

Abstract
HIV infection is associated with a rapid and sustained inversion of the Vδ1:Vδ2 T‐cell ratio in peripheral blood. Studies of antiretroviral therapy (ART)‐treated cohorts suggest that ART is insufficient to reconstitute either the frequency or function of the γδ T‐cell subset. Recent advances are now beginning to shed light on the relationship between microbial translocation, chronic inflammation, immune ageing and γδ T‐cell immunology. Here, we review the impact of acute, chronic untreated and treated HIV infection on circulating and mucosal γδ T‐cell subsets and highlight novel approaches to harness γδ T cells as components of anti‐HIV immunotherapy.
Keywords: γδ, gut, HIV, SIV, Vδ1, Vδ2
Introduction
γδ T cells are a subset of T cells that express a distinct T‐cell receptor (TCR) consisting of a γ and a δ‐chain. This allows γδ T cells to respond rapidly to nonpeptide antigens without the requirement of MHC presentation. In humans, γδ T cells have a relatively restricted repertoire of V gene segments and the most commonly used Vδ gene segments are Vδ1, Vδ2 and Vδ3. Human Vδ1 T cells predominantly reside in tissue and can make up to 40% of the intraepithelial lymphocytes (IEL) in the gut epithelia.1 Vδ1 T cells are also distributed in other tissues including dermis, spleen and liver, where they are involved in sustaining homeostasis and maintaining epithelial tissue integrity.2 Although Vδ1 T cells are also present in peripheral blood, Vδ2 T cells constitute the majority of human blood γδ T cells3 where they almost exclusively associate with the Vγ9 chain. Unlike Vδ1 T cells, which typically recognise CD1c and CD1d via the TCR, the Vγ9Vδ2 T cells recognise intermediate metabolites from the isoprenoid biosynthesis pathway, such as the host molecule isopentenyl pyrophosphate (IPP) or the pathogen‐associated molecule (E)‐4‐hydroxy‐3‐methyl‐but‐2‐enyl pyrophosphate (HMBPP).4, 5, 6, 7, 8 These phosphoantigens bind the protein BTN3A1, triggering an ‘inside‐out’ conformational change that promotes BTN3A1 binding to the Vγ9Vδ2 TCR and subsequent T‐cell activation.9 Human Vδ3 T cells have also been reported to recognise CD1d, but are usually only present in peripheral blood at low frequencies.10 However, they have been observed to expand in the blood in patients with cytomegalovirus infection, CD4 T‐cell deficiency and B‐cell leukaemia.11, 12, 13, 14, 15
HIV‐1 (herein referred to as HIV) infection still remains one of the most challenging health issues worldwide. In 2017, an estimated 36.9 million people were currently infected with the virus of which 1.8 million were children under the age of 15. Despite increasing awareness of the disease and improved access to antiretroviral therapy (ART), approximately 5000 individuals become newly infected every day.16 In addition to infecting and depleting CD4+ T cells, HIV infection also has a wide overall effect on the immune system, mediated largely by the phenomenon of microbial translocation.17 Rapid replication of HIV in gut‐associated lymphoid tissue (GALT) results in substantial damage to the gut epithelial barrier and the subsequent translocation of microbial products such as LPS into the circulation.18 This results in chronic immune activation and, consequently, dysfunction of conventional, bystander αβT cells. Upregulation of HLA‐DR and CD38 by T cells is associated with chronic immune activation and has proven to be strong predictors of disease progression.19, 20, 21, 22 Additionally, markers such as PD‐1, CD57 and CD100 have been used to define terminally differentiated, exhausted or dysfunctional T cells23, 24, 25, 26, 27 and are now the target of immunotherapies aimed at reducing T‐cell exhaustion.28
While HIV is well known to impact the function and distribution of conventional T‐cell subsets, the impact of the disease on γδ T‐cell subsets has been an ongoing subject of research since the late 1980s.29 Multiple reports have since described an inversion of the typical Vδ2:Vδ1 T‐cell ratio in the peripheral blood of HIV‐infected/AIDS patients,30, 31, 32, 33 which was quickly determined to represent an increase in Vδ1 T‐cell frequency30, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44 and a depletion of Vδ2Vγ9 T cells.31, 32, 33, 35, 38, 41, 42, 43, 44, 45, 46, 47, 48 Subsequent investigations have focused on the mechanisms behind this expansion/depletion, as well as the relationship of γδ T subsets to HIV disease progression. Here, we focus on reviewing the effect of HIV infection and antiretroviral therapy on Vδ1 and Vδ2 T‐cell subsets, with a primary emphasis on observations obtained from human cohorts (summarised in Figure 1). Approaches to rescue and harness γδ T‐cell responses as a means of anti‐HIV immunotherapy are discussed, as well as the future directions of this rapidly evolving field.
Figure 1.
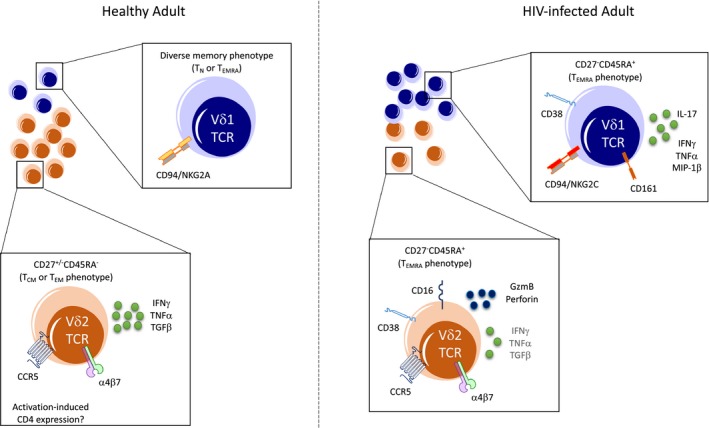
Circulating γδ T‐cell subsets in healthy and HIV‐infected adults. In healthy individuals, Vδ2 T cells (orange) comprise the majority of the circulating γδ population, with only a minority of cells expressing a Vδ1 TCR (blue). The predominant memory phenotype, surface receptor expression and functional profile of each subset are indicated for healthy (left) and HIV‐infected (right) individuals.
Progressive HIV‐1 infection
Peripheral Vδ2Vγ9 T cells
Loss of Vδ2 T cells during untreated HIV disease correlates strongly with CD4 count and viral load and occurs quickly after infection.46 Depletion preferentially affects Vδ2 cells with a Vγ9‐Jγ1.2 TCR rearrangement49 associated with the major circulating phosphoantigen‐reactive Vγ9+ Vδ2+ cell population.50 Given early reports that Vδ2 T cells lacked expression of CD451 and were resistant to direct HIV infection,52 there has been a strong emphasis on determining the mechanism of peripheral depletion, albeit with little consensus. Li and Pauza presented evidence implicating the HIV gp120 binding proteins α4β7 and CCR5 in mediating Vδ2 depletion in viremic patients.53 gp120‐induced cross‐linking of CCR5 and α4β7 on CD4− Vδ2 T cells results in the activation of the p38 caspase pathway and eventual apoptosis, without productive infection of the cell. To date, this mechanism remains to be confirmed, but analysis of existing data from preclinical studies of α4β7 monoclonal antibody (mAb) blockade54, 55, 56 may provide interesting insights into this phenomenon. An alternative hypothesis suggests that microbial translocation associated with acute HIV infection could drive Vδ2 T‐cell activation and apoptosis. Analysis of a cohort of 79 acutely infected men suggested, however, that there is no relationship between biomarkers of microbial translocation and Vδ2 frequency.32 While two studies have reported a correlation between microbial translocation and bulk γδ T‐cell activation, it is unclear whether this relationship reflects activation of the Vδ1 or Vδ2 population, or simply the change in Vδ1:Vδ2 ratio.32, 33
Vδ2 depletion is also associated with a reduction of antigen‐induced IFNγ46/TNFα57/TGF‐β47 production and proliferative/cytotoxic capacity48, 58, 59 in the residual Vδ2 population. Changes in Vδ2 cytokine production may be related in part to the relative expansion of terminally differentiated memory (TEMRA) cells and loss of the central memory (TCM) subset during chronic infection.32, 33, 47 Although all Vδ2 memory subsets exhibit significant increases in activation during acute and chronic HIV infection,33, 47 TEMRA cells tend to exhibit the highest levels of activation as measured by CD38 expression.33 However, no studies have conclusively demonstrated a causal relationship between memory differentiation or activation and loss of antigen‐induced cytokine responses in HIV.
Interesting data regarding the relationship between HIV viremia and Vδ2 T cells come from a study of structured treatment interruption (STI), where participants receiving antiretroviral therapy (ART) ceased treatment for 4–6 weeks. Within the first month following viral rebound, the Vδ2 compartment lost nearly all capacity for IFNγ production in response to antigen stimulation.60 While slower, Vδ2 T‐cell counts and the frequency of TCM cells both declined by the end of the STI. All perturbations in the Vδ2 population were restored within a month of resumption of ART, demonstrating the rapid and reversible nature of the Vδ2 response to viremia.60 Although a mechanistic explanation of this relationship is still lacking, it is possible that Vδ2 T‐cell anergy is induced by productive HIV infection of DCs, which can inhibit Vδ2 responses to phosphoantigen in vitro 61 in a contact‐dependent manner.62
Peripheral Vδ1 T cells
The destructive impact of HIV infection on Vδ2 T cells stands in direct contrast to the observed expansion of peripheral Vδ1 T cells,32, 36, 37, 38 which occurs rapidly during acute infection (and prior to HIV seroconversion38). Several studies have confirmed that the relative enrichment of Vδ1 cells as a proportion of the total γδ T‐cell population is also reflected as an increase in absolute Vδ133, 38 (or Vδ2‐47) T‐cell count in the periphery. Similar to Vδ2 cells, Vδ1 cells are significantly more activated in HIV‐infected subjects compared with controls.37, 40, 47 In acute, chronic and naturally controlled infection, the majority of Vδ1 cells exhibit a TEMRA phenotype,37, 40, 41, 44 which correlates with absolute Vδ1 T‐cell counts47 and is suggestive of antigen‐driven proliferation and activation.41 However, observations that Vδ1 T‐cell frequency is increased even in elite or viremic controllers (with low or undetectable viral loads)40, 44 suggest that HIV replication itself does not drive this expansion. Rather, Vδ1 expansion may be linked to microbial translocation, as nonhuman primate models have shown a correlation between lymph node Eschericia coli levels and peripheral Vδ1 frequency.63 Another potential mechanism for the accumulation of peripheral Vδ1 cells lies in the ability of Tat peptides to block Vδ1 chemotaxis,36 which may reduce Vδ1 cell recruitment from the periphery to mucosal sites, a phenomenon which is supported by some evidence from nonhuman primates (NHPs).64 Notably, however, evidence exists for the simultaneous accumulation of Vδ1 cells in both the periphery and gut mucosa in humans,38, 65 as well as the periphery and multiple tissues in NHP models,63 implying that the mechanisms underlying Vδ1 expansion are likely multifactorial.
