

Abstract
Despite considerable advances made in understanding of lung cancer biology, there has been meek improvement in lung cancer treatment outcome with 4% to 5% increase in 5-year survival rates in the last four decades. Underlying problem of lung cancer recurrence and poor prognosis is attributed to the presence of cancer stem cells (CSCs) which possess the potential to differentiate, proliferate and trigger chemo-resistance, tumor progression and metastasis, despite initial elimination of the tumor. To address specific targeting of CSCs, we investigated the effects of a small molecule Verrucarin J (VJ) on lung cancer cell lines A549 and H1793. VJ significantly inhibited cell proliferation of both cell lines, with IC50 values of approximately 10 nM for A549 and 20 nM for H1793 respectively after 48 h of treatment. A549 cell line when treated with VJ, induced cell apoptosis with concomitant down regulation of key CSC specific genes- ALDH1, LGR5, OCT4 and CD133 in a dose-dependent manner. To delineate the molecular mechanism by which VJ targets lung cancer cells and CSCs, we determined the effects of VJ on CSC self-renewal pathways Wnt1/β-catenin and Notch1. Treatment of A549 cell line with VJ inhibited significantly both the signalling pathways, suggesting inhibition of expression of CSC genes by VJ through the inhibition of CSC self-renewal signalling pathways. Taken together, our results suggest that VJ may serve as a potent anticancer drug to target cancer cells and CSCs.
Keywords: lung cancer, lung cancer stem cells, Verrucarin J, cancer stem cell self-renewal, anti-cancer drug
INTRODUCTION
Lung cancer is the leading cause of cancer mortality in the United States for both genders, thus posing a major global health issue [1, 2] with estimated ~2,22,500 new cases diagnosed in 2017 alone [1]. Two major subtypes of lung cancer are classified as non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC). NSCLC is the most common type accounting for 85% of lung cancer [3]. SCLC is a more aggressive form and constitutes 15% of the lung cancer. Last four to five decades witnessed strides in therapeutic advancement in lung cancer, with FDA approvals for several drugs including etoposide, carboplatin, paclitaxel, vinorelbine, Gem, Pem, docetaxel, gefitinib, erlotinib, bevacizumab, and topotecan catering to both SCLC and NSCLC [4]. Surgery, radiation and chemotherapy being the mainstay of primary oncological treatment modality, these strategies have proven to be effective for early stage of lung cancer, but fail to curb the advanced stage disease [5] (https://www.cancer.gov/about-cancer/treatment/types) which could be due to the presence of tumor initiating CSCs.
CSCs constituting a small sub-fraction of the tumor bulk are reported to be a major contributor to onco-therapeutic resistance thus permitting lung cancer recurrence and tumor progression [6–9] with a low 5-year survival rate of only 18% [1]. It has been demonstrated that CSCs resist current treatments due to their activation of anti-apoptotic pathways, increased telomere length, increased membrane transporter activity and CSC ability to migrate and metastasize [10, 11]. A varied repertoire of cell surface and other biomarkers have been reported in lung cancer with CSC specific genes such as CD44, CD90, CD117, BMI-1, EpCAM, FZD, SP and ALDH in NSCLC. Other gene signatures defining pluripotent characteristics such as OCT4, NANOG and SOX2 particularly in CD44+ populations in NSCLC were depicted [12]. Up-regulated expression of mesenchymal markers N-Cadherin and Vimentin, in addition to stem cell markers as a hallmark feature of CD44+/CD90+ sub-population in NSCLC has been reported [13]. Several characteristics including spheroid formation, chemotherapy (cisplatin)-resistance, cellular expansion, and in vivo tumorigenic potential of CSCs in lung cancer leading to poor prognosis and overall poor patient survival have been demonstrated by several investigators [14]. Moreover the stem cell/cancer stem cell compartment within cancer cells is dynamic, fluctuating and transit [15–17] and lack of effective therapeutics faithfully targeting those, poses a serious bottleneck [18].
To date, none of the commonly used treatment modalities including targeted therapies such as immunotherapy have witnessed sufficient success in targeting of lung CSCs, and ultimately avoiding tumor metastasis and relapse. [4, 19].Therefore, there is urgent unmet need for more potent and efficacious drugs that target both tumor bulk cells as well as CSCs to avoid tumor progression and recurrence. Previous study from our lab revealed that a Myrothecium verrucaria fungus metabolite “Verrucarin J” (VJ) effectively targeted ovarian cancer cells and CSCs both [20]. This quest led us to examine the anti-cancer activity of VJ in lung cancer and to understand the underlying molecular mechanisms operating therein. Our studies revealed that VJ specifically inhibited cell proliferation of lung cancer cells, induced cell apoptosis, and suppressed CSC gene expression by inhibiting canonical WNT/β-catenin and Notch1 signaling pathways.
MATERIALS AND METHODS
Human NSCLC cell lines A549 and H1793 were obtained from Dr. Mariusz Ratajczak, University of Louisville. Cell lines were cultured and maintained according to instructions from American Type Culture Collection (ATCC). Verrucarin J was purchased from AnalytiCon Discovery and dimethyl sulfoxide (DMSO) was purchased from MilliporeSigma. Verrucarin J was dissolved in DMSO.
Cell Proliferation Assay:
Lung cancer cell lines, A549 and H1793 growing in log phase were rinsed with phosphate-buffered saline (PBS) (MilliporeSigma) and trypsinized and seeded into 96 well plates as described previously [19]. After 24 h of plating, cells were treated with different concentrations of VJ (final concentrations 0, 1, 5, 10, 20 or 50 nM) and incubated for 24 h, 48 h or 72 h at 37°C in a cell culture incubator. Cell proliferation was measured at each time point using MTT reagent [3-(4, 5‑dimethylthiazol-2-yl)-2H-tetrazolium bromide] from Promega. The formazan product after incubation for 30 to 60 min was analysed by an ELISA reader at 490 nm as described previously [20].
Apoptosis assay by measuring Annexin V using flow cytometry:
Since, both A549 and H1793 cell lines provided similar results in response to VJ with respect to inhibition of cell proliferation, therefore, we selected A549 cell line for subsequent analysis. Apoptosis assays were conducted to determine if VJ induced apoptosis in lung cancer cells. For this purpose A549 cells were plated into T-75 flasks and incubated at 37°C for 24 h. Cells were treated with VJ at concentrations 0, 5, 10, 20 or 50 nM for 24 h and were harvested by centrifugation at 1,500 rpm for 5 min and re-suspended in Annexin V binding buffer from FITC conjugated Annexin V Apoptosis Detection Kit (BD Pharmingen) according to supplier’s instructions. To ensure a single cell suspension and to avoid clumps, cells were passed through a 40 µm nylon mesh and diluted to a final concentration of 107 cells/ml. Each sample was analysed using 100 μL of cell suspension. For each concentration of VJ, an independent control was used. To each sample except controls, 2 μL of Annexin V was added, and incubated for 15 min at room temperature in dark. After completion of incubation, 400 μL of binding buffer was added and immediately analysed by FACS analysis using the FACSCalibur (BD Biosciences). The stained and unstained cells were plotted and analysed using FlowJo software as described previously [20].
