medicine and Biology DOI: 10.1007/978-3-319-57127-0_1 [DOI] [Google Scholar]
- [2].Holt JE, Lane SIR, Jones KT.. Chapter seven - The control of meiotic maturation in mammalian oocytes In: Wassarman PM, editor. Current topics in developmental biology. Vol. 102 Gametogenesis: Academic Press; 2013. p. 207–226. [DOI] [PubMed] [Google Scholar]
- [3].Herbert M, Kalleas D, Cooney D, et al. Meiosis and maternal aging: insights from aneuploid oocytes and trisomy births. Cold Spring Harb Perspect Biol. 2015;7(4):a017970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Templado C, Uroz L, Estop A. New insights on the origin and relevance of aneuploidy in human spermatozoa. Mol Hum Reprod. 2013;19(10):634–643. [DOI] [PubMed] [Google Scholar]
- [5].Hassold T, Hunt P. To Err (Meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet. 2001;2(4):280. [DOI] [PubMed] [Google Scholar]
- [6].Nagaoka SI, Hassold TJ, Hunt PA. Human aneuploidy: mechanisms and new insights into an age-old problem. Nat Rev Genet. 2012;13(7):493–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Touati SA, Wassmann K. How oocytes try to get it right: spindle checkpoint control in meiosis. Chromosoma. 2016;125(2):321–335. [DOI] [PubMed] [Google Scholar]
- [8].Capalbo A, Hoffmann ER, Cimadomo D, et al. Human female meiosis revised: new insights into the mechanisms of chromosome segregation and aneuploidies from advanced genomics and time-lapse imaging. Hum Reprod Update. 2017;23(6):706–722. [DOI] [PubMed] [Google Scholar]
- [9].Vázquez-Diez C, FitzHarris G. Causes and consequences of chromosome segregation error in preimplantation embryos. Reproduction. 2018;155(1):R63–76. [DOI] [PubMed] [Google Scholar]
- [10].Schneider I, Ellenberg J. Mysteries in embryonic development: how can errors arise so frequently at the beginning of mammalian life? PLoS Biol. 2019;17(3):e3000173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Sanders JR, Jones KT. Regulation of the meiotic divisions of mammalian oocytes and eggs. Biochem Soc Trans. 2018;46(4):797–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Wassmann K. Sister chromatid segregation in meiosis II : deprotection through phosphorylation. Cell Cycle. 2013;12(9):1352–1359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Pines J, Hunter T. Isolation of a human cyclin CDNA: evidence for cyclin MRNA and protein regulation in the cell cycle and for interaction with P34cdc2. Cell. 1989;58(5):833–846. [DOI] [PubMed] [Google Scholar]
- [14].Chapman DL, Wolgemuth DJ. Identification of a Mouse B-type cyclin which exhibits developmentally regulated expression in the germ line. Mol Reprod Dev. 1992;33(3):259–269. [DOI] [PubMed] [Google Scholar]
- [15].Nieduszynski CA, Murray J, Carrington M. Whole-genome analysis of animal A- and B-type cyclins. Genome Biol. 2002;3(12). research0070.1 DOI: 10.1186/gb-2002-3-12-research0070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Jian L, Tang J-X, Cheng J-M, et al. Cyclin B2 can compensate for cyclin B1 in oocyte meiosis I. J Cell Biol. 2018;217(11):3901–3911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Karasu ME, Keeney S. Cyclin B3 is dispensable for mouse spermatogenesis. BioRxiv. 2019. April;608315 DOI: 10.1101/608315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Karasu ME, Bouftas N, Keeney S, et al. Cyclin B3 promotes anaphase I onset in oocyte meiosis. J Cell Biol. 2019. February:201808091 jcb DOI: 10.1083/jcb.201808091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Yufei L, Wang L, Zhang L, et al. Cyclin B3 is required for metaphase to anaphase transition in oocyte meiosis I. J Cell Biol. 2019. February:201808088 jcb DOI: 10.1083/jcb.201808088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Brandeis M, Rosewell I, Carrington M, et al. Cyclin B2-null mice develop normally and are fertile whereas cyclin B1-null mice die in utero. Proc Nat Acad Sci. 1998;95(8):4344–4349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Daldello EM, Luong XG, Yang C-R, et al. Cyclin B2 is required for progression through meiosis in mouse oocytes. Development. 2019;146:172734 April, dev. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Gorbsky GJ. The spindle checkpoint and chromosome segregation in meiosis. Febs J. 2015;282(13):2458–2474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Homer H. The APC/C in female mammalian meiosis I. Reproduction. 2013;146(2):R61–71. [DOI] [PubMed] [Google Scholar]
