

Abstract
Accurate chromosome segregation is essential for cell proliferation. The centromere is a specialized chromosomal locus, on which the kinetochore structure is formed. The centromere/kinetochore is required for the equal separation of sister chromatids to daughter cells. Here, we review recent findings on centromere-specific chromatin, including its constitutive protein components, its de novo formation and maintenance mechanisms, and our progress in analyses with synthetic human artificial chromosomes (HACs).
Keywords: Centromere, Chromatin, Heterochromatin, Alphoid DNA, Human artificial chromosome (HAC)
Introduction
A number of specific proteins assemble on the centromere to form a specialized chromatin structure and maintain its functions. During the mitotic phase, kinetochore components assemble on the centromere chromatin. This centromere chromatin/kinetochore structure is essential for the equal segregation of sister chromatids to each daughter cell by generating accurate attachments to the mitotic spindle that produce mobile forces and determining the separation timing by monitoring the quality of interactions with microtubules. Dysfunctions in centromere chromatin lead to chromosomal instability. Lagging and unsegregated chromosomes form micronuclei, where DNA fragmentation may occur. Re-incorporation of damaged DNA from micronuclei into the nucleus leads to chromosomal DNA rearrangements during subsequent cell cycles [1–3]. Chromosomal instability is considered to be one of the major causes of genomic instability and cancer progression. Therefore, mechanisms for assembling, maintaining, and regulating centromere chromatin are crucially important in assuring proper chromosome segregation and avoiding chromosomal/genomic instability.
Constitutive components and mechanism of maintaining centromere chromatin
In all normal human chromosomes, centromere proteins assemble on a typical centromere specific sequence of tandem 171-bp repeats, termed alpha-satellite (alphoid) DNA, where they form centromere-specific chromatin (Figure 1a). The first centromere proteins discovered were CENP-A, CENP-B, and CENP-C [4,5].
Figure 1:

Proteins involved in centromere/kinetochore organization and maintenance.
(a) Centromere DNA and the centromere/kinetochore protein and interaction network. (Left panel) Box shows the key for the component symbols; (middle panel) centromere structure; (right panel) model of centromere/kinetochore interactions. CENP-A and CENP-B assemble on alphoid DNA with H3 nucleosomes, and CCAN proteins assemble on CENP-A nucleosomes. Kinetochore proteins assemble in the mitotic phase to regulate the equal segregation of sister chromosomes through interactions with microtubules, (b) CENP-A replenishing proteins and a model of balancing centromere chromatin and heterochromatin. The M18BP1 complex assembles on alphoid DNA through interactions with CENP-C and CENP-I, and recruits proteins, such as the HJURP-CENP-A-H4 complex and KAT7. RSF1 assembles on alphoid DNA through acetylation and promotes the histone exchange reaction [80]. In this histone exchange, old heterochromatinized nucleosomes are removed, and heterochromatin invasion into the centromere chromatin is prevented. This opens a space for the deposition of a new CENP-A. (c) Model of SUV39H1 overexpression in KAT7-knockout cells. When the mechanism for preventing heterochromatin invasion is lost, due to the lack of KAT7, SUV39H1 overexpression causes CENP-A reduction and the loss of centromere function. (Bottom right) Box shows the key for the component symbols.
CENP-A, a histone H3 variant, is a key epigenetic centromere marker. It forms a centromere-specific nucleosome with the canonical histones, H2A, H2B, and H4. A network of constitutive centromere-associated network proteins (termed the CCAN, ICEN, or CENP-A NAC/CAD proteins) is associated with the CENP-A nucleosome [6–12] (Figure 1a). These CCAN proteins are localized on centromere chromatin throughout the cell cycle. Among the CCAN proteins, CENP-C and CENP-N bind directly to CENP-A nucleosomes and stabilize their three-dimensional structure [6–12] (Figure 1a). In addition, compared to the canonical histone H3 nucleosome, the CENP-A nucleosome has flexibility in DNA binding, in the vicinity of the linker region [13,14]. When amino acid sequences in the cN region of the CENP-A protein, which is involved in the flexibility describing above, were replaced with the corresponding sequences in canonical histone H3, histone H1 became bound and CENP-C assembly with the centromere was lost [15].
CENP-B is the only protein that directly binds to a specific sequence in alphoid DNA - the 17 bp DNA motif known as the CENP-B box. The CENP-B box motif appears most commonly once every 340 bp in alphoid DNA dimers in centromeric alpha satellite higher order repeat domains [5,16]. The CENP-B protein has both a DNA binding domain and a CENP-A interaction domain at the N terminus [17,18]. CENP-B forms homodimers at the C-terminus and interacts with CENP-C via an acidic region near the C-terminus [19,20]. CENP-B is required for efficient de novo CENP-A chromatin assembly, when naked alphoid DNA is introduced into cells [21,22,], but it is dispensable in the Y chromosome centromere and in neocentromeres [23], and mice can survive without it [24–26]. CENP-B also enhances heterochromatin formation at ectopic integration sites of alphoid DNA in chromosomal arms that do not assemble other centromere components [22]. Consequently, it is extremely interesting how this functional centromere chromatin switch is regulated.
CENP-C is the key platform protein necessary both for CENP-A chromatin assembly and kinetochore formation. CENP-C interacts with CENP-A, CENP-B, CCAN proteins (CENP-I, CENP-L), a kinetochore protein, MIS12, and a CENP-A replenishment factor, M18BP1 [27–32]. Two CENP-C assembly pathways on alphoid DNA have been identified [33]. In one, CENP-C directly binds to the CENP-A C-terminus region in CENP-A nucleosomes. In the other pathway, CENP-C is assembled through an interaction with CENP-B. When only one of these pathways is blocked, centromere function is preserved, but when both pathways are blocked, centromere function is lost [33].
Currently, more than one hundred centromere/kinetochore protein components have been revealed. Many are highly conserved in various species, including human and yeast [34,35].
In addition to the kinetochore proteins, CENP-A replenishing factors also assemble on the centromere chromatin. Although deposition timing of canonical histone H3.1 is coupled with DNA replication, other H3 variants (i.e. H3.3 and CENP-A) are independent of DNA replication. In human cells, newly synthesized CENP-A deposition (replenishment) occurs in G1 phase [36]. The MIS18 complex (M18BP1, MIS18q and MIS180, which is involved in CENP-A replenishment, assembles at the centromere through an interaction with CENP-C from telophase to G1 phase (Figure 1b). The CENP-A chaperone, HJURP, is recruited through an interaction with MIS18aand MIS180[37–41]. M18BP1, MIS18q and MIS180form a complex with a 2:4:2 stoichiometry, respectively. The timing of MIS18 complex assembly at the centromere is regulated through modulation by protein kinases CDK and PLK1 [42–47]. In addition, DNA binding ability of HJURP is regulated by phosphorylation; this DNA binding activity and homodimer formation were reported to be important for CENP-A deposition [41,48,49].