Recently, studies have begun to assess comprehensively Vδ1 phenotype and function during HIV infection. Fenoglio and colleagues demonstrated that expanded Vδ1 cells in HIV‐infected subjects respond to Candida albicans stimulation and coexpress IFNγ and IL‐17. This is associated with TBX21 (Tbet), RORC, CD161, CCR4 and CCR6 expression.66 Interestingly, a substantial proportion (mean ~40%) of Vδ1 cells from this HIV‐infected cohort expressed IFNγ directly ex vivo, suggesting that circulating Vδ1 cells exist in a highly activated state. These data are consistent with those of Olson et al.,40 which demonstrated a 15‐fold elevation in mitogen‐induced IFNγ + TNFα + MIP‐1β+ ‘proinflammatory’ Vδ1 cells in viremic HIV‐infected subjects. Further characterisation of the reactivity of the expanded Vδ1 cells will be required, however, as Olson et al. failed to identify any IL‐17 production or Candida albicans reactivity by the Vδ1 subset, in direct contrast to the results of Fenoglio et al. Furthermore, a study in SIV‐infected NHPs found that Vδ1 cells did express low levels of IL‐17 in response to mitogen, but that IL‐17 production was significantly reduced in SIV+ animals compared with controls.63 Differences in antigen reactivity and IL‐17 production may be related to the duration of stimulation and cell culture, particularly in the case of Candida, but will need to be resolved in future studies.
In addition to cytokine production, clinically relevant characteristics of Vδ1 cells include the expression of NK cell receptors (NKR) and the capacity to mediate cellular cytotoxicity of HIV‐infected target and bystander cells.43, 67 Assessment of NKG2A and NKG2C expression, which initiates inhibitory or activating signals in response to HLA‐E binding, respectively, demonstrated that Vδ1 cells from HIV‐infected subjects progressively lose NKG2A expression and acquire NKG2C.43 In vitro, these NKG2C+ Vδ1 cells can recognise and kill HIV‐infected CD4+ T cells.43 The modulation of CD94, NKG2A and NKG2C on Vδ1 cells in HIV has interesting implications for the regulation of Vδ1 cell function. Studies of NK cells have shown that the NKG2A+ NK cell subset contains the highest frequency of NK cells capable of recognising autologous HIV‐infected CD4+ T cells,68 possibly due to HLA‐E presentation of a capsid‐derived peptide that blocks NKG2A inhibitory signalling.69 Conversely, elevation of HLA‐A, and subsequently HLA‐E, expression during infection is associated with poor immunological control of HIV, which is speculated to occur due to NKG2A‐mediated inhibition of NK cell function.70 Blockade of NKG2A in vitro suggests that Vδ1 cells, unlike NK cells, might be relatively resistant to NKG2A‐mediated inhibitory signalling.43 The role for CD94/NKG2A+ Vδ1 cells to control HIV replication or to be inhibited by HLA‐E expression during disease therefore remains to be determined.
Mucosal γδ T‐cell subsets
While studies of peripheral blood samples provide important insights into γδ T‐cell biology, Vδ1 cells are naturally enriched in the same mucosal tissues that support HIV replication (i.e. the gut mucosa38, 41 and female reproductive tract42). Numbers (and frequency) of duodenal γδ T cells (mostly Vδ1+) are significantly increased among HIV‐infected subjects compared with controls.65 This was confirmed by a detailed study from Poles and colleagues, who compared Vδ1 and Vδ2 subset frequencies in the peripheral blood and rectal mucosa of healthy and HIV‐infected participants.38 γδ T‐cell dynamics in the gut reflected those of the peripheral blood, with significant increases in Vδ1 and decreases in Vδ2 frequency during infection. Despite the parallel dynamics of the γδ T‐cell populations at these two site, analysis of CDR3 length showed little overlap between the two anatomical sites for either Vδ1 or Vδ2 subsets, as well as evidence of private, polyclonal expansions.38 In contrast to these results, a study of 15 acutely and 14 chronically infected participants found a significant loss of Vδ1 cells in the duodenum during chronic infection, with no change in Vδ2 frequency.41 Duodenal Vδ1 cells of chronically‐infected participants exhibited an increase in TEMRA differentiation compared with controls, although mucosal Vδ1 cells were predominately TEM phenotype, which is distinct from the peripheral blood. Beyond differences in anatomical sampling location (duodenum versus rectum), there are limited data available to explain the discrepancies in these studies.
To date, only one study has assessed the impact of HIV infection on γδ T cells at the female reproductive tract and involved mostly participants receiving ART. In this group, HIV infection was associated with a significant reduction in both Vδ1 and Vδ2 frequencies at the endocervix,42 but memory distribution, NKR expression or function was not assessed.
Impact of ART on γδ T‐cell populations
Vδ2 T cells
Numerous studies have assessed Vδ1 and Vδ2 T‐cell frequencies in ART‐treated cohorts, although substantially fewer have provided more comprehensive data regarding phenotype and function. Both cross‐sectional and longitudinal cohort studies find that ART fails to restore normal frequencies or numbers of Vδ2 T cells.32, 33, 38, 71, 72, 73 This observation is corroborated by evidence that ART only partially restores the depletion of Jγ1.2 TCR repertoire, with almost no subjects exhibiting a typical frequency of Jγ1.2 chains within the Vδ2 subset49, 73, 74, 75 and few intraparticipant changes in a longitudinal study.76 Phenotypically, more studies report residual activation of the Vδ2 subset during ART compared with healthy controls33, 71 than normalisation of activation.47 Data on memory subset distribution is more controversial, with some evidence that the expanded TEMRA population persists during ART,71, 73 while other studies show a reduction in TEMRA frequencies that closely resemble uninfected controls.32, 33, 47 Functionally, the majority of evidence suggests that Vδ2 cytokine production,47, 57, 71, 72 GzmB expression/cytotoxicity73 and proliferative capacity75 remain compromised during ART, with only a single study showing a beneficial impact of ART on Vδ2 proliferation and TNFα secretion.59
Vδ1 T cells
Cross‐sectional data support the maintenance of an expanded Vδ1 cell population during viral suppression,32, 33, 38, 40, 47, 57, 71 an observation that was also confirmed in the longitudinal follow‐up of 8 subjects from the day of ART initiation through day 540 on therapy.38 At mucosal sites, the population of expanded γδ T cells is maintained during ART,77 with only modest normalisation in some individuals.38 The peripheral Vδ1 subset in ART cohorts retains the TEMRA phenotype associated with untreated infection40, 47 and is reported to express elevated levels of PD‐1 compared with healthy controls.71 Whether ART reduces Vδ1 activation is unresolved,47, 71 but evidence suggests ongoing Vδ1 proliferation during viral suppression47 and the maintenance of a large proinflammatory IFNγ + TNFα + MIP‐1β+ polyfunctional population.40 A single study of NKR expression reported elevated levels of CD94/NKG2A, CD158a, CD158b and NKB1 on Vδ1 cells compared with healthy controls,78 which was accompanied by a loss of CD28 expression and an upregulation of CD45RO.78
Microbial translocation, immune activation and apoptosis
The relationship between microbial translocation, systemic immune activation and perturbations of γδ T‐cell subsets has been a common and enduring thread throughout the studies described above. Microbial translocation occurs as a result of the massive depletion of CD4+ T cells at the gut mucosa during acute HIV or SIV infection.79, 80 This includes the preferential loss of Th17 cells,81 which contribute to the maintenance of epithelial barrier integrity and wound healing in the gastrointestinal mucosa. The dysregulation of mucosal immunity and loss of epithelial integrity allow the translocation of microbial products such as LPS into the circulation, resulting in systemic immune activation and inflammation17 (recently reviewed in References 80 and 82). Unfortunately, suppressive ART is unable to fully restore the mucosal CD4+ T‐cell compartment and abolish systemic inflammation.80
The disruption of mucosal immunity not only allows microbial products to translocate into the circulation, but also alters the composition of the gut microbiota. A number of human cohort studies have consistently shown an enrichment of proinflammatory bacteria such as the Enterobacteriaceae family and a loss of clades such as Bacteriodes.80 Enterobacteriaceae are particularly likely to translocate across the mucosal barrier and induce the production of reactive oxygen species by innate immune cells, driving inflammation. At least one member of this family, E. coli, activates Vδ2 T cells in vitro, raising the possibility that sustained exposure to translocated microbes could drive apoptosis of γδ T cells, similar to the apoptosis of bystander CD4+ in the lamina propria that occurs by Fas‐FasL interactions.79 While the impact of microbiome perturbations on the gut‐resident Vδ1 T‐cell population remains to be understood, data have shown a correlation between iNKT cells in the gut mucosa and the prevalence of Bacteroides and Prevotella microbes.83 Thus, γδ T cells may be directly impacted by changes in the microbial community at the gut mucosa, the translocation of proinflammatory products into the circulation, or dysregulation of innate and adaptive immune cells during both untreated and treated HIV infection.
