

Abstract
Actins play essential roles in cellular morphogenesis. In mice, the T-actin1 and 2 genes, which encode actin-like proteins, are specifically expressed in haploid germ cells. Both T-ACTIN1/ACTLB and T-ACTIN2/ACTL7A have also been cloned and studied. The orthologous genes in humans are present on chromosome 9q31.3 as intronless genes. Defects of germ cell-specific genes can introduce infertility without somatic function impairment. We determined T- ACTIN1 and 2, specifically expressed in the testis using reverse-transcription polymerase chain reaction (RT-PCR). To examine whether genetic polymorphisms of the T-ACTIN1 and 2 genes are associated with male infertility, we screened for T-ACTIN1 and 2 polymorphisms by direct sequencing of DNA from 282 sterile and 89 fertile Japanese men. We identified five and six single nucleotide polymorphisms (SNPs) in the T-ACTIN1 and 2 regions of the sterile and fertile subjects respectively. Among these genetic polymorphisms was a novel SNP that was not in the National Center for Biotechnology Information SNP database. Although we could not determine whether these SNPs cause infertility, the prevalence of these genetic polymorphisms may be useful for analyzing polymorphisms in future large- scale genetic analyses.
Keywords: Germ Cell, Infertility, Single Nucleotide Polymorphism, Sperm, Testis
After meiosis, round spermatids undergo a dramatic change to develop the specific morphology of the mature sperm. Actin proteins play important functions in this process (1). We developed a mouse subtracted library including genes specifically expressed in spermatogenesis and cloned and characterized these genes (2). Among these genes were T-actin1 and 2, which encode actin-like proteins and are specifically expressed in haploid germ cells. T-actin1 is located in the cytoplasm while T-actin2 is localized in the nuclei of testicular haploid germ cells and is present only in the heads and tails of sperm (3). In both the mouse and human genome, T-ACTIN1 and 2 are positioned head-to-head and lack introns (4, 5). The resulting amino acid sequences, genomic construction, and cAMP response elements (CRE) consensus DNA sequence of the promoters of T-actin1 and 2 are conserved in mice (4). These genes have been reported to cause infertility by inducing autoimmunity to sperm (6). Human T-ACTINs may play important roles in the specific morphogenesis of spermatozoa during spermiogenesis, as well as in sperm function.
We investigated genetic polymorphisms in the DNA sequences of germ cell-specific genes in infertile male patients and male volunteers with confirmed fertility (7- 19) to identify polymorphisms potentially linked to male infertility (7, 8, 15, 19). In this study, we report our analysis of genetic polymorphisms in T-ACTIN1/ACTLB and T-ACTIN2/ACTL7A in Japanese men.
Defects in germ cell-specific genes may be a cause of idiopathic infertility. To detect the presence of small amounts of transcripts, we examined tissue-specific expression patterns of T-ACTIN1 and 2 by reverse-transcription polymerase chain reaction (RT-PCR) using cDNA from various organs and a Rapid-Scan gene expression panel containing cDNA from different human tissues (OriGene Technologies, Rockville, MD, USA) (20). The specific primers:
-
TACT1-RTF:
5'-ATGGCGACAAGGAACAGCCCCATG-3'
-
TACT1-RTR:
5'-TCAGCACTTGCTGTAGATGGCCAC-3'
for T-ACTIN1
-
TACT2-RTF:
5'-ATGTGGGCTCCACCAGCAGCAATC-3'
-
TACT2-RTR:
5'-TCAGAAGCACCTTCTGTAGAGGAAG-3'
for T-ACTIN2
were designed to amplify fragments from the open reading frames. Polymerase chain reaction (PCR) was performed using Gflex Hot Start (Takara, Japan). The cycling conditions were 96°C for 2 minutes, followed by 35 cycles of denaturation at 96°C for 45 seconds, annealing at 58°C for 45 seconds, and extension at 68°C for 90 seconds. As a control, ß-actin was also amplified using primers:
-
ACTBF:
5'-ACCGAGGCCCCCCTGAACCC-3'
-
ACTBR:
5'-TCCATCATGAAGTGTGACGT-3'
according to the manufacturer’s protocol. T-ACTINs were specifically detected only in the testis (Fig .1).
Fig 1.
