

Abstract
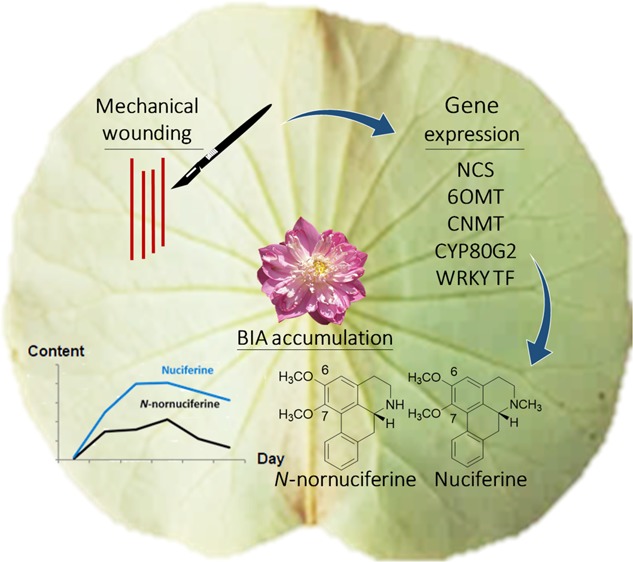
Lotus (Nelumbo nucifera Gaertn.) contains various bioactive compounds, with benzylisoquinoline alkaloids (BIAs) as one of the major groups. The biosynthetic pathways of two major bioactive BIAs in this plant, nuciferine and N-nornuciferine, are still not clear. Therefore, several genes related to BIA biosynthesis were searched from the lotus database to study the role of key genes in regulating these pathways. In this study, the expression profiles of NCS, CNMT, 6OMT, CYP80G2, and WRKY TFs were investigated in mechanically wounded lotus leaves. It was found that the accumulation of nuciferine and N-nornuciferine significantly increased in the mechanically wounded lotus leaves in accordance with the relative expression of putative CYP80G2 and one WRKY transcription factor (NNU_24385), with the coregulation of CNMT. Furthermore, the role of methyltransferase-related genes in this study suggested that methylation of the isoquinoline nucleus to yield a methylated-BIA structure may occur at the N position before the O position. Altogether, this study provides improved understanding of the genes regulating BIA biosynthesis under stressed conditions, which could lead to improvements in BIA production from the commercial lotus.
Introduction
The content of benzylisoquinoline alkaloids (BIAs) in plants is known to be closely related to their pharmaceutical values, such as analgesics, morphine, and codeine from Papaver somniferum, berberine from Coptis japonica, and the antibacterial agent sanguinarine from P. somniferum and Eschscholzia californica. Similarly, lotus (Nelumbo nucifera Gaertn.), which contains BIAs, has also been reported to possess interesting biological activities. For example, nuciferine and N-nornuciferine from lotus provide antidiabetic agents, anti-HIV agents, and melanogenesis inhibitory activities.1−3 An aquatic perennial lotus plant is distributed throughout Asia and Oceania and is used as a raw material in folk medicine.4 Phytochemically, N. nucifera has been shown to contain several bioactive alkaloids, with the majority being two aporphine-type BIAs in the leaves: N-nornuciferine and nuciferine.1,2,5 Other BIAs, such as lotusine, liensinine, isoliensinine, and neferine, have been found in trace amounts in the leaf sap, seed, embryo, and tender rhizome.6 Biosynthetically, the pathways leading to the formation of many BIAs in various alkaloid-producing plant species have been successfully elucidated, with identification of the corresponding genes for the purpose of pharmaceutical usage; however, little is known about the biosynthetic pathways of aporphine-type BIAs in plants, including bioactive nuciferine and N-nornuciferine in lotus.5,7,8 Recently, two aporphine-type BIAs, corytuberine from C. japonica and magnoflorine from Magnolia grandiflora, have had their possible biosynthetic pathways documented in C. japonica and P. somniferum, respectively.9,10 CYP80-based reactions have been employed in many steps of simple BIA production.8−11 In this pathway, CYP80G2 is responsible for hydroxylation and phenol C–C coupling. It has been reported that CYP80G2 catalyzes the phenol C–C coupling reaction, by which (S)-reticuline is converted to (S)-corytuberine.10−12 The presence of aporphine-type BIAs in lotus suggests that it possibly possesses a CYP80G2 analog that catalyzes the formation of the aporphine backbone.10,13 In this study, we predicted that this putative CYP80G2 sequence would be involved in this reaction. The putative CYP80G2 was then searched for in the lotus database.
In the literature, apart from their chemical structures, nuciferine and N-nornuciferine biosynthetic pathways have not yet been reported. It would be interesting and useful for more attention to be given to their biosynthetic pathways. Thus, we gathered this information, along with the reported structure of various aporphine-type BIAs from lotus organs, to draw a proposed biosynthetic pathway of nuciferine and N-nornuciferine (Figure 1). Generally, the BIA biosynthetic pathway begins with the condensation of dopamine and 4-hydroxyphenyl acetaldehyde catalyzed by norcoclaurine synthase (NCS). Then, a series of subsequent enzymes, including S-adenosyl-l-methionine-dependent methyltransferases 6-O-methyltransferase (6OMT), 7-O-methyltransferase (7OMT), coclaurine-N-methyltransferase (CNMT), and P450 C–C phenol-coupling enzyme (corytuberine synthase (CTS) or CYP80G2), probably perform catalytic activity to yield nuciferine and N-nornuciferine structures.10,11,14
Figure 1.