Clinical impact of γδ T‐cell perturbations during HIV infection
Contribution to HIV control
Whether γδ T‐cell activation and cytolytic capacity during HIV infection can actually contribute to control of viremia or disease progression remains an open question. γδ cells can undoubtedly control HIV replication through multiple mechanisms in vitro, including direct cytotoxicity of infected cells.43, 84 Vδ2 T cells can be recruited to HIV‐infected DCs via CCL4 production, where they control viral replication and reduce HIV transmission to bystander CD4+ T cells.61 β‐Chemokine production by both Vδ185 and Vδ286 cells can block HIV infection of target cells. However, whether this in vitro activity translates into in vivo control of viremia remains unresolved. Although HIV elite/viral controllers exhibit Vδ2 depletion relative to healthy controls, they maintain Vδ2 frequencies that are significantly higher than either untreated or ART‐treated subjects.44, 87 These cells predominately exhibit a TCM phenotype87 and produce more IL‐17 than cells from viremic patients.44 Unfortunately, such studies are confounded by an inability to determine whether viral control preserves γδ T‐cell ratios and phenotypes, or whether maintenance of a TCM/IL‐17‐expressing Vδ2 population contributes to the control of viral replication. Nonhuman primates offer a unique opportunity to longitudinally compare preinfection γδ populations to infection susceptibility and viral load setpoint. Although Tuero et al.88 reported an inverse correlation between endocervical Vδ2 T‐cell frequency and chronic viral load (VL) in SIV‐infected macaques, supporting a potential protective role for γδ T cells in this animal model, these studies are still lacking in the literature and this should be further investigated in future studies.
Impact on coinfection
While suppressive ART successfully controls HIV replication regardless of γδ T‐cell reconstitution, residual impairment of the Vδ2 subset likely has profound implications for immunity against a number HIV coinfections. Tuberculosis is currently the leading cause of death among HIV‐infected individuals, and evidence suggests that active TB and HIV infection have additive effects on peripheral Vδ2 depletion89 and dysfunction.90 At the site of TB infection, however, it is unclear what impact HIV has on lung γδ T‐cell populations. Only a single study has reported BAL γδ T‐cell numbers in healthy and HIV‐infected participants where there was a significant increase in total γδ cells during HIV infection, but the delta chain usage was not determined.91 However, the possible impact of HIV infection on Vδ2 responses to Mycobacterium has been clearly demonstrated in NHP models. Naïve macaques are able to induce robust primary and recall Vδ2 responses to BCG vaccination in the periphery and lung, while SIV‐infected macaques showed no response to BCG in either site.92 Encouragingly, administration of ART improved NHP Vδ2 responses to BCG, possibly due to reconstitution of Mtb‐specific CD4+ T cells.93 NHP studies will be critical in determining whether host‐directed therapy targeting Vδ2 T cells94 can enhance protection against TB reactivation in HIV‐infected populations.
Similarly, the expansion of Vδ1 cells during HIV infection may impact coinfection with several herpesviruses. Cytomegalovirus (CMV) infection is a widespread pathogen that usually causes asymptomatic infections. However, in HIV‐infected individuals, this pathogen can result in clinical manifestations including chorioretinitis and CMV enterocolitis. Similar to HIV infection, there is a selective expansion of Vδ2− cells during CMV infection.95 These cells are suggested to participate in the control of CMV replication and display potent anti‐CMV responses in vitro.12 Although the expansion of Vδ1 cells as a result of HIV infection would presumably be beneficial for control of CMV infection, CMV replication is enhanced by inflammatory stimuli. Since it is reported that the functional characteristics of the expanded Vδ1 cells in HIV+ individuals are skewed towards a proinflammatory profile, this may instead contribute to CMV‐associated morbidity, although this remains to be determined.96, 97, 98, 99 Furthermore, it is unclear whether the high prevalence of CMV infection among HIV+ individuals is a driver of the Vδ1 T‐cell expansion observed during chronic infection.
Human herpesvirus 8 (HHV8) is also a virus which has increased seroprevalence in HIV‐infected individuals and can cause significant disease in the form of Kaposi's sarcoma (KS).100 Although the effector populations involved in control of this virus remain elusive, HHV8 infection is also associated with an expansion of Vδ1 cells that respond to HHV8‐infected cells and prevent virus release in immunocompetent hosts.101 The role γδ T cells play in promoting HHV‐8 to progress to KS is currently unknown, but considering that inflammatory cytokines including IFN‐γ, IL‐6, IL‐1β and TNF‐α are produced by infiltrating cells in lesions of KS, expanded Vδ1 cells in HIV+ individuals may potentially contribute to progression of clinical symptoms in a similar way as with CMV.102
Other common coinfections among HIV‐infected populations include Cryptococcus, viral hepatitis and malaria. There are little data available regarding γδ T‐cell responses to Cryptococcus infection in HIV‐infected human cohorts, but murine studies have established a role for γδ T cells in Cryptococcal immunity in the lung.103, 104 Hepatitis B and C infections have a deleterious impact on γδ T cells, similar to HIV infection, which is discussed more fully below. Studies in malaria‐endemic populations have revealed a substantial role for γδ T cells in immunity to Plasmodium spp. This topic has been recently reviewed in References 105 and 106, which highlight the protective and immunoregulatory roles of both Vδ2 and Vδ1 γδ subsets. Surprisingly, however, there are no studies of HIV/malaria coinfection that report on γδ T cells, which would represent an interesting focus for future clinical cohorts.
Impact on immune cell crosstalk
γδ T cells can not only exert direct antimicrobial activity, but can also orchestrate and regulate the activation, maturation and recruitment of a variety of other immune cells.107, 108 Activated Vδ2 cells from healthy individuals can induce the maturation of neutrophils,109 DCs and B cells into APCs.110 Some evidence that this function is compromised during HIV infection, as HIV‐infected APCs cannot undergo full γδ T cell‐induced maturation in vitro, leading to high‐residual CCR5 expression and low CD86 and HLA‐DR expression.62 This impairment of crosstalk may enhance DC susceptibility to infection through CCR5 expression, as well as compromise CD4+ T‐cell responses that rely on DC‐mediated antigen presentation. To date, however, follow‐up on these observations is lacking. Additionally, Vδ2 cells can themselves present antigen and act as APCs for conventional αβ T cells.111 Antigen‐activated Vδ2 cells express HLA‐DR, CD80, CD86, CD40 and CD54 at levels comparable to LPS‐matured DCs and can induce primary αβ T‐cell responses.111 Transient activation‐induced upregulation of CCR7 implies that Vδ2 may home to draining lymph nodes during infection initiate adaptive immune responses. Considering the residual depletion, terminal differentiation and dysfunction that characterise Vδ2 cells in ART‐treated subjects, it is likely that acquisition of APC function following antigen stimulation is compromised in the context of HIV infection. The contribution of such dysfunction to poor antimicrobial immunity or vaccination in ART‐treated subjects should be further explored, as in vivo Vδ2 immunotherapy could be considered to address these defects in immune function.
More recently, Vδ2 cells have been recognised to provide CD40L‐dependent help to B cells.112, 113, 114, 115, 116 As noted above, the transient expression of CCR7 after activation allows Vδ2 cells to traffic to secondary lymphoid tissues, where they cluster within the germinal centre of mucosal B‐cell follicles.116 Stimulation with the phosphoantigen IPP is sufficient to elicit the delayed but robust expression of surface molecules involved in B‐cell help; 36–84 h poststimulation, Vδ2 T cells express CD40L, ICOS, OX‐40 and CD70.116 In vitro coculture assays indicate that activated Vδ2 cells can promote B‐cell antibody secretion to a similar, or even increased, degree as TFH cells.116 Further studies have suggested that antigen exposure in the presence of IL‐21 is required to induce the expression of CD40L and ICOS on circulating Vδ2 cells.113, 114 Interestingly, treatment of macaques with intravenous HMBPP and IL‐2 during chronic SHIV infection resulted a prolonged boosting of SHIVenv‐specific antibody titres, suggesting that Vδ2 cells can contribute to humoral immunity in vivo. Despite these fascinating observations, further data regarding the impact of Vδ2 depletion on humoral immunity during HIV infection are lacking.
γδ T cells in anti‐HIV immunotherapy
Interest is increasing in developing host‐directed immunotherapies to either supplement or replace current ART. Studies which have investigated the use of Vδ2 cells for anti‐HIV immunotherapy are summarised in Table 1. At the simplest level, methods for recovering Vδ2 responses to phosphoantigen among HIV‐infected donors include cytokine supplementation with IL‐18117 or IL‐12.61 Such an approach may improve Vδ2‐mediated immune responses against Mycobacterium or other bacterial infections. More complex interventions designed to specifically target HIV‐infected cells include an effort to harness the ability of Vδ2 T cells to perform antibody‐dependent cellular cytotoxicity (ADCC) via CD16 expression. CD16+ Vδ2 cells exhibit poor responses to phosphoantigen, but respond robustly to antibody‐coated target cells.118 These responses are largely maintained, if not slightly enhanced, in ART‐treated subjects, suggesting that Vδ2 cells derived from HIV‐infected individuals could contribute to the killing of HIV‐infected target cells.118 More encouragingly, ex vivo expansion of Vδ2 cells from HIV‐infected subjects results in an upregulation of CD16 expression and quantifiable ADCC of antibody‐coated targets.119
Table 1.