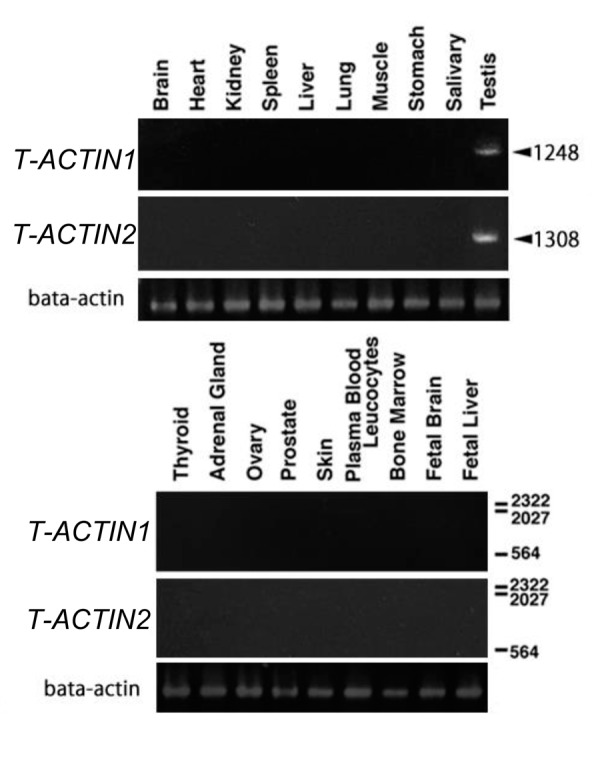
mRNA expression of T-ACTIN1 and 2 in various human organs. Multiple human tissue cDNAs were subjected to polymerase chain reaction analysis. Fragments of T-ACTIN1 and 2 were specifically detected in the testes. Numbers in the right-hand margin indicate the lengths of the amplified fragments and DNA ladder makers. The expression of actin mRNA was also examined as a control.
The entire coding sequences of T-ACTIN1 and 2 (National Center for Biotechnology Information [NCBI] accession number: chromosome 9, NC_000009.12 (108862228..108863755), Fig.2) are intronless, similar to mouse T-actin1 and 2. As T-ACTINs are expressed at high levels in the human testis (Fig .1), we investigated whether genetic polymorphisms in T-ACTINs are associated with male infertility.
Infertile Japanese subjects (n=282) were divided into subgroups according to the degree of defective spermatogenesis: 192 patients (68%) had non-obstructive azoospermia, and 90 (32%) had severe oligospermia (<5×106 cells/mL), according to the criteria of the World Health Organization (Table 1). All patients had idiopathic infertility based on cytogenetic analysis and no history of other medical conditions, including cryptorchidism, recurrent infections, trauma, orchitis, varicocele, and others. The control group consisted of fertile males who had fathered children born at a maternity clinic (n=89). All donors were informed of the purpose of the study and gave permission for use of their blood for genomic DNA data. This study was approved by the institutional review board and independent ethics committee of Osaka University.
Table 1.
Backgrounds of 371 Japanese men
| Status | n (%) |
|---|---|
| Azoospermia | 192 (68) |
| Severe oligospermia | 90 (32) |
| Total infertile | 282 (100) |
| Fertile control | 89 |
Genomic DNA was isolated from blood samples by protease treatment and phenol extraction. T-ACTIN1 and 2 sequences were amplified through PCR using the following primers:
-
TACT1F:
5'-GTGGATCCCTGGATGGTCCGCTGTGCGG- 3'
-
TACT1R:
5'-GGCCTGTGCCATCTGTGCTGGAGG-3'
for T-ACTIN1,
TACT2F: 5'-CTTTCAGGCCTTGAATCCAGTGGG-3'
TACT2R: 5'-GGTAGGCACTGCCAGTGCAGTGTC-3' for T-ACTIN 2 (Fig .2).
Fig 2.

Schematic view of the T-ACTIN1 and 2 genes. The T-ACTIN1 and 2 intronless genes are located on chromosome 9 (NCBI accession number: NC_000009.12). The box indicates the transcribed region of the T-ACTIN1 and 2 genes. The open reading frame is shaded. T-ACTIN1 is transcribed to the right and T-ACTIN2 to the left. The small horizontal arrows in the box indicate the locations of the polymerase chain reaction (PCR) and DNA-sequencing primers. The arrowheads indicate the positions of genetic polymorphisms. The NCBI accession numbers of T-ACTIN1 and 2 are CCDS4295.1 and CCDS6772.1, respectively.