Proposed biosynthetic pathway of nuciferine and N-nornuciferine. Target genes and products examined in this study are highlighted in red. (A) Possible biosynthetic pathway of BIAs in lotus mapped with various related enzymes and reported BIA structures found in lotus. (B) Possible pathway to nuciferine and N-nornuciferine synthesis in lotus drawn following relative gene expression patterns from the mechanical wounding experiment where CYP80G2 showed the first response to wounding, followed by CNMT and 6OM.
In addition, BIA-related genes are regulated by specific WRKY transcription factors (WRKY TFs), which have multiple functions in response to biotic and abiotic stresses, such as pathogens, drought, and wounds.15,16 There is evidence suggesting that wound-induced WRKY TF1 could subsequently regulate BIA pathways in P. somniferum.17 Similarly, WRKY TF1 from C. japonica has been found to induce the biosynthetic gene expression of berberine, which results in significant increases in berberine production in its tissues.15,18 In this study, we isolated and investigated the expressions of some biosynthetic genes, including NCS, CNMT, 6OMT, and CYP80G2, and the WRKY family of TFs in response to a mechanical wounding stress, along with quantitative analyses of nuciferine and N-nornuciferine accumulation in lotus leaves. Our findings could provide information on the regulation of key enzymes in BIA biosynthetic pathways and be helpful for developing lotus varieties with high levels of bioactive BIAs.
Results
Isolation of Genes Encoding BIA Biosynthetic Enzymes and Transcription Factor
Most of the BIA-related biosynthetic enzymes in lotus, except NCS, are predicted to be present. Recently, five isoforms of NCS have been isolated from the lotus genome and found to be related to the total content of alkaloid in lotus organs,19 and therefore these were used in this study. Results from amino acid sequence alignment indicated that the five identified NnNCS shared a conserved region in the glycine-rich loop (GDTGVGT), which was also shared with known NCS from other plant species retrieved from GenBank (Figure 2A). Searching for candidate 6OMT genes in the lotus genome led to the finding of four genes closely related to the 6OMT of C. japonica (accession no. BAB08004) (Figure S3). They were isolated from template sequences of NNU_19035, NNU_03165, NNU_03166, and NNU_23168 presented in the lotus genome and were hence named as NnOMT35, NnOMT65, NnOMT66, and NnOMT68, respectively. They shared 61, 63, 67, and 69% identity in amino acid sequence, respectively, with Cj6OMT. Sequence analysis suggested that they contained 1056, 1044, 1041, and 1041 base pairs, which encode 351, 347, 346, and 346 theoretical amino acids with deduced isoelectric points (pI) at pH of 3.53, 5.85, 5.26, and 6.07, respectively.
Figure 2.

Deduced amino acid sequence alignment of NCS, 2 methyltransferases, CYP80G2, and WRKY TFs from lotus (N. nucifera) with sequences retrieved from the GenBank database. (A) Five targeted NCS in bold present a glycine-rich loop, the ligand binding domain of the Bet V1 protein family. (B–C) Sequence motifs A, B, and C of plant S-adenosyl-l-methionine-dependent methyltransferases (SAM) present in targeted CNMT (NnCNMT) and 6OMT (Nn6OMT); NnCNMT possesses Motif A alone. (D) Putative NnCYP80Gs present the conserved eukaryotic P450 regions consisting of a helical K region, an aromatic region, and a heme-binding region. The asterisk located before a heme-binding region indicates the position of the proline of the CYP80G2 substitute by alanine/glycine. (E) A total of 10 putative WRKY TFs possess the β-sheet WRKY DNA binding domains (DBDs), which are located by the line. The other highlighted areas present the zinc-finger motif. Asterisks in all highlighted regions indicate conserved amino acid residues.
Three putative NnCNMT sequences were isolated using NNU_11880 as template and named as follows: NnCNMT1, NnCNMT2, and NnCNMT3. They shared 51% amino acid sequence identity with CjCNMT (accession no. BAB71802) and contained 1074, 1388, and 1224 base pairs, respectively (Figure S3). A stop codon influenced by alternative splicing was found in NnCNMT2, resulting in premature protein termination; therefore, NnCNMT2 was not used for gene expression analysis. NnCNMT1 and NnCNMT3 encoded 358 and 408 theoretical amino acids with a deduced pI at pH of 7.03 and 7.55, respectively.