Gene expression analysis by real-time PCR:
To determine targeting of CSC genes by VJ, we examined the expression of CSC genes (ALDH1, LGR5, OCT4 and CD133) by real-time PCR. A549 cells were plated into T-75 flasks. After 24 h of plating, cells were treated with VJ to a final concentration of 10 nM or 20 nM. After 48 h of treatment, cells were harvested and total RNA was purified and quantitated using Nanodrop spectrophotometer (Thermo Fisher). First strand cDNA was synthesized from 1 µg of RNA from each sample using iScript cDNA synthesis kit from BioRad. First strand cDNA was subjected to amplification for each gene by real-time PCR (Applied Biosystem) using the standard protocol as described previously [20]. Ct values were normalized with Ct values for GAPDH used as an internal control. The specific primers used for each gene are listed in Table 1.
Table 1.
Primers sequences for various genes employed in real-time PCR
| Gene | Sense (Forward) | Anti-Sense (Reverse) |
|---|---|---|
| WNT1 | CTCTCTTCTTCCCCTTTGTC | AACTCGTGGCTCTGTATCC |
| β -catenin | TGGATGGCCTGCCTCCAGGTGAC | ACCAGCCCCTCGAGCCC |
| TCF-4 | TATGCTCCATCAGCAAGCACTG | TGGATGCAGGCTACAGTAGCTG |
| Notch 1 | TCAGCGGGATCCACTGTGAG | ACACAGGCAGGTGAACGAGTTG |
| HEY1 | TGGATCACCTGAAAATGCTG | TTGTTGAGATGCGAAACCAG |
| LGR5 | GCAAACCTACGTCTGGACAA | TGATGCTGGAGCTGGTAAAG |
| ALDH1 | GCACGCCAGACTTACCTGTC | CCACTCACTGAATCATGCCA |
| Oct-4 | CGCTGGCTTATAGAAGGT | ACAGGTGTCATAAGAATGGATA |
| CD133 | AGTGGCATCGTGCAAACCTG | CTCCGAATCCATTCGACGATAGTA |
| c-Myc | GGACGACGAGACCTTCATCAA | CCAGCTTCTCTGAGACGAGCTT |
| Cyclin D1 | GCATGTTCGTGGCCTCTAAGTA | TCGGTGTAGATGCACACAGC |
| GAPDH | TGATGACATCAAGAAGGTGGT | TCCTTGGAGGCCATGTGGGCC |
Determine the effect of VJ on CSC self-renewal mechanisms:
To determine if VJ regulates the CSC gene expression through the regulation of CSC self-renewal pathways, we studied the effect of VJ on Wnt1/β-catenin and downstream signalling gene TCF-4. Wnt1/β-catenin self-renewal pathway is reported to be one of the major pathway in regulating the expression of CSC genes [21]. In the off state, Wnt1/β-catenin pathway results in phosphorylation of β-catenin by GSK-3β leading to degradation of β-catenin. In contrast, activation of Wnt1/ β-catenin pathway results in inhibition of phosphorylation of β-catenin, which then translocates to the nucleus and activates the expression of TCF4/LEF and other downstream signaling genes including CSC genes [21]. We treated the A549 cells with two concentrations of VJ (10 nM or 20 nM) and determined the expression of Wnt1, β-catenin and TCF-4 genes. To confirm the involvement of GSK-3β in regulation of Wnt1/β-catenin pathway, we treated the A549 cells with various concentrations of GSK-3β-specific inhibitor Chir99021 (MilliporeSigma) alone or in combination with VJ and determined the expression of β-catenin, TCF-4, Cyclin D1 and LGR5 using specific primers (Table 1) by employing real-time PCR.
In an independent experiment, we also determined the effect of VJ on Notch1 (another self-renewal mechanism). We treated the A549 cells with VJ (10 nM or 20 nM) for 24 h and determined the expression of Notch1 and its downstream signalling gene Hey1 using specific primers in real-time PCR.
Statistical analysis:
Student’s t-test was performed to assess the significance between controls and the treated groups. Difference between the two groups was considered statistically significant at p ≤ 0.05, and highly significant at p ≤ 0.001. Standard deviation (SD) of three independent experiments was calculated using GraphPad software (version 4.03, San Diego California USA, http://www.graphpad.com)
RESULTS
Inhibition of cell proliferation in A549 and H1793 cell lines by VJ:
The potency and efficiency of VJ to target lung cancer cells (A549 and H1793) was tested by performing cell viability/proliferation assays. Treatment of cells with VJ at concentrations of 0, 1, 2, 5, 10, 20, or 50 nM for 24 h, 48 h or 72 h showed a significant dose- and time-dependent inhibition of cell proliferation of both lung cancer cell lines (Figs. 1, 2). A549 and H1793 cell lines revealed IC50 values of approximately 10 nM and 20 nM respectively after 48 h of treatment. These results suggest profound anti-proliferative effect of VJ on lung cancer cells thus highlighting its anti-cancer potential.
Figure 1. Effect of VJ on cell viability of A549 cells:
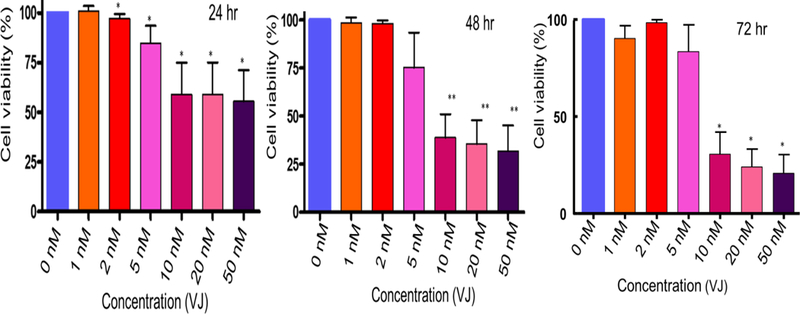
Lung cancer cells (A549) were treated with various concentrations of VJ for 24, 48 or 72 hours. Cell viability was determined by using MTT assays. Data shown is mean ± S.D (standard deviation) for three independent experiments. * represents significant (P ≤ 0.05) and ** represent highly significant (p ≤ 0.001). IC50 value after 48 hours of treatment with VJ was 10 nM.