PCR was performed using Ex Taq Hot Start (Takara, Japan) and consisted of 40 cycles of 96ºC for 45 seconds, 65ºC for 45 seconds, and 72ºC for 90 seconds. PCR-amplified fragments were purified using SUPREC PCR spin columns (Takara). The resulting DNA fragments were sequenced independently from both ends by the same PCR protocol using thermal cycle sequencing kits (Applied Biosystems, Foster City, CA, USA). Internal primers:
-
TACT1F2:
5'-GCCTGTGCCATCTGTGCTGG-3'
-
TACT2F2:
5'-TCTCAAGCTGGTTAACCCTCTGCG-3'
-
TACT2R2:
5'-AGGCACTGCCAGTGCAGTGT-3'
were used to confirm T-ACTIN genes with ambiguous identifications. The reaction products were analyzed using an ABI-PRISM 310 Genetic Analyzer (Applied Biosystems). Differences in variables between the experimental and control conditions were compared using Fisher’s exact test (P<0.05).
Nucleic acid base exchanges introducing one nonsense mutation and four silent mutations were found in the T-ACTIN1 open reading frame (Table 2). Single nucleotide polymorphisms (SNPs) were found in three silent mutations (48C>T, 561C>T, 870C>T) as minor genotypes in the entire Japanese cohort. The minor 1137 C>T homozygous alleles on T-ACTIN1 was not detected in the infertile group. One nonsense mutation was found in the volunteer group. The translated region of T-ACTIN1 is 1248 bp long, and the nonsense mutation appears at base pair 1,171, near the C-terminus. This mutation thus has little influence on the function of the translated protein, making it unlikely to be a cause of infertility.
Table 2.
Prevalence of single nucleotide polymorphisms (SNPs) in T-ACTIN1 in infertile or proven fertile populations
| Position | Genotype | Number (%) of SNP | Reference | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Nucleotide* | Amino acid | Infertile (%) | Proven fertile (%) | (NCBI dbSNP rs#) | ||||||
| T-ACTIN1/ACTL7B | 48 | 16 | D | C/C | 161 (57.1) | 54 (60.7) | rs3750468 | |||
| C/T | 102 (36.2) | 28 (31.5) | ||||||||
| T/T | 19 (6.7) | 7 (7.9) | P=0.74 | |||||||
| 561 | 187 | Y | C/C | 161 (57.1) | 54 (60.7) | rs11543179 | ||||
| C/T | 102 (36.2) | 28 (31.5) | ||||||||
| T/T | 19 (6.7) | 7 (7.9) | P=0.74 | |||||||
| 870 | 290 | T | C/C | 218 (77.7) | 66 (74.2) | rs3750467 | ||||
| C/T | 62 (22.0) | 21 (23.6) | ||||||||
| T/T | 2 (0.7) | 2 (2.2) | P=0.23 | |||||||
| 1137 | 379 | S | C/C | 282 (100) | 87 (97.8) | rs769443334 | ||||
| C/T | 0 (0) | 2 (2.2) | ||||||||
| T/T | 0 (0) | 0 (0) | ||||||||
| 1171 | 391 | Q | C/C | 282 (100) | 88 (98.9) | rs750564969 | ||||
| Q/Ter | C/T | 0 (0) | 1 (1.1) | |||||||
| Ter | T/T | 0 (0) | 0 (0) | |||||||
| Total | 282 | 89 | ||||||||
D; Aspartate, Y; Tyrosine, T; Threonine,S; Serine, Q; Glutamine, Ter; Termination, and *; The nucleotide positions relative to the first methionine.
Two nucleic acid base exchanges introducing amino acid substitutions and four silent mutations were found in the T-ACTIN2 open reading frame (Table 3). The frequency of minor genotypes associated with T-ACTIN2 nucleotide polymorphisms was low in Japanese males. One silent mutation, 153A>G, in T-ACTIN2 was not registered in the NCBI SNP database (dbSNP), marking a novel discovery in our Japanese cohort.
Table 3.