The coding sequences of isolated NnOMT35, NnOMT65, NnOMT66, NnOMT68, NnCNMT1, and NnCNMT3 were deposited in GenBank with accession nos MG517490, MG517491, MG517492, MG517493, MG517494, and MG517495, respectively. Amino acid sequence alignment of all isolated 6OMT presented a conserved region as reported in Cj6OMT, indicated by the motifs A, B, and C of a conserved S-adenosylmethionine (SAM)-binding domain located at the C-terminal end containing three conserved glycine residues (Figure 2B). However, the isolated CNMT gene possessed motif A alone (Figure 2C).20,21
One putative CYP80G2 gene from this study was named NnCYP80G2. It contained 1,458 base pairs that encode 519 theoretical amino acids, with a deduced pI at a pH of 5.70, and possessed conserved eukaryotic P450 regions consisting of a helix K region, an aromatic region, and a heme-binding region at the C-terminal end (Figure 2D). It shared over 80% amino acid similarity to known CYP80G2. The highest similarity (%) was to corytuberine synthase (CTS) from Podophyllum peltatum (accession no. AJD20230).10,11
For the WRKY transcript factors, all selected WRKY TFs presented the WRKY TF conserved region, as shown in Figure 2E, in which the β-sheet WRKY DNA binding domain (DBD) is highlighted. WRKYGQK and WRKYGKK, which are primary motifs of the DNA binding domain, were present in eight sequences.15,17,22 The other two putative WRKY TFs isolated from two identified regions (NNU_11881RA and NNU_05136RA) were found to possess an unlikely WKKY anomaly of the primary WRKYGKK motif of DBD.15
Increased BIA Content and Related Gene Expression in Mechanically Wounded (W) Lotus Leaves
Plants have evolved defense mechanisms to respond to wounds and pathogen infection by releasing endogenous molecules from wounded areas. Consequently, damage-associated molecular patterns (DAMPs) activate the plant innate immune system and the expression of defense-related genes.17 Defense responses mediated by wounding are similar to those mediated by DAMPs or microbe-associated molecular patterns. Thus, mechanical wounding has become a powerful method used to study cellular activity in plants.23 In this study, four structural genes were found to display quite different expression patterns during observation after wounding. CYP80G2 apparently responded first to mechanical wounding, with an over 10-fold increase in its relative expression level on day 0. NCS and CNMT slightly increased on day 2, while 6OMT expressed a low and late response. It is worth noting that CNMT and CYP80G2 shared parallel expression patterns during days 2–7 (Figure 3A).
Figure 3.

Quantitative real-time polymerase chain reaction (qRT-PCR) and high-performance liquid chromatography (HPLC) analyses of wounded lotus leaves collected on various observation days. (A) Bar graph shows the relative expression of putative norcoclaurine synthase (NCS), norcoclaurine 6-O-methyltransferase (6OMT), coclaurine-N-methyltransferase (CNMT), and CYP80G2 in wounded lotus leaves. (B) Bar graph shows the relative expression of five putative WRKY TFs. (C) Sum of nuciferine and N-nornuciferine contents in the wounded lotus leaves. Error bars show the standard deviation (SD) of the mean. The different lowercase letters indicate significant differences at p < 0.05.
Apart from the structural genes, WRKY TFs also showed promising behavior in this mechanism. Regarding the predominantly detectable signal from qRT-PCR analysis, five putative WRKY TFs (NNU_09891, NNU_02028, NNU_05834, NNU_24385, and NNU_12194) were selected for this study. On day 0, their relative expression levels in the wounded leaves were elevated by over 10-fold compared to the control group. Two putative WRKY TFs (NNU_09891 and NNU_24385) were recognized to have the greatest response, while other candidates displayed small changes throughout a week of observations (Figure 3B). Overall, the order of BIA-related gene expression after wounding was as follows: WRKY TFs (NNU_24385), CYP80G2, NCS, CNMT, and 6OMT.
Based on these results, CYP80G2 and WRKY TFs (NNU_24385) were deemed to be the most sensitive wound-responding genes (Figure 3A,B). However, NCS and CNMT probably played an important role on day 2 because they increased in accordance with the significant increase of the total BIA content (Figure 3C). The expression of both genes was correlated well with the accumulation of nuciferine, which appeared to be about 3 times higher than that of N-nornuciferine. For the two WRKY TFs, the fluctuation in their expression patterns between days 0 and 3 in the wounded leaves suggested that there was a limitation in the BIA production. Consequently, analysis of the events in the neighboring, nonwounded (NW) tissues was conducted to better understand these results.
Wounding Effects on Gene Expression and Bioactive Compound Accumulation in Neighboring Tissues
Wound responses are known to occur in both the wounded leaves (local response) and nonwounded leaves (systematic response), which are generated by hormones such as jasmonic acid or typical signaling of wounding, such as cell wall-derived oligogalacturonides and so on.24 Therefore, the results obtained from the wounded leaves were combined and analyzed here. The relative expression levels of the structural genes and WRKY TFs in the nonwounded leaves exhibited massive, significant differences compared to those in the wounded and control leaves (Figure 4A,B). It was also found that variegated gene expression patterns of structural genes were observed during days 0–4, whereas similar expression patterns of all structural genes were observed on day 7. The structural genes in the wounded leaves seemed to play a more sensitive role in this response over those in the nonwounded leaves, as was suggested by the increase of NCS and CNMT on day 2 in the wounded leaves (Figure 4A). The transcript levels of WRKY TFs in the nonwounded leaves apparently displayed enormous increases over those in the wounded and control leaves; they also remained relatively high throughout the observation period. NNU_24385 was again recognized to be the predominant WRKY TF. It increased considerably on day 0 after wounding, reached its highest transcript level on day 3, and then gradually increased until day 7 (Figure 4B). Taken together, all target genes exhibited the same expression order as what was observed in the wounded leaves, but they performed their roles at different time points in the nonwounded leaves. Interestingly, the increases in CNMT and NNU_24385 on day 3 in the nonwounded leaves correlated well with the highest total alkaloid accumulation (Figure 4A–C). Moreover, we could observe the fluctuating accumulation of total alkaloid content between days and leaves, as was clearly seen on days 2 and 3 (Figure 4C).
Figure 4.

Comparison of the relative gene expression and aporphine-type alkaloid accumulation in the wounded (W) and nonwounded (NW) leaves. (A) Relative expressions of putative NCS, 6OMT, CNMT, and CYP80G2. (B) Relative expression of five putative WRKY TFs. (C) Sum of aporphine-type alkaloid contents. (D) Abaxial site of commercial Rosem Plenum lotus leaf: (A) wounded site; and (B) nonwounded site. The different lowercase letters indicate significant differences at p < 0.05.