Figure 2. Effect of VJ on cell viability of H1793 cells:
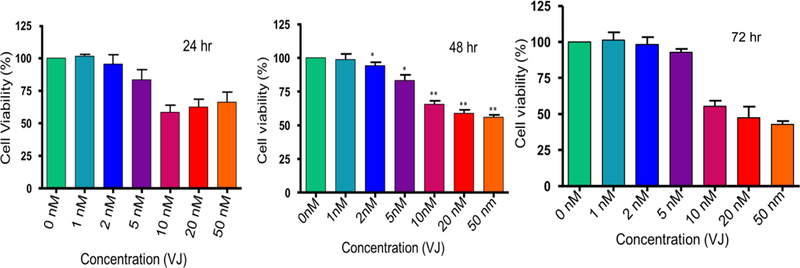
Lung cancer cells (H1793) were treated with various concentrations of VJ for 24, 48, or 72 hours. The data shown is mean ± S.D (standard deviation) of three independent experiments. * represents significant (p ≤ 0.05) and ** represent highly significant (p ≤ 0.001) values. IC50 value after 48 hours of treatment was 20 nM.
Induction of apoptosis in A549 cells by VJ:
While apoptosis or programmed cell death is a process that regulates cellular development. Cancer cells often reveal deregulation of such apoptotic pathways that allow uncontrolled cell proliferation [22]. Essentially anti-cancer drugs are known to induce apoptosis in cancer cells, but fail to stop tumor progression by sparing tumor initiating CSCs. To determine the induction of apoptosis by VJ, we performed apoptosis assays as described previously [20]. Treatment of A549 cells with various concentrations of VJ showed a significant increase in the percentage of apoptotic cells. VJ at a concentration of 10 nM was found to be saturated and resulted in approximately 22% apoptotic (FITC labeled) cells after 24 h of treatment detected by flow cytometry. These results (Fig. 3) suggested that VJ induced apoptosis in lung cancer cells (A549) in a dose-dependent manner similar to that in ovarian cancer cells [20].
Figure 3. Effect of VJ on apoptosis of lung cancer cells (A549):

Induction of apoptosis after 24 hour of treatment of lung cancer cells (A549) with VJ at concentrations of 0, 5, 10, 20 and 50 nM was examined by A) FACS analyses. Data in B represents quantitative analysis. Cells gated in blue (in A) represent apoptotic cells (Annexin-FITC+/PI+/−). The data shown is mean ± S.D. (std. deviation) of two independent experiments plotted using GraphPad. * indicates significance (p ≤ 0.05).
Inhibition of expression of CSC genes:
To determine if VJ targets CSCs in lung cancer, we determined the expression of CSC genes (ALDH1, LGR5, OCT4 and CD133) in lung cancer cell line (A549) by exposing the cells to VJ at final concentrations of 10 nM or 20 nM for 48 h. RNA was purified and subjected to gene amplification by employing real-time PCR. As indicated in Fig. 4, VJ significanlty inhibited the expression of ALDH1, LGR5, OCT4 and CD133 at both the concentrations. VJ concentration of 20 nM was found to be more potent in inhibiting the expression of ALDH1 expression compared to 10 nM. These results clearly suggest that VJ targets CSC populations by regulating expression of CSC genes at transcriptional level.
Figure 4. Effect of VJ on expression of CSC genes:
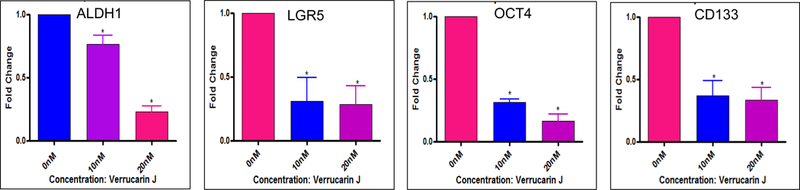
Lung cancer cells (A549) were treated with 10 nM or 20 nM of VJ for 48 hour. Total RNA was purified and subjected to real-time PCR for amplification of CSCs genes using specific primers. The data shown is fold changes compared to control (vehicle treated cells) and is mean ± S.D for three independent experiments. * represents significant (p ≤ 0.05).
Inhibition of expression of CSC genes and self-renewal pathways by VJ:
Differentiation and self renewal are the basic characteristics of both stem cells and CSCs. Some of these self-renewal mechanisms include Wnt1/β-catenin, Notch1, Sonic Hedgehog (SHH), STAT3 and NFkβ [23–29]. Several signaling pathways form complex signaling networks to contribute to stem cell diversity during development of organism as well in CSC biology by co-operative inter pathway interaction of various ligands and gene targets [30, 31]. Wnt1/β-catenin signaling pathway plays critical roles in development, tissue homoeostasis and self-renewal of stem cells/CSCs. To determine if the regulation of CSC genes (shown in Fig. 4) is through the regulation of Wnt1/β-catenin pathway, we determined the effect of Wnt1, β-catenin and downstream effector gene (TCF-4) (Fig. 5) using specific primers (Table 1) employed in real-time PCR analysis. Our results revealed that VJ inhibited Wnt1, β-catenin and TCF-4 (transcription) factor expression in a dose dependent manner, suggesting that inhibition of expression of ALDH1, LGR5, OCT4 and CD133 CSC genes could be achieved through the inhibition of Wnt1/β-catenin self-renewal pathway.
Figure 5. Effect of VJ on Wnt1/β-catenin signaling:
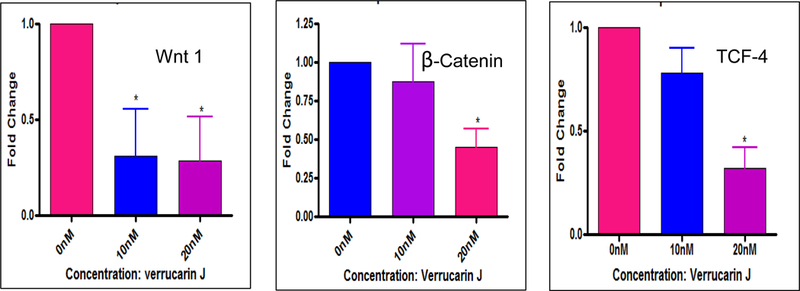
Lung cancer cells (A549) were treated with 10 nM or 20 nM of VJ for 48 hour. Gene expression was performed using real-time PCR using the specific primers for each gene. Wnt1/β-catenin signaling genes were evaluated for their expression in response to VJ treatment. * represents significance (p ≤ 0.05).