Prevalence of single nucleotide polymorphisms (SNPs) in T-ACTIN1 in infertile or proven fertile populations
| Position | Genotype | Number (%) of SNP | Reference | ||||
|---|---|---|---|---|---|---|---|
| Nucleotide* | Amino acid | Infertile (%) | Proven fertile (%) | (NCBI dbSNP rs#) | |||
| T-ACTIN1/ACTL7B | 118 | 40 | R | C/C | 28 (99.6) | 89 (100) | rs201549336 |
| C/A | 1 (0.4) | 0 (0) | |||||
| A/A | 0 (0) | 0 (0) | |||||
| 133 | 45 | R | C/C | 282 (100) | 88 (98.9) | rs368653764 | |
| R/C | C/T | 0 (0) | 1 (1.1) | ||||
| C | T/T | 0 (0) | 0 (0) | ||||
| 153 | 51 | P | A/A | 218 (99.6) | 89 (100) | In present study | |
| A/ | 1 (0.4) | 0 (0) | |||||
| G/G | 0 (0) | 0 (0) | |||||
| 528 | 176 | P | A/A | 278 (98.6) | 88 (98.9) | rs3739692 | |
| A/T | 4 (4.1) | 1 (1.1) | |||||
| T/T | 0 (0) | 0 (0) | |||||
| 657 | 219 | V | G/G | 261 (92.6) | 82 (92.1) | rs3739693 | |
| G/A | 21 (7.4) | 4 (4.5) | |||||
| AA | 0 (0) | 3 (3.4) | |||||
| 1018 | 340 | V | G/G | 261 (92.6) | 82 (92.1) | rs7872077 | |
| V/M | G/A | 21 (7.4) | 4 (4.5) | ||||
| M | A/A | 0 (0) | 3 (3.4) | ||||
| Total | 282 | 89 | |||||
R; Arginine, C; Cysteine, P; Proline, V; Proline, M; Methionine, and *; The nucleotide positions relative to the first methionine.
Logistic regression modeling of the prevalence of haplotypes, including SNPs, revealed no significant differences between major and minor alleles lacking the 1,018 G>A on T-ACTIN2 SNP in males proven to be fertile. The minor 1,018 G>A homozygous alleles on T-ACTIN2 in males proven to be fertile is considered to be due to an error made by the sequencer.
The appearance of 48 C>T and 561 C>T in T-ACTIN1 was linked; as was the appearance of 657 G>A and 1,018 G>A in T-ACTIN2. Thus, the SNPs in these two genes may have the same origin.
Although many SNPs have been registered in the NCBI dbSNP, we detected only 11 genetic polymorphisms in the open reading frames of the T-ACTIN genes among 371 Japanese men. Finally, z χ2-test was used to compare genotype distributions between infertile males and proven fertile controls. There were no significant differences for the minor genotypes (P>0.05).
Our research group has focused on cloning and analyzing germ cell-specific genes. Chromosome mapping of these genes revealed that they are distributed across various chromosomes, and that many are intronless (21). T-ACTINs are among these intronless genes and are specifically expressed in the testis (Fig .2). The dysfunction of germ cell-specific genes does not affect ontogeny and may be a cause of unexplained male infertility. The dysfunction of these genes in mice has been shown to lead to infertility (22). Dominant-negative gene mutations are not passed on to the next generation, however other gene mutations can be inherited from a heterozygous male parent or from the female parent. More than 20% of married couples in Japan are affected by infertility and the male partner is responsible in two-thirds of these cases (23). We undertook an extensive analysis of genetic polymorphisms in germ cell-specific genes and of the relationship between gene polymorphisms and infertility (7-19). We found potential relationships between infertility in Japanese men and genetic polymorphisms or mutations in PRM2, TP1, PGAM4, and SCOT-T (7, 8, 15, 19). We analyzed SNPs in germ cell-specific genes and found that some included genetic polymorphisms with single amino acid substitutions, whereas other specific genes had few genetic polymorphisms. Most genes having several genetic polymorphisms encoded in proteins were involved in signal transduction or regulation, whereas those with few genetic polymorphisms were more likely to encode structural proteins (12). In this study, we discovered several different SNPs in T-ACTIN1 and 2 in a cohort of Japanese men. The similar frequencies of these polymorphisms between the fertile and infertile groups in this study imply that these mutations are not associated with male infertility. However, the prevalence data for these genetic polymorphisms might be useful when analyzing the association of traits and genetic polymorphisms in further large-scale genetic analyses.