Role of BIA-Related Genes in Normal and Abiotic Stress Conditions
To better understand the role of BIA-related genes in mechanical wound response, we also studied the relationship between nuciferine and N-nornuciferine accumulation and the expression levels of their corresponding genes in normal leaves of commercial lotus used in a previous study and also in nine varieties of wild Thai lotus. Size-matched leaves were used, and wild lotuses were classified based on their flower morphology (Figure 5A). The results suggested that, in normal conditions, all subjected lotuses maintained high relative expressions (%) of CYP80G2 and NCS and had quite low proportional expressions of the two methyltransferases (Figure 5B). Interestingly, WRKY TFs (NNU_24385) were a major component of the transcripts, as was also found in the mechanical wounding experiment (Figure 5C). Altogether, the relative expression levels (%) of NCS and WRKY TFs (NNU_24385) seemed to correlate well with the accumulation of target compounds, as shown in Figure 5D.
Figure 5.

Gene expression profiles of BIA-related genes in mature leaves of nine wild lotus varieties. (A) Leaf at mature stage and flower morphology of the examined wild and commercial lotuses. (B) Relative expression (%) of four putative structural genes. (C) Relative expression (%) of putative WRKY TFs. (D) Bar graph showing the comparison of total BIA content in two organs between the commercial lotus and the wild lotus. Error bars show the SD of the mean.
Discussion
In this study, we isolated the sequences that were deemed to encode 6OMT, CNMT, and CYP80G2 from lotus cDNA and retrieved five putative sequences that were proposed to encode NCS from the GenBank database. Results from amino acid sequence alignment of all isolated CNMT genes showed that they possessed a conserved region, as documented in CjCNMT. This was indicated by the motif A of a conserved S-adenosylmethionine (SAM)-binding domain, located at the C-terminal end containing three conserved glycine residues.20,21 Unlike CNMT, all isolated 6OMT possessed motifs A, B, and C, with their conserved amino acid residues (Figure 2B).25 This may suggest that 6OMT and CNMT can be easily classified by the presence of these motifs. Phylogenetic analysis also clearly indicated that Cj6OMT belongs to a different branch from that of CjCNMT (Figure S3). The lotus 6OMT shared a close relationship to Cj6OMT, while the lotus CNMT shared a close relationship to CjCNMT. The putative NnCYP80G2 shared over 80% amino acid identity with corytuberine synthase (CTS or CYP80G2) from P. peltatum. These possessed conserved eukaryotic P450 regions consisting of a helix K region, an aromatic region, and a heme-binding region at the C-terminal end (Figure 2D).10,11 Thus, NnCYP80G2 in this study was deemed to be involved in BIA biosynthesis.
WRKY transcription factors (TFs), which control overall BIA biosynthetic pathways, were also characterized. Generally, WRKY TFs contain approximately 60 amino acids with four-stranded β-sheet WRKY DNA binding domains (DBDs) and zinc-finger motifs. WRKY TFs are divided into three groups: group I (2 WRKY DBDs), group II (single DBD with different C2H2 zinc finger), and group III (single DBD with C2HC zinc finger) (Figure 2E).15 All putative WRKY TFs in this study possessed a single DBD together with a zinc-finger motif; thus, they are characteristic of the group III type of WRKY proteins. Such a characteristic has been reported in other alkaloid-producing plant species, including AtWRKY70 in Arabidopsis thaliana (GenBank accession no. AF421157), GmWRKY58 in Glycine max (GenBank accession no. EU375357), and NtWRKY3 in Nicotiana tabacum (GenBank accession no. AF193770).15
All of the above sequences were used to study their role in mechanically wounded lotus leaves. Overall, we observed an enormous difference in the transcript level between the control and the wounded leaf. According to the previous study of the wound-associated signaling pathway in Arabidopsis leaf, it was found that the wounding alerted the intra- and interdefense mechanism in leaves on the same plant.24 Thus, we designed to separately collect the control leaf from the different plant to avoid this mechanism. The control leaf used in this study represented normal growth condition with a normal transcript level of all target genes. As a result, the gene expressions in the control leaf maintained quite low transcript levels when compared with those in the wounded leaf. Among the target genes, the putative CYP80G2 and one candidate WRKY TF, NNU_24385, were identified to be promising genes because they clearly exhibited the most sensitivity in the wound response system, as indicated by their massive transcript levels immediately after wounding. Interestingly, increases in the expression of three structural genes, NCS, CNMT, and CYP80G2, on day 2 correlated well with BIA accumulation in wounded lotus leaves. NCS might have both a major and a minor role in the accumulation of target compounds because it is located in the initial step of the BIA pathway, providing a simple BIA skeleton as a precursor for various BIA structures in the complex array.26 This was clearly demonstrated in the day 2 wounded leaves, in which NCS performed its role in the initial days after wounding and then maintained high transcript levels until day 7. The downstream methyltransferase enzymes CNMT and 6OMT then subsequently took on the role for the modification step on either the same or the following days. CNMT performed well as a cofactor with CYP80G2, as indicated by the rapid formation of nuciferine and N-nornuciferine products. We suggest that coexpressions of CNMT and CYP80G2 in plant or bacterial systems would clarify the collaboration of these genes. Moreover, a slight increase of 6OMT during days 3–7 may suggest two possibilities. (1) Conversion of N-nornuciferine and nuciferine to dehydronuciferine: this compound presents in lotus leaf and is structurally formed via dehydroxylation reaction as proposed in Figure 1B.5 This could result in the decrease of nuciferine and N-nornuciferine contents. (2) Migration of compounds to neighboring organs: high accumulation of nuciferine and N-nornuciferine after mechanical wounding might be toxic to the leaf tissues. The presence of these compounds in their petiole may support that these overproduced metabolites translocate to the petiole to decrease the chance of toxicity in the leaf tissues.24,28 Further investigations on these events are interesting.