Summary of HIV immunotherapy studies using Vδ2 T cells
| Outcome | Study | Species | HIV Status | Cell Type | Results |
|---|---|---|---|---|---|
| Antigen responses | Murday et al.116 | Human | HIV+ ART | Ex vivo Vδ2 | IL‐18 stimulation improves IPP‐induced Vδ2 proliferation in HIV+ individuals |
| Cardone61 | Human | Healthy | Ex vivo Vδ2 and HIV‐infected monocyte‐derived DC | Vδ2 cell phosphoantigen responses in the presence of HIV‐infected DC are inhibited due to poor IL‐12 secretion by the DCs. Responses can be restored by addition of IL‐12 to Vδ2/DC cocultures | |
| ADCC | He et al.117 | Human | HIV+ ART | Ex vivo Vδ2 | Vδ2 cells from ART‐treated individuals exhibit CD16 expression and degranulate in response to CD16‐mediated activation |
| Poonia et al.118 | Human | Healthy, HIV+ ART | IPP/zoledronate + IL‐2 expanded PBMC | Expanded Vδ2 cells expressed CD16 and were capable of killing antibody‐coated target cells | |
| Direct cytotoxicity | Poonia et al.118 | Human | Healthy, HIV+ ART | IPP/zoledronate + IL‐2 expanded PBMC | Vδ2 cells exhibited direct cytotoxicity against Daudi cells. IPP‐expanded cells were more potent killers than zoledronate‐expanded cells |
| Garrido et al.119 | Human | HIV+ ART | Bisphosphonate pamidronate (PAM) + IL‐2 expanded PBMC | PAM‐expanded cells degranulated and inhibited in vitro HIV replication in CD4+ T cells. p24 production following vorinostat‐mediated reactivation of latent HIV from primary CD4+ T cells was suppressed in the presence of PAM‐expanded Vδ2 cells | |
| In vivo expansion | Ali et al.120 | Macaque | SHIV‐infected (acute or chronic) | Injection of HMBPP + IL‐2 in vivo | Treatment resulted in expansion and activation of Vδ2 cells. Treatment during acute infection exacerbated viral replication and disease progression in an IL‐2‐dependent manner. Treatment during chronic infection boosted Env‐specific antibody titres but did not impact viral load or disease progression |
| Poonia et al.121 | Humanised mice | HIV+ | Adoptive transfer of zoledronate + IL‐2 expanded PBMC | No impact of expanded Vδ2 T‐cell transfer on CD4+ T‐cell loss, CD4:CD8 T‐cell ratio or viral load |
Any immunotherapy‐based intervention using autologous Vδ2 cells will require in vitro expansion to generate sufficiently large numbers of cells for reinfusion. Ex vivo antigen‐driven Vδ2 T‐cell expansion appears to be a viable and reproducible strategy for the production of large numbers of autologous Vδ2 cells from ART‐treated HIV‐infected individuals, although optimal expansion culture conditions may differ between healthy and infected groups.120 Expanded cells express low levels of inhibitory surface receptors and can kill latently infected CD4+ T cells after latency reversal with vorinostat in vitro.120 In vivo, however, results of γδ T‐cell immunotherapy are varied. Administration of HMBPP and IL‐2 to chronically SIV‐infected macaques expanded and activated Vδ2 T cells, transiently boosted SHIV‐specific CD8+ T‐cell responses and resulted in a sustained increase of SHIV‐specific antibody titres.121 Nonetheless, there was no impact of Vδ2 expansion on viral load or disease progression during chronic infection, and a negative impact of Vδ2 expansion during acute infection. These results were mirrored in a study of humanised mice treated with expanded Vδ2 cells, which similarly observed no protective effect of Vδ2 treatment on viral replication or CD4+ T‐cell depletion.122
Similarities with other chronic inflammatory diseases
The hallmark impacts of HIV infection on γδ T cells (Vδ2:Vδ1 ratio inversion, activation and terminal differentiation, functional defects) are, in fact, not unique to HIV infection itself. Other chronic inflammatory diseases are associated with similar effects, including kidney disease, viral hepatitis and obesity. Understanding the commonalities in pathogenesis between these diverse conditions may provide further insight into the mechanisms of γδ perturbation and identify useful therapeutic targets.
Chronic kidney disease (CKD) is a progressive condition in which the loss of renal function results in the accumulation of uraemic toxins and proinflammatory cytokines (reviewed in Reference 123). End‐stage renal disease (ESRD), the final stage of CKD, is associated with high levels of immune activation, poor responses to immunisation and high susceptibility to infection. Matsumoto first reported a significant loss of γδ T cells among CKD patients requiring hemodialysis.124 They speculated that γδ depletion likely occurred because of Fas‐ and LFA‐1‐dependent apoptosis related to uraemia. Similarly, we observed a significant loss of phosphoantigen‐reactive γδ T cells in ESRD patients compared with healthy controls.125 Surprisingly, however, there was no relationship between plasma proinflammatory cytokine levels and γδ frequency or dysfunction,125 raising the question of what drives γδ T‐cell loss during ESRD. Similar to HIV infection, it is unclear whether low peripheral γδ frequencies truly reflect apoptosis or, instead, recruitment to inflamed tissues. γδ frequencies only partially normalise following kidney transplantation, with Vδ2 frequencies remaining significantly lower than healthy controls.126 The fact that uraemia‐associated changes in the γδ repertoire are not effectively reversed upon transplantation suggests that transplant patients may exhibit long‐term susceptibility to some infections, similar to HIV ART‐treated patients.
Viral hepatitis is also associated with changes in the circulating γδ repertoire that are highly reminiscent of HIV infection. Chronic HCV infection is associated with peripheral Vδ2 depletion,127, 128 acquisition of an activated/TEMRA phenotype,128, 129 upregulation of CD16 and granzyme129 and functional impairment.128, 129 HBV‐infected subjects exhibit loss of peripheral Vδ2 cells130, 131 and/or expansion of peripheral Vδ1 cells,132 which correlates with serum ALT levels.130, 131 Residual Vδ2 cells from these individuals are impaired for IFNγ production, cytotoxicity130 and proliferation131 and exhibit an activated, TEMRA surface phenotype.131 Expression of granzyme and cytotoxic markers is particularly enhanced in HCV‐infected subjects with greater degrees of liver damage, suggesting the potential involvement of γδ T cells in mediating immunopathology during infection. Despite this, phosphoantigen‐activated Vδ2 cells can restrict in vitro HCV replication in an IFNγ‐dependent manner,133 making them potential immunotherapeutic targets for HCV treatment. In vitro studies suggested that Vδ2 dysfunction may be at least partially abrogated by treatment with IFNα, which boosts phosphoantigen responses in γδ T cells from both healthy and HCV‐infected subjects128 (although this effect was not replicated by others129). In vivo, however, a standard course of Peg‐IFNα and ribavirin therapy resulted in pronounced Vδ2 anergy after 4 weeks of treatment in two studies of chronically HCV‐infected patients.129, 134 Notably, although Vδ2 IFNγ responses were decreased/almost absent after treatment, perforin and degranulation responses were elevated, suggesting the possibility that IFNα drives a transition of Vδ2 cells away from cytokine responses and towards cytotoxicity.129 The results of these clinical trials highlight important considerations for the in vivo use of drugs to promote Vδ2 activation and proliferation during treated HIV infection, and the need to assess anergy at multiple timepoints after therapy.
Similar to CKD patients, obese adults exhibit increased susceptibility to infection associated with chronic inflammation.135, 136 Peripheral Vδ2 T cells are depleted in obese individuals (in a BMI‐dependent manner),137, 138 are more likely to exhibit a TEMRA phenotype and respond poorly to influenza‐infected APCs,138 mirroring the effects of HIV infection, HBV/HCV infection and renal disease on this compartment. As expected, cytokine supplementation overcomes some of the Vδ2 function defect, with IL‐2 boosting in vitro Vδ2 function among obese subjects.138
Interestingly, these three conditions, as well as HIV infection, all involve some degree of gut dysbiosis. As previously discussed, HIV infection results in substantial damage to the gut epithelium and microbial translocation, which causes widespread immune activation.139 Microbial translocation has also been reported in ESRD/CKD cohorts140, 141, 142 and is increasingly being recognised as an important driver of T‐cell dysfunction and chronic inflammation.143, 144 Indeed, we found that plasma sCD14 levels in ESRD patients correlate with HMBPP‐induced IFNg production by γδ T cells, suggesting a possible link between microbial translocation and Vδ2 T‐cell function.125 Similar observations have been made in the context of chronic HCV/HBV infection and liver cirrhosis, where overgrowth of pathogenic gut bacteria increases gut permeability and allows translocation of bacterial products into the liver via the portal vein.139, 145, 146 Finally, obese individuals also exhibit elevated levels of LPS and other markers of microbial translocation, supporting a putative link between gut permeability, low‐grade inflammation, and γδ T‐cell depletion and dysfunction.147, 148
Unresolved questions and future directions
As the field of HIV immunology moves forward from studies of HIV pathogenesis towards a focus on inflammation and immune ageing among ART‐treated populations,149 there are several key questions surrounding γδ T‐cell immunology that remain unanswered. First, the link between poor reconstitution of the γδ compartment and persistent innate immune activation and inflammation during ART is poorly understood. In a fascinating study, Belkina et al. comprehensively assessed the expression of inhibitory surface receptors on a wide range of lymphocyte subsets including two NK cell populations, conventional T cells, Tregs, iNKT cells and γδT cells in ART‐treated and control participants.150 Importantly, these cohorts were stratified for age, allowing for a simultaneous assessment of immune ageing in each group. Among all lymphocyte subsets, only γδ phenotype was sufficient to distinguish between the control and infected groups. A transition of the γδ compartment from ‘resting’ CD160+ phenotype to an ‘activated/exhausted’ TIGIT + PD‐1+ phenotype was associated with plasma‐derived proinflammatory profile.150 While an inversion of the Vδ2:Vδ1 ratio was confirmed for a subset of HIV‐infected study participants, no data were available to assess the separate contribution of Vδ1 and Vδ2 cells to the HIV‐associated inflammation and ageing. Such information will be critical to understanding which γδ subset primarily expressed the TIGIT + PD‐1+ phenotype and/or correlates mostly strongly with plasma inflammatory biomarkers. In addition to these data, a transcriptomics study of mitogen‐activated lymphocyte responses identified γδ T‐cell differentiation as a differentially regulated pathway between healthy control and long‐term ART cohorts.149
Second, the field will benefit from a better understanding of the composition of the expanded Vδ1 subset in ART cohorts. Vδ1 T cells include CD1‐restricted, lipid‐reactive T cells, as well as cells with undefined antigen specificity and numerous mechanisms to sense host cell stress. As studies undertake novel approaches to defining Vδ1 T‐cell subsets with different functions and phenotypes,151 we will move closer to understanding what drives the dramatic peripheral expansion and proinflammatory cytokine profile of these cells. It is interesting to note that the phenomena of microbial translocation and gut dysbiosis are common to many chronic inflammatory diseases associated with changes in the Vδ1:Vδ2 T‐cell ratio. Recent data from murine models highlight the physiological importance of gut‐derived γδ cells and their ability to traffic to inflamed tissues, including the brain.152 Whether gut dysbiosis is a predominate driver of γδ dysfunction and accumulation during HIV or other viral infections remains to be fully investigated.