Acknowledgments
We thank Ms. H. Nishimuna, Mr. H. Kawaguchi, and Ms. M. Kanzaki for their technical assistance with the sequence analysis of human DNA samples. The funding sources played no role in the study design; the collection, analyses, or interpretation of data; or in the writing or publication of the manuscript. The authors declare no conflict of interest.
Author’s Contributions
H.T.; Contributed to conception and design, all experimental work, data and statistical analysis, and interpretation of data, and wrote the manuscript. Y.M., A.T., M.W.; Contributed to materials and analyzed the data. All authors read and approved the final manuscript.
References
- 1.Soda T, Miyagawa Y, Ueda N, Takezawa K, Okuda H, Fukuhara S, et al. Systematic characterization of human testis-specific actin capping protein β3 as a possible biomarker for male infertility. Hum Reprod. 2017;32(3):514–522. doi: 10.1093/humrep/dew353. [DOI] [PubMed] [Google Scholar]
- 2.Fujii T, Tamura K, Masai K, Tanaka H, Nishimune Y, Nojima H. Use of stepwise subtraction to comprehensively isolate mouse genes whose transcription is up-regulated during spermiogenesis. EMBO Rep. 2002;3(4):367–372. doi: 10.1093/embo-reports/kvf073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tanaka H, Iguchi N, Egydio de Carvalho C, Tadokoro Y, Yomogida K, Nishimune Y. Novel actin-like proteins T-ACTIN 1 and T-ACTIN 2 are differentially expressed in the cytoplasm and nucleus of mouse haploid germ cells. Biol Reprod. 2003;69(2):475–482. doi: 10.1095/biolreprod.103.015867. [DOI] [PubMed] [Google Scholar]
- 4.Hisano M, Yamada S, Tanaka H, Nishimune Y, Nozaki M. Genomic structure and promoter activity of the testis haploid germ cell-specific intronless genes, Tact1 and Tact2. Mol Reprod Dev. 2003;65(2):148–156. doi: 10.1002/mrd.10276. [DOI] [PubMed] [Google Scholar]
- 5.Chadwick BP, Mull J, Helbling LA, Gill S, Leyne M, Robbins CM, et al. Cloning, mapping, and expression of two novel actin genes, actin-like-7A (ACTL7A) and actin-like-7B (ACTL7B), from the familial dysautonomia candidate region on 9q31. Genomics. 1999;58(3):302–309. doi: 10.1006/geno.1999.5848. [DOI] [PubMed] [Google Scholar]
- 6.Fu J, Yao R, Luo Y, Yang D, Cao Y, Qiu Y, et al. Immune Infertility Should Be Positively Diagnosed Using an Accurate Method by Monitoring the Level of Anti-ACTL7a Antibody. Sci Rep. 2016;6:22844–22844. doi: 10.1038/srep22844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tanaka H, Miyagawa Y, Tsujimura A, Matsumiya K, Okuyama A, Nishimune Y. Single nucleotide polymorphisms in the protamine-1 and -2 genes of fertile and infertile human male populations. Mol Hum Reprod. 2003;9(2):69–73. doi: 10.1093/molehr/gag010. [DOI] [PubMed] [Google Scholar]
- 8.Miyagawa Y, Nishimura H, Tsujimura A, Matsuoka Y, Matsumiya K, Okuyama A, et al. Single-nucleotide polymorphisms and mutation analyses in the TNP1 and TNP2 genes of fertile and infertile human male populations. J Androl. 2005;26(6):779–786. doi: 10.2164/jandrol.05069. [DOI] [PubMed] [Google Scholar]
- 9.Nishimune Y, Tanaka H. Infertility caused by polymorphisms or mutations in spermatogenesis-specific genes. J Androl. 2006;27(3):326–334. doi: 10.2164/jandrol.05162. [DOI] [PubMed] [Google Scholar]
- 10.Tanaka H, Matsuoka Y, Onishi Y, Kitamura K, Miyagawa Y, Nishimura H, et al. Expression profiles and single-nucleotide polymorphism analysis of human HANP1/H1T2 encoding a histone H1-like protein. Int J Androl. 2006;29(2):353–359. doi: 10.1111/j.1365-2605.2005.00600.x. [DOI] [PubMed] [Google Scholar]