Mechanical wounding affected related gene expression and BIA accumulation not only in the wounded tissues, but also in the neighboring tissues. Briefly, two structural genes (CYP80G2 and CNMT) and one putative WRKY TF (NNU_24385) played an important role on day 3 in the nonwounded leaf when the highest total BIA content was detected, particularly nuciferine. The normal behavior of CNMT in the mature leaves of all studied wild Thai lotuses may support the conclusion that CNMT is another key gene for nuciferine production. Similarly, CNMT has been reported to maintain low transcript levels in alkaloid-producing plant species and is also involved in the bottleneck step of alkaloid biosynthesis.21,26 The results herein underlined the fact that CNMT is a promising gene, which may have a potent role in nuciferine accumulation in lotus leaves.
Besides the biosynthetic genes, TFs are known to play an important role in the regulation of BIA production. Molecular study showed the regulation of WRKY TFs on secondary metabolite production, such as that of phenolic compounds and alkaloids.15 Therefore, additional studies of transcript levels of WRKY TFs along with other biosynthetic genes will be helpful for the improvement of alkaloid production. We found that NNU_24385 seemed to be a predominant WRKY TF among all those examined because its expression level was related to the highest accumulation of target compounds in both wounded and nonwounded leaves (Figure 4B). According to our phylogenetic analysis, NNU_24385 shares the highest percent similarity in amino acid sequences with CjWRKY1, which regulates BIA biosynthesis in C. japonica.18 WRKY1 from alkaloid-producing species, including C. roseus, C. japonica, and A. thaliana, has been reported to have potential function in BIA biosynthesis, and wounding could also induce the WRKY TF expression, which subsequently regulates the BIA pathway in P. somniferum.15,17 In addition, WRKY1 from Arabidopsis triggered the transcript level of the cytochrome P450-dependent oxidase berbamunine synthase (CYP80A1), leading to increased alkaloid production in callus culture of California poppy.10 Likewise, NNU_24385 was found to be the most active transcript in the mature leaf of most selected wild Thai lotuses (Figure 5A,B). This may suggest that NNU_24385 is a wound-related WRKY TF and is responsible for BIA accumulation in lotus leaves. Additionally, metabolite translocation may occur in the tissues as a result of the fluctuation in BIA content among days because high quantities of secondary metabolites can be toxic to plant tissues themselves.24,27,28
Our study provides some insight into the biosynthetic pathway of nuciferine and N-nornuciferine in lotus. To date, the chemical structures of various BIAs have been reported in Nelumbo organs, for example in the embryo, rhizome, flower, and leaf; however, their biosynthesis pathways are not yet documented.1,3,29−31 Accordingly, we mapped the reported BIAs’ structures elucidated from those organs and known BIA-related enzymes together, as shown in Figure 1A.1,10,11,14,29 Normally, the order of structural enzymes in simple BIA complex arrays occurs with the use of NCS, 6OMT, and CNMT to generate norcoclaurine, (S)-coclaurine, and methylcoclaurine, respectively.29,32,33 We further predicted that CYP80G2 might later occupy the BIA backbone to yield lirinidine, and then 7OMT will form nuciferine (Figure 1A). Alternatively, N-nornuciferine might be constructed via a different array by CYP80G2, 7OMT, and 6OMT, respectively (Figure 1B). The roles of CYP80G2 and CNMT in the mechanically wounded lotus leaf suggested that C–C coupling and methylation at the N position of the isoquinoline nucleus may occur before O-methylation; thus, the proposed biosynthetic pathway following these results has been drawn (Figure 1B). Determining the structures of asimilobine and O-nornuciferine could support the results from our mechanical wounding experiment and lead to clarification of the possible pathway of nuciferine. Nuciferine is probably produced via CYP80G2, CNMT, and 6OMT, respectively. Similarly, nuciferine could be directly generated from asimilobine and N-nornuciferine via CYP80G2, 6OMT, and CNMT, respectively. Our findings on asimilobine, O-nornuciferine, and N-nornuciferine structures suggested that methylation on the N or O position of the isoquinoline nucleus might not occur sequentially; hence, we proposed two possible pathways that lead to the formation of nuciferine. Our results of relative gene expression using the mechanical wounding method provide information on some parts of this mechanism. Further study on the function of these genes is required for better understanding and confirmation of the pathways.
Methods
Plant Materials
Commercial lotuses purchased from a well-known market in the Bangkok, Thailand, area were used for the mechanical wounding experiment on lotus leaves. A total of nine wild Thai lotuses were collected from the germplasm located in the collection of Rajamangkala University of Technology Tawan-ok, Chonburi, Thailand, and used to study gene expression profiles and BIA accumulation in lotus leaves under normal conditions. According to the taxonomic description retrieved from The Botanical Garden Organization, Ministry of Natural Resources and Environment (http://www.qsbg.org/webBGO/database.html), the commercial lotus is classified as Album Plenum, while the wild lotuses were classified into three groups: Album Plenum, Rosem Plenum lotus, and Bua Khem Chin.