- [24].Emilie TM, Wassmann K, Waizenegger I, et al. The Meiosis I-to-Meiosis II transition in mouse oocytes requires separase activity. Curr Biol. 2003;13(20):1797–1802. [DOI] [PubMed] [Google Scholar]
- [25].Herbert M, Levasseur M, Homer H, et al. Homologue disjunction in mouse oocytes requires proteolysis of securin and cyclin B1. Nat Cell Biol. 2003;5(11):1023–1025. [DOI] [PubMed] [Google Scholar]
- [26].Kudo NR, Wassmann K, Anger M, et al. Resolution of chiasmata in oocytes requires separase-mediated proteolysis. Cell. 2006;126:135–146. [DOI] [PubMed] [Google Scholar]
- [27].Runft LL, Jaffe LA, Mehlmann LM. Egg activation at fertilization: where it all begins. Dev Biol. 2002;245(2):237–254. [DOI] [PubMed] [Google Scholar]
- [28].Perry ACF, Verlhac M-H. Second meiotic arrest and exit in frogs and mice. EMBO Rep. 2008;9(3):246–251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Kishimoto T. Entry into mitosis: a solution to the decades-long enigma of MPF. Chromosoma. 2015;124(4):417–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Vigneron S, Robert P, Hached K, et al. The master greatwall kinase, a critical regulator of mitosis and meiosis. Int J Dev Biol. 2016;60(7–8–9):245–254. [DOI] [PubMed] [Google Scholar]
- [31].Adhikari D, Liu K. The regulation of maturation promoting factor during Prophase I arrest and meiotic entry in mammalian oocytes. Mol Cell Endocrinol. 2014;382(1):480–487. [DOI] [PubMed] [Google Scholar]
- [32].Han SJ, Conti M. New pathways from PKA to the Cdc2/Cyclin B complex in oocytes: wee1B as a potential PKA substrate. Cell Cycle. 2006;5(3):227–231. [DOI] [PubMed] [Google Scholar]
- [33].Solc P, Schultz RM, Motlik J. Prophase I arrest and progression to metaphase I in mouse oocytes: comparison of resumption of meiosis and recovery from G2-arrest in somatic cells. Mol Hum Reprod. 2010;16(9):654–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Richardson H, Lew DJ, Henze M, et al. Cyclin-B homologs in saccharomyces cerevisiae function in S phase and in G2. Genes Dev. 1992;6(11):2021–2034. [DOI] [PubMed] [Google Scholar]
- [35].Gutiérrez-Escribano P, Nurse P. A single cyclin–CDK complex is sufficient for both mitotic and meiotic progression in fission yeast. Nat Commun. 2015;6(1):6871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Fisher DL, Nurse P. A single fission yeast mitotic cyclin B P34cdc2 kinase promotes both S-phase and mitosis in the absence of G1 cyclins. Embo J. 1996;15(4):850–860. [PMC free article] [PubMed] [Google Scholar]
- [37].McGuinness BE, Anger M, Anna Kouznetsova AM, et al. Regulation of APC/C activity in oocytes by a Bub1-dependent spindle assembly checkpoint. Curr Biol. 2009;19(5):369–380. [DOI] [PubMed] [Google Scholar]
- [38].Wassmann K, Niault T, Maro B. Metaphase I arrest upon activation of the Mad2-dependent spindle checkpoint in mouse oocytes. Curr Biol. 2003;13(18):1596–1608. [DOI] [PubMed] [Google Scholar]
- [39].Ledan E, Polanski Z, Terret M-E, et al. Meiotic maturation of the mouse oocyte requires an equilibrium between cyclin B synthesis and degradation. Dev Biol. 2001;232(2):400–413. [DOI] [PubMed] [Google Scholar]
- [40].Reis A, Madgwick S, Chang H-Y, et al. Prometaphase APCcdh1 activity prevents non-disjunction in mammalian oocytes. Nat Cell Biol. 2007;9(10):1192–1198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Jones KT. Mammalian egg activation: from Ca2+ spiking to cell cycle progression. Reproduction. 2005;130(6):813–823. [DOI] [PubMed] [Google Scholar]
- [42].Vantéry C, de Gavin AC, Vassalli JD, et al. An accumulation of P34cdc2at the end of mouse oocyte growth correlates with the acquisition of meiotic competence. Dev Biol. 1996;174(2):335–344. [DOI] [PubMed] [Google Scholar]
- [43].Hashimoto N, Kishimoto T. Regulation of meiotic metaphase by a cytoplasmic maturation-promoting factor during mouse oocyte maturation. Dev Biol. 1988;126(2):242–252. [DOI] [PubMed] [Google Scholar]
- [44].Levasseur MD, Thomas C, Davies OR, et al. Aneuploidy in oocytes is prevented by sustained CDK1 activity through degron masking in cyclin B1. Dev Cell. 2019February;48:672–684.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Arooz T, Yam CH, Siu WY, et al. On the concentrations of cyclins and cyclin-dependent kinases in extracts of cultured human cells. Biochemistry. 2000;39(31):9494–9501. [DOI] [PubMed] [Google Scholar]