Finally, the question of whether productive HIV infection of Vδ2 T cells occurs in vivo remains a matter of debate. Commonly, Vδ2 T cells from healthy individuals are reported to be CD4‐ 51, 53, 75, 153, 154, ostensibly rendering them impermissible to infection. Surprisingly, however, Wallace et al.58 reported in 1997 that Vδ2 T‐cell lines could be productively infected with HIV in vitro over the course of 18 days. Similarly, coinfection with human herpesvirus 6 can induce CD4 expression on γδ T cells in vitro, rendering them susceptible to HIV infection.52 Since then, some studies have reported low‐level CD4 expression on peripheral γδ T cells ex vivo, 155 which is sufficient to mediate CD4‐dependent productive HIV infection. Similarly, humanised mice produce thymic γδ T cells that express CD4, CCR5 and CXCR4 and are susceptible to infection by multiple HIV isolates.156 Perhaps most intriguingly, mucosal Vδ2 T cells at the female reproductive tract are reported to be predominately CD4+.42 The relevance of Vδ2 T‐cell infection by HIV was recently highlighted by data indicating that circulating Vδ2 cells are a reservoir for replication‐competent HIV in ART‐suppressed patients.154 In this study, exposure of Vδ2 T cells to IL‐2 was sufficient to induce CD4 expression on an average of 15% of isolated Vδ2 cells. Three acutely HIV‐infected subjects (<23 days postinfection) also exhibited a similar level of CD4 expression on their Vδ2 T cells, suggesting that immune activation is sufficient to promote productive infection of Vδ2 T cells in vivo. What impact this might have on Vδ2‐based immunotherapies and latency reactivation‐based HIV cure strategies is currently unknown, but should be considered in future studies.
Conflict of interest
The authors declare no conflict of interest.
Author contributions
JAJ and EME wrote and revised the manuscript.
Acknowledgments
JAJ is funded by a NHMRC fellowship.
References
- 1. Marsal J, Agace WW. Targeting T‐cell migration in inflammatory bowel disease. J Intern Med 2012; 272: 411–429. [DOI] [PubMed] [Google Scholar]
- 2. Bonneville M, O'Brien RL, Born WK. γδ T cell effector functions: a blend of innate programming and acquired plasticity. Nat Rev Immunol 2010; 10: 467–478. [DOI] [PubMed] [Google Scholar]
- 3. Vantourout P, Hayday A. Six‐of‐the‐best: unique contributions of γδ T cells to immunology. Nat Rev Immunol 2013; 13: 88–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Spada FM, Grant EP, Peters PJ et al Self‐recognition of CD1 by γ/δ T cells: implications for innate immunity. J Exp Med 2000; 191: 937–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Porcelli S, Brenner MB, Greenstein JL, Balk SP, Terhorst C, Bleicher PA. Recognition of cluster of differentiation 1 antigens by human CD4–CD8–cytolytic T lymphocytes. Nature 1989; 341: 447–450. [DOI] [PubMed] [Google Scholar]
- 6. Faure F, Jitsukawa S, Miossec C, Hercend T. CD1c as a target recognition structure for human T lymphocytes: analysis with peripheral blood γ/δ cells. Eur J Immunol 1990; 20: 703–706. [DOI] [PubMed] [Google Scholar]
- 7. Bai L, Picard D, Anderson B et al The majority of CD1d‐sulfatide‐specific T cells in human blood use a semiinvariant Vδ1 TCR. Eur J Immunol 2012; 42: 2505–2510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Gober HJ, Kistowska M, Angman L, Jeno P, Mori L, De Libero G. Human T cell receptor γδ cells recognize endogenous mevalonate metabolites in tumor cells. J Exp Med 2003; 197: 163–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Yang Y, Li L, Yuan L et al A structural change in butyrophilin upon phosphoantigen binding underlies phosphoantigen‐mediated Vγ9Vδ2 T cell activation. Immunity 2019; 50: 1043–1053.e5. [DOI] [PubMed] [Google Scholar]
- 10. Mangan BA, Dunne MR, O'Reilly VP et al Cutting edge: CD1d restriction and Th1/Th2/Th17 cytokine secretion by human Vδ3 T cells. J Immunol 2013; 191: 30–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Dechanet J, Merville P, Lim A et al Implication of γδ T cells in the human immune response to cytomegalovirus. J Clin Investig 1999; 103: 1437–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Halary F, Pitard V, Dlubek D et al Shared reactivity of Vδ2neg γδ T cells against cytomegalovirus‐infected cells and tumor intestinal epithelial cells. J Exp Med 2005; 201: 1567–1578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Knight A, Madrigal AJ, Grace S et al The role of Vδ2‐negative γδ T cells during cytomegalovirus reactivation in recipients of allogeneic stem cell transplantation. Blood 2010; 116: 2164–2172. [DOI] [PubMed] [Google Scholar]
- 14. Bartkowiak J, Kulczyck‐Wojdala D, Blonski JZ, Robak T. Molecular diversity of γδ T cells in peripheral blood from patients with B‐cell chronic lymphocytic leukaemia. Neoplasma 2002; 49: 86–90. [PubMed] [Google Scholar]
- 15. Kabelitz D, Hinz T, Dobmeyer T et al Clonal expansion of Vγ3/Vδ3‐expressing γδ T cells in an HIV‐1/2‐negative patient with CD4 T‐cell deficiency. Br J Haematol 1997; 96: 266–271. [DOI] [PubMed] [Google Scholar]
- 16. UNAIDS . UNAIDS Data 2018. 2018. ed2018. p. 1–376.
- 17. Brenchley JM, Price DA, Schacker TW et al Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med 2006; 12: 1365–1371. [DOI] [PubMed] [Google Scholar]
- 18. Dillon SM, Frank DN, Wilson CC. The gut microbiome and HIV‐1 pathogenesis: a two way street. AIDS 2016; 30: 2737–2751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Deeks SG, Kitchen CM, Liu L et al Immune activation set point during early HIV infection predicts subsequent CD4+ T‐cell changes independent of viral load. Blood 2004; 104: 942–947. [DOI] [PubMed] [Google Scholar]
- 20. Giorgi JV, Hultin LE, McKeating JA et al Shorter survival in advanced human immunodeficiency virus type 1 infection is more closely associated with T lymphocyte activation than with plasma virus burden or virus chemokine coreceptor usage. J Infect Dis 1999; 179: 859–870. [DOI] [PubMed] [Google Scholar]
- 21. Giorgi JV, Lyles RH, Matud JL et al Predictive value of immunologic and virologic markers after long or short duration of HIV‐1 infection. J Acquir Immune Defic Syndr 2002; 29: 346–355. [DOI] [PubMed] [Google Scholar]
- 22. Liu Z, Cumberland WG, Hultin LE, Kaplan AH, Detels R, Giorgi JV. CD8+ T‐lymphocyte activation in HIV‐1 disease reflects an aspect of pathogenesis distinct from viral burden and immunodeficiency. J Acquir Immune Defic Syndr Hum Retrovirol 1998; 18: 332–340. [DOI] [PubMed] [Google Scholar]
- 23. Brenchley JM, Karandikar NJ, Betts MR et al Expression of CD57 defines replicative senescence and antigen‐induced apoptotic death of CD8+ T cells. Blood 2003; 101: 2711–2720. [DOI] [PubMed] [Google Scholar]
- 24. Day CL, Kaufmann DE, Kiepiela P et al PD‐1 expression on HIV‐specific T cells is associated with T‐cell exhaustion and disease progression. Nature 2006; 443: 350–354. [DOI] [PubMed] [Google Scholar]
- 25. Petrovas C, Casazza JP, Brenchley JM et al PD‐1 is a regulator of virus‐specific CD8+ T cell survival in HIV infection. J Exp Med 2006; 203: 2281–2292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Petrovas C, Chaon B, Ambrozak DR et al Differential association of programmed death‐1 and CD57 with ex vivo survival of CD8+ T cells in HIV infection. J Immunol 2009; 183: 1120–1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Eriksson EM, Milush JM, Ho EL et al Expansion of CD8+ T cells lacking Sema4D/CD100 during HIV‐1 infection identifies a subset of T cells with decreased functional capacity. Blood 2012; 119: 745–755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Wykes MN, Lewin SR. Immune checkpoint blockade in infectious diseases. Nat Rev Immunol 2018; 18: 91–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Autran B, Triebel F, Katlama C, Rozenbaum W, Hercend T, Debre P. T cell receptor γ/δ+ lymphocyte subsets during HIV infection. Clin Exp Immunol 1989; 75: 206–210. [PMC free article] [PubMed] [Google Scholar]
- 30. De Maria A, Ferrazin A, Ferrini S, Ciccone E, Terragna A, Moretta L. Selective increase of a subset of T cell receptor γδ T lymphocytes in the peripheral blood of patients with human immunodeficiency virus type 1 infection. J Infect Dis 1992; 165: 917–919. [DOI] [PubMed] [Google Scholar]
- 31. Hinz T, Wesch D, Friese K, Reckziegel A, Arden B, Kabelitz D. T cell receptor γδ repertoire in HIV‐1‐infected individuals. Eur J Immunol 1994; 24: 3044–3049. [DOI] [PubMed] [Google Scholar]
- 32. Li Z, Li W, Li N et al γδ T cells are involved in acute HIV infection and associated with AIDS progression. PLoS One 2014; 9: e106064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Li Z, Jiao Y, Hu Y et al Distortion of memory Vδ2 γδ T cells contributes to immune dysfunction in chronic HIV infection. Cell Mol Immunol 2015; 12: 604–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. De Paoli P, Gennari D, Martelli P et al A subset of γδ lymphocytes is increased during HIV‐1 infection. Clin Exp Immunol 1991; 83: 187–191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Wesch D, Hinz T, Kabelitz D. Analysis of the TCR Vγ repertoire in healthy donors and HIV‐1‐infected individuals. Int Immunol 1998; 10: 1067–1075. [DOI] [PubMed] [Google Scholar]