Each new CENP-A molecule that is recruited to the centromere by HJURP is then stabilized as a newly assembled CENP-A nucleosome by chromatin remodeling factors, such as RSF1 [50]. RNA also plays an important role in centromere assembly that is still being actively studied. In human and fly cells, a cis-acting centromere transcript was also shown to stabilize CENP-A nucleosomes [51,52]. On the other hand, a trans-acting centromere RNA, ectopically transcribed from an introduced DNA vector, was reported to destabilize chromosome segregation [53]. CENP-C was reported to bind to RNA [54,55]. The Drosophila melanogaster HJURP homolog, CAL1, interacts with a chromatin remodeling factor complex, FACT, which promotes transcription [56]. In human cells, FACT was reported to interact with the CENP-T/W complex [57]. The newly assembled CENP-A nucleosomes might be stabilized by chromatin remodeling factors, CCAN protein assembly, and/or possibly by centromere DNA transcripts.
Alphoid DNA is competent in both de novo centromere assembly and HAC formation
Two types of alphoid DNA arrays have been identified at many human centromeres. The first is based on the higher-order repeat (HOR) units of alphoid DNA. The HOR unit is repeated homogeneously in tandem, to sizes on the order of a mega-base, with more than 97% identity (though the constituent monomers are much more variable). The second type of array is composed of divergent irregular 171-bp monomers. By comparing these two types of alphoid repeats in X chromosome centromeres, evolutionarily ancient transposon sequences were frequently found in the monomeric array, but not in the HOR. Thus, the HOR is considered to be evolutionarily newer than the monomeric alphoid arrays [58]. In addition, all the HORs of human chromosomes contain a CENP-B box, except the alphoid HOR on the Y chromosome.
When a HOR with several ten-kb sized components was introduced into human HT1080 cells, de novo centromere assembly occurred, and a human artificial chromosome (HAC) was formed via amplification of the introduced DNA (Figure 2a) [59–60]. Strikingly, no de novo centromere assembly or HAC formation occurred with alphoid DNA that lacked a CENP-B box (i.e., the Y alphoid HOR, the monomeric alphoid DNA of chromosome 21, and a synthetic alphoid DNA with a mutation in all CENP-B boxes). Thus, CENP-B protein and the CENP-B box are indispensable for efficient de novo centromere assembly and HAC formation [21,22,59–62]. It appears that the relevant function of CENP-B is to assemble CENP-A de novo on input alphoid DNAs. Newly assembled CENP-A is structurally stabilized by an interaction with the CENP-B N-terminal domain (Figure 2b) [17]. The assembly of CCAN proteins and/or alphoid RNA might also contribute to the stabilization of the newly assembled CENP-A nucleosome.
Figure 2:

A model of de novo centromere formation.
(a) Model of human artificial chromosome (HAC) formation. (Left) When an alphoid DNA molecule is introduced into cells, it multimerizes and the formation of a de novo centromere chromatin and heterochromatin occurs on the alphoid DNA. (Right) Fluorescence image shows stable centromeres, and the introduced alphoid DNA is maintained as a HAC. (b) Model of CENP-B function in de novo chromatin formation. CENP-B has multiple functions: it facilitates de novo CENP-A assembly and stabilizes the CENP-A nucleosome. CENP-B also promotes heterochromatinization, when alphoid DNA does not form a centromere, but integrates into a host chromosomal arm. Boxes show the keys for the component symbols.
Compared to the evolutionarily conserved centromere proteins, centromere DNAs evolved rapidly and diverged among species. Some strains of mice have chromosomes with expanded centromere repeats (minor satellite DNA) that contain CENP-B boxes, but no other sequences in common with primate alpha satellite DNA. These arrays produce stronger centromeres that assemble more CENP-A than observed in other mouse strains. Chromosomes with stronger centromeres were reported to be more likely to distribute to the egg-proximal side during female meiosis by the inter-strain crosses, and thus, they have a better chance of succeeding in the egg (that is, they will pass to the next generation, rather than being eliminated in the polar body) [63]. Acquisition of a CENP-B box on centromeric satellite DNA could confer an advantage to chromosome survival in female meiosis, as described in the centromere-drive/meiotic-drive theory [64]. This hypothesis could possibly explain why only Y centromere DNAs have no CENP-B box, both in human and mouse genomes, because the Y chromosome is not subject to female meiosis. In recent years, centromere DNAs with and without CENP-B boxes have been found in new-world monkey species [65,66]. It is an interesting puzzle how the CENP-B box was acquired through evolution.
It should be noted that, although de novo centromere assembly could occurr when a CENP-B box was combined with alphoid DNA, no centromere assembly occurred when a GC-rich repeat derived from a pBR plasmid fragment was combined with a CENP-B box [21]. Thus, in addition to the CENP-B box, alphoid DNA might have other advantageous properties for de novo centromere assembly. These could include AT-rich sequences and/or the ability to form non-B-form DNA upon CENP-B binding [67]).
The discovery that HACs could be generated from synthetic alphoid DNA has advanced the functional analysis of centromeres. Moreover, HACs have been applied as chromosomal vectors [68]. For example, synthetic HACs are useful for manipulating the epigenetic landscape of chromatin. These HACs contain synthetic alphoid DNA monomers with a tetracycline operator (tetO) sequence alternating with monomers bearing CENP-B boxes (tetO-alphoid DNA). The synthetic arrays can be targeted in vivo with tetracycline repressor (tetR) proteins fused to any protein or protein domain of interest. Binding to the tetO site thus tethers the chimeric protein to the tetO-alphoid DNA within the centromere. This approach enables removal or addition of histone modifications as well as the recruitment of other proteins that interact with the chimeric protein (Figure 3) [69–71]. The functional consequences are readily assayed by scoring for HAC loss and levels of critical proteins such as CENP-A and CENP-C. HACs are also useful as markers of chromosomal instability and as vectors for gene therapy [72,73].