- 11.Tanaka H, Hirose M, Tokuhiro K, Matsuoka Y, Miyagawa Y, Tsujimura A, et al. Single nucleotide polymorphisms: discovery of the genetic causes of male infertility. Soc Reprod Fertil Suppl. 2007;65:531–534. [PubMed] [Google Scholar]
- 12.Tokuhiro K, Hirose M, Miyagawa Y, Tsujimura A, Irie S, Isotani A, et al. Meichroacidin containing membrane occupation and nexus motif is essential for spermatozoa morphogenesis. J Biol Chem. 2008;283(27):19039–19048. doi: 10.1074/jbc.M708590200. [DOI] [PubMed] [Google Scholar]
- 13.Irie S, Tsujimura A, Miyagawa Y, Ueda T, Matsuoka Y, Matsui Y, et al. Single nucleotide polymorphisms in PRDM9 (MEISETZ) in patients with nonobstructive azoospermia. J Androl. 2009;30(4):426–431. doi: 10.2164/jandrol.108.006262. [DOI] [PubMed] [Google Scholar]
- 14.Irie S, Nakamura J, Miyagawa Y, Tsujimura A, Okuda H, Yamamoto K, et al. Primary screening of single nucleotide polymorphisms in human calreticulin 3 (CALR3) The Open Andrology Journal. 2010;3:30–35. [Google Scholar]
- 15.Okuda H, Tsujimura A, Irie S, Yamamoto K, Fukuhara S, Matsuoka Y, et al. A single nucleotide polymorphism within the novel sex-linked testis-specific retrotransposed PGAM4 gene influences human male fertility. PLoS One. 2012;7(5):e35195–e35195. doi: 10.1371/journal.pone.0035195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tanaka H, Tsujimura A, Miyagawa Y, Kohama Y, Aramaki Y, Araki Y, et al. Nucleotide Polymorphism Analysis of Testis-specific CETN1 in Human Male Infertility. The Open Andrology Journal. 2013;5:1–5. [Google Scholar]
- 17.Tanaka H, Tsujimura A, Miyagawa Y, Wada M. Genetic Variation in the testis-specific Poly(A) Polymerase Beta (PAPOLB) gene among japanese males. The Open Reproductive Science Journal. 2015;7:1–4. [Google Scholar]
- 18.Tanaka H, Tsujimura A, Miyagwa Y, Oh D, Choi D, Wada M, et al. Genetic variation in the testis-specific HASPIN gene encoding a serine/threonine protein kinase in infertile Japanese males. Advances in Sexual Medicine. 2016;6(3):19–25. [Google Scholar]
- 19.Tanaka H, Miyagwa Y, Tsujimura A, Nishimune Y. Male infertility and single-nucleotide polymorphisms in the testis-specific succinyl CoA: 3-oxoacid CoA transferase (SCOT-T/OXCT2) gene in a Japanese cohort. Int J Reprod Fertil Sex Health. 2018;001(S1: 02):1–6. [Google Scholar]
- 20.Kitamura K, Nishimura H, Nishimune Y, Tanaka H. Identification of human HAPRIN potentially involved in the acrosome reaction. J Androl. 2005;26(4):511–518. doi: 10.2164/jandrol.04189. [DOI] [PubMed] [Google Scholar]
- 21.Tanaka H, Baba Y. Gene Expression in spermiogenesis. Cell Mol Life Sci. 2005;62(3):344–354. doi: 10.1007/s00018-004-4394-y. [DOI] [PubMed] [Google Scholar]
- 22.Matzuk MM, Lamb DJ. The biology of infertility: research advances and clinical challenges. Nat Med. 2008;14(11):1197–1213. doi: 10.1038/nm.f.1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Barratt CLR, Bjorndahl L, De Jonge CJ, Lamb DJ, Osorio Martini F, McLachlan R, et al. The diagnosis of male infertility: an analysis of the evidence to support the development of global WHO guidance-challenges and future research opportunities. Hum Reprod Update. 2017;23(6):660–680. doi: 10.1093/humupd/dmx021. [DOI] [PMC free article] [PubMed] [Google Scholar]