- 36. Poggi A, Carosio R, Fenoglio D et al Migration of Vδ1 and Vδ2 T cells in response to CXCR18 and CXCR18 ligands in healthy donors and HIV‐1‐infected patients: competition by HIV‐1 Tat. Blood 2004; 103: 2205–2213. [DOI] [PubMed] [Google Scholar]
- 37. Rossol R, Dobmeyer JM, Dobmeyer TS et al Increase in Vδ1+ γδ T cells in the peripheral blood and bone marrow as a selective feature of HIV‐1 but not other virus infections. Br J Haematol 1998; 100: 728–734. [DOI] [PubMed] [Google Scholar]
- 38. Poles MA, Barsoum S, Yu W et al Human immunodeficiency virus type 1 induces persistent changes in mucosal and blood γδ T cells despite suppressive therapy. J Virol 2003; 77: 10456–10467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Li Z, Lu X, Hu Z et al Syphilis infection differentially regulates the phenotype and function of γδ T cells in HIV‐1‐infected patients depends on the HIV‐1 disease stage. Front Immunol 2017; 8: 991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Olson GS, Moore SW, Richter JM et al Increased frequency of systemic pro‐inflammatory Vδ1+ γδ T cells in HIV elite controllers correlates with gut viral load. Sci Rep 2018; 8: 16471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Cimini E, Agrati C, D'Offizi G et al Primary and chronic HIV infection differently modulates mucosal Vδ1 and Vδ2 T‐cells differentiation profile and effector functions. PLoS One 2015; 10: e0129771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Strbo N, Alcaide ML, Romero L et al Loss of intra‐epithelial endocervical γδ (GD) 1 T cells in HIV‐infected women. Am J Reprod Immunol 2016; 75: 134–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Fausther‐Bovendo H, Wauquier N, Cherfils‐Vicini J, Cremer I, Debre P, Vieillard V. NKG2C is a major triggering receptor involved in the Vδ1 T cell‐mediated cytotoxicity against HIV‐infected CD4 T cells. AIDS 2008; 22: 217–226. [DOI] [PubMed] [Google Scholar]
- 44. Chevalier MF, Bhatnagar N, Didier C et al γδ T‐cell subsets in HIV controllers: potential role of Tγδ17 cells in the regulation of chronic immune activation. AIDS 2019; 33: 1283–1292. [DOI] [PubMed] [Google Scholar]
- 45. Enders PJ, Yin C, Martini F et al HIV‐mediated γδ T cell depletion is specific for Vγ2+ cells expressing the Jγ1.2 segment. AIDS Res Hum Retroviruses 2003; 19: 21–29. [DOI] [PubMed] [Google Scholar]
- 46. Li H, Peng H, Ma P et al Association between Vγ2Vδ2 T cells and disease progression after infection with closely related strains of HIV in China. Clin Infect Dis 2008; 46: 1466–1472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Bhatnagar N, Girard PM, Lopez‐Gonzalez M et al Potential role of Vδ2+ γδ T cells in regulation of immune activation in primary HIV infection. Front Immunol 2017; 8: 1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Poccia F, Boullier S, Lecoeur H et al Peripheral V γ 9/V δ 2 T cell deletion and anergy to nonpeptidic mycobacterial antigens in asymptomatic HIV‐1‐infected persons. J Immunol 1996; 157: 449–461. [PubMed] [Google Scholar]
- 49. Chaudhry S, Cairo C, Venturi V, Pauza CD. The γδ T‐cell receptor repertoire is reconstituted in HIV patients after prolonged antiretroviral therapy. AIDS 2013; 27: 1557–1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Davodeau F, Peyrat MA, Hallet MM et al Close correlation between Daudi and mycobacterial antigen recognition by human γδ T cells and expression of V9JPC1 γ/V2DJC δ‐encoded T cell receptors. J Immunol 1993; 151: 1214–1223. [PubMed] [Google Scholar]
- 51. Brenner MB, McLean J, Scheft H et al Two forms of the T‐cell receptor γ protein found on peripheral blood cytotoxic T lymphocytes. Nature 1987; 325: 689–694. [DOI] [PubMed] [Google Scholar]
- 52. Lusso P, Garzino‐Demo A, Crowley RW, Malnati MS. Infection of γ/δ T lymphocytes by human herpesvirus 6: transcriptional induction of CD4 and susceptibility to HIV infection. J Exp Med 1995; 181: 1303–1310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Li H, Pauza CD. HIV envelope‐mediated, CCR5/α4β7‐dependent killing of CD4‐negative γδ T cells which are lost during progression to AIDS. Blood 2011; 118: 5824–5831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Calenda G, Keawvichit R, Arrode‐Bruses G et al Integrin α4β7 blockade preferentially impacts CCR6+ lymphocyte subsets in blood and mucosal tissues of naive rhesus macaques. J Immunol 2018; 200: 810–820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Byrareddy SN, Arthos J, Cicala C et al Sustained virologic control in SIV+ macaques after antiretroviral and α4β7 antibody therapy. Science 2016; 354: 197–202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Santangelo PJ, Cicala C, Byrareddy SN et al Early treatment of SIV+ macaques with an α4β7 mAb alters virus distribution and preserves CD4+ T cells in later stages of infection. Mucosal Immunol 2018; 11: 932–946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Kosub DA, Lehrman G, Milush JM et al γ/δ T‐cell functional responses differ after pathogenic human immunodeficiency virus and nonpathogenic simian immunodeficiency virus infections. J Virol 2008; 82: 1155–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Wallace M, Scharko AM, Pauza CD et al Functional γδ T‐lymphocyte defect associated with human immunodeficiency virus infections. Mol Med 1997; 3: 60–71. [PMC free article] [PubMed] [Google Scholar]
- 59. Martini F, Urso R, Gioia C et al γδ T‐cell anergy in human immunodeficiency virus‐infected persons with opportunistic infections and recovery after highly active antiretroviral therapy. Immunology 2000; 100: 481–486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Martini F, Poccia F, Goletti D et al Acute human immunodeficiency virus replication causes a rapid and persistent impairment of Vγ9Vδ2 T cells in chronically infected patients undergoing structured treatment interruption. J Infect Dis 2002; 186: 847–850. [DOI] [PubMed] [Google Scholar]
- 61. Cardone M, Ikeda KN, Varano B, Gessani S, Conti L. HIV‐1‐induced impairment of dendritic cell cross talk with γδ T lymphocytes. J Virol 2015; 89: 4798–4808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Sacchi A, Rinaldi A, Tumino N et al HIV infection of monocytes‐derived dendritic cells inhibits Vγ9Vδ2 T cells functions. PLoS One 2014; 9: e111095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Harris LD, Klatt NR, Vinton C et al Mechanisms underlying γδ T‐cell subset perturbations in SIV‐infected Asian rhesus macaques. Blood 2010; 116: 4148–4157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Kosub DA, Durudas A, Lehrman G et al γ/δ T cell mRNA levels decrease at mucosal sites and increase at lymphoid sites following an oral SIV infection of macaques. Curr HIV Res 2008; 6: 520–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Nilssen DE, Muller F, Oktedalen O et al Intraepithelial γ/δ T cells in duodenal mucosa are related to the immune state and survival time in AIDS. J Virol 1996; 70: 3545–3550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Fenoglio D, Poggi A, Catellani S et al Vδ1 T lymphocytes producing IFN‐γ and IL‐17 are expanded in HIV‐1‐infected patients and respond to Candida albicans. Blood 2009; 113: 6611–6618. [DOI] [PubMed] [Google Scholar]
- 67. Sindhu ST, Ahmad R, Morisset R, Ahmad A, Menezes J. Peripheral blood cytotoxic γδ T lymphocytes from patients with human immunodeficiency virus type 1 infection and AIDS lyse uninfected CD4+ T cells, and their cytocidal potential correlates with viral load. J Virol 2003; 77: 1848–1855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Lisovsky I, Isitman G, Song R et al A higher frequency of NKG2A+ than of NKG2A‐ NK cells responds to autologous HIV‐infected CD4 cells irrespective of whether or not they coexpress KIR3DL1. J Virol 2015; 89: 9909–9919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Davis ZB, Cogswell A, Scott H et al A conserved HIV‐1‐derived peptide presented by HLA‐E renders infected T‐cells highly susceptible to attack by NKG2A/CD94‐bearing natural killer cells. PLoS Pathog 2016; 12: e1005421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Ramsuran V, Naranbhai V, Horowitz A et al Elevated HLA‐A expression impairs HIV control through inhibition of NKG2A‐expressing cells. Science 2018; 359: 86–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Negash M, Tsegaye A, Wassie L, Howe R. Phenotypic and functional heterogeneity of peripheral γδ T cells in pulmonary TB and HIV patients in Addis Ababa, Ethiopia. BMC Infect Dis 2018; 18: 464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Casetti R, De Simone G, Sacchi A et al Vγ9Vδ2 T‐cell polyfunctionality is differently modulated in HAART‐treated HIV patients according to CD4 T‐cell count. PLoS One 2015; 10: e0132291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Cummings JS, Cairo C, Armstrong C, Davis CE, Pauza CD. Impacts of HIV infection on Vγ2Vδ2 T cell phenotype and function: a mechanism for reduced tumor immunity in AIDS. J Leukoc Biol 2008; 84: 371–379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Bordon J, Evans PS, Propp N, Davis CE Jr, Redfield RR, Pauza CD. Association between longer duration of HIV‐suppressive therapy and partial recovery of the Vγ2 T cell receptor repertoire. J Infect Dis 2004; 189: 1482–1486. [DOI] [PubMed] [Google Scholar]