Figure 3:
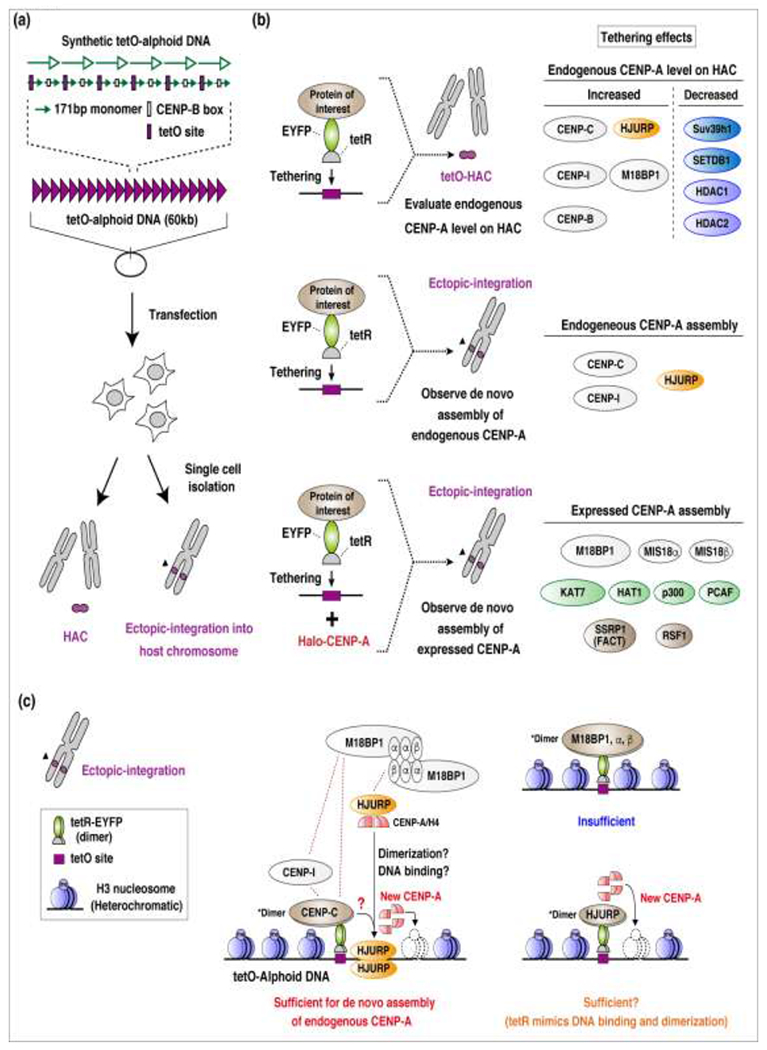
Schematic diagrams show evaluations of CENP-A assembly activity in the tethering assay.
(a, top) Structure of a synthetic tetO-alphoid DNA dimer-repeat unit. One monomer of the dimer has a CENP-B box; the other monomer has the tetO operator sequence, which binds the tet repressor protein (tetR). Dimers are ligated in tandem to form a 60 kb plasmid, and (middle) introduced into cells. The cells generate either (bottom) a tetO-HAC or a tetO site ectopically integrated into the host chromosome. (b) A schematic diagram shows three assays where the protein of interest is fused to the tetR sequence; upon tetR binding, the fused protein is tethered to the tetO-alphoid site (top) in the HAC or (middle and bottom) to the ectopic integration site in the chromosome. Assays were conducted without (middle) or with (bottom) added exogenous CENP-A (Halo-CENP-A). (Right) Summaries of the results (from Shono et al. [70]) are shown for each assay, (c) Model of de novo assembly of endogenous CENP-Ato the tetO-alphoid DNA integration site. (Left) The tetO-alphoid DNA integration site is highly heterochromatinized, and no CENP-A assembly is detected. (Middle) CENP-C tethering is sufficient forde novo assembly of endogenous CENP-A on the integration site. (Right, top) However, tethering the downstream factors, M18BP1, MIS18a, or MIS188is not sufficient for de novo CENP-A assembly. These results suggested that CENP-C might control either the activity of the MIS18 complex or HJURP binding to induce de novo endogenous CENP-A assembly on the ectopic integration site, without the centromere. (Right, bottom) Since the tetR binds to the tetO site as homodimers, tetR-fused HJURP protein binds to the tetO-alphoid DNA integration site as dimer. This tetR-fused HJURP dimer binding might be sufficient to drive de novo CENP-A assembly. (Bottom left) Box shows the key for the component symbols.
Chromatin assembly balance determines centromere assembly on alphoid DNA
In addition to CENP-A chromatin, heterochromatin is also assembled on alphoid DNA, when it carries histone H3 methylated on the K9 residue (H3K9me3). The H3K9me3 modification serves as a binding site for HP1 assembly. HP1 is reported to recruit cohesin and Aurora B in the chromosomal passenger complex (CPC). Cohesin maintains sister chromosome cohesion and the CPC improves the accuracy of chromosome segregation [74,75]. Heterochromatin can also spread to adjacent chromatin regions [76,77]. When heterochromatin invades the centromere, CENP-A chromatin and kinetochore functions are no longer maintained, and eventually, they are inactivated [69,78]. Interestingly, levels of H3K9me3 modification on alphoid DNA differ, depending on cell type; for example, the activity is high in HeLa cells and low in HT1080 cells. In HeLa cells, high H3K9me3 modification activity on alphoid DNA inhibits de novo centromere assembly on introduced alphoid DNA. Conversely, in HT1080 cells, low H3K9me3 modification activity permits de novo centromere assembly on introduced alphoid DNA [79]. Therefore, forde novo assembly of CENP-A chromatin, the epigenetic chromatin modification status against heterochromatinization is important. Established centromere chromatin might possess some type of barrier mechanism to protect against heterochromatin invasion.
Indeed, heterochromatinization was blocked by acetylation of tetO-alphoid chromatin by a tethered HAT (histone acetyl transferase) in the synthetic HAC system [79]. Recent studies have identified endogenous acetylating enzymes involved in the CENP-A chromatin assembly process. KAT7 is an acetylating enzyme active in the G1 phase; it is recruited onto alphoid DNA through an interaction with M18BP1 [80]. Centromeres in KAT7-knockout cells were sensitive to overexpression of the H3K9 tri-methylating enzyme, SUV39H1, which forms pericentromere heterochromatin. Specifically, the overexpression of SUV39H1 in KAT7-knockout cells resulted in high frequencies of chromosome loss and micronucleus formation (Figure 1c). Histone H3 acetylation, induced by the tethering of KAT7 onto tetO-alphoid DNA, recruits a remodeling factor, RSF1, and enhances histone turnover. This reduces pre-existing H3K9me3 nucleosomes and promotes assembly of ectopically expressed histone H3 variants (i.e., H3.3 and CENP-A). Thus, KAT7 can prevent heterochromatin from spreading into the centromere by promoting a histone exchange reaction through RSF1 (Figure 1b). This histone exchange reaction may also be involved in the CENP-A deposition pathway. KAT7 also induces H3K14 acetylation (H3K14ac) [81,82]. H3K14ac was sufficient for evicting histone through a histone chaperone, NAP1, in vitro [83]. Thus, KAT7 induction of H3K14ac might also be involved in the histone exchange process on alphoid DNA. HAT1 is another acetyltransferase involved in CENP-A assembly. Chicken HAT1 induced the acetylation of histones H4K5 and H4K12. This recruited a prenucleosomal complex composed of HJURP, CENP-A, and histone H4 to the centromere [84]. Drosophila melanogaster HAT1 is also involved in the new CENP-A assembly pathway [85].