As depicted in Fig. 6, Wnt1/β-catenin signaling pathway, in its off state shows APC, Axin, GSK-3β and β-catenin complex formation which leads to phosphorylation and subsequent degradaton of β-catenin. In the, on state however, β-catenin fails to undergo phosphyrylation and is translocated to the nucleus in order to regulate the transcription of TCF-4/LEF (transcription factors) and other genes including CSC genes, Cyclin D1 and c-Myc [21]. GSK-3 (glycogen synthase kinase 3) is known to regulate transcription and cell proliferation via β-catenin and the Wnt canonical pathway [32]. Hence, blocking the acivity of GSK-3β with specific inhibitor CHIR99021 revealed upregulation of β-catenin regulatory genes (TCF-4, c-Myc, Cyclin D1 and LGR5) (Fig. 7) thus confirming the importance of GSK-3β in regulating Wnt1/β-catenin signaling pathway. To delineate if the effect of VJ is mediated through the regulation of GSK-3β activity, we treated A549 cells with 10 nM VJ, 1 µM or 3 µM of CHIR99021, 1 µM CHIR99021 plus 10 nM VJ or 3 µM CHIR99021 plus 10 nM VJ. As expected, VJ induced a significant inhibition of expression of TCF-4 and Cyclin D1, whereas CHIR99021 upregulated the expression of both TCF-4 and Cyclin D1 (Fig. 8). Treatment of A549 cells with VJ in combination with CHIR99021 attenuated the effect of CHIR99021, suggesting that regulation of CSC genes by VJ is achieved through the inhibition of Wnt1/β-catenin self-renewal pathway. These results are very interesting and to the best of our knowledge, the first observation being reported, indicating the regulation of CSC population via the Wnt1/β-catenin signaling mechanism by VJ thus strongly crediting its anti-cancer property being regulated via self-renewal pathways.
Figure 6: Schematic of Wnt1/β-catenin signaling pathway:
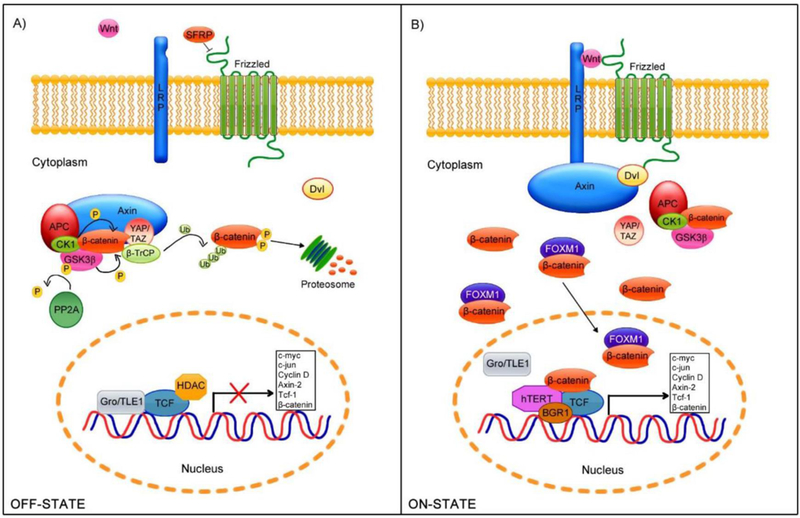
A. In the absence of stimuli (off-state), β-catenin undergoes phosphorylation at specific residues by the GSK3β and CK1 kinases. Phosphorylated β-catenin undergoes proteosomal degradation, therefore does not translocate to the nucleus. B. In the presence of Wnt ligands (on-state), LRP5/6 and Fz, allowing Axin-Dv1 binding and the disassembly of the β-catenin degradation complex. β-catenin is released in the cytoplasm and translocated to the nucleus. In the nucleus β-catenin binds to TCF/LEF and activates several downstream genes. The figure is reproduced from Bello et al [72]. Appropriate credit is provided by provided the reference as per Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Figure 7. Effect of GSK3β inhibitor CHIR99021 on downstream signaling genes of Wnt1/β-catenin pathway and LGR5:
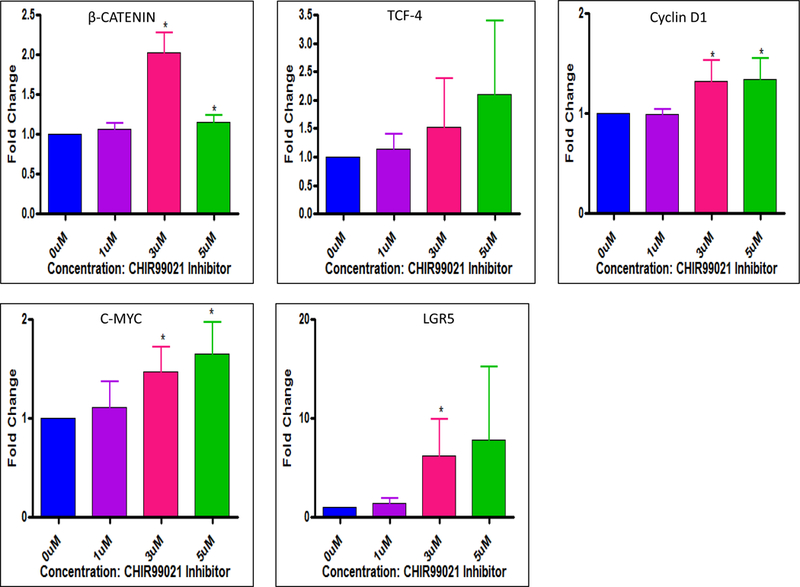
Lung cancer cells (A549) were treated with various concentrations of GSK3β inhibitor CHIR99021 for 48 h. Effect of VJ on downstream signaling genes (β-catenin, TCF-4, and Cyclin D1, c-Myc) and LGR5 was evaluated using real-time PCR and specific primers for each gene. CHIR99021 at a concentration of 3 μM was found to be significant. * indicates significance (p ≤0.05). The data shown is mean ± S.D for three independent experiments. * indicates significance (p ≤0.05).
Figure 8. Effect of VJ (10 nM), GSK3β inhibitor CHIR99021 (1 µM, 3 µM) and combination of GSK3β inhibitor CHIR99021 and VJ on downstream signaling genes (TCF-4 and Cyclin D1):
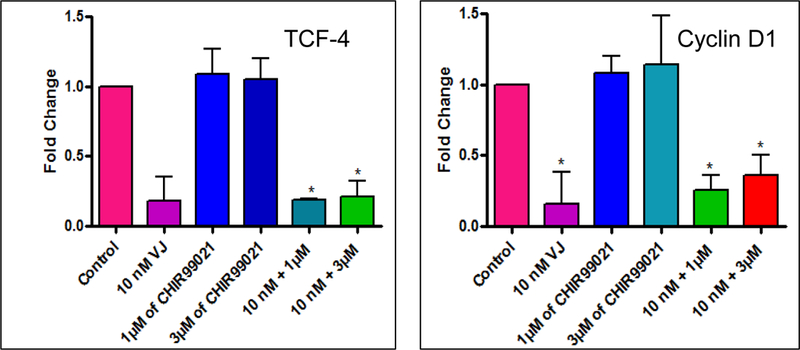
Lung cancer cells (A549) were treated with various concentrations of GSK3β inhibitor CHIR99021 or VJ or combination of both for 48 h. Expression of downstream signaling genes TCF-4 and Cyclin D1 was evaluated by real-time PCR using specific primers. * indicates significance (p ≤0.05).