- 75. Boudova S, Li H, Sajadi MM, Redfield RR, Pauza CD. Impact of persistent HIV replication on CD4 negative Vγ2Vδ2 T cells. J Infect Dis 2012; 205: 1448–1455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Hebbeler AM, Propp N, Cairo C et al Failure to restore the Vγ2‐Jγ1.2 repertoire in HIV‐infected men receiving highly active antiretroviral therapy (HAART). Clin Immunol 2008; 128: 349–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Nilssen DE, Brandtzaeg P. Intraepithelial γδ T cells remain increased in the duodenum of AIDS patients despite antiretroviral treatment. PLoS One 2012; 7: e29066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Wesch D, Kabelitz D. Differential expression of natural killer receptors on Vδ1 γδ T cells in HIV‐1‐infected individuals. J Acquir Immune Defic Syndr 2003; 33: 420–425. [DOI] [PubMed] [Google Scholar]
- 79. Li Q, Duan L, Estes JD et al Peak SIV replication in resting memory CD4+ T cells depletes gut lamina propria CD4+ T cells. Nature 2005; 434: 1148–1152. [DOI] [PubMed] [Google Scholar]
- 80. Vujkovic‐Cvijin I, Somsouk M. HIV and the gut microbiota: composition, consequences, and avenues for amelioration. Curr HIV/AIDS Rep 2019; 16: 204–213. e‐pub ahead of print Apr 29; 10.1007/s11904-019-00441-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Brenchley JM, Paiardini M, Knox KS et al Differential Th17 CD4 T‐cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood 2008; 112: 2826–2835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Tincati C, Douek DC, Marchetti G. Gut barrier structure, mucosal immunity and intestinal microbiota in the pathogenesis and treatment of HIV infection. AIDS Res Ther 2016; 13: 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Ibarrondo FJ, Wilson SB, Hultin LE et al Preferential depletion of gut CD4‐expressing iNKT cells contributes to systemic immune activation in HIV‐1 infection. Mucosal Immunol 2013; 6: 591–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Poccia F, Cipriani B, Vendetti S et al CD94/NKG2 inhibitory receptor complex modulates both anti‐viral and anti‐tumoral responses of polyclonal phosphoantigen‐reactive V γ 9V δ 2 T lymphocytes. J Immunol 1997; 159: 6009–6017. [PubMed] [Google Scholar]
- 85. Omi K, Shimizu M, Watanabe E et al Inhibition of R5‐tropic HIV type‐1 replication in CD4+ natural killer T cells by γδ T lymphocytes. Immunology 2014; 141: 596–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Poccia F, Battistini L, Cipriani B et al Phosphoantigen‐reactive Vγ9Vδ2 T lymphocytes suppress in vitro human immunodeficiency virus type 1 replication by cell‐released antiviral factors including CC chemokines. J Infect Dis 1999; 180: 858–861. [DOI] [PubMed] [Google Scholar]
- 87. Riedel DJ, Sajadi MM, Armstrong CL et al Natural viral suppressors of HIV‐1 have a unique capacity to maintain γδ T cells. AIDS 2009; 23: 1955–1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Tuero I, Venzon D, Robert‐Guroff M. Mucosal and systemic γδ+ T cells associated with control of simian immunodeficiency virus infection. J Immunol 2016; 197: 4686–4695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Shao L, Zhang W, Zhang S et al Potent immune responses of Ag‐specific Vγ2Vδ2+ T cells and CD8+ T cells associated with latent stage of Mycobacterium tuberculosis coinfection in HIV‐1‐infected humans. AIDS 2008; 22: 2241–2250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Rojas RE, Chervenak KA, Thomas J et al Vδ2+γδ T cell function in Mycobacterium tuberculosis‐ and HIV‐1‐positive patients in the United States and Uganda: application of a whole‐blood assay. J Infect Dis 2005; 192: 1806–1814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Mwale A, Hummel A, Mvaya L et al The B cell, CD8+ T cell and γδ T cell infiltration alters alveolar immune cell homeostasis in HIV‐infected Malawian adults. Wellcome Open Res 2017; 2: 105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Zhou D, Lai X, Shen Y et al Inhibition of adaptive Vγ2Vδ2+ T‐cell responses during active mycobacterial coinfection of simian immunodeficiency virus SIVmac‐infected monkeys. J Virol 2003; 77: 2998–3006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Shen L, Shen Y, Huang D et al Development of Vγ2Vδ2+ T cell responses during active mycobacterial coinfection of simian immunodeficiency virus‐infected macaques requires control of viral infection and immune competence of CD4+ T cells. J Infect Dis 2004; 190: 1438–1447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Shen L, Frencher J, Huang D et al Immunization of Vγ2Vδ2 T cells programs sustained effector memory responses that control tuberculosis in nonhuman primates. Proc Natl Acad Sci USA 2019; 116: 6371–6378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Pitard V, Roumanes D, Lafarge X et al Long‐term expansion of effector/memory Vδ2−γδ T cells is a specific blood signature of CMV infection. Blood 2008; 112: 1317–1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Soderberg‐Naucler C. Treatment of cytomegalovirus infections beyond acute disease to improve human health. Expert Rev Anti Infect Ther 2014; 12: 211–222. [DOI] [PubMed] [Google Scholar]
- 97. Dobmeyer TS, Dobmeyer R, Wesch D, Helm EB, Hoelzer D, Kabelitz D. Reciprocal alterations of Th1/Th2 function in γδ T‐cell subsets of human immunodeficiency virus‐1‐infected patients. Br J Haematol 2002; 118: 282–288. [DOI] [PubMed] [Google Scholar]
- 98. Boullier S, Dadaglio G, Lafeuillade A, Debord T, Gougeon ML. V δ 1 T cells expanded in the blood throughout HIV infection display a cytotoxic activity and are primed for TNF‐α and IFN‐γ production but are not selected in lymph nodes. J Immunol 1997; 159: 3629–3637. [PubMed] [Google Scholar]
- 99. Hudspeth K, Fogli M, Correia DV et al Engagement of NKp30 on Vδ1 T cells induces the production of CCL3, CCL4, and CCL5 and suppresses HIV‐1 replication. Blood 2012; 119: 4013–4016. [DOI] [PubMed] [Google Scholar]
- 100. Rohner E, Wyss N, Heg Z et al HIV and human herpesvirus 8 co‐infection across the globe: systematic review and meta‐analysis. Int J Cancer 2016; 138: 45–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Barcy S, De Rosa SC, Vieira J et al γδ+ T cells involvement in viral immune control of chronic human herpesvirus 8 infection. J Immunol 2008; 180: 3417–3425. [DOI] [PubMed] [Google Scholar]
- 102. Ensoli B, Barillari G, Buonaguro L, Gallo RC. Molecular mechanisms in the pathogenesis of AIDS‐associated Kaposi's sarcoma. Adv Exp Med Biol 1991; 303: 27–38. [DOI] [PubMed] [Google Scholar]
- 103. Uezu K, Kawakami K, Miyagi K et al Accumulation of γδ T cells in the lungs and their regulatory roles in Th1 response and host defense against pulmonary infection with Cryptococcus neoformans . J Immunol 2004; 172: 7629–7634. [DOI] [PubMed] [Google Scholar]
- 104. Wozniak KL, Kolls JK, Wormley FL Jr. Depletion of neutrophils in a protective model of pulmonary cryptococcosis results in increased IL‐17A production by γδ T cells. BMC Immunol 2012; 13: 65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Dantzler KW, Jagannathan P. γδ T cells in antimalarial immunity: new insights into their diverse functions in protection and tolerance. Front Immunol 2018; 9: 2445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Deroost K, Langhorne J. γ/δ T cells and their role in protection against Malaria. Front Immunol 2018; 9: 2973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. He Y, Wu K, Hu Y et al γδ T cell and other immune cells crosstalk in cellular immunity. J Immunol Res 2014; 2014: 960252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Peters C, Kabelitz D, Wesch D. Regulatory functions of γδ T cells. Cell Mol Life Sci 2018; 75: 2125–2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Davey MS, Morgan MP, Liuzzi AR et al Microbe‐specific unconventional T cells induce human neutrophil differentiation into antigen cross‐presenting cells. J Immunol 2014; 193: 3704–3716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Petrasca A, Doherty DG. Human Vδ2+ γδ T cells differentially induce maturation, cytokine production, and alloreactive T cell stimulation by dendritic cells and B cells. Front Immunol 2014; 5: 650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Brandes M, Willimann K, Moser B. Professional antigen‐presentation function by human γδ T Cells. Science 2005; 309: 264–268. [DOI] [PubMed] [Google Scholar]
- 112. Born WK, Huang Y, Reinhardt RL, Huang H, Sun D, O'Brien RL. γδ T cells and B cells. Adv Immunol 2017; 134: 1–45. [DOI] [PubMed] [Google Scholar]
- 113. Caccamo N, Todaro M, La Manna MP, Sireci G, Stassi G, Dieli F. IL‐21 regulates the differentiation of a human γδ T cell subset equipped with B cell helper activity. PLoS One 2012; 7: e41940. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 114. Bansal RR, Mackay CR, Moser B, Eberl M. IL‐21 enhances the potential of human γδ T cells to provide B‐cell help. Eur J Immunol 2012; 42: 110–119. [DOI] [PubMed] [Google Scholar]
- 115. Caccamo N, Battistini L, Bonneville M et al CXCR1 identifies a subset of Vγ9Vδ2 T cells which secrete IL‐4 and IL‐10 and help B cells for antibody production. J Immunol 2006; 177: 5290–5295. [DOI] [PubMed] [Google Scholar]