It is becoming clear that transcriptionally active or neutral modifications, such as H3K9ac, H3K4me2, H3K36me2, and H4K20me1 promote centromere assembly [80,86–90]. In addition, the modification of CENP-A through aamino trimethylation is also involved in centromere assembly [91,92]. Transcriptional activity also affects centromere maintenance. Strong transcription causes loss of centromere activity, but moderate levels of transcription are compatible with centromere activity [93]. Based on the evidence that cis-transcripts stabilize the CENP-A nucleosome, it is reasonable to postulate that an unknown mechanism might exist for initiating transcription within centromere DNA. Further studies are necessary to elucidate a mechanism that can explain how histone modifications, chromatin proteins, and transcription events are involved in centromere assembly and maintenance.
Chromatin factors involved in CENP-A assembly and maintenance
Two HeLa cell lines were generated to evaluate how chromatin factors and histone modifiers affected CENP-A assembly on alphoid DNA. One cell line contained the HAC, and the other contained tetO-alphoid DNA integrated into a host chromosome (Figure 3a) [70]. In the first assay (Figure 3b), the tetR was fused with several chromatin proteins of interest. Upon tetR binding to the tetO site on the HAC (tetO-HAC) [66], each protein of interest was tethered to that site. Then, the effect of each protein on levels of endogenous CENP-A at the tetO-HAC centromere was quantified. In other assays, tetR-fused proteins were tethered to the ectopic integration site of the tetO-alphoid DNA. Each construct was then assessed for whether it induced the assembly of either de novo endogenous CENP-A or exogenously over-expressed CENP-A (Figure 3b). The results revealed that CENP-C, CENP-I, and HJURP contributed to the assembly of endogenous CENP-A on the ectopic tetO-alphoid DNA site. In contrast, other MIS18 complex proteins (M18BP1, MIS18q MIS18$, which assemble to the centromere downstream of CENP-C, were not sufficient to assemble endogenous CENP-A on the ectopic tetO-alphoid integration site (Figure 3c). These results suggest that the presence of CENP-C and CENP-I might regulate the activity of MIS18 or the HJURP complex in de novo endogenous CENP-A deposition, even on a non-centromeric tetO-alphoid site. Alternatively, HJURP dimerization and binding through the tetR might mimic or bypass the regulation by CENP-C and CENP-I. Taken together, these findings indicate that CENP-C and CENP-I might promote HJURP dimerization and binding to DNA. However, it remains unclear whether CENP-C and CENP-I directly regulate this process, or whether some other proteins (or modifications) mediate the process.
In these experiments, most tethered proteins that promote open chromatin, such as acetyltransferase subunits or the RSF1 remodeling factor, could promote the assembly of exogenously over-expressed CENP-A or a placeholder histone H3.3 on the ectopic tetO-alphoid DNA integration site (Figure 3b). A potential mechanism could be that these chromatin modifiers and the remodeling factor promote histone exchange or turnover on the tetO-alphoid DNA site. This would create openings for the deposition of a new histone H3.3 or CENP-A. Interestingly, some of these factors (M18BP1, SSRP1, HAT1) selectively assembled CENP-A rather than histone H3.1 or H3.3 at the ectopic tetO-alphoid DNA integration site [70]. Indeed, in chicken and Drosophila melanogaster cells, FACT (SSRP, SPT16) and HAT1 are normally involved in CENP-A assembly [56,84,85]. These findings suggested that open chromatin modifications and/or remodeling mechanisms are very likely involved in the CENP-A deposition process.
In the same experiments, tethered heterochromatin factors caused reductions in CENP-A levels on HAC centromeres (Figure 3b) [70]. When HP1 was tethered to the tetO-HAC, a hierarchical centromere protein reduction occurred on the HAC centromere, in the following order: CENP-H, CENP-C, then CENP-A [94]. In addition, at chicken centromeres, CENP-T and CENP-H levels fell when H4K20me1 was removed [87]. It is interesting that heterochromatin spreading destabilized CCAN protein assembly, which consequently, destabilized CENP-A.
Closing remarks
By now, the list of structural components that assemble on centromere chromatin is largely complete. In recent years, the functions of these core components have been studied by a wide range of methods, including in vitro reconstitution, crystal structures, and other analyses. Parallel studies of natural and synthetic human artificial chromosomes (HACs) have focused on characterizing the chromatin environment that is essential for kinetochore assembly. In particular, the tethering of various chromatin modifiers to the HAC and ectopic integration sites has been applied to investigate de novo centromere assembly and maintenance in vivo. This approach has led to new insights that have suggested that centromere chromatin assembly and maintenance are controlled positively and negatively on repetitive DNA through chromatin modifications and remodeling activities. Thus, the tethering approach is a powerful method for evaluating how chromatin modifications and remodeling can affect centromere organization and establishment. Together these studies are beginning to reveal how the centromere is organized dynamically on repetitive DNA.
Acknowledgements
This work was supported by MEXT KAKENHI [grant numbers 16H04747, 16H01414, and 18H04721] and the Kazusa DNA Research Institute Foundation (to H. Masumoto). Work in the W.C. Earnshaw laboratory was funded by the Wellcome Trust, of which W.C. Earnshaw is a Principal Research Fellow [grant number 073915]. Work in the V. Larionov laboratory was supported by an Intramural Research Program of the National Institutes of Health, the National Cancer Institute, and the Center for Cancer Research.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of interest
The authors declare no conflict of interest.
Contributor Information
Junichirou Ohzeki, Email: johzeki@kazusa.or.jp.
Vladimir Larionov, Email: larionov@mail.nih.gov.
William C. Earnshaw, Email: Bill.Earnshaw@ed.ac.uk.
Hiroshi Masumoto, Email: masumoto@kazusa.or.jp.
References
- 1.Zhang W, Mao JH, Zhu W, Jain AK, Liu K, Brown JB, Karpen GH: Centromere and kinetochore gene misexpression predicts cancer patient survival and response to radiotherapy and chemotherapy. Nat Commun 2016, 7:12619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ly P, Teitz LS, Kim DH, Shoshani O, Skaletsky H, Fachinetti D, Page DC, Cleveland DW: Selective Y centromere inactivation triggers chromosome shattering in micronuclei and repair by non-homologous end joining. Nat Cell Biol 2017, 19:68–75. [DOI] [PMC free article] [PubMed] [Google Scholar]; **elegantly demonstrated that centromere dysfunction reads micronuclei formation, chromosomal DNA fragmentation and reassembly.