- [46].Hellmuth S, Böttger F, Pan C, et al. PP2A delays APC/C‐dependent degradation of separase‐associated but not free securin. Embo J. 2014;33(10):1134–1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Holt LJ, Krutchinsky AN, Morgan DO. Positive feedback sharpens the anaphase switch. Nature. 2008;454(7202):353–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Touati SA, Cladière D, Lister LM, et al. Cyclin A2 is required for sister chromatid segregation, but not separase control, in mouse oocyte meiosis. Cell Rep. 2012;2(5):1077–1087. [DOI] [PubMed] [Google Scholar]
- [49].Konishi M, Shindo N, Komiya M, et al. Quantitative analyses of the metaphase-to-anaphase transition reveal differential kinetic regulation for securin and cyclin B1. Biomed Res. 2018;39(2):75–85. [DOI] [PubMed] [Google Scholar]
- [50].Kubiak JZ, Weber M, de Pennart H, et al. The metaphase II arrest in mouse oocytes is controlled through microtubule-dependent destruction of cyclin B in the presence of CSF. Embo J. 1993;12(10):3773–3778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Nixon VL, Levasseur M, McDougall A, et al. Ca2+ oscillations promote APC/C-dependent cyclin B1 degradation during metaphase arrest and completion of meiosis in fertilizing mouse eggs. Curr Biol. 2002;12(9):746–750. [DOI] [PubMed] [Google Scholar]
- [52].Yamamoto TM, Iwabuchi M, Ohsumi K, et al. APC/C–cdc20-mediated degradation of cyclin B participates in CSF arrest in unfertilized xenopus eggs. Dev Biol. 2005;279(2):345–355. [DOI] [PubMed] [Google Scholar]
- [53].Oh JS, Susor A, Schindler K, et al. Cdc25A activity is required for the metaphase II arrest in mouse oocytes. J Cell Sci. 2013;126(5):1081–1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Oh JS, Susor A, Conti M. Protein tyrosine kinase Wee1B is essential for metaphase II exit in mouse oocytes. Science. 2011;332(6028):462–465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Verlhac M-H, Lefebvre C, Kubiak JZ, et al. Mos activates MAP kinase in mouse oocytes through two opposite pathways. Embo J. 2000;19(22):6065–6074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Verlhac MH, Kubiak JZ, Clarke HJ, et al. Microtubule and chromatin behavior follow MAP kinase activity but not MPF activity during meiosis in mouse oocytes. Development. 1994;120(4):1017–1025. [DOI] [PubMed] [Google Scholar]
- [57].Clift D, Schuh M. Re-starting life: fertilization and the transition from meiosis to mitosis. Nat Rev Mol Cell Biol. 2013;14(9):549–562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Strauss B, Harrison A, Coelho PA, et al. Cyclin B1 is essential for mitosis in mouse embryos, and its nuclear export sets the time for mitosis. J Cell Biol. 2018;217(1):179–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Jackman M, Firth M, Pines J. Human Cyclins B1 and B2 are localized to strikingly different structures: B1 to microtubules, B2 primarily to the golgi apparatus. Embo J. 1995;14(8):1646–1654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Marangos P, Carroll J. The dynamics of cyclin B1 distribution during meiosis I in mouse oocytes. Reproduction. 2004;128(2):153–162. [DOI] [PubMed] [Google Scholar]
- [61].Holt JE, Weaver J, Jones KT. Spatial regulation of APCCdh1-induced cyclin B1 degradation maintains G2 arrest in mouse oocytes. Development. 2010;137(8):1297–1304. [DOI] [PubMed] [Google Scholar]
- [62].Kotani T, Yasuda K, Ota R, et al. Cyclin B1 MRNA translation is temporally controlled through formation and disassembly of RNA granules. J Cell Biol. 2013;202(7):1041–1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Yang Y, Yang C-R, Han SJ, et al. Maternal MRNAs with Distinct 3′ UTRs define the temporal pattern of Ccnb1 synthesis during mouse oocyte meiotic maturation. Genes Dev. 2017;31(13):1302–1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Han SJ, Martins JPS, Yang Y, et al. The translation of cyclin B1 and B2 is differentially regulated during mouse oocyte reentry into the meiotic cell cycle. Sci Rep. 2017;7(1). DOI: 10.1038/s41598-017-13688-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Gui L, Homer H. Hec1-dependent cyclin B2 stabilization regulates the G2-M transition and early prometaphase in mouse oocytes. Dev Cell. 2013;25(1):43–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Rattani A, Vinod PK, Godwin J, et al. Dependency of the spindle assembly checkpoint on Cdk1 renders the anaphase transition irreversible. Curr Biol. 2014;24(6):630–637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Touati SA, Buffin E, Cladière D, et al. Mouse oocytes depend on BubR1 for proper chromosome segregation but not for prophase I arrest. Nat Commun. 