Gene Isolation Using Specifically Designed Primers
Four structural genes, including NCS, 6OMT, CNMT, and CYP80G2, as well as alkaloid-related WRKY TFs were targeted in this study. Five amino acid sequences of NnNCS were accessed from the GenBank database with accession nos ANI26411, ANI26412, ANI26413, AND61511, and AND61512.19 Meanwhile, target genes that are related to 6OMT, CNMT, and CYP80G2 were isolated from lotus cDNA by specific primers designed based on predicted nucleotide sequences accessed for the lotus (Table S1). These were subsequently used in quantitative real-time polymerase chain reaction (qRT-PCR), which was carried out as described in a previous study.19 The amplified cDNA fragments were inserted into pEASY-T1 vector (TransGen Biotech, Beijing, China) and subsequently sequenced. Meanwhile, a total of 60 deduced amino sequences of lotus WRKY TF were retrieved from the lotus genome (http://lotus-db.wbgcas.cn) using the BLAST analysis against C. japonica WRKY1 (GenBank accession no. AB26740).34 Due to the close relationship among known WRKY TFs in BIA-producing plant species, only 10 out of 60 sequences were selected for qRT-PCR analysis (Figure S1).
Phylogenetic Tree Analysis
Translated amino acid sequences of all target genes retrieved from the Expasy translate tool (http://web.expasy.org/translate/) were aligned with MUSCLE alignment, and they were further analyzed for their phylogenetic relationships using MEGA7 program.35 The resulting data matrix was analyzed using equally weighted maximum parsimony. Phylogenetic trees were constructed using a maximum likelihood method based on the JTT matrix-based model. The topology with the most superior log-likelihood value was selected. The bootstrap consensus tree was inferred from 1000 replicates, and branches with less than 50% bootstrap support were collapsed.
Expression Profile of BIA-Related Genes Using RT-PCR
Approximately 100 mg of each sample was finely ground in liquid nitrogen and then subjected to total RNA isolation and genomic DNA removal using an RNA prep pure kit for polysaccharides and polyphenolics-rich plants (TIANGEN China). cDNA synthesis was conducted in one step using ProtoScript II Reverse Transcriptase (NEB England). Next, 500 ng of each cDNA template was then mixed in 10 μL total of real-time PCR reaction mixture in the presence of 5 μL of 2 × iTaq Universal SYBR Green Supermix (BIO-RAD) and 0.2 μM of each primer (Tables S2 and S3). Lotus actin gene (accession no. XM_010267617) was used as a constitutive control. The amplification program was set as described in a previous study.19 The samples were prepared in triplicate, and procedures were performed in a BIO-RAD T100 Thermal Cycler (BIO-RAD).
Wounding Experiment
Fully opened leaves from lotus shoots with sizes of 11 cm × 13 cm were chosen for use in three treatments as follows: control leaf, wounded (W) leaf, and nonwounded (NW) leaf. The control leaves were selected separately, whereas the W and NW leaf treatments were separately performed on two abaxial sites of the same leaf lined by the midrib. Mechanical wounding was performed by making several parallel cuts on the wounded leaves using a sterile surgical blade no. 21 (Skidmore Instruments, England) (Figure 4D). The W and NW treatments were observed at the following set observation days: 0, 2, 3, 4, and 7 days after treatment.
Identification and Measurement of Alkaloid Composition by HPLC Analysis
Alkaloid extraction and separation methods used herein were the same as those described in a previous study.5 Separation and detection were performed by HPLC-DAD (SHIMADZU) using a TOSOH C18 column (4.6 mm × 150 mm, 3.5 μm diameter). Nuciferine and N-nornuciferine alkaloid contents were determined against the linear standard curves of nuciferine (Yuanye Biotechnology Shanghai, China) as a representative aporphine-type alkaloid (Figure S2). The peak areas of nuciferine and N-nornuciferine were pooled to calculate the total content of major BIAs.
Validation of HPLC Assay
Six concentrations of nuciferine standard, including 5, 20, 40, 60, 80, and 100 μg/mL, were prepared in extraction buffer to make a calibration curve of aporphine-type BIA standard for HPLC analysis and to measure the recoveries of alkaloids from the assay using a standard addition method. They were then subjected to alkaloid extraction and separation following the reference protocol mentioned above. Their recoveries at each concentration listed above were 65.75, 97.40, 103.15, 103.07, 103.44, and 96.38%, respectively. The relative standard deviation of interday injection repeats was 0.26%. These values suggested that the HPLC system was reliable.36,37
Statistical Analysis
Triplicates of each alkaloid standard and samples were subjected to analyses to determine statistical significance in SPSS software version 21 with one-way analysis of variance methods. Significant differences were defined as those with p < 0.05.
Acknowledgments
This research was supported by funds received from the 90th Anniversary of Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund) Batch# 37 Round 4/2017 Academic Year 2016 and new researcher funding from the National Science and Technology Development Agency (project No. SCH-NR2016-091).
Supporting Information Available
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acsomega.8b00827.
Information on the primers used for gene isolation and RT-PCR, the nucleotide sequences of putative BIA-related genes in lotus, phylogenetic tree of lotus WRKY TF, 6OMT, and CNMT (PDF)
The authors declare no competing financial interest.