- 3.Crasta K, Ganem NJ, Dagher R, Lantermann AB, Ivanova EV, Pan Y, Nezi L, Protopopov A, Chowdhury D, Pellman D: DNA breaks and chromosome pulverization from errors in mitosis. Nature 2012, 482:53–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Willard HF, Waye JS: Hierarchical order in chromosome-specific human alpha satellite DNA. Trends Genet 1987, 3:192–198. [Google Scholar]
- 5.Earnshaw WC, Rothfield N: Identification of a family of human centromere proteins using autoimmune sera from patients with scleroderma. Chromosoma 1895, 91:313–321. [DOI] [PubMed] [Google Scholar]
- 6.Cao S, Zhou K, Zhang Z, Luger K, Straight AF: Constitutive centromere-associated network contacts confer differential stability on CENP-A nucleosomes in vitro and in the cell. Mol Biol Cell 2018, 29:751–762. [DOI] [PMC free article] [PubMed] [Google Scholar]; **showed that CENP-C and CENP-N stabilized CENP-A nucleosome in vitro, but are not essential in the cell. There might be another additional CENP-A nucleosome stabilization mechanisms in vivo.
- 7.Pentakota S, Zhou K, Smith C, Maffini S, Petrovic A, Morgan GP, Weir JR, Vetter IR, Musacchio A, Luger K: Decoding the centromeric nucleosome through CENP-N. Elife 2017, 6. [DOI] [PMC free article] [PubMed] [Google Scholar]; **demonstrated how CENP-C and CENP-N recognize CENP-A nucleosome.
- 8.Chittori S, Hong J, Saunders H, Feng H, Ghirlando R, Kelly AE, Bai Y, Subramaniam S: Structural mechanisms of centromeric nucleosome recognition by the kinetochore protein CENP-N. Science 2018, 359:339–343. [DOI] [PMC free article] [PubMed] [Google Scholar]; **demonstrated that CENP-N recognize CENP-A through the L1 loop region of CENP-A.
- 9.Guo LY, Allu PK, Zandarashvili L, McKinley KL, Sekulic N, Dawicki-McKenna JM, Fachinetti D, Logsdon GA, Jamiolkowski RM, Cleveland DW, Cheeseman IM, Black BE: Centromeres are maintained by fastening CENP-A to DNA and directing an arginine anchor-dependent nucleosome transition. Nat Commun 2017, 8:15775. [DOI] [PMC free article] [PubMed] [Google Scholar]; ** demonstrated that CENP-C and CENP-N stabilize CENP-A nucleosome.
- 10.Falk SJ, Lee J, Sekulic N, Sennett MA, Lee TH, Black BE: CENP-C directs a structural transition of CENP-A nucleosomes mainly through sliding of DNA gyres. Nat Struct Mol Biol 2016, 23:204–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Weir JR, Faesen AC, Klare K, Petrovic A, Basilico F, Fischböck J, Pentakota S, Keller J, Pesenti ME, Pan D, Vogt D, Wohlgemuth S, Herzog F, Musacchio A: Insights from biochemical reconstitution into the architecture of human kinetochores. Nature 2016, 537:249–253. [DOI] [PubMed] [Google Scholar]
- 12.Pesenti ME, Prumbaum D, Auckland P, Smith CM, Faesen AC, Petrovic A, Erent M, Maffini S, Pentakota S, Weir JR, Lin YC, Raunser S, McAinsh AD, Musacchio A: Reconstitution of a 26-Subunit Human Kinetochore Reveals Cooperative Microtubule Binding by CENP-OPQUR and NDC80. Mol Cell 2018, 71:923–939.e10. [DOI] [PMC free article] [PubMed] [Google Scholar]; **reconstructed centromere/kinetochore structure using 26 of main protein components and revealed a novel function of CENP-OPQUR.
- 13.Tachiwana H, Kagawa W, Shiga T, Osakabe A, Miya Y, Saito K, Hayashi-Takanaka Y, Oda T, Sato M, Park SY, Kimura H, Kurumizaka H: Crystal structure of the human centromeric nucleosome containing CENP-A. Nature 2011, 476:232–235. [DOI] [PubMed] [Google Scholar]
- 14.Stumme-Diers MP, Banerjee S, Hashemi M, Sun Z, Lyubchenko YL: Nanoscale dynamics of centromere nucleosomes and the critical roles of CENP-A. Nucleic Acids Res 2018, 46:94–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Roulland Y, Ouararhni K, Naidenov M, Ramos L, Shuaib M, Syed SH, Lone IN, Boopathi R, Fontaine E, Papai G, Tachiwana H, Gautier T, Skoufias D, Padmanabhan K, Bednar J, Kurumizaka H, Schultz P, Angelov D, Hamiche A, Dimitrov S: The Flexible Ends of CENP-A Nucleosome Are Required for Mitotic Fidelity. Mol Cell 2016, 63:674–685. [DOI] [PubMed] [Google Scholar]
- 16.Masumoto H, Masukata H, Muro Y, Nozaki N, Okazaki T. A human centromere antigen (CENP-B) interacts with a short specific sequence in alphoid DNA, a human centromeric satellite. J Cell Biol 1989, 109:1963–1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fujita R, Otake K, Arimura Y, Horikoshi N, Miya Y, Shiga T, Osakabe A, Tachiwana H, Ohzeki J, Larionov V, Masumoto H, Kurumizaka H: Stable complex formation of CENP-B with the CENP-A nucleosome. Nucleic Acids Res 2015, 43:4909–4922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fachinetti D, Han JS, McMahon MA, Ly P, Abdullah A, Wong AJ, Cleveland DW: DNA Sequence-Specific Binding of CENP-B Enhances the Fidelity of Human Centromere Function. Dev Cell 2015, 33:314–327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kitagawa K, Masumoto H, Ikeda M, Okazaki T: Analysis of protein-DNA and protein-protein interactions of centromere protein B (CENP-B) and properties of the DNA-CENP-B complex in the cell cycle. Mol Cell Biol 1995, 15:1602–1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Suzuki N, Nakano M, Nozaki N, Egashira S, Okazaki T, Masumoto H: CENP-B interacts with CENP-C domains containing Mif2 regions responsible for centromere localization. J Biol Chem 2004, 279:5934–5946. [DOI] [PubMed] [Google Scholar]
- 21.Ohzeki J, Nakano M, Okada T, Masumoto H. CENP-B box is required for de novo centromere chromatin assembly on human alphoid DNA. J Cell Biol 2002, 159:765–775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Okada T, Ohzeki J, Nakano M, Yoda K, Brinkley WR, Larionov V, Masumoto H: CENP-B controls centromere formation depending on the chromatin context. Cell 2007, 131:1287–1300. [DOI] [PubMed] [Google Scholar]