Inhibition of Notch1 self-renwal pathway by VJ:
Notch1 signaling pathway is associated with regulation of cell fate at several distinct developmental stages and is one of the most important pathways involved in cancer initiation and progression and CSC regulation in a variety of human cancers [33–39]. Notch1 signaling is also involved in the process of EMT and hence responsible for conferring stemness property and drug resistance [40, 41]. In addition, deregulation of the Notch pathway is known to promote tumorigenesis and upregulation of genes such as Notch1 and Hey1 [42]. Treatment of A549 cell line with VJ resulted in a significant down regulation of expression of Notch1 and its downstream signaling gene Hey1 (Fig. 9), suggesting regulation of Notch1 signaling pathway by VJ. However, the comprehensive expression pattern of Notch1 signaling genes and their relationship with CSCs remains yet to be determined. A significant decrease in expression of CSC specific genes by VJ suggested that VJ could certainly target both cancer cells and CSCs.
Figure 9. Effect of VJ on Notch1 signaling:
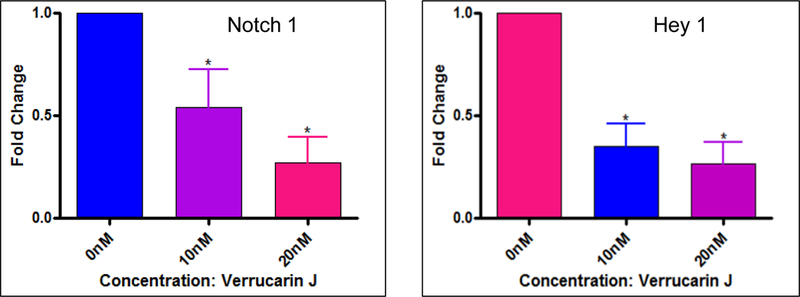
Lung cancer cells (A549) were treated with VJ at a final concentration of 10 nM or 20 nM for 48 hour. The data presented is gene expression (fold change) analyzed by real-time PCR employing specific primers for each gene. * represents significance (p ≤ 0.05).
DISCUSSION
CSCs within a tumor share their enormous capacity for self-renewal, regeneration and differentiation with the normal stem cells in adult tissues, embryonic stem cells and induced pluripotent stem cells [14, 43] besides core attributes of chemo- and radio-resistance and those that contribute to malignant progression, metastasis and cancer relapse [44]. Besides, tumor micro-environment, stem cell related signal transduction pathways including Wnt/β-catenin, Hedgehog, Notch are implicated in the phenomenon of EMT and metastasis, while CSCs are closely associated with these cellular attributes [19, 44–46]. Moreover conventional onco-therapy modalities and regimes have proven obsolete in terms of complete disease free survival while avoiding relapse. Hence, targeting of these cardinal signaling pathway components while considering the CSC subsets is of prime importance [47].
Under this backdrop of CSC biology in lung cancer and inadequate information regarding anti-metastatic effect of naturally occurring small molecule inhibitors/drugs/compounds targeting CSCs, it was envisaged to evaluate VJ over conventional chemotherapeutic drugs [48] known to impart chemo-resistance. An in vitro model comprising of advanced malignant phenotype of NSCLC A549 and H1793 cells were employed initially to determine cell viability under the influence of VJ using MTT assays and to assess their IC50 values. IC50 of VJ was ~10 nM for A549 and 20 nM for H1793 respectively after 48 h of treatment (Figs. 1, 2). Subsequently, in order to reveal how VJ reduced proliferation of these cells in a dose-and time-dependent manner, cell apoptosis was assessed by Annexin V/PI method using flow cytometry based analysis. VJ very prominently induced inhibition of cell proliferation and reduced cell viability in lung cancer cell lines A549 and H1793 through the induction of apoptosis. Increasing concentrations of VJ increased the number of apoptotic cells which substantiated the anti-cancer property of VJ (Fig. 3). Further VJ resulted in a dose dependent suppression of various CSC specific mRNA transcripts including ALDH1, LGR5, OCT4 and CD133 (Fig. 4) as assessed by real-time PCR, thus suggesting the targeting of varied CSC populations. These results are consistent with our earlier published results for ovarian cancer [20].
Interestingly our results revealed down-regulation of key CSC genes, and hence we were intrigued to investigate relevant self-renewal pathways such as Wnt1/β-catenin and Notch1, besides the expression of CSC specific gene transcripts representing varied CSC subsets in A549 lung cancer cells. Recently, the significance of CSC specific biomarkers such as ALDH1, OCT4, Sox2, Nanog, BMI1, CD133, CD44, CD166, ABCB5, AGR2, TAZ in elucidating the clinical characteristics manifested by tumors at different stages of tumorigenesis and metastasis and as prognostic determinants in multiple tumor types was reported [49]. ALDH1 is a metabolic marker involved in retinoid signaling, and is related to stemness, self-renewal, differentiation and self-protection of normal stem and CSCs both and possesses role in drug resistance [50, 51] and radio-resistance [52]. It is expressed in putative lung epithelial stem cell niches and implicated in poor overall survival outcome and disease-free survival in lung cancer and advanced stage of the disease [53]. CD133 is involved in glucose and transferrin uptake, autophagy, membrane–membrane interaction, matrix metalloproteinase functions, self-renewal, proliferation and differentiation during damage repair and responsible for tumor metastasis, chemo-resistance, radio-resistance and recurrence [54]. Besides Nanog and Sox2, OCT4 is core homeodomain transcription factor of POU family required to maintain stemness, self-renewal and pluripotency of embryonic stem cells, adult tissue stem cells, and essential for somatic cell reprogramming [55]. Overexpression of OCT4 is associated with tumorigenicity, metastasis and relapse in certain cancers [19, 56]. OCT4 expressing bronchio-alveolar stem cell-like cancer cells detected in lung adenocarcinoma reveal worse clinical outcomes [57]. LGR5, a member of G-protein-coupled receptor superfamily is a CSC surface marker initially reported in colon cancer and a target gene of Wnt signaling pathway and is implicated in tumor growth, invasion and poor prognosis and recently detected in a subset of lung adenocarcinoma [58]. Taken together, inhibition of expression of ALDH1, LGR5, OCT4 and CD133 by VJ suggests that it may possess the potential to inhibit drug-resistance and recurrence of cancer, through the regulation of expression of CSC genes, hence CSC populations. Recent research has witnessed unprecedented increase in the exploration of phytochemicals in their potential to directly influence CSC activity thus posing as attractive targets for developing CSC specific therapeutics. Currently no clarity prevails with respect to the molecular mechanisms involved and the identity of heterogeneous CSC populations being targeted which collectively warrant further comprehensive investigation [59–63].