Supplementary Material
References
- Nakamura S.; Nakashima S.; Tanabe G.; Oda Y.; Yokota N.; Fujimoto K.; Matsumoto T.; Sakuma R.; Ohta T.; Ogawa K. Alkaloid constituents from flower buds and leaves of sacred lotus (Nelumbo nucifera, Nymphaeaceae) with melanogenesis inhibitory activity in B16 melanoma cells. Bioorg. Med. Chem. 2013, 21, 779–787. 10.1016/j.bmc.2012.11.038. [DOI] [PubMed] [Google Scholar]
- Sakuljaitrong S.; Buddhakala N.; Chomko S.; Talubmook C. Effects of flower extract from lotus (Nelumbo nucifera) on hypoglycemic and hypolipidemic in streptozotocin-induced diabetic rats. Int. J. Sci. Eng. Res. 2013, 4, 1441–1446. [Google Scholar]
- Do T. C. M. V.; Nguyen T. D.; Tran H.; Stuppner H.; Ganzera M. Analysis of alkaloids in lotus (Nelumbo nucifera Gaertn.) leaves by non-aqueous capillary electrophoresis using ultraviolet and mass spectrometric detection. J. Chromatogr. A 2013, 1302, 174–180. 10.1016/j.chroma.2013.06.002. [DOI] [PubMed] [Google Scholar]
- Hu J.; Pan L.; Liu H.; Wang S.; Wu Z.; Ke W.; Ding Y. Comparative analysis of genetic diversity in sacred lotus (Nelumbo nucifera Gaertn.) using AFLP and SSR markers. Mol. Biol. Rep. 2012, 39, 3637–3647. 10.1007/s11033-011-1138-y. [DOI] [PubMed] [Google Scholar]
- Deng X.; Zhu L.; Fang T.; Vimolmangkang S.; Yang D.; Ogutu C.; Liu Y.; Han Y. Analysis of isoquinoline alkaloid composition and wound-induced variation in Nelumbo using HPLC-MS/MS. J. Agric. Food. Chem. 2016, 64, 1130–1136. 10.1021/acs.jafc.5b06099. [DOI] [PubMed] [Google Scholar]
- Duan X. H.; Jiang J. Q. A new benzylisoquinoline alkaloid from stems of Nelumbo nucifera. Chin. Chem. Lett. 2008, 19, 308–310. 10.1016/j.cclet.2008.01.013. [DOI] [Google Scholar]
- Balandrin M. F.; Klocke J.; Wurtele E. S.; Bollinger W. H. Natural plant chemicals: sources of industrial and medicinal materials. Science 1985, 228, 1154–1159. 10.1126/science.3890182. [DOI] [PubMed] [Google Scholar]
- Liscombe D. K.; Facchini P. J. Molecular cloning and characterization of tetrahydroprotoberberine cis-N-methyltransferase, an enzyme Involved in alkaloid biosynthesis in opium poppy. J. Biol. Chem. 2007, 282, 14741–14751. 10.1074/jbc.M611908200. [DOI] [PubMed] [Google Scholar]
- Chen J.; Gao K.; Liu T.; Zhao H.; Wang J.; Wu H.; Liu B.; Wang W. Aporphine alkaloids: a kind of alkaloids’ extract source, chemical constitution and pharmacological actions in different botany. Asian J. Chem. 2013, 25, 10015–10027. [Google Scholar]
- Ikezawa N.; Iwasa K.; Sato F. Molecular cloning and characterization of CYP80G2, a cytochrome P450 that catalyzes an intramolecular C–C phenol coupling of (S)-reticuline in magnoflorine biosynthesis, from cultured Coptis japonica cells. J. Biol. Chem. 2008, 283, 8810–8821. 10.1074/jbc.M705082200. [DOI] [PubMed] [Google Scholar]
- Gesell A.; Rolf M.; Ziegler J.; Chávez M. L. D.; Huang F.-C.; Kutchan T. M. CYP719B1 is salutaridine synthase, the CC phenol-coupling enzyme of morphine biosynthesis in opium poppy. J. Biol. Chem. 2009, 284, 24432–24442. 10.1074/jbc.M109.033373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou W. M.; Kutchan T. M. Enzymatic oxidations in the biosynthesis of complex alkaloids. Plant J. 1998, 15, 289–300. 10.1046/j.1365-313X.1998.00220.x. [DOI] [PubMed] [Google Scholar]
- Nelson D. R.; Schuler M. A. Cytochrome P450 genes from the sacred lotus genome. Trop. Plant Biol. 2013, 6, 138–151. 10.1007/s12042-013-9119-z. [DOI] [Google Scholar]
- Pauli H. H.; Kutchan T. M. Molecular cloning and functional heterologous expression of two alleles encoding (S)-N-methylcoclaurine 3′-hydroxylase (CYP80B1), a new methyl jasmonate-inducible cytochrome P-450-dependent mono-oxygenase of benzylisoquinoline alkaloid biosynthesis. Plant J. 1998, 13, 793–801. 10.1046/j.1365-313X.1998.00085.x. [DOI] [PubMed] [Google Scholar]
- Phukan U. J.; Jeena G. S.; Shukla R. K. WRKY transcription factors: molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760 10.3389/fpls.2016.00760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schluttenhofer C.; Yuan L. Regulation of specialized metabolism by WRKY transcription factors. Plant Physiol. 2015, 167, 295–306. 10.1104/pp.114.251769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishra S.; Triptahi V.; Singh S.; Phukan U. J.; Gupta M.; Shanker K.; Shukla R. K. Wound induced tanscriptional regulation of benzylisoquinoline pathway and characterization of wound inducible PsWRKY transcription factor from Papaver somniferum. PLoS One 2013, 8, e52784 10.1371/journal.pone.0052784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suttipanta N.; Pattanaik S.; Kulshrestha M.; Patra B.; Singh S. K.; Yuan L. The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 2011, 157, 2081–2093. 10.1104/pp.111.181834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vimolmangkang S.; Deng X.; Owiti A.; Meelaph T.; Ogutu C.; Han Y. Evolutionary origin of the NCSI gene subfamily encoding norcoclaurine synthase is associated with the biosynthesis of benzylisoquinoline alkaloids in plants. Sci. Rep. 2016, 6, 26323 10.1038/srep26323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inui T.; Tamura K.-i.; Fujii N.; Morishige T.; Sato F. Overexpression of Coptis japonica norcoclaurine 6-O-methyltransferase overcomes the rate-limiting step in benzylisoquinoline alkaloid biosynthesis in cultured Eschscholzia californica. Plant Cell Physiol. 2007, 48, 252–262. 10.1093/pcp/pcl062. [DOI] [PubMed] [Google Scholar]