- 23.du Sart D, Cancilla MR, Earle E, Mao JI, Saffery R, Tainton KM, Kalitsis P, Martyn J, Barry AE, Choo KH: A functional neo-centromere formed through activation of a latent human centromere and consisting of non-alpha-satellite DNA. Nat Genet. 1997, 16:144–153. [DOI] [PubMed] [Google Scholar]
- 24.Hudson DF, Fowler KJ, Earle E, Saffery R, Kalitsis P, Trowell H, Hill J, Wreford NG, de Kretser DM, Cancilla MR, Howman E, Hii L, Cutts SM, Irvine DV, Choo KH: Centromere protein B null mice are mitotically and meiotically normal but have lower body and testis weights. J Cell Biol. 1998, 141: 309–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Perez-Castro AV, Shamanski FL, Meneses JJ, Lovato TL, Vogel KG, Moyzis RK, Pedersen R: Centromeric protein B null mice are viable with no apparent abnormalities. Dev Biol. 1998, 201:135–143. [DOI] [PubMed] [Google Scholar]
- 26.Kapoor M, de Montes de Oca Luna R, Liu G, Lozano G, Cummings C, Mancini M, Ouspenski I, Brinkley BR, May GS: The CENPB gene is not essential in mice. Chromosoma 1998, 107:570–576. [DOI] [PubMed] [Google Scholar]
- 27.Petrovic A, Keller J, Liu Y, Overlack K, John J, Dimitrova YN, Jenni S, van Gerwen S, Stege P, Wohlgemuth S, Rombaut P, Herzog F, Harrison SC, Vetter IR, Musacchio A: Structure of the MIS12 Complex and Molecular Basis of Its Interaction with CENP-C at Human Kinetochores. Cell 2016, 167:1028–1040.e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Przewloka MR, Venkei Z, Bolanos-Garcia VM, Debski J, Dadlez M, Glover DM: CENP-C is a structural platform for kinetochore assembly. Curr Biol 2011, 21:399–405. [DOI] [PubMed] [Google Scholar]
- 29.Screpanti E, De Antoni A, Alushin GM, Petrovic A, Melis T, Nogales E, Musacchio A: Direct binding of Cenp-C to the Mis12 complex joins the inner and outer kinetochore. Curr Biol 2011, 21:391–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Milks KJ, Moree B, Straight AF: Dissection of CENP-C-directed centromere and kinetochore assembly. Mol Biol Cell 2009, 20:4246–4255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dambacher S, Deng W, Hahn M, Sadic D, Fröhlich J, Nuber A, Hoischen C, Diekmann S, Leonhardt H, Schotta G: CENP-C facilitates the recruitment of M18BP1 to centromeric chromatin. Nucleus 2012, 3:101–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Moree B, Meyer CB, Fuller CJ, Straight AF: CENP-C recruits M18BP1 to centromeres to promote CENP-A chromatin assembly. J Cell Biol 2011, 194:855–871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fachinetti D, Folco HD, Nechemia-Arbely Y, Valente LP, Nguyen K, Wong AJ, Zhu Q, Holland AJ, Desai A, Jansen LE, Cleveland DW: A two-step mechanism for epigenetic specification of centromere identity and function. Nat Cell Biol 2013, 15:1056–1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Musacchio A, Desai A: A Molecular View of Kinetochore Assembly and Function. Biology (Basel) 2017, 6 pii: E5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.van Hooff JJ, Tromer E, van Wijk LM, Snel B, Kops GJ: Evolutionary dynamics of the kinetochore network in eukaryotes as revealed by comparative genomics. EMBO Rep 2017, 18:1559–1571. [DOI] [PMC free article] [PubMed] [Google Scholar]; *demonstrated both conservation and divergence of class of centromere proteins among eukaryotes. It is curious how species lacking some class of centromere proteins maintain its centromere structure and function.
- 36.Jansen LE, Black BE, Foltz DR, Cleveland DW: Propagation of centromeric chromatin requires exit from mitosis. J Cell Biol 2007, 176:795–805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fujita Y, Hayashi T, Kiyomitsu T, Toyoda Y, Kokubu A, Obuse C, Yanagida M: Priming of centromere for CENP-A recruitment by human hMis18alpha, hMis18beta, and M18BP1. Dev Cell 2007, 12:17–30. [DOI] [PubMed] [Google Scholar]
- 38.Maddox PS, Hyndman F, Monen J, Oegema K, Desai A: Functional genomics identifies a Myb domain-containing protein family required for assembly of CENP-A chromatin. J Cell Biol 2007, 176:757–763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Foltz DR, Jansen LE, Bailey AO, Yates JR 3rd, Bassett EA, Wood S, Black BE, Cleveland DW: Centromere-specific assembly of CENP-a nucleosomes is mediated by HJURP. Cell 2009, 137:472–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dunleavy EM, Roche D, Tagami H, Lacoste N, Ray-Gallet D, Nakamura Y, Daigo Y, Nakatani Y, Almouzni-Pettinotti G: HJURP is a cell-cycle-dependent maintenance and deposition factor of CENP-A at centromeres. Cell 2009, 137:485–497. [DOI] [PubMed] [Google Scholar]
- 41.Stankovic A, Guo LY, Mata JF, Bodor DL, Cao XJ, Bailey AO, Shabanowitz J, Hunt DF, Garcia BA, Black BE, Jansen LET: A Dual Inhibitory Mechanism Sufficient to Maintain Cell-Cycle-Restricted CENP-A Assembly. Mol Cell 2017, 65:231–246. [DOI] [PMC free article] [PubMed] [Google Scholar]; **demonstrated how G1 phase specific CENP-A deposition through M18BP1 and HJURP is regulated.
- 42.Spiller F, Medina-Pritchard B, Abad MA, Wear MA, Molina O, Earnshaw WC, Jeyaprakash AA: Molecular basis for Cdk1-regulated timing of Mis18 complex assembly and CENP-A deposition. EMBO Rep 2017, 18:894–905. [DOI] [PMC free article] [PubMed] [Google Scholar]; **See note Ref. [43]
- 43.Pan D, Klare K, Petrovic A, Take A, Walstein K, Singh P, Rondelet A, Bird AW, Musacchio A: CDK-regulated dimerization of M18BP1 on a Mis18 hexamer is necessary for CENP-A loading. Elife 2017, 6 pii: e23352. [DOI] [PMC free article] [PubMed] [Google Scholar]; **Refs. 42 and 43 provide composition of human M18BP1 complex contains hetero hexamer of MIS18α and MIS18β.