Wnt/β-catenin signaling pathway is crucial for cell development, proliferation, differentiation, tissue homeostasis, self-renewal of stem cells and CSCs. Activation of Wnt1/β-catenin pathway steers the β-catenin from cytoplasm to the nucleus which further binds with T-cell factor/Lymphoid enhancer factor (TCF4/LEF) in the nucleus to regulate the expression of related target genes including Cyclin D1, c-Myc and several CSC specific genes [21, 64]. A schematic representation (Fig. 6) depicts crucial steps involved in the signaling mechanism and their respective on/off state functions. Wnt activation is reported in various malignancies including breast, lung and hematopoietic system, and implicated in chemo-resistance and tumor recurrence. Overexpression of c-Myc and Cyclin D1 is implicated in tumorigenesis. On account of the cross talk of Wnt signal transduction pathway with that of Notch and Sonic Hedgehog, Wnt signaling effectors and inhibitors may provide huge implications for therapeutic interventions in various cancers [21]. To define the mechanism involved in current study, a GSK-3 inhibitor (CHIR99021) was employed, whereas in the combination treatment group, cells were exposed first to the inhibitor at concentrations of 1 µM or 3 µM followed by VJ at concentration of 10 nM after 2 hours to prevent β-catenin from being targeted which allowed β-catenin to accumulate and enter the nucleus for transcription to be activated (GSK-3 is an inhibitor of Wnt pathway). Results revealed that increased concentration of GSK-3 inhibitor (i.e. inhibitor of Wnt pathway inhibitor), increased the expression levels of WNT1 as well as key WNT target genes downstream, including c-Myc, TCF-4, β-catenin, Cyclin D1 and LGR5 (Fig. 7). To reiterate, combination group of lung cancer cell response reflected a significant destruction of self-renewal mechanism and increased vulnerability/sensitivity to VJ in vitro (Fig 8), suggesting that VJ may inhibit CSC gene expression through the regulation of Wnt1/β-catenin self-renewal pathway.
Another important signaling pathway responsible for regulating self-renewal of stem cells and CSCs is “Notch1 signaling” which is known to regulate cell fate determination, specification, proliferation, differentiation, apoptosis, stem cell maintenance, homeostasis of multicellular organism and angiogenesis in normal physiology [65–67]. Notch1 signaling regulates tumorigenesis, progression and therapeutic resistance of NSCLC and is linked with CSC self-renewal and angiogenesis. Its up-regulation is associated with chemo- and radio-therapy resistance, EMT, tumor progression and poor prognosis in NSCLC [68]. Hey1 being a Hes-related transcriptional factor is a downstream effector gene in regulating Notch signaling pathway [64]. Unscheduled cell cycle division in lung cancer progenitor cells directed by Cyclin D1 activation is significantly associated with NSCLC tumor development [69, 70]. Besides Wnt1/β-catenin pathway, a prominent inhibition of Notch1 and Hey1 genes in response to VJ treatment was noted in current study (Fig. 9). VJ thus reflects potential to be utilized further as a potent chemotherapeutic drug either alone and/or in combination to target CSCs thus strongly validating that Wnt1/β-catenin as well as Notch1 self-renewal pathways both were involved and were activated. Targeting of specific CSC subsets by the small molecule drug VJ is the mainstay of current study and will pave novel paths in future towards personalized and precision medicine for lung cancer patients with more effective outcome. Recently bioengineered tumor models have initiated a revolution in anti-cancer drug validation process by serving as predictive models for testing drug safety and efficacy [71]. Thus the current interesting findings in addition to those in ovarian cancer [20] may lay the foundation for establishing 3D tumor model platforms for CSC targeted drug screening and discovery.
Acknowledgments
Funding: This study was funded by a grant from NIH/NCI UO1CA2177798 (subcontract, SSK).
Footnotes
Compliance with Ethical Standards:
Competing interests:
The Authors declare that they have no competing interest.
REFERENCES
- 1.Siegel RL, Miller KD and Jemal A (2017). Cancer statistics, 2017. CA: A Cancer Journal for Clinicians, 67, 7–30. [DOI] [PubMed] [Google Scholar]
- 2.Xu B, Chen Q, Yue C, et al. Prognostic value of IL-6R mRNA in lung adenocarcinoma and squamous cell carcinoma. (2018). Oncology Letters, 16(3), 2935–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tsvetkova E, and Goss GD (2012). Drug Resistance and Its Significance for Treatment Decisions in Non-Small-Cell Lung Cancer. Current Oncology, 19 Suppl 1, S45–S51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Johnson DH, Schiller JH, Bunn PA Jr. (2014). Recent clinical advances in lung cancer management. J Clin Oncol, 32(10), 973–82. [DOI] [PubMed] [Google Scholar]
- 5.National Cancer Institute. Types of Cancer Available at: https://www.cancer.gov/about-cancer/treatment/types. Accessed on August 13, 2018.
- 6.MacDonagh L, Gray SG, Breen E, Cuffe S, Finn SP, O’Byrne KJ, Barr MP (2016). Lung cancer stem cells: The root of resistance. Cancer Letters, 372(2), 147–56. [DOI] [PubMed] [Google Scholar]
- 7.Shanker M, Willcutts D, Roth JA, & Ramesh R (2010). Drug resistance in lung cancer. Lung Cancer (Auckland, N.Z.), 1, 23–36. [PMC free article] [PubMed] [Google Scholar]
- 8.Arriagada R, Bergman B, Dunant A, Le Chevalier T, Pignon JP, Vansteenkiste J, International Adjuvant Lung Cancer Trial Collaborative Group. (2004). Cisplatin-based adjuvant chemotherapy in patients with completely resected non-small-cell lung cancer. N Engl J Med, 350(4), 351–60. [DOI] [PubMed] [Google Scholar]
- 9.Christen RD, Isonishi S, Jones JA, Jekunen AP, Hom DK, Kröning R, Gately DP, Thiebaut FB, Los G, Howell SB (1994). Signaling and drug sensitivity. Cancer Metastasis Rev, 13(2), 175–89. [DOI] [PubMed] [Google Scholar]