- Choi K. B.; Morishige T.; Shitan N.; Yazaki K.; Sato F. Molecular cloning and characterization of coclaurine N-methyltransferase from cultured cells of Coptis japonica. J. Biol. Chem. 2002, 277, 830–835. 10.1074/jbc.M106405200. [DOI] [PubMed] [Google Scholar]
- Xie Z.; Zhang Z.-L.; Zou X.; Huang J.; Ruas P.; Thompson D.; Shen Q. J. Annotations and functional analyses of the rice WRKY gene superfamily reveal positive and negative regulators of abscisic acid signaling in aleurone cells. Plant Physiol. 2005, 137, 176–189. 10.1104/pp.104.054312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rehrig E. M.; Appel H. M.; Jones A. D.; Schultz J. C. Roles for jasmonate-and ethylene-induced transcription factors in the ability of Arabidopsis to respond differentially to damage caused by two insect herbivores. Front. Plant Sci. 2014, 5, 407 10.3389/fpls.2014.00407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savatin D. V.; Gramegna G.; Modesti V.; Cervone F. Wounding in the plant tissue: the defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470 10.3389/fpls.2014.00470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morishige T.; Tsujita T.; Yamada Y.; Sato F. Molecular characterization of the S-adenosyl-L-methionine: 3′-hydroxy-N-methylcoclaurine 4′-O-methyltransferase involved in isoquinoline alkaloid biosynthesis in Coptis Japonica. J. Biol. Chem. 2000, 275, 23398–23405. 10.1074/jbc.M002439200. [DOI] [PubMed] [Google Scholar]
- Staniek A.; Bouwmeester H.; Fraser P. D.; Kayser O.; Martens S.; Tissier A.; van der Krol S.; Wessjohann L.; Warzecha H. Natural products-modifying metabolite pathways in plants. Biotechnol. J. 2013, 8, 1159–1171. 10.1002/biot.201300224. [DOI] [PubMed] [Google Scholar]
- Fürstenberg-Hägg J.; Zagrobelny M.; Bak S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. 10.3390/ijms140510242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green T. R.; Ryan C. A. Wound-induced proteinase inhibitor in plant leaves: a possible defense mechanism against insects. Science 1972, 175, 776–777. 10.1126/science.175.4023.776. [DOI] [PubMed] [Google Scholar]
- Facchini P. J. Alkaloid biosynthesis in plants: biochemistry, cell biology, molecular regulation, and metabolic engineering applications. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 29–66. 10.1146/annurev.arplant.52.1.29. [DOI] [PubMed] [Google Scholar]
- Kashiwada Y.; Aoshima A.; Ikeshiro Y.; Chen Y.-P.; Furukawa H.; Itoigawa M.; Fujioka T.; Mihashi K.; Cosentino L. M.; Morris-Natschke S. L.; et al. Anti-HIV benzylisoquinoline alkaloids and flavonoids from the leaves of Nelumbo nucifera, and structure–activity correlations with related alkaloids. Bioorg. Med. Chem. 2005, 13, 443–448. 10.1016/j.bmc.2004.10.020. [DOI] [PubMed] [Google Scholar]
- JI Y.; Yu M.; Wang B.; Zhang Y. The extraction, separation and purification of alkaloids in the natural medicine. J. Chem. Pharm. Res. 2014, 6, 338–345. [Google Scholar]
- Stadler R.; Kutchan T. M.; Zenk M. H. (S)-norcoclaurine is the central intermediate in benzylisoquinoline alkaloid biosynthesis. Phytochemistry 1989, 28, 1083–1086. 10.1016/0031-9422(89)80187-6. [DOI] [Google Scholar]
- Hashimoto T.; Yamada Y. Alkaloid biogenesis: molecular aspects. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 257–285. 10.1146/annurev.pp.45.060194.001353. [DOI] [Google Scholar]
- Wang K.; Deng J.; Damaris R. N.; Yang M.; Xu L.; Yang P. LOTUS-DB: an integrative and interactive database for Nelumbo nucifera study. Database 2015, 2015, bav023 10.1093/database/bav023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S.; Stecher G.; Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papadoyannis I. N.; Samanidou V. F. Validation of HPLC instrumentation. J. Liq. Chromatogr. Relat. Technol. 2004, 27, 753–783. 10.1081/JLC-120029697. [DOI] [Google Scholar]
- Taverniers I.; De Loose M.; Van Bockstaele E. Trends in quality in the analytical laboratory. II. analytical method validation and quality assurance. TrAC, Trends Anal. Chem. 2004, 23, 535–552. 10.1016/j.trac.2004.04.001. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.