- 44.Stellfox ME, Nardi IK, Knippler CM, Foltz DR: Differential Binding Partners of the Mis18α/βYIPPEE Domains Regulate Mis18 Complex Recruitment to Centromeres. Cell Rep 2016, 15:2127–2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Subramanian L, Medina-Pritchard B, Barton R, Spiller F, Kulasegaran-Shylini R, Radaviciute G, Allshire RC, Arockia Jeyaprakash A: Centromere localization and function of Mis18 requires Yippee-like domain-mediated oligomerization. EMBO Rep 2016, 17:496–507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.McKinley KL, Cheeseman IM: Polo-like kinase 1 licenses CENP-A deposition at centromeres. Cell 2014, 158:397–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Silva MC, Bodor DL, Stellfox ME, Martins NM, Hochegger H, Foltz DR, Jansen LE: Cdk activity couples epigenetic centromere inheritance to cell cycle progression. Dev Cell 2012, 22:52–63. [DOI] [PubMed] [Google Scholar]
- 48.Müller S, Montes de Oca R, Lacoste N, Dingli F, Loew D, Almouzni G: Phosphorylation and DNA binding of HJURP determine its centromeric recruitment and function in CenH3(CENP-A) loading. Cell Rep 2014, 8:190–203. [DOI] [PubMed] [Google Scholar]
- 49.Zasadzińska E, Barnhart-Dailey MC, Kuich PH, Foltz DR: Dimerization of the CENP-A assembly factor HJURP is required for centromeric nucleosome deposition. EMBO J 2013, 32:2113–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Perpelescu M, Nozaki N, Obuse C, Yang H, Yoda K: Active establishment of centromeric CENP-A chromatin by RSF complex. J Cell Biol 2009, 185:397–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.McNulty SM, Sullivan LL, Sullivan BA: Human Centromeres Produce Chromosome-Specific and Array-Specific Alpha Satellite Transcripts that Are Complexed with CENP-A and CENP-C. Dev Cell 2017, 42:226–240.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]; **See note Ref. [52]
- 52.Bobkov GOM, Gilbert N, Heun P: Centromere transcription allows CENP-A to transit from chromatin association to stable incorporation. J Cell Biol 2018, 217:1957–1972. [DOI] [PMC free article] [PubMed] [Google Scholar]; **Refs. 51 and 52 provide evidences that cis-acting centromere transcript is involved in a stable CENP-A assembly.
- 53.Chan DYL, Moralli D, Khoja S, Monaco ZL: Noncoding Centromeric RNA Expression Impairs Chromosome Stability in Human and Murine Stem Cells. Dis Markers 2017, 2017: [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Du Y, Topp CN, Dawe RK: DNA binding of centromere protein C (CENPC) is stabilized by single-stranded RNA. PLoS Genet 2010, 6:e1000835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wong LH, Brettingham-Moore KH, Chan L, Quach JM, Anderson MA, Northrop EL, Hannan R, Saffery R, Shaw ML, Williams E, Choo KH: Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res 2007, 17:1146–1160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chen CC, Bowers S, Lipinszki Z, Palladino J, Trusiak S, Bettini E, Rosin L, Przewloka MR, Glover DM, O’Neill RJ, Mellone BG: Establishment of Centromeric Chromatin by the CENP-A Assembly Factor CAL1 Requires FACT-Mediated Transcription. Dev Cell 2015, 34:73–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Prendergast L, Müller S, Liu Y, Huang H, Dingli F, Loew D, Vassias I, Patel DJ, Sullivan KF, Almouzni G: The CENP-T/-W complex is a binding partner of the histone chaperone FACT. Genes Dev 2016, 30:1313–1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Schueler MG, Higgins AW, Rudd MK, Gustashaw K, Willard HF: Genomic and genetic definition of a functional human centromere. Science 2001, 294:109–115. [DOI] [PubMed] [Google Scholar]
- 59.Harrington JJ, Van Bokkelen G, Mays RW, Gustashaw K, Willard HF: Formation of de novo centromeres and construction of first-generation human artificial microchromosomes. Nat Genet 1997, 15:345–355. [DOI] [PubMed] [Google Scholar]
- 60.Ikeno M, Grimes B, Okazaki T, Nakano M, Saitoh K, Hoshino H, McGill NI, Cooke H, Masumoto H: Construction of YAC-based mammalian artificial chromosomes. Nat Biotechnol 1998, 16:431–439. [DOI] [PubMed] [Google Scholar]
- 61.Kouprina N, Samoshkin A, Erliandri I, Nakano M, Lee HS, Fu H, Iida Y, Aladjem M, Oshimura M, Masumoto H, Earnshaw WC, Larionov V: Organization of synthetic alphoid DNA array in human artificial chromosome (HAC) with a conditional centromere. ACS Synth Biol. 2012, 1:590–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Basu J, Stromberg G, Compitello G, Willard HF, Van Bokkelen G: Rapid creation of BAC-based human artificial chromosome vectors by transposition with synthetic alpha-satellite arrays. Nucleic Acids Res 2005, 33:587–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Iwata-Otsubo A, Dawicki-McKenna JM, Akera T, Falk SJ, Chmátal L, Yang K, Sullivan BA, Schultz RM, Lampson MA, Black BE: Expanded Satellite Repeats Amplify a Discrete CENP-A Nucleosome Assembly Site on Chromosomes that Drive in Female Meiosis. Curr Biol 2017, 27:2365–2373.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]; **showed an experimental evidence how centromeric DNA has been selected for evolution.
- 64.Drinnenberg IA, Henikoff S, Malik HS: Evolutionary Turnover of Kinetochore Proteins: A Ship of Theseus? Trends Cell Biol. 2016. July;26(7):498–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kugou K, Hirai H, Masumoto H, Koga A: Formation of functional CENP-B boxes at diverse locations in repeat units of centromeric DNA in New World monkeys. Sci Rep 2016, 6:27833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Suntronpong A, Kugou K, Masumoto H, Srikulnath K, Ohshima K, Hirai H, Koga A: CENP-B box, a nucleotide motif involved in centromere formation, occurs in a New World monkey. Biol Lett 2016, 12: [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kasinathan S, Henikoff S: Non-B-Form DNA Is Enriched at Centromeres. Mol Biol Evol. 2018, 35:949–962. [DOI] [PMC free article] [PubMed] [Google Scholar]; **showed DNA sequence features favorable for centromere formation of non-B-form DNA which is enriched at centromeres, and suggests CENP-B contribution in the induced non-B form DNA formation.