- 10.Lathia JD, Venere M, Rao MS, & Rich JN (2011). Seeing is believing: are cancer stem cells the Loch Ness monster of tumor biology? Stem Cell Reviews and Reports, 7(2), 227–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Prabavathy D, Swarnalatha Y, Ramadoss N (2018). Lung cancer stem cells-origin, characteristics and therapy. Stem Cell Investigation, 5:6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Leung EL, Fiscus RR, Tung JW, Tin VP, Cheng LC, Sihoe AD, Fink LM, Ma Y, … Wong MP (2010). Non-small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties. PloS One, 5(11), e14062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang P, Gao Q, Suo Z, Munthe E, et al. (2013). Identification and characterization of cells with cancer stem cell properties in human primary lung cancer cell lines. PloS One, 8(3), e57020. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 14.Hardavella G, George R, & Sethi T (2016). Lung cancer stem cells-characteristics, phenotype. Translational lung cancer research, 5(3), 272–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li Y, & Laterra J (2012). Cancer stem cells: distinct entities or dynamically regulated phenotypes?. Cancer research, 72(3), 576–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sellers ZP, Schneider G, Bujko K, Suszynska M, Pedziwiatr D (2017). Do Cancer Cell Lines Have Fixed or Fluctuating Stem Cell Phenotypes? – Studies with the NTera2 Cell Line. Stem Cell Reviews and Reports, 13(5), 603–610. [DOI] [PubMed] [Google Scholar]
- 17.Gunjal P, Pedziwiatr D, Ismail AA, Kakar SS, Ratajczak MZ (2015). An emerging question about putative cancer stem cells in established cell lines-are they true stem cells or a fluctuating cell phenotype? J Cancer Stem Cell Res 3 pii: e1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sell S (2007). Cancer and stem cell signalling: a guide to preventive and therapeutic strategies for cancer stem cells. Stem Cell Rev, 3(1), 1–6. [DOI] [PubMed] [Google Scholar]
- 19.Maiuthed A, Chantarawong W, Chanvorachote P (2018). Lung Cancer Stem Cells and Cancer Stem Cell-targeting Natural Compounds. Anticancer Res, 38(7):3797–809. [DOI] [PubMed] [Google Scholar]
- 20.Carter K, Rameshwar P, Ratajczak MZ, & Kakar SS (2017). Verrucarin J inhibits ovarian cancer and targets cancer stem cells. Oncotarget, 8(54), 92743–92756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Krishnamurthy N, Kurzrock R (2018). Targeting the Wnt/β-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat Rev 62, 50–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mohammad RM, Muqbil I, Lowe L, et al. (2015). Broad targeting of resistance to apoptosis in cancer. Semin Cancer Biol, 35 Suppl(0):S78–S103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Koduru S, Kumar R, Srinivasan S, Evers MB, Damodaran C (2010). Notch-1 inhibition by Withaferin-A: a therapeutic target against colon carcinogenesis. Mol Cancer Ther, 9(1), 202–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Reya T, Clevers H (2005). Wnt signaling in stem cells and cancer. Nature 434(7035), 843. [DOI] [PubMed] [Google Scholar]
- 25.Liu S, Dontu ID, Mantle S, Patel NS, Ahn KW, Jackson, et al. (2006) Hedgehog signaling and Bmi-1 regulate self-renewal of normal and malignant human mammary stem cells. Cancer Res, 66, 6063–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dontu G, Jackson KW, McNicholas E, Kawamura MJ, Abdallah WM, Wicha MS (2004). Role of Notch signaling in cell-fate determination of human mammary stem/progenitor cells. Breast Cancer Res 6, 605–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Giles RH, van ESJH, Clevers H (2003). Caught up in a Wnt storm: wnt signaling in cancer. Biochim Biophys Acta, 1653, 1–24. [DOI] [PubMed] [Google Scholar]
- 28.Abubaker K, Luwor RB, Zhu H, McNally O, Quinn MA, Burns CJ, Thompson EW, Findlay JK, Ahmed N (2014). Inhibition of the JAK2/STAT3 pathway in ovarian cancer results in the loss of cancer stem celllike characteristics and a reduced tumor burden. BMC Cancer, 14, 317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chefetz I, Holmberg JC, Alvero AB, Visintin I, & Mor G (2011). Inhibition of Aurora-A kinase induces cell cycle arrest in epithelial ovarian cancer stem cells by affecting NFĸB pathway. Cell cycle (Georgetown, Tex.), 10(13), 2206–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang Z, Li B, Zhou L, Yu S, Su Z, Song J, Sun Q, Sha O, Wang X, Jiang W, Willert K, Wei L, Carson DA, … Lu D (2016). Prodigiosin inhibits Wnt/β-catenin signaling and exerts anticancer activity in breast cancer cells. Proceedings of the National Academy of Sciences of the United States of America, 113(46), 13150–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.MacDonald BT, Tamai K, & He X (2009). Wnt/beta-catenin signaling: components, mechanisms, and diseases. Developmental cell, 17(1), 9–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Patel S, & Woodgett J (2008). Glycogen synthase kinase-3 and cancer: good cop, bad cop?. Cancer cell, 14(5), 351–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Suman S, Das TP, Damodaran C (2013). Silencing NOTCH signaling causes growth arrest in both breast cancer stem cells and breast cancer cells. Br J Cancer, 109, 2587–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dontu G, Jackson KW, McNicholas E, Kawamura MJ, Abdallah WM, & Wicha MS (2004). Role of Notch signaling in cell-fate determination of human mammary stem/progenitor cells. Breast cancer research : BCR, 6(6), R605–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang M, Wang J, Wang L, Wu L, Xin X (2010). Notch1 expression correlates with tumor differentiation status in ovarian carcinoma. Med Oncol, 27,1329–35. [DOI] [PubMed] [Google Scholar]
- 36.Hori K, Sen A, Artavanis-Tsakonas S (2013). Notch signaling at a glance. J Cell Sci, 26, 2135–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hopfer O, Zwahlen D, Fey MF, Aebi S (2005). The Notch pathway in ovarian carcinomas and adenomas. Br J Cancer, 93, 709–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gao Y, Rankin GO, Tu Y, & Chen YC (2015). Theaflavin-3, 3’-digallate decreases human ovarian carcinoma OVCAR-3 cell-induced angiogenesis via Akt and Notch-1 pathways, not via MAPK pathways. International journal of oncology, 48(1), 281–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yabuuchi S, Pai SG, Campbell NR, de Wilde RF, De Oliveira E, Korangath P, Streppel MM, Rasheed ZA, Hidalgo M, Maitra A, … Rajeshkumar NV (2013). Notch signaling pathway targeted therapy suppresses tumor progression and metastatic spread in pancreatic cancer. Cancer letters, 335(1), 41–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang L, Sha J, Yang G, Huang X, Bo J, & Huang Y (2017). Activation of Notch pathway is linked with epithelial-mesenchymal transition in prostate cancer cells. Cell cycle (Georgetown, Tex.), 16(10), 999–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kong D, Banerjee S, Ahmad A et al. (2010). Epithelial to mesenchymal transition is mechanistically linked with stem cell signatures in prostate cancer cells. PLoS ONE 5:e12445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yuan X, Wu H, Han N, et al. , (2014). Notch signaling and EMT in non-small cell lung cancer: biological significance and therapeutic application. Journal of Hematology & Oncology, 7, 87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ma I & Allan AL (2011). The Role of Human Aldehyde Dehydrogenase in Normal and Cancer Stem Cells. Stem Cell Rev and Rep, 7: 292. [DOI] [PubMed] [Google Scholar]