- 116. Brandes M, Willimann K, Lang AB et al Flexible migration program regulates γδ T‐cell involvement in humoral immunity. Blood 2003; 102: 3693–3701. [DOI] [PubMed] [Google Scholar]
- 117. Murday AS, Chaudhry S, Pauza CD. Interleukin‐18 activates Vγ9Vδ2+ T cells from HIV‐positive individuals: recovering the response to phosphoantigen. Immunology 2017; 151: 385–394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. He X, Liang H, Hong K et al The potential role of CD16+ Vγ2Vδ2 T cell‐mediated antibody‐dependent cell‐mediated cytotoxicity in control of HIV type 1 disease. AIDS Res Hum Retroviruses 2013; 29: 1562–1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Poonia B, Pauza CD. γδ T cells from HIV+ donors can be expanded in vitro by zoledronate/interleukin‐2 to become cytotoxic effectors for antibody‐dependent cellular cytotoxicity. Cytotherapy 2012; 14: 173–181. [DOI] [PubMed] [Google Scholar]
- 120. Garrido C, Clohosey ML, Whitworth CP, Hudgens M, Margolis DM, Soriano‐Sarabia N. γδ T cells: an immunotherapeutic approach for HIV cure strategies. JCI Insight 2018; 3: e120121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Ali Z, Yan L, Plagman N et al γδ T cell immune manipulation during chronic phase of simian‐human immunodeficiency virus infection [corrected] confers immunological benefits. J Immunol 2009; 183: 5407–5417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Poonia B. Adoptive transfer of aminobisphonate‐expanded Vγ9Vδ2+ T cells does not control HIV replication in a humanized mouse model. Immunotherapy 2016; 8: 521–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Betjes MG. Immune cell dysfunction and inflammation in end‐stage renal disease. Nat Rev Nephrol 2013; 9: 255–265. [DOI] [PubMed] [Google Scholar]
- 124. Matsumoto Y, Shinzato T, Takai I et al Peripheral deletion of γδ T cells in haemodialysis patients. Nephrol Dial Transplant 1998; 13: 2861–2866. [DOI] [PubMed] [Google Scholar]
- 125. Juno JA, Waruk JLM, Harris A et al γδ T‐cell function is inhibited in end‐stage renal disease and impacted by latent tuberculosis infection. Kidney Int 2017; 92: 1003–1014. e‐pub ahead of print Jun 24; 10.1016/j.kint.2017.03.036 [DOI] [PubMed] [Google Scholar]
- 126. Zhuang Q, Peng B, Wei W et al The detailed distribution of T cell subpopulations in immune‐stable renal allograft recipients: a single center study. PeerJ 2019; 7: e6417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Par G, Rukavina D, Podack ER et al Decrease in CD3‐negative‐CD8dim+ and Vδ2/Vγ9 TcR+ peripheral blood lymphocyte counts, low perforin expression and the impairment of natural killer cell activity is associated with chronic hepatitis C virus infection. J Hepatol 2002; 37: 514–522. [DOI] [PubMed] [Google Scholar]
- 128. Cimini E, Bonnafous C, Bordoni V et al Interferon‐α improves phosphoantigen‐induced Vγ9Vδ2 T‐cells interferon‐γ production during chronic HCV infection. PLoS One 2012; 7: e37014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Yin W, Tong S, Zhang Q et al Functional dichotomy of Vδ2 γδ T cells in chronic hepatitis C virus infections: role in cytotoxicity but not for IFN‐γ production. Sci Rep 2016; 6: 26296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Chen M, Zhang D, Zhen W et al Characteristics of circulating T cell receptor γ‐δ T cells from individuals chronically infected with hepatitis B virus (HBV): an association between Vδ2 subtype and chronic HBV infection. J Infect Dis 2008; 198: 1643–1650. [DOI] [PubMed] [Google Scholar]
- 131. Wu X, Zhang JY, Huang A et al Decreased Vδ2 γδ T cells associated with liver damage by regulation of Th17 response in patients with chronic hepatitis B. J Infect Dis 2013; 208: 1294–1304. [DOI] [PubMed] [Google Scholar]
- 132. Chen M, Hu P, Peng H et al Enhanced peripheral γδT cells cytotoxicity potential in patients with HBV‐associated acute‐on‐chronic liver failure might contribute to the disease progression. J Clin Immunol 2012; 32: 877–885. [DOI] [PubMed] [Google Scholar]
- 133. Agrati C, Alonzi T, De Santis R et al Activation of Vγ9Vδ2 T cells by non‐peptidic antigens induces the inhibition of subgenomic HCV replication. Int Immunol 2006; 18: 11–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Cimini E, Bonnafous C, Sicard H et al In vivo interferon‐α/ribavirin treatment modulates Vγ9Vδ2 T‐cell function during chronic HCV infection. J Interferon Cytokine Res 2013; 33: 136–141. [DOI] [PubMed] [Google Scholar]
- 135. Fay NS, Larson EC, Jameson JM. Chronic Inflammation and γδ T cells. Front Immunol 2016; 7: 210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Lackey DE, Olefsky JM. Regulation of metabolism by the innate immune system. Nat Rev Endocrinol 2016; 12: 15–28. [DOI] [PubMed] [Google Scholar]
- 137. Donninelli G, Del Corno M, Pierdominici M et al Distinct blood and visceral adipose tissue regulatory T cell and innate lymphocyte profiles characterize obesity and colorectal cancer. Front Immunol 2017; 8: 643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Costanzo AE, Taylor KR, Dutt S, Han PP, Fujioka K, Jameson JM. Obesity impairs γδ T cell homeostasis and antiviral function in humans. PLoS One 2015; 10: e0120918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol 2012; 30: 149–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Shi K, Wang F, Jiang H et al Gut bacterial translocation may aggravate microinflammation in hemodialysis patients. Dig Dis Sci 2014; 59: 2109–2117. [DOI] [PubMed] [Google Scholar]
- 141. Wang F, Jiang H, Shi K, Ren Y, Zhang P, Cheng S. Gut bacterial translocation is associated with microinflammation in end‐stage renal disease patients. Nephrology (Carlton) 2012; 17: 733–738. [DOI] [PubMed] [Google Scholar]
- 142. Anders HJ, Andersen K, Stecher B. The intestinal microbiota, a leaky gut, and abnormal immunity in kidney disease. Kidney Int 2013; 83: 1010–1016. [DOI] [PubMed] [Google Scholar]
- 143. Pan W, Kang Y. Gut microbiota and chronic kidney disease: implications for novel mechanistic insights and therapeutic strategies. Int Urol Nephrol 2018; 50: 289–299. [DOI] [PubMed] [Google Scholar]
- 144. Vaziri ND, Zhao YY, Pahl MV. Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: the nature, mechanisms, consequences and potential treatment. Nephrol Dial Transplant 2016; 31: 737–746. [DOI] [PubMed] [Google Scholar]
- 145. Yang R, Xu Y, Dai Z, Lin X, Wang H. The Immunologic role of gut microbiota in patients with chronic HBV infection. J Immunol Res 2018; 2018: 2361963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Milosevic I, Vujovic A, Barac A et al Gut‐liver axis, gut microbiota, and its modulation in the management of liver diseases: a review of the literature. Int J Mol Sci 2019; 20: E395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Troseid M, Nestvold TK, Rudi K, Thoresen H, Nielsen EW, Lappegard KT. Plasma lipopolysaccharide is closely associated with glycemic control and abdominal obesity: evidence from bariatric surgery. Diabetes Care 2013; 36: 3627–3632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Tuomi K, Logomarsino JV. Bacterial lipopolysaccharide, lipopolysaccharide‐binding protein, and other inflammatory markers in obesity and after bariatric surgery. Metab Syndr Relat Disord 2016; 14: 279–288. [DOI] [PubMed] [Google Scholar]
- 149. Rhoades N, Mendoza N, Jankeel A et al Altered immunity and microbial dysbiosis in aged individuals with long‐term controlled HIV infection. Front Immunol 2019; 10: 463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Belkina AC, Starchenko A, Drake KA et al Multivariate computational analysis of γ δ T cell inhibitory receptor signatures reveals the divergence of healthy and ART‐suppressed HIV+ aging. Front Immunol 2018; 9: 2783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Dunne PJ, Maher CO, Freeley M et al CD3ε expression defines functionally distinct subsets of Vδ1 T cells in patients with human immunodeficiency virus infection. Front Immunol 2018; 9: 940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Benakis C, Brea D, Caballero S et al Commensal microbiota affects ischemic stroke outcome by regulating intestinal γδ T cells. Nat Med 2016; 22: 516–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Bank I, Marcu‐Malina V. Quantitative peripheral blood perturbations of γδ T cells in human disease and their clinical implications. Clin Rev Allergy Immunol 2014; 47: 311–333. [DOI] [PubMed] [Google Scholar]
- 154. Soriano‐Sarabia N, Archin NM, Bateson R et al Peripheral Vγ9Vδ2 T cells are a novel reservoir of latent HIV infection. PLoS Pathog 2015; 11: e1005201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Imlach S, Leen C, Bell JE, Simmonds P. Phenotypic analysis of peripheral blood γδ T lymphocytes and their targeting by human immunodeficiency virus type 1 in vivo . Virology 2003; 305: 415–427. [DOI] [PubMed] [Google Scholar]
- 156. Gurney KB, Yang OO, Wilson SB, Uittenbogaart CH. TCRγδ+ and CD161+ thymocytes express HIV‐1 in the SCID‐hu mouse, potentially contributing to immune dysfunction in HIV infection. J Immunol 2002; 169: 5338–5346. [DOI] [PubMed] [Google Scholar]