2015;6(April):6946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Gallant P, Nigg EA. Identification of a novel vertebrate cyclin: cyclin B3 shares properties with both A- and B-type cyclins. Embo J. 1994;13(3):595–605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Sigrist S, Jacobs H, Stratmann R, et al. Exit from mitosis is regulated by drosophila fizzy and the sequential destruction of cyclins A, B and B3. Embo J. 1995;14(19):4827–4838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Lozano J-C, Perret E, Schatt P, et al. Molecular cloning, gene localization, and structure of human cyclin B3. Biochem Biophys Res Commun. 2002;291(2):406–413. [DOI] [PubMed] [Google Scholar]
- [71].Kreutzer MA, Richards JP, De Silva-Udawatta MN, et al. Caenorhabditis elegans cyclin A- and B-type genes: a cyclin A multigene family, an ancestral cyclin B3 and differential germline expression. J Cell Sci. 1995;108(Pt 6):2415–2424. [DOI] [PubMed] [Google Scholar]
- [72].Lozano JC, Vergé V, Schatt P, et al. Evolution of cyclin B3 shows an abrupt three-fold size increase, due to the extension of a single exon in placental mammals, allowing for new protein-protein interactions. Mol Biol Evol. 2012;29:3855–3871. [DOI] [PubMed] [Google Scholar]
- [73].Yuan K, O’Farrell PH. Cyclin B3 is a mitotic cyclin that promotes the metaphase-anaphase transition. Curr Biol. 2015;25:811–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Jacobs HW, Knoblich JA, Lehner CF. Drosophila cyclin B3 is required for female fertility and is dispensable for mitosis like cyclin B. Genes Dev. 1998;12(23):3741–3751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Bourouh M, Dhaliwal R, Rana K, et al. Distinct and overlapping requirements for cyclins A, B and B3 in drosophila female meiosis. G3: Genes Genomes Genet. 2016;6:3711–3724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Matthew MW. Cyclin CYB-3 controls both S-phase and mitosis and is asymmetrically distributed in the early C. Elegans Embryo. Development. 2016;143(17):3119–3127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Deyter GMR, Furuta T, Kurasawa Y, et al. Caenorhabditis Elegans Cyclin B3 is required for multiple mitotic processes including alleviation of a spindle checkpoint–dependent block in anaphase chromosome segregation. PLoS Genet. 2010;6(11):e1001218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Pierron G, Tirode F, Lucchesi C, et al. A new subtype of bone sarcoma defined by BCOR-CCNB3 gene fusion. Nat Genet. 2012;44(4):461–466. [DOI] [PubMed] [Google Scholar]
- [79].Wan-Shan L, Chuang Liao I, Wen M-C, et al. BCOR-CCNB3-positive soft tissue sarcoma with round-cell and spindle-cell histology: a series of four cases highlighting the pitfall of mimicking poorly differentiated synovial sarcoma. Histopathology. 2016;69(5):792–801. [DOI] [PubMed] [Google Scholar]
- [80].Cohen-Gogo S, Cellier C, Coindre J-M, et al. Ewing-like Sarcomas with BCOR-CCNB3 fusion transcript: a clinical, radiological and pathological retrospective study from the Société Française Des Cancers de L’Enfant. Pediatr Blood Cancer. 2014;61(12):2191–2198. [DOI] [PubMed] [Google Scholar]
- [81].Voet MVD, Lorson MA, Srinivasan DG, et al. C. Elegans mitotic cyclins have distinct as well as overlapping functions in chromosome segregation. Cell Cycle. 2009;8(24):4091–4102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Nguyen TB, Manova K, Capodieci P, et al. Characterization and expression of mammalian cyclin B3, a prepachytene meiotic cyclin. J Biol Chem. 2002;277(44):41960–41969. [DOI] [PubMed] [Google Scholar]
- [83].Treen N, Heist T, Wang W, et al. Depletion of maternal cyclin B3 contributes to zygotic genome activation in the ciona embryo. Curr Biol. 2018;28(7):1150–1156.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Zhang T, Qi ST, Huang L, et al. Cyclin B3 controls anaphase onset independent of spindle assembly checkpoint in meiotic oocytes. Cell Cycle. 2015;14(16):2648–2654. [DOI] [PMC free article] [PubMed] [Google Scholar]