- 44.Marquardt S, Solanki M, Spitschak A, Vera J, Pützer BM (2018). Emerging functional markers for cancer stem cell-based therapies: Understanding signaling networks for targeting metastasis. Semin Cancer Biol, June 30 pii: S1044–579X(18)30018-X. [DOI] [PubMed] [Google Scholar]
- 45.Moustakas A and Heldin CH (2007). Signaling networks guiding epithelial–mesenchymal transitions during embryogenesis and cancer progression. Cancer Sci, 98(10),1512–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Vermeulen L, De Sousa EMF, van der Heijden M, et al. , (2010). Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat Cell Biol, 12, 468–76. [DOI] [PubMed] [Google Scholar]
- 47.Abetov D, Mustapova Z, Saliev T, Bulanin D, Batyrbekov K, Gilman CP (2015). Novel Small Molecule Inhibitors of Cancer Stem Cell Signaling Pathways. Stem Cell Reviews and Reports, 11(6), 909–18. [DOI] [PubMed] [Google Scholar]
- 48.Kornakiewicz A, Czarnecka AM, Khan MI, Krasowski P, Kotrys AV, Szczylik C (2018). Effect of Everolimus on Heterogenous Renal Cancer Cells Populations Including Renal Cancer Stem Cells. Stem Cell Reviews and Reports, 14(3), 385–97. [DOI] [PubMed] [Google Scholar]
- 49.Curtarelli RB, Gonçalves JM, Dos Santos LGP, Savim MG, Nör JE, Mezzomo LAM, Rodríguez Cordeiro MM (2018). Expression of Cancer Stem Cell Biomarkers in Human Head and Neck Carcinomas: a Systematic Review. Stem Cell Reviews and Reports, 14(6), 769–84. [DOI] [PubMed] [Google Scholar]
- 50.Tomita H, Tanaka K, Tanaka T and Hara A (2016). Aldehyde dehydrogenase 1A1 in stem cells and cancer. Oncotarget, 7(10), 11018–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Suresh R, Ali S, Ahmad A, Philip PA and Sarkar FH (2016). The role of cancer stem cells in recurrent and drug-resistant lung cancer. Adv Exp Med Biol, 890, 57–74. [DOI] [PubMed] [Google Scholar]
- 52.Miyata T, Oyama T, Yoshimatsu T, Higa H, Kawano D, Sekimura A, Yamashita N, So T, Gotoh A (2017). The Clinical Significance of Cancer Stem Cell Markers ALDH1A1 and CD133 in Lung Adenocarcinoma. Anticancer Res, 37(5), 2541–47. [DOI] [PubMed] [Google Scholar]
- 53.Li X, Wan WL, Geng J, Wu CL and Bai X (2012). Aldehyde dehydrogenase 1A1 possesses stem-like properties and predicts lung cancer patient outcome. J Thorac Oncol, 7, 1235–45. [DOI] [PubMed] [Google Scholar]
- 54.Li Z CD133. (2013). A stem cell biomarker and beyond. Exp Hematol Oncol, 2(1), 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Liedtke S, Stephan M, Kögler G (2008). Oct4 expression revisited: potential pitfalls for data misinterpretation in stem cell research. Biol Chem, 389(7):845–50. [DOI] [PubMed] [Google Scholar]
- 56.Hayashi H, Arao T, Togashi Y, Kato H, Fujita Y, et al. , and Nishio K (2015). The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene, 34, 199–208. [DOI] [PubMed] [Google Scholar]
- 57.Zhang X, Han B, Huang J, Zheng B, Geng Q, Aziz F, Dong Q (2010). Prognostic significance of OCT4 expression in adenocarcinoma of the lung. Jpn J Clin Oncol, 40(10), 961–6. [DOI] [PubMed] [Google Scholar]
- 58.Ryuge S, Sato Y, Jiang SX, Wang G, Kobayashi M, Nagashio R, Katono K, Iyoda A, Satoh Y, Masuda N (2013). The clinicopathological significance of LGR5 expression in lung adenocarcinoma. Lung Cancer, 82, 143e148. [DOI] [PubMed] [Google Scholar]
- 59.Dandawate PR, Subramaniam D, Jensen RA, & Anant S (2016). Targeting cancer stem cells and signaling pathways by phytochemicals: Novel approach for breast cancer therapy. Seminars in cancer biology, 40–41, 192–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Oh J, Hlatky L, Jeong YS, & Kim D (2016). Therapeutic Effectiveness of Anticancer Phytochemicals on Cancer Stem Cells. Toxins, 8(7), 199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Singh A, Sharma N, Ghosh M, Park YH Jeong DK (2017). Emerging importance of dietary phytochemicals in fight against cancer: Role in targeting cancer stem cells. Crit Rev Food Sci Nutr, 57(16), 3449–63. [DOI] [PubMed] [Google Scholar]
- 62.Zhu JY, Yang X, Chen Y, Jiang Y, Wang SJ, Li Y, Wang XQ, Meng Y, Zhu MM, Ma X, Huang C, Wu R, Xie CF, Li XT, Geng SS, Wu JS, Zhong CY, Han HY (2017). Curcumin Suppresses Lung Cancer Stem Cells via Inhibiting Wnt/β-catenin and Sonic Hedgehog Pathways. Phytother Research, 31(4), 680–88. [DOI] [PubMed] [Google Scholar]
- 63.Zhang L, Wen X, Li M, Li S, Zhao H (2018). Targeting cancer stem cells and signaling pathways by resveratrol and pterostilbene. Biofactors, 44(1), 61–68. [DOI] [PubMed] [Google Scholar]
- 64.Rapp J, Jaromi L, Kvell K, Miskei G, Pongracz JE (2017). WNT signaling - lung cancer is no exception. Respir Research, 18(1),167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Westhoff B, Colaluca IN, D’Ario G, et al. (2009). Alterations of the Notch pathway in lung cancer. Proc Natl Acad Sci U S A, 106(52), 22293–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hassan KA, Wang L, Korkaya H, et al. (2013). Notch pathway activity identifies cells with cancer stem cell-like properties and correlates with worse survival in lung adenocarcinoma. Clin Cancer Research, 19(8),1972–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Venkatesh V, Nataraj R, Thangaraj GS, et al. (2018). Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig, 5, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zou B, Zhou XL, Lai SQ, & Liu JC (2018). Notch signaling and non-small cell lung cancer. Oncology letters, 15(3), 3415–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wang P, Gao Q, Suo Z, Munthe E, Solberg S, Ma L, Wang M, Westerdaal NA, Kvalheim G, Gaudernack G (2013). Identification and characterization of cells with cancer stem cell properties in human primary lung cancer cell lines. PloS one, 8(3), e57020. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 70.Casimiro MC, Velasco-Velázquez M, Aguirre-Alvarado C, Pestell RG (2014). Overview of cyclins D1 function in cancer and the CDK inhibitor landscape: past and present. Expert Opin Investig Drugs, 23, 295–304. [DOI] [PubMed] [Google Scholar]
- 71.Carvalho MR, Lima D, Reis RL, Oliveira JM, Correlo VM (2017). Anti-Cancer Drug Validation: the Contribution of Tissue Engineered Models. Stem Cell Reviews and Reports, 13(3), 347–63. [DOI] [PubMed] [Google Scholar]
- 72.Bello JO, Nieva LO, Paredes AC, Gonzalez AM, Zavaleta LR, Lizano M (2015). Regulation of the Wnt/β-Catenin Signaling Pathway by Human Papillomavirus E6 and E7 Oncoproteins. Viruses, 7(8):4734–55. [DOI] [PMC free article] [PubMed] [Google Scholar]