- 68.Kouprina N, Earnshaw WC, Masumoto H, Larionov V: A new generation of human artificial chromosomes for functional genomics and gene therapy. Cell Mol Life Sci. 2013, 70:1135–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nakano M, Cardinale S, Noskov VN, Gassmann R, Vagnarelli P, Kandels-Lewis S, Larionov V, Earnshaw WC, Masumoto H: Inactivation of a human kinetochore by specific targeting of chromatin modifiers. Dev Cell 2008, 14:507–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shono N, Ohzeki J, Otake K, Martins NM, Nagase T, Kimura H, Larionov V, Earnshaw WC, Masumoto H: CENP-C and CENP-I are key connecting factors for kinetochore and CENP-A assembly. J Cell Sci 2015, 128:4572–4587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Molina O, Kouprina N, Masumoto H, Larionov V, Earnshaw WC: Using human artificial chromosomes to study centromere assembly and function. Chromosoma 2017, 126:559–575. [DOI] [PubMed] [Google Scholar]
- 72.Kouprina N, Petrov N, Molina O, Liskovykh M, Pesenti E, Ohzeki JI, Masumoto H, Earnshaw WC, Larionov V: Human Artificial Chromosome with Regulated Centromere: A Tool for Genome and Cancer Studies. ACS Synth Biol 2018, 7:1974–1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Oshimura M, Uno N, Kazuki Y, Katoh M, Inoue T: A pathway from chromosome transfer to engineering resulting in human and mouse artificial chromosomes for a variety of applications to bio-medical challenges. Chromosome Res 2015, 23:111–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Yi Q, Chen Q, Liang C, Yan H, Zhang Z, Xiang X, Zhang M, Qi F, Zhou L, Wang F. HP1 links centromeric heterochromatin to centromere cohesion in mammals. EMBO Rep 2018, 19 pii: e45484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Abe Y, Sako K, Takagaki K, Hirayama Y, Uchida KS, Herman JA, DeLuca JG, Hirota T: HP1-Assisted Aurora B Kinase Activity Prevents Chromosome Segregation Errors. Dev Cell 2016, 36:487–497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Allshire RC, Javerzat JP, Redhead NJ, Cranston G: Position effect variegation at fission yeast centromeres. Cell 1994, 76:157–169. [DOI] [PubMed] [Google Scholar]
- 77.Talbert PB, Henikoff S: Spreading of silent chromatin: inaction at a distance. Nat Rev Genet 2006, 7:793–803. [DOI] [PubMed] [Google Scholar]
- 78.Scott KC, Merrett SL, Willard HF: A heterochromatin barrier partitions the fission yeast centromere into discrete chromatin domains. Curr Biol 2006, 16:119–129. [DOI] [PubMed] [Google Scholar]
- 79.Ohzeki J, Bergmann JH, Kouprina N, Noskov VN, Nakano M, Kimura H, Earnshaw WC, Larionov V, Masumoto H: Breaking the HAC Barrier: histone H3K9 acetyl/methyl balance regulates CENP-A assembly. EMBO J 2012, 31:2391–2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ohzeki J, Shono N, Otake K, Martins NM, Kugou K, Kimura H, Nagase T, Larionov V, Earnshaw WC, Masumoto H: KAT7/HBO1/MYST2 Regulates CENP-A Chromatin Assembly by Antagonizing Suv39h1-Mediated Centromere Inactivation. Dev Cell 2016, 37:413–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kueh AJ, Dixon MP, Voss AK, Thomas T: HBO1 is required for H3K14 acetylation and normal transcriptional activity during embryonic development. Mol Cell Biol 2011, 31:845–860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Mishima Y, Miyagi S, Saraya A, Negishi M, Endoh M, Endo TA, Toyoda T, Shinga J, Katsumoto T, Chiba T, Yamaguchi N, Kitabayashi I, Koseki H, Iwama A: The Hbo1-Brd1/Brpf2 complex is responsible for global acetylation of H3K14 and required for fetal liver erythropoiesis. Blood 2011, 118:2443–2453. [DOI] [PubMed] [Google Scholar]
- 83.Luebben WR, Sharma N, Nyborg JK: Nucleosome eviction and activated transcription require p300 acetylation of histone H3 lysine 14. Proc Natl Acad Sci U S A 2010, 107:19254–19259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Shang WH, Hori T, Westhorpe FG, Godek KM, Toyoda A, Misu S, Monma N, Ikeo K, Carroll CW, Takami Y, Fujiyama A, Kimura H, Straight AF, Fukagawa T: Acetylation of histone H4 lysine 5 and 12 is required for CENP-A deposition into centromeres. Nat Commun 2016, 7:13465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Boltengagen M, Huang A, Boltengagen A, Trixl L, Lindner H, Kremser L, Offterdinger M, Lusser A: A novel role for the histone acetyltransferase Hat1 in the CENP-A/CID assembly pathway in Drosophila melanogaster. Nucleic Acids Res 2016, 44:2145–2159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zhu J, Cheng KCL, Yuen KWY: Histone H3K9 and H4 Acetylations and Transcription Facilitate the Initial CENP-AHCP-3 Deposition and De Novo Centromere Establishment in Caenorhabditis elegans Artificial Chromosomes. Epigenetics Chromatin 2018, 11:16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Hori T, Shang WH, Toyoda A, Misu S, Monma N, Ikeo K, Molina O, Vargiu G, Fujiyama A, Kimura H, Earnshaw WC, Fukagawa T: Histone H4 Lys 20 monomethylation of the CENP-A nucleosome is essential for kinetochore assembly. Dev Cell 2014, 29:740–749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Molina O, Vargiu G, Abad MA, Zhiteneva A, Jeyaprakash AA, Masumoto H, Kouprina N, Larionov V, Earnshaw WC: Epigenetic engineering reveals a balance between histone modifications and transcription in kinetochore maintenance. Nat Commun 2016, 7:13334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Bergmann JH, Rodríguez MG, Martins NM, Kimura H, Kelly DA, Masumoto H, Larionov V, Jansen LE, Earnshaw WC: Epigenetic engineering shows H3K4me2 is required for HJURP targeting and CENP-A assembly on a synthetic human kinetochore. EMBO J 2011, 30:328–3240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Pesenti E, Kouprina N, Liskovykh M, Aurich-Costa J, Larionov V, Masumoto H, Earnshaw WC, Molina O: Generation of a Synthetic Human Chromosome with Two Centromeric Domains for Advanced Epigenetic Engineering Studies. ACS Synth Biol 2018, 7:1116–1130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Srivastava S, Zasadzińska E, Foltz DR: Posttranslational mechanisms controlling centromere function and assembly. Curr Opin Cell Biol 2018, 52:126–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sathyan KM, Fachinetti D, Foltz DR: α-amino trimethylation of CENP-A by NRMT is required for full recruitment of the centromere. Nat Commun 2017, 8:14678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Bergmann JH, Jakubsche JN, Martins NM, Kagansky A, Nakano M, Kimura H, Kelly DA, Turner BM, Masumoto H, Larionov V, Earnshaw WC: Epigenetic engineering: histone H3K9 acetylation is compatible with kinetochore structure and function. J Cell Sci 2012, 125:411–421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Cardinale S, Bergmann JH, Kelly D, Nakano M, Valdivia MM, Kimura H, Masumoto H, Larionov V, Earnshaw WC: Hierarchical inactivation of a synthetic human kinetochore by a chromatin modifier. Mol Biol Cell 2009, 20:4194–4204. [DOI] [PMC free article] [PubMed] [Google Scholar]